FliZ Regulated the Biofilm Formation of Bacillus subtilis Bs916 and Its Biocontrol Efficacy on Rice Sheath Blight
ZHOU HuaFei1,2, YANG HongFu1, YAO KeBing1, ZHUANG YiQing1, SHU ZhaoLin1, CHEN ZhiYi,31 Zhenjiang Institute of Agricultural Sciences in Hilly Region of Jiangsu, Jurong 212400, Jiangsu 2 College of Plant Protection, Nanjing Agricultural University, Nanjing 210095 3 Institute of Plant Protection, Jiangsu Academy of Agricultural Sciences, Nanjing 210014
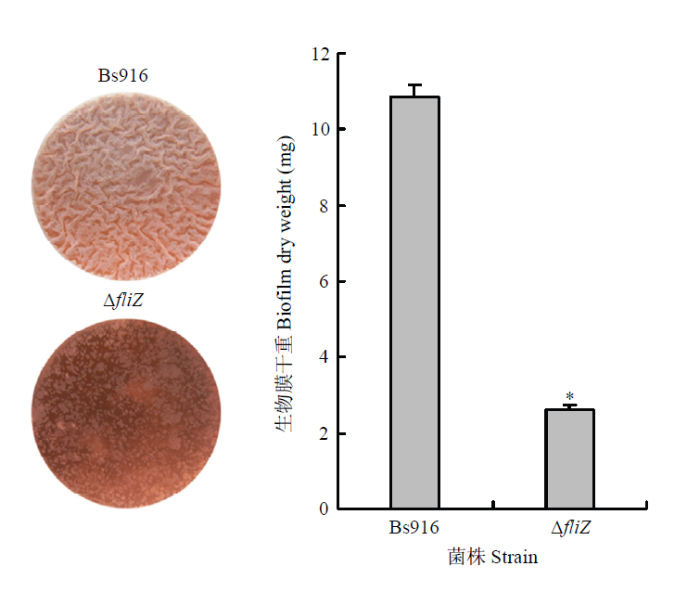
Abstract 【Objective】The objective of this study is to discover and identify new regulatory genes on biofilm formation of Bacillus subtilis Bs916, detect its effect on biofilm formation of Bs916 and biocontrol efficacy on rice sheath blight.【Method】The single knockout mutant of Bs916 at fliZ was construct by homologous recombination, and its defects in biofilm formation were verified by dry weight analysis. The anti-bacterial effect of fliZ mutant and Bs916 on rice sheath blight pathogen (Rhizoctonia solani) was detected by flat panel. The relative production of 3 lipopeptide antibiotics (LPs) surfactin, bacillomycin L, and fengycin in fliZ mutant and Bs916 was detected by HPLC. The GFP-labeled strains of Bs916 and fliZ mutant were constructed by green fluorescent labeling, the colonization ability of them in rice stalks was observed, and the biocontrol efficacy of fliZ mutant and Bs916 on rice sheath blight was detected.【Result】The single knockout mutant of Bs916 at fliZ was successfully constructed. Compared with the three-dimensional structure biofilm of the control group Bs916, fliZ mutant only formed a planar two-dimensional structure biofilm, appeared broken form, which proved that it had significant defects in biofilm formation. Quantitative analysis of the dry weight of biofilms showed that the biofilm dry weight of fliZ mutant was only 23% of the control group Bs916, which further verified that the biofilm formation ability of fliZ mutant was significantly decreased. The motility test found that the expanded diameter of fliZ mutant was only 32% of Bs916, which proved that the swimming ability of fliZ mutant was significantly reduced. The bacteriostatic test showed that the antibacterial bandwidth of the two strains was basically the same, and it is proved that the antibacterial activity of fliZ mutant against R. solani was not significantly different from that of Bs916. The relative production of three LPs bacillomycin L, surfactin, and fengycin in fliZ mutant and Bs916 was successfully detected. Compared with Bs916, the relative production of bacillomycin L was significantly increased by 1 time in fliZ mutant, but the relative production of surfactin and fengycin was not significantly different from that of Bs916. The colonization test of rice stalks showed that the number of fliZ mutant was significantly lower than that of Bs916, and there was no significant aggregation effect near the rice sheath blight lesions, and presented an unordered state, which proved that the colonization ability of fliZ mutant on rice stalks was significantly lower than that of Bs916. The field biocontrol trials against rice sheath blight showed that biocontrol efficacy of fliZ mutant ranged from 6.0% to 20.7% on days 6-15, which was significantly lower than that of Bs916 (36.0%-57.6%). It was proved that the biocontrol efficacy of fliZ mutant on rice sheath blight was significantly reduced.【Conclusion】The new regulatory gene fliZ of Bs916 biofilm identified in this study is located in the signal pathway controlling flagellar movement, directly acts on swimming and expansion of the bacteria, and can significantly control the biofilm formation and its biocontrol efficacy on rice sheath blight. Keywords:Bacillus subtilis Bs916;regulated genes;biofilm;lipopeptide antibiotics (LPs);colonization;rice sheath blight;biocontrol efficacy
PDF (1747KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 周华飞, 杨红福, 姚克兵, 庄义庆, 束兆林, 陈志谊. FliZ调控枯草芽孢杆菌Bs916生物膜形成 及其对水稻纹枯病的防治效果[J]. 中国农业科学, 2020, 53(1): 55-64 doi:10.3864/j.issn.0578-1752.2020.01.005 ZHOU HuaFei, YANG HongFu, YAO KeBing, ZHUANG YiQing, SHU ZhaoLin, CHEN ZhiYi. FliZ Regulated the Biofilm Formation of Bacillus subtilis Bs916 and Its Biocontrol Efficacy on Rice Sheath Blight[J]. Scientia Acricultura Sinica, 2020, 53(1): 55-64 doi:10.3864/j.issn.0578-1752.2020.01.005
A—C图蓝色代表Bs916,红色代表ΔfliZ突变株 In fig A-C, blue represents Bs916, red represents ΔfliZ mutant. A:杆菌霉素L 相对产量Relative production of bacillomycin L;B:表面活性素相对产量Relative production of surfactin;C:泛革素相对产量Relative production of fengycin;D:3种脂肽抗生素相对产量柱形图Histogram of relative production of 3 lipopeptide antibiotics. *: P<0.05 Fig. 4Relative production of 3 lipopeptide antibiotics of ΔfliZ mutant and Bs916
KUMAR K VK, REDDY MS, KLOEPPER JW, LAWRENCE KS, GROTH DE, MILLER ME . Sheath blight disease of rice (Oryza sativa L.)-An overview Biosciences, Biotechnology Research Asia, 2009,6(2):465-480. [本文引用: 1]
CHOUDHURYD, ANAND YR, KUNDUS, NATHR, KOLE RK, SAHAJ . Effect of plant extracts against sheath blight of rice caused by Rhizoctonia solani Journal of Pharmacognosy and Phytochemistry, 2017,6(4):399-404. [本文引用: 1]
BONANOMIG, CESARANOG, ANTIGNANIV, DI MAIOC, DE FILIPPISF, SCALAF . Conventional farming impairs Rhizoctonia solani disease suppression by disrupting soil food web Journal of Phytopathology, 2018,166(9):663-673. [本文引用: 1]
LUOC, ZHOUH, ZOUJ, WANGX, ZHANGR, XIANGY, CHENZ . Bacillomycin L and surfactin contribute synergistically to the phenotypic features of Bacillus subtilis 916 and the biocontrol of rice sheath blight induced by Rhizoctonia solani Applied Microbiology and Biotechnology, 2015,99(4):1897-1910. [本文引用: 4]
SHRESTHA BK, KARKI HS, GROTH DE, JUNGKHUNN, HAM JH . Biological control activities of rice-associated Bacillus sp. strains against sheath blight and bacterial panicle blight of rice PLoS ONE, 2016,11(1):e0146764. [本文引用: 1]
QIZ, YUJ, SHENL, YUZ, YUM, DUY, ZHANGR, SONGT, YINX, ZHOUY, LIH, WEIQ, LIUY . Enhanced resistance to rice blast and sheath blight in rice (Oryza sativa L.) by expressing the oxalate decarboxylase protein Bacisubin from Bacillus subtilis Plant Science, 2017,265:51-60. [本文引用: 1]
XUZ, ZHANGH, SUNX, LIUY, YANW, XUNW, SHENQ, ZHANGR . Bacillus velezensis wall teichoic acids are required for biofilm formation and root colonization Applied and Environmental Microbiology, 2019,85(5):e02116-18. [本文引用: 1]
GUTTENPLAN SB, BLAIR KM, KEARNS DB . The EpsE flagellar clutch is bifunctional and synergizes with EPS biosynthesis to promote Bacillus subtilis biofilm formation PLoS Genetics, 2010,6(12):e1001243. [本文引用: 2]
CZACZYKK, MYSZKAK . Biosynthesis of extracellular polymeric substances (EPS) and its role in microbial biofilm formation Polish Journal of Environmental Studies, 2007,16(6):799-806. [本文引用: 1]
MARVASIM, VISSCHER PT, CASILLAS MARTINEZL . Exopolymeric substances (EPS) from Bacillus subtilis: Polymers and genes encoding their synthesis FEMS Microbiology Letters, 2010,313(1):1-9. [本文引用: 1]
DIEHLA, ROSKEY, BALLL, CHOWDHURYA, HILLERM, MOLIèREN, KRAMERR, ST?PPLERD, WORTH CL, SCHLEGELB, LEIDERTM, CREMERN, ERDMANNN, LOPEZD, STEPHANOWITZH, KRAUSEE, VAN ROSSUM BJ, SCHMIEDERP, HEINEMANNU, TURGAYK, AKBEYü, OSCHKINATH . Structural changes of TasA in biofilm formation of Bacillus subtilis Proceedings of the National Academy of Sciences of the United States of America, 2018,115(13):3237-3242. [本文引用: 2]
ZHOUH, LUOC, FANGX, XIANGY, WANGX, ZHANGR, CHENZ . Loss of GltB inhibits biofilm formation and biocontrol efficiency of Bacillus subtilis Bs916 by altering the production of γ-polyglutamate and three lipopeptides. PLoS ONE, 2016,11(5):e0156247. [本文引用: 4]
BISCHOFF DS, WEINREICH MD, ORDAL GW . Nucleotide sequences of Bacillus subtilis flagellar biosynthetic genes fliP and fliQ and identification of a novel flagellar gene, fliZ Journal of Bacteriology, 1992,174(12):4017-4025. [本文引用: 1]
XU SJ, PENGZ, CUIB, WANGT, SONGY, ZHANGL, WEIG, WANGY, SHENX . FliS modulates FlgM activity by acting as a non-canonical chaperone to control late flagellar gene expression, motility and biofilm formation in Yersinia pseudotuberculosis Environmental Microbiology, 2014,16(4):1090-1104. [本文引用: 1]
YANG BY, FUD, HU FP, CAI XQ . Function analysis of flagellar gene fliS in Acidovorax citrulli. Scientia Agricultura Sinica, 2017,50(15):2946-2956. (in Chinese) [本文引用: 1]
BRANDA SS, VIK?, FRIEDMANL, KOLTERR . Biofilms: the matrix revisited TRENDS in Microbiology, 2005,13(1):20-26. [本文引用: 1]
BRANDA SS, CHUF, KEARNS DB, LOSICKR, KOLTERR . A major protein component of the Bacillus subtilis biofilm matrix Molecular Microbiology, 2006,59(4):1229-1238.
ROMEROD, AGUILARC, LOSICKR, KOLTERR . Amyloid fibers provide structural integrity to Bacillus subtilis biofilms Proceedings of the National Academy of Sciences of the United States of America, 2010,107(5):2230-2234. [本文引用: 1]
KOBAYASHIK, IWANOM . BslA (YuaB) forms a hydrophobic layer on the surface of Bacillus subtilis biofilms Molecular Microbiology, 2012,85(1):51-66. [本文引用: 1]
KEARNS DB, CHUF, BRANDA SS, KOLTERR, LOSICKR . A master regulator for biofilm formation by Bacillus subtilis Molecular Microbiology, 2005,55(3):739-749. [本文引用: 1]
DELOUGHERYA, DENGLERV, CHAIY, LOSICKR . Biofilm formation by Bacillus subtilis requires an endoribonuclease-containing multisubunit complex that controls mRNA levels for the matrix gene repressor SinR Molecular Microbiology, 2016,99(2):425-437. [本文引用: 1]
KAMPFJ, GERWIGJ, KRUSEK, CLEVERLEYR, DORMEYERM, GRüNBERGERA, KOHLHEYERD, COMMICHAU FM, LEWIS RJ, STüLKEJ, . Selective pressure for biofilm formation in Bacillus subtilis: Differential effect of mutations in the master regulator SinR on bistability MBio, 2018,9(5):e01464-18. [本文引用: 1]
HAMON MA, STANLEY NR, BRITTON RA, GROSSMAN AD, LAZAZZERA BA . Identification of AbrB-regulated genes involved in biofilm formation by Bacillus subtilis. Molecular Microbiology, 2004,52(3):847-860. [本文引用: 2]
MURRAY EJ, STRAUCH MA, STANLEY-WALL NR . σ X is involved in controllingBacillus subtilis biofilm architecture through the AbrB homologue Abh. Journal of Bacteriology, 2009,191(22):6822-6832. [本文引用: 2]
CHUF, KEARNS DB, BRANDA SS, KOLTERR, LOSICKR . Targets of the master regulator of biofilm formation in Bacillus subtilis Molecular Microbiology, 2006,59(4):1216-1228. [本文引用: 1]
STRAUCH MA, BOBAY BG, CAVANAGHJ, YAOF, WILSONA, LE BRETONY . Abh and AbrB control of Bacillus subtilis antimicrobial gene expression Journal of Bacteriology, 2007,189(21):7720-7732. [本文引用: 1]
FUJITAM, LOSICKR . Evidence that entry into sporulation in Bacillus subtilis is governed by a gradual increase in the level and activity of the master regulator Spo0A Genes and Development, 2005,19(18):2236-2244. [本文引用: 1]
HAMON MA, LAZAZZERA BA . The sporulation transcription factor Spo0A is required for biofilm development in Bacillus subtilis Molecular Microbiology, 2001,42(5):1199-1209.
DUBNAU EJ, CARABETTA VJ, TANNER AW, MIRASM, DIETHMAIERC, DUBNAUD . A protein complex supports the production of Spo0A‐P and plays additional roles for biofilms and the K‐state in Bacillus subtilis Molecular Microbiology, 2016,101(4):606-624.
BANSE AV, CHASTANETA, RAHN-LEEL, HOBBS EC, LOSICKR . Parallel pathways of repression and antirepression governing the transition to stationary phase inBacillus subtilis Proceedings of the National Academy of Sciences of the United States of America, 2008,105(40):15547-15552. [本文引用: 1]
SAIFUDDINN, WONG CW, YASUMIRA AA . Rapid biosynthesis of silver nanoparticles using culture supernatant of bacteria with microwave irradiation Journal of Chemistry, 2009,6(1):61-70. [本文引用: 1]
ZHOU HF, LUO CP, WANG XY, ZHANG RS, CHEN ZY . Construction of Bacillus subtilis Bs916 mutant libraries by transposon tagging and cloning the genes to the organism’s anti-bacterial activities. Scientia Agricultura Sinica, 2013,46(11):2232-2239. (in Chinese) [本文引用: 4]
GRABAM, SAUVAGES, MOULIN FY, URREAG, SABATERS, SANCHEZ-PéREZ JM . Interaction between local hydrodynamics and algal community in epilithic biofilm Water Research, 2013,47(7):2153-2163. [本文引用: 1]
LUOC, LIUX, ZHOUH, WANGX, CHENZ . Nonribosomal peptide synthase gene clusters for lipopeptide biosynthesis in Bacillus subtilis 916 and their phenotypic functions Applied and Environmental Microbiology, 2015,81(1):422-431. [本文引用: 2]
ZHOU EX, CAO JX, YANGM, ZHU XR . Studies on the genetic diversity of Rhizoctonia solani AG-1-IA from six provinces in the southern China. Journal of Nanjing Agricultural University, 2002,25(3):36-40. (in Chinese) [本文引用: 1]
ZOU CJ, TANGF, YANGM, HE XX, LI XJ, ZHOU EX . Studies on biological characteristics and pathogenicity differentiation of rice sheath blight pathogen from three provinces in South China Chinese Journal of Rice Science, 2011,25(2):206-212. (in Chinese) [本文引用: 1]
XIANG YP, CHEN ZY, LUO CP, ZHOU HF, LIU YF . The antifungal activities of Bacillus spp. and its relationship with lipopeptide antibiotics produced by Bacillus spp Scientia Agricultura Sinica, 2015,48(20):4064-4076. (in Chinese) [本文引用: 2]
BIANCIOTTOV, ANDREOTTIS, BALESTRINIR, BONFANTEP, PEROTTOS . Mucoid mutants of the biocontrol strain Pseudomonas fluorescens CHA0 show increased ability in biofilm formation on mycorrhizal and nonmycorrhizal carrot roots Molecular Plant-Microbe Interactions, 2001,14(2):255-260. [本文引用: 2]
BAIS HP, FALLR, VIVANCO JM . Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production Plant Physiology, 2004,134(1):307-319. [本文引用: 2]
TIMMUSKS, GRANTCHAROVAN, WAGNER E GH . Paenibacillus polymyxa invades plant roots and forms biofilms Applied and Environmental Microbiology, 2005,71(11):7292-7300. [本文引用: 2]
CHENY, CAOS, CHAIY, CLARDYJ, KOLTERR, GUO JH, LOSICKR . A Bacillus subtilis sensor kinase involved in triggering biofilm formation on the roots of tomato plants Molecular Microbiology, 2012,85(3):418-430. [本文引用: 2]
CHEN ZY, LIU YF, LUF . Study on key technology in the industrialized production of Bacillus subtilis Bs-916, the rice sheath blight control agent. Acta Phytophylacica Sinica, 2004,31(3):230-234. (in Chinese) [本文引用: 1]
WANGX, LUOC, CHENZ . Genome sequence of the plant growth-promoting rhizobacterium Bacillus sp. strain 916 Journal of Bacteriology, 2012,194(19):5467-5468. [本文引用: 1]
IYODAS, KAMIDOIT, HIROSEK, KUTSUKAKEK, WATANABEH . A flagellar gene fliZ regulates the expression of invasion genes and virulence phenotype in Salmonella enterica serovar Typhimurium Microbial Pathogenesis, 2001,30(2):81-90. [本文引用: 1]
KUTSUKAKEK, IKEBET, YAMAMOTOS . Two novel regulatory genes, fliT and fliZ, in the flagellar regulon of Salmonella Genes and Genetics Systems, 1999,74(6):287-292. [本文引用: 1]
WANGY, DING LS, HU YB, ZHANGY, YANG BY, SHEN SY . The flhDC gene affects motility and biofilm formation in Yersinia pseudotuberculosis Science in China Series C: Life Sciences, 2007,50(6):814-821. [本文引用: 1]
ZHUOT, WEIR, SONGX, GUOJ, FANX, KAMAU GG, ZOUH . Molecular study on the carAB operon reveals that carB gene is required for swimming and biofilm formation in Xanthomonas citri subsp. citri BMC Microbiology, 2015,15:225. [本文引用: 1]
SANCHEZ-TORRESV, HUH, WOOD TK . GGDEF proteins YeaI, YedQ, and YfiN reduce early biofilm formation and swimming motility in Escherichia coli Applied and Microbiology Biotechnology, 2011,90(2):651-658. [本文引用: 1]
GAO XW, YAO SY, PHAMH, VATERJ, WANG JS . Lipopeptide antibiotic produced by the engineered strain Bacillus subtilis GEB3 and detection of its bioactivity. Scientia Agricultura Sinica, 2003,36(12):1496-1501. (in Chinese) [本文引用: 1]
 ,3
,3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT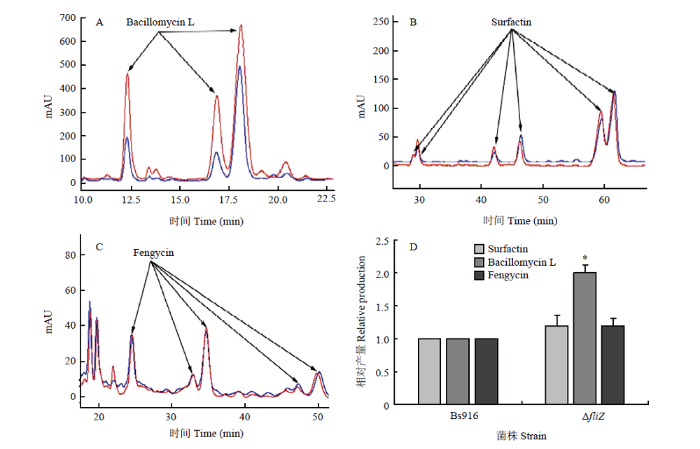 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}