 ,1
,1Expression and Structural Analysis of SC MI390-5p and Its Target Genes in Potato Response to Low Temperature
XIE Jie1, WANG Ming1, DING HongYing1, LI Qing1, WANG WanXing2, XIONG XingYao2, QIN YuZhi,1ͨѶ����:
���α༭: ������
�ո�����:2019-01-3��������:2019-05-13�����������:2019-07-01
| ��������: |
Received:2019-01-3Accepted:2019-05-13Online:2019-07-01
����� About authors
�,Tel��0731-84618171;E-mail��358562505@qq.com

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (4805KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
л��, ����, ����ӳ, ����, ������, ����ҫ, ����֥. ������������Ӧ��ScmiR390-5p����л��������ṹ����[J]. �й�ũҵ��ѧ, 2019, 52(13): 2295-2308 doi:10.3864/j.issn.0578-1752.2019.13.009
XIE Jie, WANG Ming, DING HongYing, LI Qing, WANG WanXing, XIONG XingYao, QIN YuZhi.
0 ����
���о����塿MicroRNA390��miR390��������һ�������Ҹ߶ȱ��صļ���,�㷺����ֲ��������������������ټ����γɡ��������γɼ�в����Ӧ [1]�������������ظ�����ĵ���ø�ڲ���ֲ����Է�Ӧ���������ؼ����ص��ź�ת���ȹ����з�������[2]�����÷�������ѧ�ֶ�̽��в����ػ���ı�����������MicroRNA��miRNAs�����ص�ģʽ,�Խ������û����ֶν������澳������Ʒ�ֵ�ѡ�������������Ҫ�����塣��ǰ���о���չ��miRNA��һ��ת¼���������,ͨ����mRNA�����и����ɷ�����������ưб����ı���[3],ʵ�ֶԻ���в���źŵ���Ӧ[4,5,6]���о�����miR390����RISC��ͨ����Two-Hit��������miR390�İ���λ�㣩�Ļ���������AtTAS1��,�յ���������tasiARF��tasiRNA����ARF����,������tasiARF��������,������������AtARF2/3/4��,��mRNA�����и��������ʹAGO7��Ĭ [7]��ֲ������Ȼ�����¾����ܵ��䡢����NaCl�����ء��ؽ����ȶ��ַ��������ص�в��,Ŀǰ,��miR390��Ӧ���¼����ַ�����в�����в����о���ľ���ڵ���в����miR390�ı������½�[8],��ɽ��Եֲ���ɽ���ӽ���miR390ͨ�����ذл���ı���Ե���������Ӧ[9],miR390�ڵ���в�ȴ�������������ҶƬ�б�������,���������miR390��ë������4����´��������з�����õı��������Ϊ����[10]�����̲ݽ��иɺ�в�Ⱥͻָ���ˮ�����о���ʾ,miR390�ڲ�ͬ�ɺ������³��ֲ������[11],����miR390���ܲ���ֲ���NaClв����Ӧ���̡��о���Աͨ������NaClв�������ڸ���֯�Ľ������Ŀ�,��miR390�İб���������ø�����仯[12],�ڸ���[13]���̲�[14]�����Ͻ�[15]�ʹ���[16]��ֲ���ж�������������ø��NaClв���±����������ı�,�Ʋ�miR390��ͨ��������������ø�ı���������ֲ���NaClв����Ӧ;���о��з���,miR390������ͨ������������RDR6,����ֲ���NaClв����Ӧ [17]������в��ͬ��Ӱ��miR390�ı���,BA�������ٽ���ȫ���ǡ���ݮ�Թ�����miR390�ı���;IAAҲ�������������miR390�ı�����[18]��YOON��[19]�о�ָ��,�ڸ�IAAŨ���£�10 ��mol��L-1��50 ��mol��L-1������12��24 h,���Ͻ�miR390�ı���ˮƽ��������,���ڵ�Ũ�ȣ�10 nmol��L-1���´�����ͬʱ��,������û���������졣�ؽ���в�������������������е���ҪӰ������֮һ,DING��[20]�о�����Cdв����miR390������ת������ϵ��OsHMA2��OsNramp5�ı�����ǿ,�ٽ���ˮ�����ϲ����ӻ����������ӡ���Cdв����������ϵ��miR390�������µ�[21]��������miR390-TAS3�ܱ���ת¼��,�Ʋ�miR390ͨ����������ֲ�������������Alв����������[22]�����о���ScmiR390��һ��Ԥ��л���SCLRRK1�������������ظ�����ĵ���ø,���൰��ø��ֲ��Ӧ���澳�ͷ�����ع��̵���Ҫø�ࡣLEE��[23]����ˮ���и����������ظ�����ĵ���øOsRLK1�����º�NaClв���յ�����,JUNG��[24]����������CALRR1�ڸ�NaCl�������ᣨABA���ȷ�����в���±�������о�����㡿ScmiR390/SCLRRK1����ģ����Ӧ���µȷ�����в�ȵ��о�Ŀǰ��δ�����������о���ͼͨ������RT-qPCR����¡��RLM-5�� RACE�ȷ�������ѧ�ֶ�̽����������ScmiR390/SCLRRK1����ģ������ģʽ���������Ĺؼ����⡿��¡������SCLRRK1���нṹ�빦��,���о�ScmiR390/SCLRRK1����ģ���ڵ��¡�NaCl��6-BA��ABA��GA3��IAA��PEGв���µ���Ӧ���,ͬʱ����ScmiR390��SCLRRK1����������ͬ��֯�еĵ��ع�ϵ��1 �����뷽��
1.1 ֲ�����
������GS393��Solanum commersonii-LZ3.4��,��������˹���������������ʿ⣨http://www.ars-grin. gov/npgs/acc/acc_queries.html���������ͺ�����������,��-4�����,��ѱ�������-6�档GS393�����簴���������19�棩,������������3��4�����ڻ����¡��RLM-5��RACE���顣1.2 ���鴦��
2016��12��,��60ƿ������һ�µ������������ѱ����5�棩��˪��������-4�棩,���������������һ�µ������������ա�������ѱ��ǰ��ȡ�����ΪCK;5��ѱ����1��2��3��Ļ����ΪTLX1������ΪCLX1��;5��ѱ����8��9��10��Ļ����ΪTLX2������ΪCLX2��;5��ѱ��10 d��-4��˪��������ǰ1��2��3 h�Ļ����ΪTLH1������ΪCLH1��;5��ѱ��10 d��-4��˪��������ǰ7��8��9 h�Ļ����ΪTLH2������ΪCLH2����2016��12�µײ���,����������,��2017��4�·ֱ�ȡ��������Ҷ�����뾥�Ϳ龥,����ScmiR390-5p��SCLRRK1�ĸ���֯�����Ա��������2017��11�¶�180ƿ������һ�µ�������GS393������ֱ���100 mg?L-1 IAA��100 mg?L-1 ABA��100 mg?L-1 6-BA��100 mg?L-1 GA3��11.7 g?L-1 NaCl��20 g?L-1 PEGв�ȴ���,����ScmiR390-5p��SCLRRK1�ı���������������������3���ظ�,ȡ�ĺ���Һ��Ѹ���䶳, -80�泬���±����б��汸�á�1.3 ����
����Primer 5.0�������,���Ϻ�������˾�ϳ�,�����м�����Table 1
��1
��1���������������
Table 1
| ���� Primer | ���У�5���3�䣩 Sequence (5���3��) | ��; Use |
|---|---|---|
| SCLRRK1 F | ATGACACTTTTGAGGTTTCTGTTAA | SCLRRK1 CDSƬ�ο�¡ |
| SCLRRK1 R | TTAGCTTTCCGTGATTTCTTGA | Clone of SCLRRK1 CDS fragments |
| SCLRRK1 F1 | GTTTTCTTCATTTCTTGATATGGC | SCLRRK1DNAȫ������ |
| SCLRRK1 R1 | GTTGAATAGTAGCATTCCCTTAATC | Amplification of Full-length DNA of SCLRRK1 |
| SCLRRK1 F2 | GCTTGAGATCCTAACCACCCG | SCLRRK1ʵʱ����PCR |
| SCLRRK1 R2 | TCTTGCAGCATTTCTACCACCTT | Quantitative real-time PCR of SCLRRK1 |
| SCLRRK1 outer F3 | GCTGATGGCGATGAATGAATGAACACTG | RLM-5��RACE ��������� |
| SCLRRK1 outer R3 | CAACAAAGACGTTTCCCGCCAGT | Rapid amplification of cDNA 5�� ends |
| SCLRRK1 inner F4 | CGCGGATCCGAACACTGCGTTTGCTGGCTTTGATG | RLM-5��RACE ��������� |
| SCLRRK1 inner R4 | CCTGAGGCAATTTTCCGTGGAGA | Rapid amplification of cDNA 5�� ends |
| ScmiR390-5p | CCAGCGTGAAGCUCAGGAGGG | ScmiR390-5pʵʱӫ�ⶨ��PCR |
| ScmiR390-5p | CCAGCGTGCGCUAUCCAUCUU | Quantitative real-time PCR of ScmiR390-5p |
�´��ڴ�|����CSV
1.4 RNA����ȡ
��RNA����ȡ���������˾�Ķ��Ƕ��ֲ����RNA��ȡ�Լ�����ȡ,ScmiR390-5p����ȡ���������˾��miRcutemiRNA��ȡ�����Լ��С�����1.0%��֬��������Ӿ�����RNA��������,�õ¹�Eppendorf Biospectrometer? basic����ɼ���ӫ��ֹ��ȼƣ�Eppendorf Biospectromete? basic,Germany���������Ũ�Ⱥʹ��ȡ�1.5 RT-PCR
��RNA�ķ�ת¼���ñ��������ҵ���\������˾�Ľ�����ת¼�Լ��У�GoldenstarTMRT6 cDNA Synthesis Kit��,ScmiR390-5p�ķ�ת¼���������˾��miRcute��ǿ��miRNAcDNA��һ���ϳ��Լ��С�1.6 ��¡
����DNA���������Լ��д���֬�������л�������Ƭ��,��pEASY?-T5Zero Cloning Vector��������,Ȼ��ת�����˾�Trans-T1����̬ϸ����,ͨ�����װ�ɸѡPCR�������Կ�¡��DNA���������Լ��й��Ա��������ҵ���\������˾,pEASY?- T5Zero Cloning Kit���幺��̩˼�¹�˾,����ҩƷΪ������������1.7 ʵʱӫ�ⶨ��PCR
SCLRRK1��RT-qPCR�����ñ��������ҵ���\������˾��2��T5 Fast RT-qPCRMix,ScmiR390-5p��RT-qPCR�����������˾��miRcute��ǿ��miRNAӫ�ⶨ������Լ��С�ʹ��ABI��˾��7500����PCR��,96�װ��ѧĤ��ɶ���PCR��Ӧ����18S rRNA������Ϊ�ܼһ���,��ˮģ��Ϊ���Զ���,����2-����Ct��������Ա������ĸߵ͡�1.8 RLM-5�� RACE
5�� RACE����FirstChoice?RLM-RACE Kit�Լ��еķ�������,��Ӧø����Promega��˾,��ͷ�����ɱ��������ҵ���\������˾�ϳɡ�1.9 ���вⶨ������
������ί�б��������ҵ���\������˾���С�����NCBI ��https://www.ncbi.nlm.nih.gov/������BLAST�ȶԺͻ��������ORF��Ԥ��;ʹ��SMART��http://smart.embl-heidelberg.de/����SCLRRK1���нṹ��Ԥ��;ʹ��portparam��http://web.expasy.org/ protparam/�����߷���SCLRRK1����������;ʹ��TMHMM��http://www.cbs.dtu.dk/services/TMHMM/����SCLRRK1���п�Ĥ������;ʹ��signalP��http:// www.cbs.dtu.dk/services/SignalP-3.0/����SCLRRK1���ź��Ľ���Ԥ��;ʹ��NEBcutter��http://nc2.neb. com/NEBcutter2/index.php����SCLRRK1���е�������ø��λ����з���;ʹ��predictprotein��https:// www.predictprotein.org/������������������SCLRRK1�Ķ����ṹ;ʹ��SWISS-MODEL��http://www.swissmodel. expasy.org/�����߷���������SCLRRK1��ģ����������ṹģ�͡�2 ���
2.1 ScmiR390-5p������SCLRRK1��������Ӧ��������
2.1.1 ������ת¼�鼰miRNA������� ����Illumina HiSeqTM2500����ƽ̨����������ת¼���miRNA��ͨ������,����������PGSC_DM_v4.03��Ϊ�ο�������,����|log2 FC|��1��FDR��0.01�Ƚϡ�ɸѡ����������Differentially expressed genes,DEGs��,��miRBase���ݿ���бȶԡ�ע��,��������miRBase�и�miRNA���������н�������ȶ�,�ȶ��ϵ����ж���Ϊ��֪��miRNA�����Դ�Ϊ������ΪmiRNA�ı�����ͳ��,����DEGs�������������,����ѱ�����ڣ�8��9��10 d,ͼ1-A��106��miRNA�ϵ�,11��miRNA�µ�������8���ϵ�miRNA�İл������Ϊ�����µ�;3���ϵ�miRNA�İл�����ת¼������б���Ϊ�����ϵ�;11���µ�miRNA�İл�������2����ת¼������б���Ϊ�����ϵ�������˪������,����ղ��ϱȽ�,ѱ����IJ���˪���������ڣ�ǰ1��2��3 h,ͼ1-B��65��miRNA�ϵ�,238��miRNA�µ�������4���ϵ�miRNA�İл�����ת¼������б���Ϊ�����µ�;6���ϵ�miRNA�İл�����ת¼������б���Ϊ�����ϵ���ͼ1
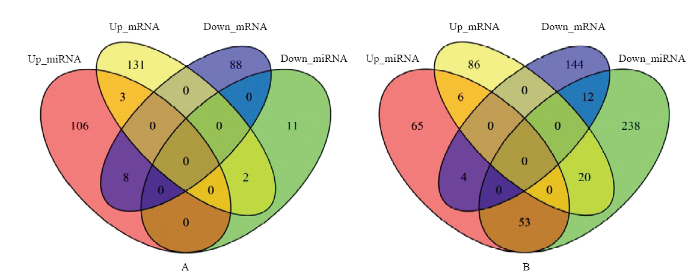 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1��ͬ���������������mRNA��miRNA����л���Աȷ���
Fig. 1Comparative analysis of mRNA, miRNA and its target genes with significant differences between different treatments
A: TLX2-CLX2; B: TLH1-CLH1
2.1.2 ScmiR390-5p�л���Ԥ�� ����http://plantgrn. noble.org/psRNATarget/�ϵķ�������miRNA�л���Ԥ��,������ת¼�������������������������������,���о���ע��ScmiR390-5p������������İл�������������2����ǰ�ڹ۲쵽ѱ�����ںͽ���˪��ǰ��ScmiR390-5p������Ϊ�ϵ�,��Ԥ��л���PGSC0003DMT400070158��SCLRRK1����PGSC0003DMT400010618��PGSC0003DMT400034044�����Ϊ�����µ�,���о�ѡ��PGSC0003DMT400070158��SCLRRK1����Ϊ����ScmiR390-5p������ػ����о�����,���л����¡�������ϵ��в����Ӧ������
Table 2
��2
��2��ͬ������ScmiR390-5p������������İл���������
Table 2
| ������ ID | TLX2/CLX2 | TLH1/CLH1 | ������ ID | ������ Gene name | TLX2/CLX2 | TLH1/CLH1 |
|---|---|---|---|---|---|---|
| �ϵ�_�µ� Up_down | �ϵ�_�µ� Up_down | �ϵ�_�µ� Up_down | �ϵ�_�µ� Up_down | |||
| ScmiR390-5p | �ϵ� Up | �ϵ� Up | PGSC0003DMT 400070158 SCLRRK1 | Leucine-rich repeat receptor kinase | �µ� Down | �µ� Down |
| �ϵ� Up | �ϵ� Up | PGSC0003DMT 400010618 | Receptor protein kinase CLAVATA1 | �µ� Down | �µ� Down | |
| �ϵ� Up | �ϵ� Up | PGSC0003DMT 400034044 | Unknown | �µ� Down | �µ� Down | |
| �ϵ� Up | �ϵ� Up | PGSC0003DMT 400009701 | Endo-1,3-1,4-beta-d-glucanase | �ϵ� Up | �ϵ� Up |
�´��ڴ�|����CSV
2.1.3 ����в����ScmiR390-5p��SCLRRK1�ı��� ͨ��������ϢѧԤ��õ�ScmiR390-5p�İл���SCLRRK1��MG763883��,RT-qPCR�Ͱ붨��PCR�����ͼ2��ʾ�����ŵ���5��ѱ��ʱ�������ScmiR390-5p�ı���������,�л���SCLRRK1�ı�������½�,��δ��������GS393��ȣ�CK��,ѱ������SCLRRK1�ı�������0.05��ͼ2-B,AΪ���գ�;δ��ѱ���IJ�����˪����-4�棩в����ScmiR390-5p�ı�������������,�л���SCLRRK1�ı����½�,���½�����û��ѱ��в�����ԣ�ͼ2-C��;ѱ������˪������ScmiR390-5p�ı����½�,Ԥ��л���SCLRRK1�ı����������,ѱ������˪����������SCLRRK1�ı������ﵽ272��ͼ2-D�����������,����в����ScmiR390-5p��SCLRRK1�ı��������ڸ�����ԡ�
ͼ2
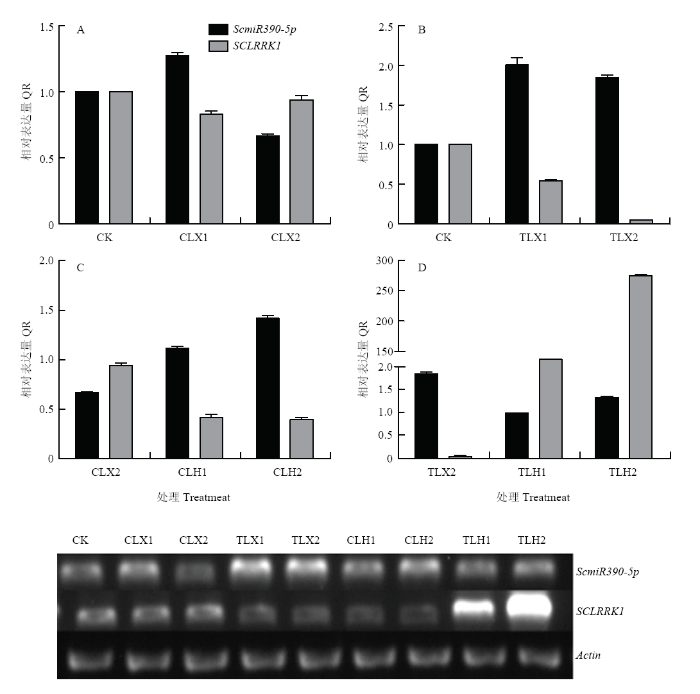 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2SCLRRK1��Ӧ���µ�ʵʱӫ�ⶨ�������Ͱ붨������
CK��δ����������ȡ�����;TLX1��5��ѱ����1��2��3��Ļ����������CLX1��;TLX2��5��ѱ����8��9��10��Ļ����������CLX2��;TLH1��5��ѱ��10 d��-4��˪��������ǰ1��2��3 h�Ļ����������CLH1��;TLH2��5��ѱ��10 d��-4��˪��������ǰ7��8��9 h�Ļ����������CLH2��
Fig. 2The analysis of the SCLRRK1 by Real-time PCR and Semi quantitative in low temperature
CK: The mixture samples without treatment; TLX1: The mixed samples which acclimated at 5�� of 1th, 2th and 3th days (control is CLX1); TLX2: The mixed samples of 8th, 9th, and 10th days after 5�� acclimation (control is CLX2); TLH1: The mixed samples acclimated at 5�� for 10 days and frozen at -4�� for the first 1th, 2th and 3th hours (control is CLH1); TLH2: The mixed samples acclimated at 5�� for 10 days and frozen at -4�� for the 7th, 8th and 9th hours (control is CLH2)
2.1.4 ����в����ScmiR390-5p����ͨ�����SCLRRK1���и�λ����ظû������ ����Ԥ������λ�����������������,���ó���PCR����Ŀ�Ļ���Ƭ��,��ͼ3��ʾ���л���SCLRRK1�����������IJ����ϴ�,�����СΪ415 bp,��Ԥ����һ��,������PCR�����н�����ֱ����T��������,ɸѡ������������˾����,��¡Ƭ��������Ԥ����һ��,ScmiR390-5p�����ڰл���SCLRRK1��GGAACTATTCCT//CCTGAGTTTƬ�Ρ�ScmiR390-5p��SCLRRK1���ڰ����ϵ,֤������������в����Ӧ�д���ScmiR390-5p/SCLRRK1����ģ�顣
ͼ3
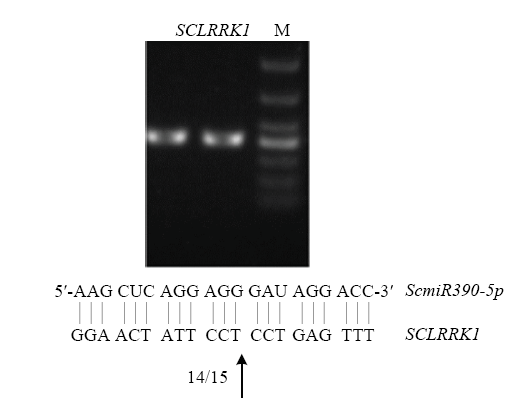 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3ScmiR390-5p��SCLRRK1����λ���RLM-RACE��֤
Fig. 3Validation of ScmiR390-5p guided cleavage of SCLRRK1 by RLM-RACE
2.2 ������SCLRRK1�����
2.2.1 ������SCLRRK1 CDS���з��� ��NCBI��EST���ݿ���������EST���ݿ���м������SCLRRK1��������Ϣ����CDS���е������������,��������������GS393��cDNA��һ��Ϊģ�����RT-PCR����,�õ�1������Լ2 685 bp������������ͼ4-A��,����������,��Ƭ�γ���Ϊ2 685 bp,��������������ε�ͬԴ��Ϊ99%,ScmiR390-5p�İ����д���CDS�����ڲ���960��981 bp,GGAACTA TTCCTCCTGAGTTT����2.2.2 ������SCLRRK1 DNA���з��� ��������������GS393��DNAΪģ�����PCR����,�õ�1��Լ3 549 bp������������ͼ4-B��������������,��Ƭ�γ���Ϊ3 564 bp,��������������ε�ͬԴ��Ϊ95%��
ͼ4
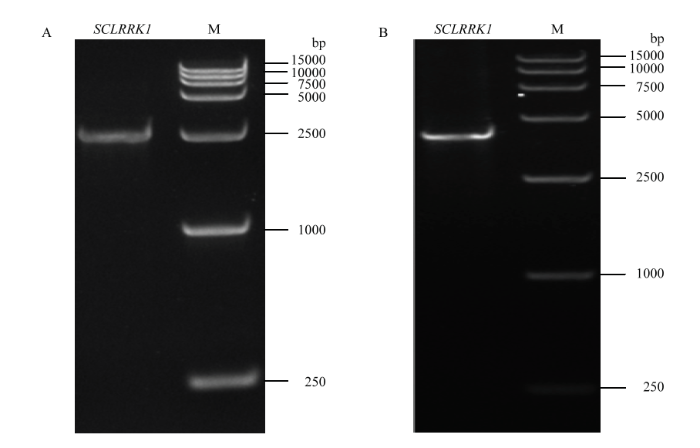 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4������SCLLRK1�����������
Fig. 4Products of amplification of SCLLRK1 gene from Solanum commersonii-LZ3.4
2.2.3 SCLRRK1�Ľṹ���� ����Portparam����������SCLRRK1��һ���ṹ���з�����ʾ���������ԼΪ223 kD,���۵ȵ��pIΪ4.92,�ܹ�����28 902��ԭ��,����ʽΪC8300H13915N2687O3542S458���հ��ᣨThr���ĺ������,�ﵽ32.18%���䲻�ȶ�ָ��Ϊ38.9,֬��ָ��Ϊ28.31,����Guruprasad��������SCLRRK1Ϊ�ȶ����ס��ܵ���ˮ��ƽ��ϵ��Ϊ0.62,Ԥ��õ���������ˮ�Ե��ס�Pedictprotein���߷���SCLRRK1�Ķ����ṹ��ʾ���-����ռ38.7%,������ռ17.23%,��-�۵�ռ8.28%,�������ռ35.79%��SWISS-MODEL���߶�SCLRRK1�������ṹ���з�����SCLRRK1������ִ������Ľṹ,��Ҫ�ɦ�-����������������,��-��������������м�������������,���������Ħ�-�۵�,��������ṹ��Ԥ��������һ�¡�
2.2.4 SCLRRK1�Ĺ�������� ͨ������ProtScale����Ԥ��SCLRRK1������ˮ��,SCLRRK1�ʰ�����л��а�������ˮ�����ֵΪ3.494,��СֵΪ-2.526,��ˮ���������Ŀ������ˮ���������Ŀ,����540��562λ�����ᣨIVLAVIGSGLAVFVCVTVVVLLF��֮�京��һ�����͵���ˮ����,�ɼ�SCLRRK1Ϊ��ˮ�Ե��ף�ͼ5����ʹ��TMHMM���߷���������SCLRRK1���п�Ĥ��������ʾSCLRRK1���������д������ԵĿ�Ĥ�ṹ����,1��539λ������λ��ϸ��Ĥ����,540��562λ������֮���γ�һ�����͵Ŀ�Ĥ������,563��694λ������λ��ϸ����,��õ�����ˮ����������������һ��,�����õ�������һ����ϸ���źŴ����йص�Ĥ���嵰�ף�ͼ6����ʹ��SignalP4.1server��������SCLRRK1�����ź���Ԥ����ʾC score��ʾ����λ���ֵ��Cֵ��,S score��ʾ�ź��ķ�ֵ��Sֵ��,Y score��ʾ�ۺϼ���λ���ֵ��Yֵ����SCLRRK1��Sƽ��ֵ����0.5,Ԥ��û���Ϊ���ڵ����Ҵ����ź��ġ���ͼ7�п������Կ���N���ź���,���е�λ��25��26λ������֮�䡣
ͼ5
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5SCLRRK1��ˮ��/��ˮ��Ԥ��
Fig. 5Hydrophobicity/hydrophilicity prediction of SCLRRK1
ͼ6
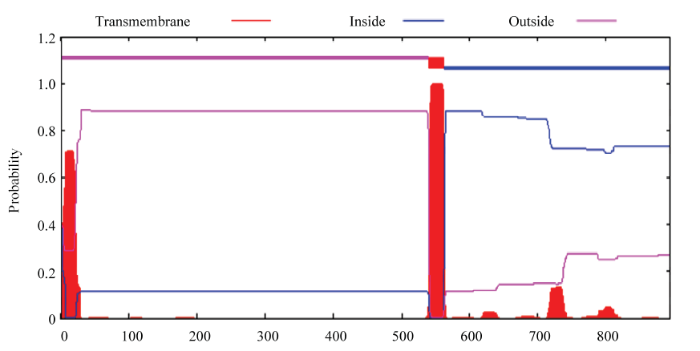 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6SCLRRK1�Ŀ�Ĥ��Ԥ��
Fig. 6Predicted transmembrane domain of prediction of SCLRRK1
ͼ7
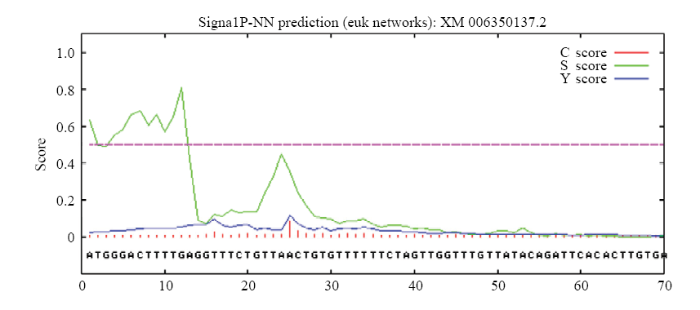 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ7SCLRRK1���ź���Ԥ��
Fig. 7Predicted signal peptide of SCLRRK1
2.2.5 SCLRRK1�İ��������з��� SMART�ṹ��Ԥ��,��ϰ��������з�����ͼ8��,��֪SCLRRK1��һ��LRR����Ser/Thr/Tyr�ĵ���ø,��2��17��������͵�566��576�İ�����京���������Ӷ�����,��90��113�������ᡢ��114��137�������ᡢ��138��162�������ᡢ��210��234�������ᡢ��330��354��������͵�426��451��������京��6��LRR����,�Ǹ�����������ظ�����,��540��562��������京��һ����Ĥ�ṹ��,��613��888��������京��1������ø����STYKc��¡�����ϵͳ��������NCBI�Ͻ��л���ע�ᣨMG763883��,������ΪSCLRRK1��
ͼ8
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ8SCLRRK1�ĺ��������м�����������
������Ϊ���Ӷ�����;���»���ΪLRR����;˫�»���Ϊ��Ĥ������;��Ӱ����Ϊ����ø����STYKc��;��Բ����ScmiR390-5p��ʶ��λ��
Fig. 8Nucleotide sequence and amino acid sequence of SCLRRK1
The box is containing Low complexity region; the one underline area is LRR domain; the two underline area is transmembrane helix region; Protein Kinase STYKc domain is shaded area; The ellipse is containing the ScmiR390-5p recognition sites
2.3 ScmiR390-5p��SCLRRK1����֯����������
��ͼ9��ʾ,ScmiR390-5p��Ҷ�еı��������,����֮,�ڿ龥�����뾥�еı������ϵ�;��SCLRRK1�ı��������ScmiR390-5p�෴,����Ҷ�еı������,�����뾥�еı������,�龥��֮���ý�������ģ����ScmiR390-5p������SCLRRK1��ϵ�����ͼ9
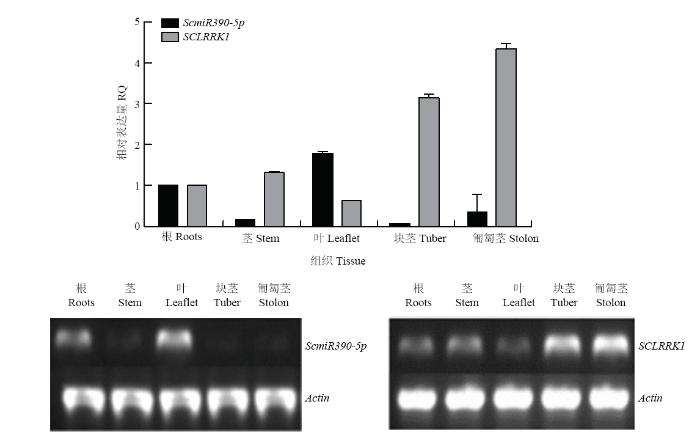 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ9 ScmiR390-5p��SCLRRK1�ڸ���֯�б����ʵʱӫ�ⶨ�������Ͱ붨������
Fig. 9The specific expression of the ScmiR390-5p and SCLRRK1 by Real-time PCR in different tissue form Solanum tuberosum and semi quantitative PCR in different tissue
2.4 ������SCLRRK1�IJ��ַ�����в����Ӧ
��������,ABA��6-BA����24 hʱScmiR390-5p���������,֮��ʲ���С���ص�;6-BA����8 h,SCLRRK1�ﵽ�����ֵ,֮��40 h��ά�ָ�λˮƽ,ABA����36 h SCLRRK1����ʲ�����������ֵ������½�;GA3��PEG��IAA����8 h ScmiR390-5p������ﵽ��ֵ,GA3��PEG��IAA���������յ�SCLRRK1����,����仯������ScmiR390-5p����Բ�ǿ;ScmiR390-5p��NaCl����8 hʱ���ֱ����½�,֮�������ͼ10����NaCl��6-BAв�ȴ����ĸ�ʱ�����,SCLRRK1�������ƶ���ScmiR390-5p�෴,������ΪScmiR390-5p������SCLRRK1��ת¼��ˮƽ��NaClв��������Ӧ,��������ScmiR390-5p/SCLRRK1����ģ����NaCl��6-BAв����Ӧ��������,������ӦABA��GA3��IAA��PEGв��ʱ,ScmiR390-5p ��SCLRRK1���ֱܷ��ܵ����Ե�ת¼ˮƽ���ء�ͼ10
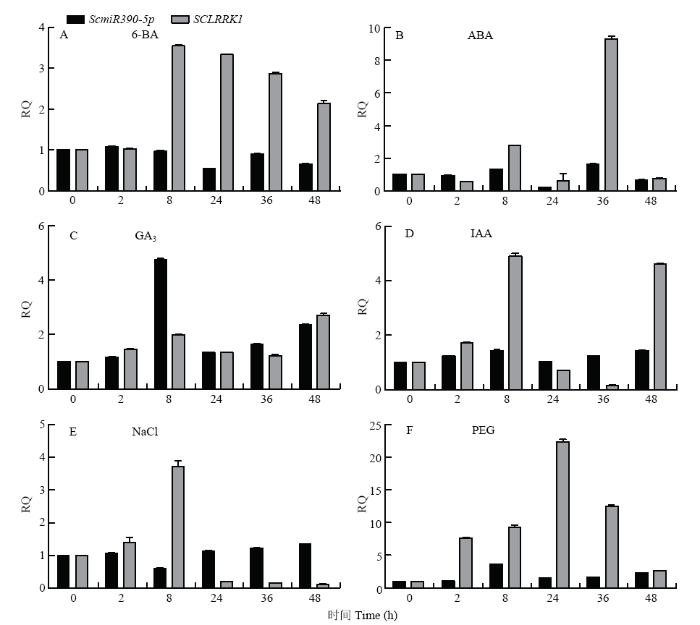 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ10ScmiR390-5p��SCLRRK1�ڸ��ַ�����в���µ�ʵʱӫ�ⶨ��PCR����
Fig. 10Expression of the SCLRRK1 and ScmiR390-5p by real-time PCR in different abiotic stress
3 ����
�����о�������ֲ������澳в�ȶ�������Ӧ�Ե���������miRNA����ͨ�����ذл���ı���������ֲ���澳�ĵ���[25,26]�����澳в����,ֲ�������γɷ�����в�����ء�miRNA/�л����������miRNA�ȶ�����ط�����·����[27,28]��miR390/tasiRNA/ ARF��Ҷ�����ٵļ��Է���[29],�ر��ֲ��������ٷ���������ʱ�ڵ�ת��[30]�ȷ�������Ҫ��������,miR390/TAS3/ARF2/ARF3/ARF4�������Ʋ������������ģʽ�Ѿ��õ�֤ʵ[31]��miR390��Ӧ����[8,9,10]�����ַ�����в��[15,16,17,18,19,20,21,22]Ҳ�в����о�,��miR390����Ӧ������в�ȹ����еľ�����������������������ϢԤ���,�����о������뼰�������Ϸ�չ,�ھ�miRNAs��DZ�ڰл�����֤�书�ܶ��ڲ���miRNAs���ػ��������ش����о�ͨ����ScmiR390-5p�İл������������ϢѧԤ����һ�����Ұ��������������ظ��������嵰��ø�е���SCLRRK1,��ͨ��RLM- 5��RACE��֤��ScmiR390-5p������SCLRRK1�İе�,����ϵͳ��������,SCLRRK1��Ҫ�������ڼ�ø��cytoplasmic protein kinase domain��������ṹ��extracellular domain���Ϳ�Ĥ�ṹ��transmembran domain��[32]�ȶ����ź�ʶ��;���еĹؼ����,������Ϊ��Ĥ�����ܹ��������̼�,ͨ�����ữ���ò�������źŴ��ݡ�LRR�������嵰��ø��ֲ������֪������һ���Ĥ�����嵰��ø,����ֲ���������������ź�ת���Լ�в�ȷ�Ӧ�о�����Ҫ�ĵ��ع��ܡ���LRR�������嵰��ø��N-ĩ�����ź��ģ�signal peptide����������������leucine zipper motif��,������������LRR����,ÿ�������б�������lxxlxxlxxlx-lxxnxlvgxip,��ø��Cĩ��[33,34,35,36,37,38]��LEE��[23]����ˮ��LRR�����嵰��øOs-RLK1�ܹ��������յ�����,��յ�[39]���õ����ij��LRK����AcLRR���뵽��ƽ�ĸ��,�������ڵ��µ�в���µĴ�������Ը��ڶ���,��������нϺõ��͵��µ����������о�Ҳ����SCLRRK1������������в�������±����ܵ�����,֤ʵLRR�������嵰��ø�������в����Ӧ���̡��о�ͬʱ���ֵ����յ�ScmiR390-5p����,SCLRRK1������֮�����,��ScmiR390-5p/SCLRRK1ģ��Ե���������Ӧ������,����5����ѱв�ȴ���10 d���������ڽ���˪������,SCLRRK1�ı���������,��û������ѱ���IJ��ϱ��ֳ��෴�ı���ģʽ,�Ʋ��������в����ScmiR390-5p/SCLRRK1ģ�����ŵ�������ʱ����ӳ����ܴ��ڷ������ڡ���һ������ScmiR390-5p/SCLRRK1ģ�����֯������,�������ScmiR390-5p��Ҷ�еı��������,����֮,�ڿ龥�����뾥�еı������ϵ�;��SCLRRK1�ı��������ScmiR390-5p�෴���ý����ʾScmiR390- 5p/SCLRRK1ģ��Ҳ���ܲ������������ٷ���,�����д���һ�������о�֤ʵ��
�����������ظ��������嵰��ø������������в�ȷ�Ӧ��Ҳ��������Ҫ������,��������е�LRRs�����ܹ������������ź�,���˼��ص�С���ӻ���������,�����ܽ���һЩС���Ӷ����༤�ص��źš�HONG��[40]�����Ͻ�ֲ���¡�õ�RPKI,�ڸɺ����NaClв������������������ߡ����о�������cmiR390-5p������SCLRRK1,��ת¼��ˮƽ��NaCl��6-BAв��������Ӧ,��������ScmiR390-5p/ SCLRRK1����ģ����NaCl��6-BAв����Ӧ��������,������ӦABA��GA3��IAA��PEGв��ʱ,ScmiR390-5p��SCLRRK1���ֱܷ��ܵ����Ե�ת¼ˮƽ���ء�
4 ����
RT-qPCR�Ͱ붨��PCR����,�ڵ���в����,ScmiR390-5p������SCLRRK1����,RLM-5��RACE֤ʵScmiR390-5p����SCLRRK1ת¼���и�λ����ATTCCT//CCTGAGTT����¡���SCLRRK1��CDS���к�DNA����,������Ϣѧ��������SCLRRK1��һ����������ظ������൰��ø,���ڵ��͵Ŀ�Ĥ��ˮ�Ե���,���ڶ������LRR�ṹ��1�����ο�Ĥ���Լ�1�����ڼ�ø�ṹ��ScmiR390-5p/SCLRRK1����ģ������������֯�����о߱���������;ScmiR390-5p��ת¼��ˮƽͨ������������SCLRRK1�ı���Ե���в��������Ӧ��ͬʱ,������ScmiR390-5p/SCLRRK1����ģ����NaCl��6-BAв����Ӧ�������á�ScmiR390-5p/SCLRRK1����ģ�鲻��ABA��GA3��IAA��PEGв��������Ӧ,ScmiR390-5p��SCLRRK1�ֱ���ת¼ˮƽ�ܵ������źŵ��ء�
�ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.3724/SP.J.1005.2013.00315Magsci [��������: 1]

����miRNA��ֲ������������е���Ҫ����, �����Ѿ���չ��ֲ��miRNA��Ԥ�⡢���������ܺͽ����ȷ�����о�����������ģʽֲ��������������, miRNA�Ļ�����ѧ�ͽ�����Ϣ������ΪmiRNA����Դ�ͽ����о��ṩ��Խ��Խ���֤�ݺͼ�˵, Ȼ����δ������ֲ��miRNA���������ϵͳ���������´�miRNA����Դ�Լ���Ӧ�ļ��ּ�˵��miRNA�IJ�����������miRNA�Ĺ��ܽ����ȼ�����������������ֲ��miRNA�������о���չ��
DOI:10.3724/SP.J.1005.2013.00315Magsci [��������: 1]
����miRNA��ֲ������������е���Ҫ����, �����Ѿ���չ��ֲ��miRNA��Ԥ�⡢���������ܺͽ����ȷ�����о�����������ģʽֲ��������������, miRNA�Ļ�����ѧ�ͽ�����Ϣ������ΪmiRNA����Դ�ͽ����о��ṩ��Խ��Խ���֤�ݺͼ�˵, Ȼ����δ������ֲ��miRNA���������ϵͳ���������´�miRNA����Դ�Լ���Ӧ�ļ��ּ�˵��miRNA�IJ�����������miRNA�Ĺ��ܽ����ȼ�����������������ֲ��miRNA�������о���չ��
[��������: 1]
DOI:10.1016/j.bbagrm.2011.05.001URL [��������: 1]
[��������: 1]
[��������: 1]
[D].
[��������: 2]
[D].
[��������: 2]
[D].
[��������: 2]
[D].
[��������: 2]
[��������: 2]
[��������: 2]
DOI:10.1186/s12864-016-3372-0URL [��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[��������: 1]
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 2]
[D].
[��������: 2]
[D].
[��������: 2]
[D].
[��������: 2]
[��������: 2]
.
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
DOI:10.1016/j.plantsci.2003.08.008URL [��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1093/jxb/ert164URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1126/science.3291115URL [��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1016/0092-8674(90)90801-KURL [��������: 1]
DOI:10.1104/pp.108.2.451URL [��������: 1]
[��������: 1]
DOI:10.1016/j.plantsci.2008.09.004URL [��������: 1]
[��������: 1]
Magsci [��������: 1]
<p>���ij��(��Atriplex canescens��)cDNA�Ŀ��еõ�һ�������ḻ���ظ����������嵰��ø(Leucine��rich repeat receptor��like protein<br /> kinases, LRR��RLKs)��ȫ��cDNA����, ����Ϊ��AcLRR��(��¼�ţ� JN974247). Ϊ�о���AcLRR���Ŀ��湦��, ���乹������ĸ��������pYES��DEST52, ��ת������ƽ�ĸINVSc1������, ��������ĸINVSc1(pYES����AcLRR��), ��������Ρ� � �ɺ��� ���¡� �䶳�ͻ�������в��, �����ڲ�ͬ�澳в���µı�������. ��ʵ����������, INVSc1(pYES����AcLRR��)�ڸ���в���µĴ�������Ը��ڶ���, ������AcLRR��������п�NaCl,KCl,NaHCO<sub>3</sub>,Na<sub>2</sub>CO<sub>3</sub>�� �ɺ��� ���¡� �䶳�ͻ�������в�ȵ�����, �ر���䶳�ͻ��������ֳ����Կ���. �ɴ˿��Ʋ�û���������ij���Ŀ�����ع���.</p>
Magsci [��������: 1]
<p>���ij��(��Atriplex canescens��)cDNA�Ŀ��еõ�һ�������ḻ���ظ����������嵰��ø(Leucine��rich repeat receptor��like protein<br /> kinases, LRR��RLKs)��ȫ��cDNA����, ����Ϊ��AcLRR��(��¼�ţ� JN974247). Ϊ�о���AcLRR���Ŀ��湦��, ���乹������ĸ��������pYES��DEST52, ��ת������ƽ�ĸINVSc1������, ��������ĸINVSc1(pYES����AcLRR��), ��������Ρ� � �ɺ��� ���¡� �䶳�ͻ�������в��, �����ڲ�ͬ�澳в���µı�������. ��ʵ����������, INVSc1(pYES����AcLRR��)�ڸ���в���µĴ�������Ը��ڶ���, ������AcLRR��������п�NaCl,KCl,NaHCO<sub>3</sub>,Na<sub>2</sub>CO<sub>3</sub>�� �ɺ��� ���¡� �䶳�ͻ�������в�ȵ�����, �ر���䶳�ͻ��������ֳ����Կ���. �ɴ˿��Ʋ�û���������ij���Ŀ�����ع���.</p>
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}