The Variation/Heteroplasmy of Chicken Mitochondrial ND1 Gene and Its Association with Traits
XU YuanYuan1, HOU LingLing2, JI JieFei1, WANG HuanJie1, LI SiLu1, CHEN Wen1, KANG XiangTao1, HUANG YanQun,11 College of Livestock Husbandry and Veterinary Engineering, Henan Agricultural University, Zhengzhou 450002 2 Xuchang Animal Husbandry Bureau (Municipal Animal Health Inspection Institute) , Xuchang 461099, Henan
Abstract 【Objective】 The purpose of this study was to investigate the heteroplasmy of mitochondrial ND1 gene in chickens and to study the distribution of mitochondrial heteroplasmy among different breeds and Gushi chicken resource population, and furthermore, to analysis the effects of mitochondrial heteroplasmy on valuable economic traits. 【Method】 DNA extracted from blood samples of eight chicken breeds including Gushi chicken, Arbor Acres broiler chicken, Henan Cockfighting chicken, Lushi chicken, White Leghorn chicken, White Plymouth Rock chicken, Silkie chicken and Tibetan chicken, were used to study the distribution of mt.A4589G mutation/ heteroplasmy in natural populations by PCR-RFLP. In addition, DNA extracted from blood samples of Gushi chicken resource population constructed previously were used to study the distribution of mt.A4589G mutation/ heteroplasmy in Gushi chicken resource population by PCR-RFLP and to study the association analysis between mt.A4589G mutation/ heteroplasmy and F2 traits of Gushi chicken resource population, including carcass traits, growth traits, meat quality indicators and blood biochemical indicators, thereby grasping the heteroplasmy of chicken mitochondrial ND1 gene and its potential effects; 【Result】1) Among eight breeds, one synonymous mutation mt.A4589G was detected from the full length of mitochondrial ND1 gene, and its distribution showed obvious variety characteristics. The dominant alleles of exotic breeds such as broiler chicken, White Plymouth Rock chicken and White Leghorn chicken were A, while domestic local breeds such as Silky chicken, Henan Cockfighting chicken and Lushi chicken were G. (2) The heteroplasmy of mt.A4589G in different breeds and Gushi chicken resource population was detected by PCR-RFLP. Among eight breeds, only five AG heterogeneous individuals were detected in Gushi chicken, Henan Cockfighting chicken and broiler chicken. In the Gushi chicken resource population, mt.A4589G had a wider heteroplasmy and the frequencies of heteroplasmy in F0, F1 and F2 generations were 0.81, 0.41 and 0.70, respectively. (3) The correlation analysis between mt.A4589G heteroplasmy and F2 traits of Gushi chicken resource population showed that mt.A4589G heteroplasmy was significantly correlated with abdominal fat, sebum thickness eight-week-old shank length, diameter of leg muscle fiber and glucose levels in the blood (P<0.05). (4) The correlation analysis between the levels of heteroplasmy of mt.A4589G and F2 traits of Gushi chicken resource population indicated that the levels of heteroplasmy was significantly correlated with carcass traits, growth traits and serum biochemical indicators (P<0.05). 【Conclusion】 mt.A4589G is a heteroplasmic variation, and its heteroplasmic distribution exhibits obvious variety characteristics. The mt.A4589G heteroplasmy of the natural population occurs less frequently, while mt.A4589G has a wider heteroplasmy in the Gushi chicken resource population. Furthermore, through the correlation analysis between the heteroplasmy of mt.A4589G and the F2 traits of Gushi chicken resource population, it was found that the mt.A4589G mutation/heterogeneity was significantly correlated with the abdominal fat and the glucose level in the blood (P<0.05). Keywords:chicken;ND1 gene;PCR-RFLP;heteroplasmy;association analysis
PDF (2113KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 徐媛媛, 侯玲灵, 姬杰菲, 王焕杰, 李思露, 陈文, 康相涛, 黄艳群. 鸡线粒体ND1基因变异/异质性及其与性状的关联分析[J]. 中国农业科学, 2019, 52(16): 2880-2890 doi:10.3864/j.issn.0578-1752.2019.16.013 XU YuanYuan, HOU LingLing, JI JieFei, WANG HuanJie, LI SiLu, CHEN Wen, KANG XiangTao, HUANG YanQun. The Variation/Heteroplasmy of Chicken Mitochondrial ND1 Gene and Its Association with Traits[J]. Scientia Acricultura Sinica, 2019, 52(16): 2880-2890 doi:10.3864/j.issn.0578-1752.2019.16.013
Table 2 表2 表2线粒体mt.A4589G变异/异质性在8个鸡品种的分布 Table 2Distribution of mitochondrial mt.A4589G variation/heteroplasmy in eight breeds of chicken
基因型 Genotype
频率Frequency
品种(样本数)Breed 1)(sample number)
GS (10)
BC (12)
HC (8)
LS (10)
WL (10)
WP (10)
SK (10)
TB (10)
AA
0.375
0.0(0)
0.75(9)
0.0(0)
0.1(1)
0.7(7)
1.0(10)
0.0(0)
0.3(3)
AG 2)
0.063
0.2(2)
0.08(1)
0.25(2)
0.0(0)
0.0(0)
0.0(0)
0.0(0)
0.0(0)
GG
0.562
0.8(8)
0.17(2)
0.75(6)
0.9(9)
0.3(3)
0.0(0)
1.0(10)
0.7(7)
1) The capital letters in the table indicate different breeds of chicken: GS.Gushi chicken;BC.Arbor Acres broiler chicken;HC.Henan Cockfighting chicken;LS.Lushi chicken;WL.White Leghorn chicken;WP.White Plymouth Rock chicken;SK.Silkie chicken;TB.Tibetan chicken. 2) AG represents the genotype of a heteroplasmic individual 1)表格中的大写字母分别表示不同的鸡种:GS.固始鸡;BC.肉鸡;HC.河南斗鸡;LS.卢氏鸡;WL.白来航;WP.白洛克;SK.丝羽乌骨鸡;TB.藏鸡。2)AG代表异质性个体的基因型
1)左侧是利用PCR-RFLP法检测到的5个异质性个体的品种及异质性程度,右侧是通过对其PCR产物进行Sanger测序验证得到的峰图(GS1、GS2—固始鸡,BC1—肉鸡,HC1、HC2—河南斗鸡)。2)红色箭头处为mt.A4589G位点 Fig. 5Validation of 5 heteroplasmic individuals detected by PCR-RFLP using Sanger sequencing
1) Left is the breeds and heteroplasmic level of the five heterogeneous individuals detected by the PCR-RFLP method, and right is the Sanger sequencing peak map corresponding samples (GS1, GS2 - Gushi chicken, BC1 - Broiler chicken, HC1, HC2 - Henan Cockfighting chicken). 2) The red arrow is the mt.A4589G site
Table 4 表4 表4mt.A4589G变异/异质性与固始鸡F2代资源群性状的关联分析(包含AA和AG型的家系) Table 4Correlation analysis between mt.A4589G variation/heteroplasmy and traits of Gushi chicken F2 generation resource group (families containing AA and AG genotypes)
性状Traits
AA型 AA genotype(n=63)
AG型 AG genotype(n=69)
P值P-value
出生重 BW(g)
29.495±0.200
30.180±0.194
0.0232
腿肌纤维直径 FDL (cm)
37.831±0.925
35.047±0.827
0.0388
8周胸深 CD8(cm)
6.251±0.109
6.644±0.105
0.0159
8周胸骨长 BBL8(cm)
8.718±0.079
8.983±0.077
0.0259
8周胫长 SL8(cm)
7.749±0.078
8.012±0.077
0.0266
8周胫围 SG8(cm)
3.347±0.027
3.421±0.025
0.0644
胸肌pH BpH
6.189±0.038
6.080±0.036
0.0553
盲肠长度 CL(cm)
16.453±0.262
15.689±0.256
0.0538
肌胃重率 GR(%)
2.149±0.036
2.052±0.035
0.0685
The values corresponding to the two columns AA and AG are indicated by LSM±S.E AA型、AG型两列对应的数值以LSM±S.E表示
Table 5 表5 表5mt.A4589G变异/异质性与固始鸡F2代资源群性状的关联分析(包含GG和AG型的家系) Table 5Correlation analysis between mt.A4589G variation/heteroplasmy and traits of Gushi chicken F2 generation resource group (families containing GG and AG genotypes)
性状 Traits
AG型 AG genotype (n=488)
GG型 GG genotype (n=158)
P值 P-value
腹脂 AF(g)
8.618±0.498
6.310±0.924
0.0292
腹脂率 AFR(%)
0.936±0.051
0.692±0.096
0.0264
皮下脂肪厚度 SFT(cm)
0.452±0.007
0.486±0.014
0.0339
葡萄糖 GLU(mmol·L-1)
8.617±0.164
9.471±0.319
0.0191
乳酸脱氢酶 LDH(U·L-1)
2811.195±21.164
2707.856±41.082
0.0277
盲肠长度 CL(cm)
16.125±0.117
15.619±0.218
0.0427
The values corresponding to the two columns GG and AG are indicated by LSM±S.E GG型、AG型两列对应的数值以LSM±S.E表示
Table 6 表6 表6mt.A4589G中mt.4589A的比例等级与固始鸡资源群F2代性状的关联分析 Table 6Correlation analysis between different grades of the proportion of mt.4589A in mt.A4589G and traits of Gushi chicken F2 generation resource population
性状 Traits
1 级=0% (n=165)
0%<2级≤15% (n=226)
15%<3级≤45% (n=247)
45%<4级<100% (n=67)
5级=100% (n=66)
P值P-value
腹脂重Abdominal fat (g)
5.968±0.929A
8.857±0.768
7.313±0.720a
12.004±1.392Bb
9.448±1.897
0.0048
腹脂率Abdominal fat rate(%)
0.668±0.096A
0.958±0.080
0.811±0.075a
1.284±0.145Bb
1.055±0.197
0.0067
腿肌率Leg muscle rate(%)
32.569±0.143
32.587±0.118a
32.117±0.110b
32.506±0.213
32.087±0.296
0.0157
脾重Spleen weight (g)
2.823±0.088a
2.874±0.073
2.948±0.068
3.294±0.132b
3.056±0.180
0.0661
脾率Spleen rate(%)
0.212±0.006
0.214±0.005
0.220±0.005
0.244±0.010
0.234±0.013
0.0717
肌胃率Masticatory stomach rate (%)
2.056±0.023
2.063±0.019
2.075±0.018
2.030±0.035
2.181±0.047
0.0984
盲肠长度Cecal length(cm)
15.508±0.212
16.196±0.176
15.884±0.164
15.980±0.322
16.737±0.438
0.0410
屠宰率Slaughter rate(%)
89.586±0.170
89.863±0.142
89.526±0.131
90.184±0.255
90.182±0.343
0.0910
8周胫长8-weeks shank length(cm)
7.830±0.054
7.945±0.045
7.940±0.042
8.076±0.082
7.766±0.110
0.0406
12周胸深12-weeks chest depth(cm)
7.985±0.063
7.913±0.052
7.814±0.049
8.019±0.095
7.743±0.129
0.0680
12周胸角12-week chest angle(°)
78.515±0.340
78.418±0.283
79.275±0.263
79.763±0.512
79.225±0.694
0.0704
葡萄糖GLC(mmol·L-1)
9.472±0.319
8.576±0.251
8.911±0.239
7.878±0.458
7.887±0.677
0.0370
淀粉酶Amylase(U·L-1)
410.247±18.096a
405.599±14.427a
423.412±13.735
504.657±25.784b
519.791±40.013
0.0106
乳酸脱氢酶 LDH(U·L-1)
2714.742±40.803
2761.307±32.411
2845.511±30.532
2827.621±58.904
2820.090±85.306
0.0996
1) In the same line, different small letters means significantly (P<0.05), different capital letters means highly significantly (P<0.01). 2) In the table, the heteroplasmic ratio of 0 represents GG genotype, 100% represents AA genotype, and the heteroplasmic ratio= Base A / (Base A + Base G)×100% 1)同一行不同的小写字母表示差异显著(P<0.05),不同的大写字母表示差异极显著(P<0.01)。2)表中异质性比值为0代表的是GG基因型,100%代表的是AA基因型,异质性比值=碱基A/(碱基A+碱基G)×100%
GORMANG S, SCHAEFERA M, NGY, GOMEZN, BLAKELYE L, ALSTONC L, FEENEYC, HORVATHR, YU-WAI-MANP, CHINNERYP F, TAYLORR W, TURNBULLD M, MCFARLANDR . Prevalence of nuclear and mitochondrial DNA mutations related to adult mitochondrial disease Annals of Neurology, 2015,77(5):753-759. [本文引用: 1]
BLACKSTONEN W . Book Review: Essential Cell Biology: An introduction to the molecular biology of the cell Quarterly Review of Biology, 1998,73(4). [本文引用: 1]
CROWM T, MANIK, NAMY J, KITSISR N . The mitochondrial death pathway and cardiac myocyte apoptosis Circulation Research, 2004,95(10):957-970. [本文引用: 1]
TAANMANJ W . The mitochondrial genome: structure, transcription, translation and replication Biochimica et Biophysica Acta, 1999,1410(2):103-123. [本文引用: 1]
KUKATC, WURMC A, SPAHRH, FALKENBERGM, LARSSONN G, JAKOBSS . Super-resolution microscopy reveals that mammalian mitochondrial nucleoids have a uniform size and frequently contain a single copy of mtdna Proceedings of the National Academy of Sciences of the United States of America, 2011,108(33):13534-13539. [本文引用: 2]
YEK, LUJ, MAF, KEINANA, GUZ . Extensive pathogenicity of mitochondrial heteroplasmy in healthy human individuals Proceedings of the National Academy of Sciences of the United States of America, 2014,111(29):10654-10659. [本文引用: 1]
STEWARTJ B, CHINNERYP F . The dynamics of mitochondrial DNA heteroplasmy: implications for human health and disease Nature Reviews Genetics, 2015,16(9):530-542. [本文引用: 1]
YUSNITAY, NORSIAHM D, RAHMANA J . Mutations in mitochondrial NADH dehydrogenase subunit 1 (mtnd1) gene in colorectal carcinoma The Malaysian Journal of Pathology, 2010,32(2):103-110. [本文引用: 1]
GIULIANIC, BARBIERIC, LIM, BUCCIL, MONTID, PASSARINOG, LUISELLID, FRANCESCHIC, STONEKINGM, GARAGNANIP . Transmission from centenarians to their offspring of mtdna heteroplasmy revealed by ultra-deep sequencing Aging, 2014,6(6):454-467. [本文引用: 1]
LOUTRER, HECKELA M, JEANDARDD, TARASSOVI, ENTELISN . Anti-replicative recombinant 5S rrna molecules can modulate the mtdna heteroplasmy in a glucose-dependent manner PLoS One, 2018,13(6):e0199258. [本文引用: 1]
SCHONE A, DIMAUROS, HIRANOM . Human mitochondrial DNA: roles of inherited and somatic mutations Nature Reviews Genetics, 2012,13(12):878-890. [本文引用: 1]
YUE P, BENNETTM R . Mitochondrial DNA damage and atherosclerosis Trends in Endocrinology & Metabolism Tem, 2014,25(9):481-487. [本文引用: 1]
DANCAUSEK N, VILARM G, STEFFYR, LUMJ K . Characterizing genetic diversity of contemporary pacific chickens using mitochondrial DNA analyses PLoS One, 2011,6(2):e16843. [本文引用: 1]
LIUY P, WUG S, YAOY G, MIAOY W, LUIKARTG, BAIGM, BEJA-PEREIRAA, DINGZ L, PALANICHAMYM G, ZHANGY P . Multiple maternal origins of chickens: out of the Asian jungles Molecular Phylogenetics and Evolution, 2006,38(1):12-19. [本文引用: 1]
MWACHAROJ M, BJORNSTADG, MOBEGIV, NOMURAK, HANADAH, AMANOT, JIANLINH, HANOTTEO . Mitochondrial DNA reveals multiple introductions of domestic chicken in East Africa Molecular Phylogenetics and Evolution, 2011,58(2):374-382. [本文引用: 1]
REVAYT, BODZSARN, MOBEGIV E, HANOTTEO, HIDASA . Origin of Hungarian indigenous chicken breeds inferred from mitochondrial DNA D-loop sequences Animal Genetics, 2010,41(5):548-550. [本文引用: 1]
KOMIYAMAT, IKEOK, GOJOBORIT . The evolutionary origin of long-crowing chicken: its evolutionary relationship with fighting cocks disclosed by the mtdna sequence analysis Gene, 2004,333:91-99. [本文引用: 1]
GUANX, GENGT, SILVAP, SMITHE J . Mitochondrial DNA sequence and haplotype variation analysis in the chicken (Gallus gallus) Journal of Heredity, 2007,98(7):723-726. [本文引用: 1]
LUW W, HOUL L, ZHANGW W, ZHANGP F, CHENW, KANGX, HUANGY . Study on heteroplasmic variation and the effect of chicken mitochondrial ND2 Mitochondrial DNA Part A, DNA Mapping, Sequencing, and Analysis, 2016,27(4):2303-2309. [本文引用: 3]
PRUETTC L, WINKERK . The effects of sample size on population genetic diversity estimates in song sparrows Melospiza melodia Journal of Avian Biology, 2008,39(2):252-256. [本文引用: 1]
TRASKJ A, MALHIR S, KANTHASWAMYS, JOHNSONJ, GARNICAW T, MALLADIV S, SMITHD G . The effect of SNP discovery method and sample size on estimation of population genetic data for Chinese and Indian rhesus macaques (Macaca mulatta) Primates; Journal of Primatology, 2011,52(2):129-138. [本文引用: 1]
IOMMARINIL, GHELLIA, TROPEANOC V, KURELACI, LEONEG, VIDONIS, LOMBESA, ZEVIANIM, GASPARREG, PORCELLIA M . Unravelling the effects of the mutation m.3571insc/MT-ND1 on respiratory complexes structural organization International Journal of Molecular Sciences, 2018,19(3):764-780. [本文引用: 2]
BARADARANR, BERRISFORDJ M, MINHASG S, SAZANOVL A . Crystal structure of the entire respiratory complex I Nature, 2013,494(7438):443-448. [本文引用: 1]
ANTONICKAH, OGILVIEI, TAIVASSALOT, ANITORIR P, HALLERR G, VISSINGJ, KENNAWAYN G, SHOUBRIDGEE A . Identification and characterization of a common set of complex I assembly intermediates in mitochondria from patients with complex I deficiency The Journal of Biological Chemistry, 2003,278(44):43081-43088. [本文引用: 1]
TURRENSJ F . Mitochondrial formation of reactive oxygen species The Journal of Physiology, 2003,552(2):335-344. [本文引用: 1]
MAGNACCAK N, BROWNM J . Mitochondrial heteroplasmy and DNA barcoding in Hawaiian Hylaeus (Nesoprosopis) bees (Hymenoptera: Colletidae) BMC Evolutionary Biology, 2010,10(1):174. [本文引用: 1]
REINERJ E, KISHORER B, LEVINB C, ALBANETTIT, BOIREN, KNIPEA, HELMERSONK, DECKMANK H . Detection of Heteroplasmic Mitochondrial DNA in Single Mitochondria PLoS One, 2010,5(12):e14359. [本文引用: 1]
RAMOSA, SANTOSC, MATEIUL, GONZALEZMDEL M, ALVAREZL, AZEVEDOL, AMORIMA, ALUJAM P . Frequency and pattern of heteroplasmy in the complete human mitochondrial genome PLoS One, 2013,8(10):e74636. [本文引用: 1]
WALLACED C . Mitochondrial DNA mutations in disease and aging Environmental and Molecular Mutagenesis, 2010,51(5):440-450. [本文引用: 1]
PATANANANA N, WUT H, CHIOUP Y, TEITELLM A . Modifying the Mitochondrial Genome Cell Metabolism, 2016,23(5):785-796. [本文引用: 1]
SIMOND K, MAYEUXR, MARDERK, KOWALLN W, BEALM F, JOHNSD R . Mitochondrial DNA mutations in complex I and trna genes in Parkinson's disease Neurology, 2000,54(3):703-709. [本文引用: 1]
BROWNM D, ZHADANOVS, ALLENJ C, HOSSEINIS, NEWMANN J, ATAMONOVV V, MIKHAILOVSKAYAI E, SUKERNIKR I, WALLACED C . Novel mtdna mutations and oxidative phosphorylation dysfunction in Russian LHON families Human Genetics, 2001,109(1):33-39. [本文引用: 1]
SUDOYOH, SURYADIH, LERTRITP, PRAMOONJAGOP, LYRAWATID, MARZUKIS . Asian-specific mtdna backgrounds associated with the primary G11778A mutation of Leber's hereditary optic neuropathy Journal of Human Genetics, 2002,47(11):594-604. [本文引用: 1]
HOWELLN, BINDOFFL A, MCCULLOUGHD A, KUBACKAI, POULTONJ, MACKEYD, TAYLORL, TURNBULLD M . Leber hereditary optic neuropathy: Identification of the same mitochondrial NDI mutation in six pedigrees American Journal of Human Genetics, 1991,49(5):939-950. [本文引用: 1]
ABU-AMEROK K, ALZAHRANIA S, ZOUM, SHIY . High frequency of somatic mitochondrial DNA mutations in human thyroid carcinomas and complex I respiratory defect in thyroid cancer cell lines Oncogene, 2005,24(8):1455-1460. [本文引用: 1]
WANGJ W, CHENW, KANGX T, HUANGY Q, TIANY D, WANGY B . Identification of differentially expressed genes induced by energy restriction using annealing control primer system from the liver and adipose tissues of broilers Poultry Science, 2012,91(4):972-978. [本文引用: 1]
MERLOPICH M, RAULEN, CATANIL, FAGIOLIM E, FAENZAI, COCCOL, LENAZG . Increased transcription of mitochondrial genes for Complex I in human platelets during ageing FEBS Letters, 2004,558(1-3):19-22. [本文引用: 1]
LUW W, HOUL L, ZHANGW W, ZHANGP F, CHENW, KANGX, HUANGY . Study on heteroplasmic variation and the effect of chicken mitochondrial ND2 Mitochondrial DNA Part A, DNA Mapping, Sequencing, and Analysis, 2016,27(4):2303-2309. [本文引用: 1]
 ,1
,1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT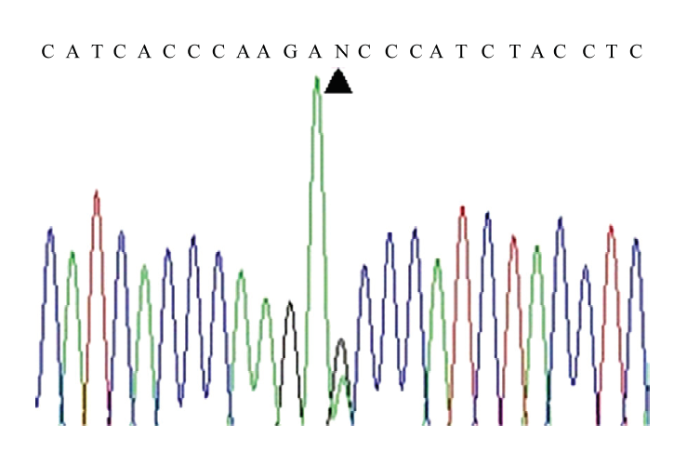 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT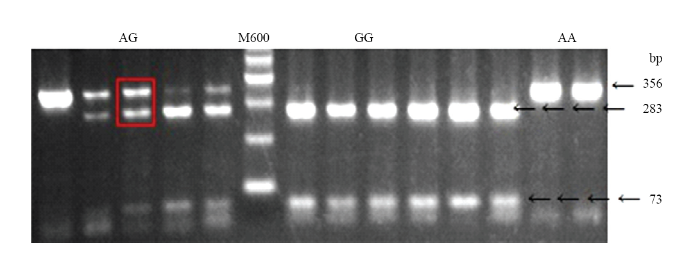 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT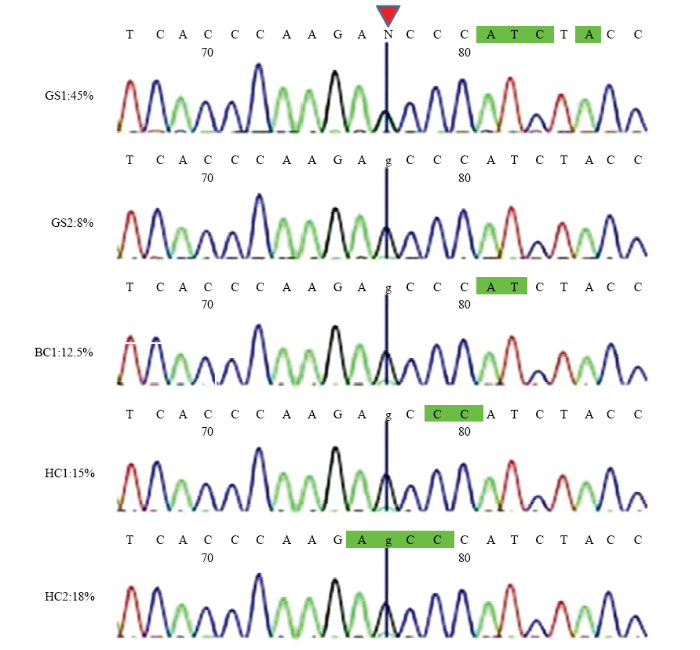 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}