Advances in Mechanisms of Biosynthesis and Regulation of Pasteurella multocida Capsule
GUAN LiJun1, XUE Yun1, DING WenWen1, ZHAO ZhanQin,1,21 Laboratory of Veterinary Biologics Engineering, College of Animal Science and Technology, Henan University of Science and Technology, Luoyang 471003, Henan 2 College of Animal Science and Technology, Henan University of Science and Technology/ Key-Disciplines Laboratory of Safety of Environment and Animal Product, Luoyang 471003, Henan
Abstract Pasteurella multocida can be widely infected with a variety of animals, causing hemorrhagic septicemia or infectious pneumonia. P. multocida possess a viscous capsular polysaccharide on the cell surface, which is a critical structural component and virulence factor, and plays an important role in the interaction between bacteria and the host, promoting the adhesion of bacteria to the host surface and enhancing the virulence of the bacteria. The molecular structure of the P. multocida capsule is similar to that of vertebrate glycosaminoglycan, which is polymerized by repeated disaccharide units to form a linear polysaccharide chain, which is an important immunological material basis for molecular mimicry, resistance to phagocytosis, and immune evasion during the infection of the host. In recent years, a series of important research advances have been made in the biosynthesis and regulation mechanism aspects of P. multocida capsule, providing a certain basic knowledge for the molecular pathogenesis of P. multocida capsule, and supplying a theoretical basis for the development of the capsular polysaccharide vaccine of P. multocida. This paper systematically illuminates the biosynthesis and expression regulation mechanisms of P. multocida capsule, including the serotyping of the capsule, the composition and structure of the capsular polysaccharide, the biosynthesis gene cluster and function of the capsule, the molecular synthesis mechanism of capsular polysaccharide, the expression regulation mechanism of capsular biosynthesis gene cluster, a total of five aspects. According to the capsular antigen, P. multocida is divided into five capsular serogroups of A, B, D, E, and F. The type A capsule GAG component is hyaluronic acid; the type D is heparosan; the type F is chondroitin, which is repeatedly composed of its corresponding disaccharide unit [β-GlcUA/β-GlcNAc], [β-GlcUA/α-GlcNAc], [β-GlcUA/β-GalNAc], respectively; the type B capsular polysaccharide is composed of arabinose, mannose and galactose in a certain structural form, and the composition and chemical structure of type E capsular polysaccharide are uncertain. Genes related to the biosynthesis of A, B, D, E and F capsules of P. multocida exist in the form of gene clusters and are divided into three distinct functional regions, R1, R2 and R3; the R1 region is responsible for transporting the capsular polysaccharide, the R2 region is responsible for the activation of the monosaccharide and the assembly of the capsular polysaccharide, and the R3 region is responsible for the modification of capsular polysaccharide (phospholipid replacement); according to the structure and the number of genes of the R2 region, the biosynthetic gene clusters of the five capsules can be divided into two categories: type A, D and F are Class I, and R2 contains 4 genes; types B and E are Class II, and R2 contains 9 genes, and using the specific gene in the R2 region to design primers, the capsular serotype of P. multocida can be rapidly identified by PCR. The capsular GAG of P. multocida is synthesized in the cytoplasm, and then exported to the cell surface via the ABC transporter formed by the protein encoded by the R1 region, and tightly bound to the cell surface by covalent attachment to the phospholipid; during the biosynthesis of the P. multocida capsular GAG, the glycosyltransferase gene located in the R2 region determines the type of activated monosaccharide and the type of capsular polysaccharide after assembly. In the biosynthetic gene cluster of the P. multocida capsule, the R1 and R2 regions form an operon with the same transcriptional direction, while the R3 transcription direction is opposite, and the promoter regions of both are located on the DNA sequence between the R2 and R3 regions; the transcriptional process of the P. multocida capsular biosynthesis gene cluster is positive regulated by the Fis protein, and the translation process is mainly positive regulated by Hfq protein. Keywords:Pasteurella multocida;capsule;glycosaminoglycan;biosynthesis;expression regulation
PDF (1337KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 关丽君, 薛云, 丁文文, 赵战勤. 多杀性巴氏杆菌荚膜的生物合成及其调控机制研究进展[J]. 中国农业科学, 2020, 53(3): 658-668 doi:10.3864/j.issn.0578-1752.2020.03.016 GUAN LiJun, XUE Yun, DING WenWen, ZHAO ZhanQin. Advances in Mechanisms of Biosynthesis and Regulation of Pasteurella multocida Capsule[J]. Scientia Acricultura Sinica, 2020, 53(3): 658-668 doi:10.3864/j.issn.0578-1752.2020.03.016
ZEIDAN AA, POULSEN VK, JANZENT, BULDOP, DERKX PM F, OREGAARDG, NEVESA R. Polysaccharide production by lactic acid bacteria: from genes to industrial applications FEMS Microbiology Reviews, 2017,41(supp_1):S168-S200. [本文引用: 3]
LISTON SD, MCMAHON SA, LE BASA, SUITS MD L, NAISMITHJ H, WHITFIELDC. Periplasmic depolymerase provides insight into ABC transporter-dependent secretion of bacterial capsular polysaccharides Proceedings of the National Academy of Sciences of the United States of America, 2018,115(21):E4870-E4879. [本文引用: 1]
CARTER GR, BIGLAND CH . Dissociation and virulence in strains of Pasteurella multocida isolated from a variety of lesions. Canadian Journal of Comparative Medicine and Veterinary Science, 1953,17(17):473-479. [本文引用: 1]
WANG KC, LU CP, FAN WX . Bacterial capsular polysaccharide Acta Microbiologica Sinica, 2011,51(12):1578-1584. (in Chinese) [本文引用: 1]
KHAMESIPOURF, MOMTAZH, MAMOREHMA . Occurrence of virulence factors and antimicrobial resistance in Pasteurella multocida strains isolated from slaughter cattle in Iran. Frontiers in Microbiology, 2014,5:536. [本文引用: 1]
FERNANDEZ-ROJAS MA, VACAS, REYES-LOPEZM, DE LA GARZAM, AGUILAR-ROMEROF, ZENTENOE, SORIANO- VARGASE, NEGRETE-ABASCALE . Outer membrane vesicles of Pasteurella multocida contain virulence factors. MicrobiologyOpen, 2014,3(5):711-717. [本文引用: 1]
AHMAD TA, RAMMAH SS, SHEWEITA SA, HAROUNM, EI-SAYEDL H. Development of immunization trials against Pasteurella multocida. Vaccine, 2014,32(8):909-917. [本文引用: 1]
LITSCHKOC, OLDRINID, BUDDEI, BERGERM, MEENSJ, GERARDY-SCHAHNR, BERTIF, SCHUBERTM, FIEBIGT . A new family of capsule polymerases generates teichoic acid-like capsule polymers in Gram-negative pathogens MBio, 2018,9(3):e00641-18. [本文引用: 1]
JIANGP . The studies on isolation, purification and antlbiofilm activities of the exopolysaccharide from Vibrio sp. QY 101[D]. Ocean University of China, 2011. ( in Chinese) [本文引用: 1]
GU HW, LU CP . Evaluation of the immunization of iron-regulated outer membrane proteins (iromps) of leporid Pasteurella multocida in mice model. Scientia Agricultura Sinica, 2007,40(5):1073-1078. (in Chinese) [本文引用: 1]
KATECHAKISN, MARAKIS, DRAMITINOUI, MAROLACHAKIE, KOUTLAC, IOANNIDOUE . An unusual case of Pasteurella multocida bacteremic meningitis. Journal of Infection and Public Health, 2019,12(1):95-96. [本文引用: 1]
TUN AE, BENEDICENTIL, GALBAN EM . Pasteurella multocida meningoencephalomyelitis in a dog secondary to severe periodontal disease Clinical Case Reports,2018,6(6):1137-1141. [本文引用: 1]
PAKS, VALENCIAD, DECKERJ, VALENCIAV, ASKAROGLUY . Pasteurella multocida pneumonia in an immunocompetent patient: case report and systematic review of literature Lung India Official Organ of Indian Chest Society, 2018,35(3):237. [本文引用: 1]
CARTER GR . The type specific capsular antigen of Pasteurella multocida. Canadian Journal of Medical Science, 1952,30(1):48-53. [本文引用: 1]
RIMLER RB, RHOADES KR . Serogroup F. a new capsule serogroup of Pasteurella multocida. Journal of Clinical Microbiology, 1987,25(4):615-618. [本文引用: 1]
PANDITt KK, SMITH JE . Capsular hyaluronic acid in Pasteurella multocida type A and its counterpart in type D. Research in Veterinary Science, 1993,54(1):20-24. [本文引用: 1]
DEANGELIS PL, GUNAY NS, TOIDAT, MAO WJ, LINHARDT RJ . Identification of the capsular polysaccharides of type D and F Pasteurella multocida as unmodified heparin and chondroitin, respectively. Carbohydrate Research, 2002,337(17):1547-1552. [本文引用: 2]
MUNIANDY JB, MUKKURT . Virulence, purification, structure and protective properties of the putative capsular polysaccharide of Pasteurella multocida type 6:B. Aciar Proceedings, 1993: 47-54. [本文引用: 1]
WILSON BA, HOM . Pasteurella multocida: from zoonosis to cellular microbiology Clinical Microbiology Reviews, 2013,26(3):631-655. [本文引用: 1]
WILKIE IW, HARPERM, BOYCE JD, ADLERB . Pasteurella multocida: diseases and pathogenesis Springer Berlin Heidelberg, 2012,361(9):1-22. [本文引用: 1]
ASKI HS, TABATABAEIM . Occurrence of virulence-associated genes in Pasteurella multocida isolates obtained from different hosts. Microb Pathog, 2016,96:52-57. [本文引用: 1]
ZHAO ZQ, LIU QY, XI XJ, WANGL, DENGW, XUEY, ZHANG CJ . A comparative study on biological characterization of Pasteurella multocida serogroups A and D isolates from swine in China. Chinese Journal of Preventive Veterinary Medicine, 2016,38(5):366-370. (in Chinese) [本文引用: 2]
ZHAO ZQ, QIAO PY, LIU QY, XUEY, DINGK . Advances in the classification and inactivated vaccine of Pasteurella multocida. Chinese Journal of Preventive Veterinary Medicine, 2017,39(7):600-604. (in Chinese) [本文引用: 1]
XI XJ, ZHAO ZQ, XUEY, LONGT, WANGL, LIU HS . Prevalence and virulence of Pasteurella multocida in pig farms in central China. Chinese Journal of Veterinary Science, 2015,35(8):1205-1210. (in Chinese) [本文引用: 1]
LIU HS, ZHAO ZQ, XI XJ, XUEQ, LONGT, XUEY . Occurrence of Pasteurella multocida among pigs with respiratory disease in China between 2011 and 2015. Irish Veterinary Journal, 2017,70:2. [本文引用: 1]
ROSNERH, GRIMMECKE HD, KNIREL YA, SHASHKOV AS . Hyaluronic acid and a (1→4)-β-d-xylan, extracellular polysaccharides of Pasteurella multocida (Carter type A) strain 880. Carbohydrate Research, 1992,223(4):329. [本文引用: 1]
DEANGELIS PL . Hyaluronan synthases: fascinating glycosyltransferases from vertebrates, bacterial pathogens, and algal viruses Cellular and Molecular Life Sciences Cmis, 1999,56(7/8):670-682. [本文引用: 2]
TOWNLEY RA, BULOW HE . Deciphering functional glycosaminoglycan motifs in development Current Opinion in Structural Biology, 2018,50:144-154. [本文引用: 1]
DEANGELIS PL . Evolution of glycosaminoglycans and their glycosyltransferases: implications for the extracellular matrices of animals and the capsules of pathogenic bacteria Anatomical Record- advances in Integrative Anatomy and Evolutionary Biology, 2010,268(3):317-326. [本文引用: 1]
BOYCE JD, CHUNG JY, ADLERB . Genetic organisation of the capsule biosynthetic locus of Pasteurella multocida M1404 (B:2). Veterinary Microbiology, 2000,72:121-134. [本文引用: 3]
CHUNG JY, ZHANGY, ADLERB . The capsule biosynthetic locus of Pasteurella multocida A:1. Fems Microbiology Letters, 1998,166(2):289-296. [本文引用: 8]
BOYCE JD, CHUNG JY, ADLERB . Pasteurella multocida capsule: composition, function and genetics Journal of Biotechnology, 2000,83(1):153-160. [本文引用: 2]
LIUW, YANGM, XUZ, ZHENGH, LIANGW, ZHOUR, WUB, CHENH . Complete genome sequence of Pasteurella multocida HN06, a toxigenic strain of serogroup D. Journal of Bacteriology, 2012,194(12):3292-3293. [本文引用: 3]
TOWNSEND KM, BOYCE JD, CHUNG JY, FROST AJ, ADLERB . Genetic organization of Pasteurella multocida cap loci and development of a multiplex capsular PCR typing system. Journal of Clinical Microbiology, 2001,39(3):924-929. [本文引用: 4]
ROBERTS IS . The biochemistry and genetics of capsular polysaccharide production in bacteria Annual Review of Microbiology, 1996,50:285-315. [本文引用: 2]
DEANGELIS PL . Enzymological characterization of the Pasteurella multocida hyaluronic acid synthase. Biochemistry, 1996,35(30):9768-9771. [本文引用: 2]
DEANGELIS PL, WHITE CL . Identification and molecular cloning of a heparosan synthase from Pasteurella multocida type D. Journal of Biological Chemistry, 2002,277(9):7209-7213. [本文引用: 1]
DEANGELIS PL, PADGETT-MCCUEA J . Identification and molecular cloning of a chondroitin synthase from Pasteurella multocida type F. Journal of Biological Chemistry, 2000,275(31):24124-24129. [本文引用: 3]
WILLIS LM, STUPAKJ, RICHARDS MR, LOWARY TL, LIJ, WHITFIELDC . Conserved glycolipid termini in capsular polysaccharides synthesized by ATP-binding cassette transporter-dependent pathways in Gram-negative pathogens Proceedings of the National Academy of Sciences of the United States of America, 2013,110(19):7868-7873. [本文引用: 1]
WILLIS LM, WHITFIELDC . KpsC and KpsS are retaining 3-deoxy-D-manno-oct-2-uloso-nic acid (Kdo) transferases involved in synthesis of bacterial capsules Proceedings of the National Academy of Sciences of the United States of America, 2013,110(51):20753-20758. [本文引用: 1]
OVCHINNIKOVA OG, MALLETTEE, KOIZUMIA, LOWARY TL, KIMBER MS, WHITFIELDC . Bacterial beta-Kdo glycosyltransferases represent a new glycosyltransferase family (GT99) Proceedings of the National Academy of Sciences of the United States of America, 2016,113(22):E3120-3129. [本文引用: 2]
DEANGELIS PL, WHITE CL . Identification of a distinct, cryptic heparosan synthase from Pasteurella multocida types A, D, and F. Journal of Bacteriology, 2004,186(24):8529-8532. [本文引用: 1]
DEANGELIS PL . Molecular directionality of polysaccharide polymerization by the Pasteurella multocida hyaluronan synthase. Journal of Biological Chemistry, 1999,274(37):26557-26562. [本文引用: 1]
JINGW, DEANGELIS PL . Dissection of the two transferase activities of the Pasteurella multocida hyaluronan synthase: two active sites exist in one polypeptide. Glycobiology, 2000,10(9):883-889. [本文引用: 1]
KANE TA, WHITE CL, DEANGELIS PL . Functional characterization of PmHS1, a Pasteurella multocida heparosan synthase. Journal of Biological Chemistry, 2006,281(44):33192-33197. [本文引用: 1]
LINHARDT RJ, DORDICK JS, DEANGELIS PL, LIUJ . Enzymatic synthesis of glycosaminoglycan heparin Seminars in Thrombosis and Hemostasis, 2007,33(5):453-465.
TRACY BS, AVCI FY, LINHARDT RJ, DeAngelisP L . Acceptor specificity of the Pasteurella hyaluronan and chondroitin synthases and production of chimeric glycosaminoglycans Journal of Biological Chemistry, 2007,282(1):337-344. [本文引用: 1]
WILLISE . Structure and biosynthesis of capsular polysaccharides synthesized via ABC transporter-dependent processes Carbohydrate Research, 2013,378:35-44. [本文引用: 1]
STEEN JA, STEEN JA, PAULH, TORSTENS, IANW, MARINAH, BENA, BOYCE JD . Fis is essential for capsule production in Pasteurella multocida and regulates expression of other important virulence factors. PLoS Pathogens, 2010,6(2):e1000750. [本文引用: 4]
DORMAN CJ, DEIGHANP . Regulation of gene expression by histone-like proteins in bacteria Current Opinion in Genetics and Development, 2003,13(2):179-184. [本文引用: 2]
BAGCHIA . Structural characterization of Fis - a transcriptional regulator from pathogenic Pasteurella multocida essential for expression of virulence factors. Gene, 2015,554(2):249-253. [本文引用: 2]
WATT JM, SWIATLOE, WADE MM, CHAMPLIN FR . Regulation of capsule biosynthesis in serotype A strains of Pasteurella multocida. Fems Microbiology Letters, 2003,225(1):9-14. [本文引用: 1]
PENGZ, LIANGW, LIUW, WUB, TANGB, TANC, ZHOUR, CHENH . Genomic characterization of Pasteurella multocida HB01, a serotype A bovine isolate from China. Gene, 2016,581(1):85-93. [本文引用: 1]
MEGROZM, KLEIFELEO, WRIGHTA, POWELLD, HARRISONP, ADLERB, HARPERM, BOYCE JD . The RNA-binding chaperone Hfq is an important global regulator of gene expression in Pasteurella multocida and plays a crucial role in production of a number of virulence factors, including hyaluronic acid capsule. Infection and Immunity, 2016,84(5):1361-1370. [本文引用: 1]
CARMICHAEL GG . Isolation of bacterial and phage proteins by homopolymer RNA-cellulose chromatography Journal of Biological Chemistry, 1975,250(15):6160-6167. [本文引用: 1]
FELICIANO JR, GRILO AM, GUERREIRO SI, Sousa SA, Leitao JH . Hfq: a multifaceted RNA chaperone involved in virulence Future Microbiology, 2016,11(1):137-151. [本文引用: 1]
GULLIVER EL, WRIGHTA, LUCAS DD, MEGROZM, KLEIFELDO, SCHITTENHELM RB, POWELL DR, SEEMANNT, BULITTA JB, HARPERM, BOYCE JD . Determination of the small RNA GcvB regulon in the Gram-negative bacterial pathogen Pasteurella multocida and identification of the GcvB seed binding region. RNA, 2018,24(5):704-720. [本文引用: 1]
FROHLICH KS, VOGELJ . Activation of gene expression by small RNA Current Opinion in Microbiology, 2009,12(6):674-682. [本文引用: 1]
ZHOUG, GROTHT . Host responses to biomaterials and anti- inflammatory design-a brief review Macromolecular Bioscience, 2018: e1800112. [本文引用: 1]
BAT-ERDENEU, QUANE, CHANK, LEE BM, MATOOKW, LEE KY, ROSALES JL . Neutrophil TLR4 and PKR are targets of breast cancer cell glycosaminoglycans and effectors of glycosaminoglycan- induced APRIL secretion Oncogenesis, 2018,7(6):45. [本文引用: 1]
JIAN WH, WANG HC, KUAN CH, Chen MH, Wu HC, Sun JS, Wang TW . Glycosaminoglycan-based hybrid hydrogel encapsulated with polyelectrolyte complex nanoparticles for endogenous stem cell regulation in central nervous system regeneration Biomaterials, 2018,174:17-30. [本文引用: 1]
FINLANDM, SUTLIFF WD . Specific antibody response of human subjects to intracutaneous injection of pneumococcus products The Journal of Experimental Medicine, 1932,55(6):853-865. [本文引用: 1]
HOAGLAND CL, BEESON PB, GOEBEL WF . The capsular polysaccharide of the type XIV pneumococcus and its relationship to the specific substances of human blood Science, 1938,88(2281):261-263. [本文引用: 1]
ROBBINS JB, AUSTRIANR, LEE CJ, RASTOGI SC, SCHIFFMANG, HENRICHSENJ, M?KEL?P H, BROOMEC V, FACKLAMR R, TIESJEMAR H . Considerations for formulating the second-generation pneumococcal capsular polysaccharide vaccine with emphasis on the cross-reactive types within groups The Journal of Infectious Disease, 1983,148(6):1136-1159. [本文引用: 1]
HARRISON LH . Prospects for vaccine prevention of meningococcal infection Clinical Microbiology Reviews, 2006,19(1):142-164. [本文引用: 1]
ZAREI AE, ALMEHDAR HA, REDWAN EM . Hib vaccines: past, present, and future perspectives Journal of Immunology Research, 2016,2016:7203587. [本文引用: 2]
CILLóNIZC, AMAROR, TORRESA . Pneumococcal vaccination Current Opinion in Infectious Diseases, 2016,29(2):187-196. [本文引用: 1]
HARRISON LH, MOHANN, KIRKPATRICKP . Meningococcal group A, C, Y and W-135 conjugate vaccine Nature Reviews Drug Discovery, 2010,9(6):429-430. [本文引用: 1]
LIU XL, HAO YQ, GUOY . Progress on capsular polysaccharides of staphylococcus aureus isolated from cow mastitis. Progress in Veterinary Medicine, 2011,32(3):117-120. (in Chinese) [本文引用: 1]
HEN, WANG CF, YANG HJ, HE HB, YANG SH, WANG LQ, GAO YD, ZHONG JF . Screening an attenuated strain and immunogenicity in mice of a bovine mastitis staphylococcus aureus mutant. Scientia Agricultura Sinica, 2010,43(10):2174-2181. (in Chinese) [本文引用: 1]
FATTOMA, FULLERS, PROPSTM, WINSTONS, MUENZL, HED, NASOR, HORWITHG . Safety and immunogenicity of a booster dose of Staphylococcus aureus types 5 and 8 capsular polysaccharide conjugate vaccine (StaphVAX) in hemodialysis patients. Vaccine, 2004,23(5):656-663. [本文引用: 1]
ZHANGZ . Study on protein conjugate polysaccharide vaccine of Staphylococcus aureus for bovine mastitis[D]. Gansu Agricultural University, 2017. ( in Chinese) [本文引用: 1]
LATTAR SM, NOTO LLANAM, DENO?LP, GERMAINS, BUZZOLAF R, LEEJ C, SORDELLID O . Protein antigens increase the protective efficacy of a capsule-based vaccine against Staphylococcus aureus in a rat model of osteomyelitis. Infection and Immunity, 2014,82(1):83-91. [本文引用: 1]
 ,1,21
,1,21 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT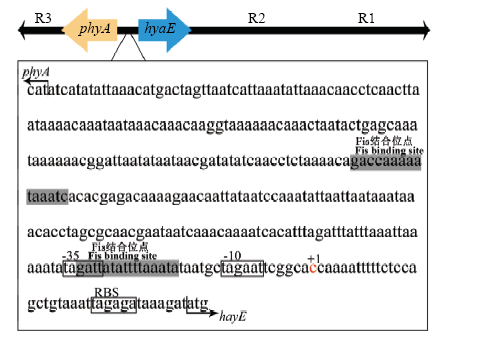 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}