 ,�й��ȴ�ũҵ��ѧԺ���������о���,�������� 571533
,�й��ȴ�ũҵ��ѧԺ���������о���,�������� 571533Effects of Salt Stress on Seedlings Growth, Photosynthesis and Chlorophyll Fluorescence of Two Species of Artocarpus
SU LanXi, BAI TingYu, YU Huan, WU Gang, TAN LeHe,Spice and Beverage Research Institute, Chinese Academy of Tropical Agricultural Sciences, Wanning 571533, HainanͨѶ����:
���α༭: ������
�ո�����:2019-01-11��������:2019-03-18�����������:2019-06-16
| ��������: |
Received:2019-01-11Accepted:2019-03-18Online:2019-06-16
����� About authors
������,Tel��0898-62553687;E-mail�� sulanxi1988@163.com��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (1188KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
������, ��ͤ��, �㻶, ���, ̷�ֺ�. ��в�ȶ�2�ֲ�������ֲ���������������ӫ�����Ե�Ӱ��[J]. �й�ũҵ��ѧ, 2019, 52(12): 2140-2150 doi:10.3864/j.issn.0578-1752.2019.12.011
SU LanXi, BAI TingYu, YU Huan, WU Gang, TAN LeHe.
0 ����
���о����塿�μ���Լռ½���������25%,�Ƿֲ�������������֮һ���ҹ������μ������ϴ�,�����˿���������ؼ��ٵ�ì������ͻ��,����������Դ,�ر����ҹ�������������Ϊһ����Ҫ�����غ���Դ,ؽ������������[1]��ɸѡ���ξ���ֲ��,��չ����ũҵ,�������μ�ص���Ч;��[2,3]��Ȼ�������μ����������������,���������ֲ�������൱����,�����������ҹ������μ�صĿ������á�����,��ɳ�����������������ڵز���ܴ�,����ȫ��ɺ���绯��ϸɰ,����ƶ�,��ˮ��Դ��ȱ,פ���ٱ�����ʳ������ˮ���������ϵ������ϵ�����������������������μ�����,�Կ������ñ���������Դ������Ҫ���塣��ǰ���о���չ�������ۺ����������ɣ�ƣ�Moraceae������������Artocarpus����ɫľ����ʳ��������۹�ʵ�к��зḻ���Ƿ֡������ʺ�֬��,Ӫ����ֵ��,���Ӹ������ۡ���������⼰���Ӹ��������ʡ�̼ˮ�����P�ḻ����ʳ��ά,������ʳ,�ɴ���ʳ�����߾��ǿɿ������õ�ľ����ʳ������Դ��Ŀǰ,���������ֲ�����в�ȵ���Ӧ��չ�˴������о�,����в�ȿ����ƴ�«έ����������SPADֵ�����ӫ�����,��������Ʒ��Ӱ��̶ȴ�������Ʒ��[4,5],����Ũ����в�ȶ�ֲ���Ӱ�첻����,��Ũ����в�ȶ�ֲ���������������[6]��һ����Ϊ��в�ȶ�ֲ����˺���Ҫ��������в�ȡ����Ӷ�����Ӫ��ʧ��3�����档��в�Ȼ����Ƹ�ϵ��ˮ������,������������ú���������Ч��;ֲ������Na+�� Cl-�Ĵ������۴�����ϸ���ڵ�����ƽ��,�ƻ�ϸ��Ĥ�ṹ����ʹ��л����;��������Ӫ��Ԫ�����������ֲ������Ӫ����ȱ,�Ӷ�Ӱ��ֲ������������ͷ���[7]�������о�����㡿��ȥ������ֲ����о��༯������ֲ������������Լ������ȷ��ȷ���[8,9,10,11,12,13,14],���ȴ�ľ����ʳ����Ŀ�������δ������,��в���²����ۺ������ҶƬ���ӫ�����ԵIJ���Ҳ����ȷ���������Ĺؼ����⡿���о��Բ����ۺ����������Ϊ�������,��ʾ��в���������ü����ָ��֮��Ĺ�ϵ,Ϊ�ȴ��μ�ؿ�������,��չ������ɫũҵ���ȴ�ľ����ʳ����ṩ�ο���1 �����뷽��
1.1 ���Բ���
1.1.1 ���Ժ�ɳ ���Ժ�ɳ���Ժ���ʡ��������¡��ʯ÷�壨18��39��43���N;110��15��47���E��,�õ������ȴ������Լ�������,�����ˮ2 135 mm,ĸ��Ϊ����dz��������[15],��ɳpH 6.85,��Ч��30 mg?kg-1,��Ч��12.68 mg?kg-1,��8.01 mg?kg-1,�л���0.5 g?kg-1��1.1.2 ���Բ��� ���Ժ�ˮΪʯ÷�庣ˮ,�õ絼�ζȼƲⶨ��ˮ�ζ�Ϊ34.0�롣���Է���Ϊţ���л��ʣ�N,15.6 g?kg-1;P2O5,47.2 g?kg-1;K2O,12.9 g?kg-1��,���Խ��պ�ϲ����Ƽ�����˾�����Բ�������Ϊ��������1�ż���,�������Ϊ�й��ȴ�ũҵ��ѧԺ���������о�������ѡ����4��Ʒ�֡�
1.2 �������
����������2018��3��6�����й��ȴ�ũҵ��ѧԺ���������о������ҽ��С�����������ɸ��2 mm����װ�������裨ֱ��18 cm,��30 cm����,ÿ��װ��6 kg,����1%�ı������Ȱ���ţ���л���60 g,����1����Ҷһ�ĵĹ��Բ�������/�������,ÿ�콽��ͬ��Ũ�ȵĺ�ˮ200 mL����Ũ�ȹ���5��ˮƽ��T0�������Ӻ�ˮ������ˮ,����ˮ�ζ�Ϊ0.05��,�˴����Բ��ƣ���T1����Ũ��Ϊ3.4 g��kg-1,900 gˮ��100 g��ˮ���,����Ũ������ͬ������T2����Ũ��Ϊ10.2 g��kg-1����T3����Ũ��Ϊ17.0 g��kg-1����T4����Ũ��Ϊ23.8 g��kg-1����Ϊ��ֹ�����ۺ�����������μ���Ӧ,���������ÿd����3.4 g��kg-1�����ķ�ʽ�ﵽԤ��Ũ�ȡ������������,ͨ�����ط�������ˮ��ά��������ˮ����75%����,ÿ�����18��00,����㷨�ɼ��������к�ˮ���ⶨ,��ʱ��ˮ��15 d��������ˮ��ϴ3��5��,����֮ǰ�ķ�ʽ�ټ��벻ͬ��Ũ�ȵĺ�ˮ,�Ա��������η�Ũ�ȵ�����ȶ���ÿ��ˮƽ����3���ظ�,ÿ���ظ�4��,�ܼ�60�ꡣ������������30 d��ⶨ���ָ�ꡣ1.3 ��Ŀ�ⶨ�뷽��
1.3.1 ����������ָ��ⶨ ��ֲ4���º�ʼ�ⶨ����ָ��,ÿ���������ѡȡ6�곤�ƽ�һ�µIJ���������������ⶨ����ָ��,���Ϊ��������ҶƬ��ߵ�ĸ߶�;����Ҷ��Ҷ������ó���ϵ�����ⶨ��Ҷ���=��������0.75,δչ��Ҷϵ��0.5��[16];�������α꿨�߲ⶨ,ͳһ�ⶨ�ྥ����2 cm����Ҷ���غ�����SPAD-502��ЯʽҶ�����ٲ��Dzⶨ��ֲ����Ʒ�ջ��,��ȥ����ˮ��ֲ���ϴ3��,�ֳɵ��ϲ��͵��²�,����105�������ɱ��30 min,ת��65���������,����ȡ������������ڱ�=������/���ϲ����ء�1.3.2 ���ָ��ⶨ ����LI-6400��ϲⶨ��,ѡ����������������һ�µĵ���Ҷ�ⶨҶƬ��������ʣ�Pn���������ȣ�Gs�����������ʣ�Tr���Ͱ���CO2Ũ�ȣ�Ci���ȹ��ָ�ꡣ��������ֵ��Ls����ʽ��Ls=1-Ci/Ca��ʽ�У�CiΪ����CO2Ũ��;CaΪ����CO2Ũ�ȣ�[17]���ⶨʱ��Ϊ�������ʵ�����9��00��11��00,CO2Ũ��Ϊ��385��5����mol?mol-1,�ⶨ��ǿΪ1 500 ��mol?m-2��s-1,ҶƬ�¶�Ϊ��25.5��2����,�������ʪ��ά����45%���ҡ����ñ�Ҷ��,����ʽ��·,���پ��趨Ϊ500 ��mol��s-1��ÿ���������ѡȡ6�곤�ƽ�һ�µ�����,ѡȡ��ͬ��λ��ҶƬ,ÿҶ�ظ��ⶨ3�Ρ�
1.3.3 Ҷ����ӫ������IJⶨ ���ó���Яʽ����ӫ����Mini-Image-PAM�ⶨ,ÿ���������ѡȡ6��ҶƬ����������,ѡȡ����Ҷ�ⶨ,ÿҶ�ظ��ⶨ3�Ρ�ҶƬ��������30 min��,�ⶨ��ʼӫ�⣨Fo�������ӫ�⣨Fm���Լ���������Ч�ʣ�Fv/Fm��;��Ȼ���»30 min��,�ⶨ����Ӧ�µ���Сӫ��Fo�䡢���ӫ��Fm�䡢��̬ӫ��Fs��
1.3.4 �����������ʲⶨ ������Ʒ���ա�����ũ��������[18]�����������ʲⶨ��
1.4 ���ݷ���
��SPSS 19.0��,ʹ��Kolmogorov-Smirnov�����Levene's������������ݽ�����̬�����ͷ������Լ��顣���õ����ط��������ANOVA���������ݱȽ�,����Duncan�¸�������鴦��������������ˮƽ��P<0.05����ʹ��R������3.3.2�����corrplot�������������Է���,BioEnv������ѡ�Բ����ۺ�������������仯��������Ӱ��Ĺ�ϲ���,��ͨ��vegan��������з���ֽ������variance partitioning analysis,VPA�����о���в�����ϲ����Լ����ǹ�ͬ�Բ����ۺ�������������仯�Ĺ��ס�2 ���
2.1 ��в�ȶԲ����ۺ����������������Ӱ��
����1���Կ���,��������,T1�������������˲������������ߡ����ֺ�Ҷ�������,�����ʷֱ��23.75%,18.23%��229.31%,T1��������Ũ�ȴ������첻������T2�����������Ʋ�����ҶƬ��Ҷ���غϳ�,�����ʴ�45.63%,T2��T3��T4�����������Բ��졣T1�����Բ����������������Ļ�������������,�Ե��ϲ����ء����²����غ��ܸ��ص������ʷֱ��32.09%,25.66%��30.02%,T4������������������͡���������,T2��T4�����������Ӳ���������ĸ��ڱ�,���ӷ��ȴ�44.68%,��T3��T4�����������Բ��졣��в������������ϲ������������Բ�ͬ�ڲ�����,��T4������Ҷ�����,������в�ȳ̶ȵ�����,��������ϲ�����ָ�����½�����,��������ղ��첻��������в�ȶ������ҶƬҶ���غ���������Ӱ�졣��������,T2��������������������ϲ����ء����²����غ��ܸ���,���ٱ����ֱ��37.57%,27.54%��35.03%,T3��������������͡�T1�������������������ٵ��²�����,���ٱ���Ϊ15.61%,�Ե��ϲ����غ��ܸ��ص�Ӱ�첻��������������,T4������������������ڱ�,������36.36%,������Ũ�ȴ���������������Բ��졣
Table 1
��1
��1��в���²����ۺ��������������ָ��仯
Table 1
| ���� Treatment | ��� Plant height (cm) | ���� Stem diameter (mm) | Ҷ��� Leaf area (cm2) | Ҷ���غ��� Chlorophyll content SPAD | ���ϲ����� Dry weight of shoot (g) | ���²����� Dry weight of root (g) | �ܸ��� Total dry weight (g) | ���ڱ� Ratio of root/ shoot (%) | |
|---|---|---|---|---|---|---|---|---|---|
| ������ Jackfruit | T0 | 40.17��4.59a | 5.76��0.44a | 160.05��44.11a | 52.97��1.15a | 8.82��0.54a | 4.17��0.34a | 12.99��0.88a | 0.47��0.01b |
| T1 | 30.63��3.37b | 4.71��0.39b | 113.14��6.37b | 53.33��4.13a | 5.99��0.27b | 3.10��0.14b | 9.09��0.39b | 0.52��0.02b | |
| T2 | 24.37��4.52b | 4.32��0.12b | 117.43��5.34b | 28.80��5.73bc | 5.06��0.15cd | 3.45��0.26b | 8.52��0.29bc | 0.68��0.06a | |
| T3 | 28.63��2.63b | 4.56��0.11b | 111.07��5.87b | 32.93��5.60b | 5.34��0.36bc | 3.33��0.14b | 8.67��0.25bc | 0.63��0.07a | |
| T4 | 31.80��3.98b | 4.31��0.30b | 77.84��17.91b | 22.67��2.67c | 4.62��0.41d | 3.12��0.14b | 7.74��0.53c | 0.68��0.04a | |
| ����� Breadfruit | T0 | 77.13��8.87a | 8.41��1.10a | 266.47��31.90a | 44.27��3.15a | 17.14��0.80a | 5.70��0.36a | 22.84��1.10a | 0.33��0.01bc |
| T1 | 80.23��4.57a | 8.62��0.75a | 293.23��44.24a | 45.73��5.61a | 16.91��1.74a | 4.81��0.47b | 21.72��2.17a | 0.28��0.01c | |
| T2 | 72.10��3.55a | 7.81��0.11a | 278.34��42.92a | 40.53��2.40a | 10.70��0.61b | 4.13��0.08c | 14.84��0.53b | 0.39��0.03b | |
| T3 | 79.17��2.70a | 7.66��0.79a | 240.91��49.80a | 47.37��1.95a | 6.89��0.98c | 2.12��0.28d | 9.00��1.25d | 0.31��0.02c | |
| T4 | 70.00��11.10a | 7.40��1.85a | 307.84��52.05a | 46.67��4.95a | 8.16��0.67c | 3.63��0.39c | 11.79��0.78c | 0.45��0.06a |
�´��ڴ�|����CSV
2.2 ��в�ȶԲ����ۺ���������������Ե�Ӱ��
2.2.1 �Ծ�������ʵ�Ӱ�� ������в�ȳ̶ȵ�����,������ҶƬ�ľ�������ʼ����½�����������,T2�����Ծ�������ʵ������ʴ�84.41%,T2��T3��T4�����������Բ��졣T1�����϶��ս��;�������ʵı���Ϊ9.56%��P<0.05���������ҶƬ�ľ������������в�ȳ̶����ӳ����½�������,T1��T4�����½��ı����ֱ�Ϊ21.79%��39.01%��61.02%��69.19%,����T1��T2��Ũ�ȴ������������Բ���,T3��T4��������첻������ͼ1-a����ͼ1
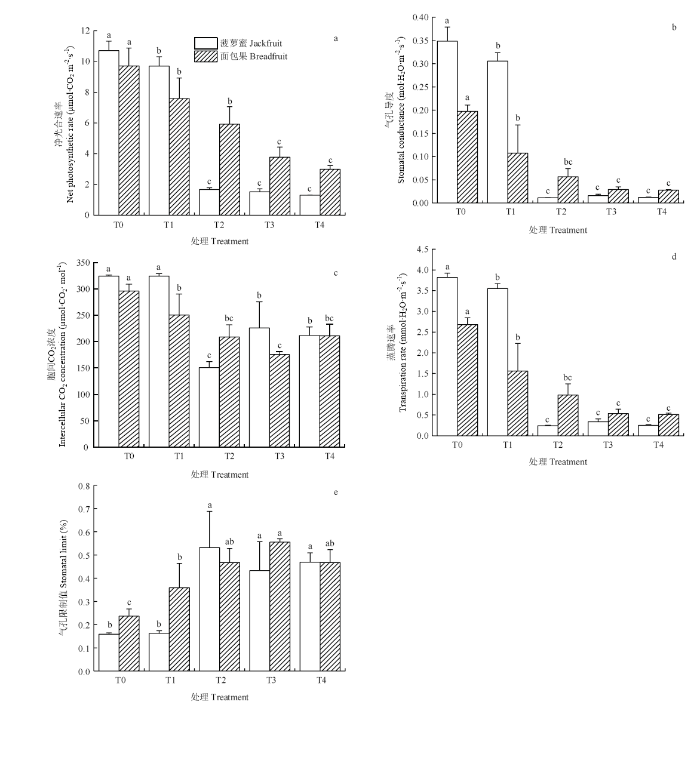 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1��в���²����ۺ����������ҶƬ�Ĺ�ϲ���
Fig. 1Photosynthetic parameters in leaves of jackfruit and breadfruit seedlings under salt stress (mean��SE)
2.2.2 ��ҶƬ�����ȵ�Ӱ�� ������в�ȳ̶ȵ�����,������ҶƬ�����ȱ仯�����뾻��������������ơ�T2�����������ȵ�����������ǿ,�϶��������ʴ�96.80%,T2��T3��T4�����������Բ��졣T1�����϶��ս��������ȵı���Ϊ12.29%��P<0.05���������ҶƬ������������в�ȳ̶����ӳ����½�������,T1�����������Ƚ϶����½��ı���Ϊ45.43%��P<0.05��,T1������������T3��T4������ͼ1-b����
2.2.3 ����CO2Ũ�ȵ�Ӱ�� �����ۺ������ҶƬ����CO2Ũ������в�ȳ̶ȵ����Ӿ������½�������������,���в���������T2����ʱ�����ֵ,���������T3����ʱ�����ֵ��T1�����IJ�����ҶƬ����CO2Ũ��������������Բ���,�����T1����ҶƬ����CO2Ũ�Ƚ϶����½��ı���Ϊ15.38%��P<0.05,ͼ1-c����
2.2.4 ��ҶƬ�������ʵ�Ӱ�� ������в�ȳ̶ȵ�����,������ҶƬ�������ʱ仯�����뾻��������������ơ���������,T2�������������ʵ������ʴ�93.58%,T2��T3��T4�����������Բ��졣T1�����϶��ս����������ʵı���Ϊ6.97%��P<0.05���������ҶƬ��������������в�ȳ̶����ӳ����½�������,T1�������������ʽ϶����½��ı���Ϊ41.73%��P<0.05��,��������T3��T4������ͼ1-d����
2.2.5 ��ҶƬ��������ֵ��Ӱ�� T2��T4�����IJ�����ҶƬ��������ֵ�������ڶ��պ�T1������������ı仯���Ƚϲ�����С,������в�ȳ̶ȵ�����,����������������ֵ�������Ӻ͵�����,���������ڶ���,T3��������������ֵ��������T1������ͼ1-e����
2.3 ��в�ȶԲ����ۺ����������Ҷ����ӫ�����Ե�Ӱ��
��2�������,������ҶƬ�⻯ѧ���ϵ����qP����DZ��������ת��Ч�ʣ�Fv/Fm����ʵ�ʹ���ת��Ч�ʣ���PSII������Ե��Ӵ���Ч�ʣ�rETR����T0��T3����֮���������Բ���,��T4�������������ڶ���,���ٵı����ֱ�Ϊ7.37%��12.66%��23.61%��23.40%����������,T4������ҶƬ��ϵͳ����ԣ�Fv/Fo���������ʴ�36.63%����в�ȶ�ҶƬ�ķǹ⻯ѧ����ϵ����NPQ���������������á���Ũ����в�ȶ������ҶƬ��Ҷ����ӫ������������Ӱ�졣T4���������ҶƬ�Ĺ⻯ѧ���ϵ�����ǹ⻯ѧ����ϵ����ʵ�ʹ���ת��Ч�ʺ���Ե��Ӵ���Ч�ʽ϶�����������,���ٵı����ֱ�Ϊ4.26%��55.56%��5.63%��5.90%��Table 2
��2
��2��в���²����ۺ����������ҶƬ��Ҷ����ӫ������
Table 2
| ���� Treatment | �⻯ѧ���ϵ�� Photochemical quenching coefficient qP | �ǹ⻯ѧ���ϵ�� Non-photochemical quenching coefficient NPQ | ��ϵͳ����� PSII activity Fv/Fo | DZ��������ת��Ч�� Potential maximum light energy conversion efficiency Fv/Fm | ʵ�ʹ���ת��Ч�� Actual light energy conversion efficiency ��PS�� | ��Ե��Ӵ���Ч�� Relative electron transfer efficiency rETR | |
|---|---|---|---|---|---|---|---|
| ������ Jackfruit | T0 | 0.95��0.02a | 0.11��0.02a | 3.74��0.03ab | 0.79��0.00a | 0.72��0.01a | 8.29��0.10a |
| T1 | 0.94��0.00a | 0.10��0.06a | 4.07��0.06a | 0.80��0.00a | 0.72��0.00a | 8.31��0.06a | |
| T2 | 0.95��0.02a | 0.18��0.05a | 3.80��0.13ab | 0.79��0.01a | 0.70��0.02a | 8.04��0.14a | |
| T3 | 0.91��0.04ab | 0.36��0.41a | 3.11��0.44bc | 0.75��0.03ab | 0.63��0.10ab | 7.19��1.18ab | |
| T4 | 0.88��0.02b | 0.44��0.32a | 2.37��1.00c | 0.69��0.09b | 0.55��0.09b | 6.35��1.03b | |
| ����� Breadfruit | T0 | 0.94��0.02a | 0.09��0.01ab | 3.78��0.20a | 0.79��0.01a | 0.71��0.02a | 5.93��0.14a |
| T1 | 0.93��0.01ab | 0.06��0.03ab | 3.93��0.17a | 0.80��0.01a | 0.70��0.01ab | 5.84��0.11ab | |
| T2 | 0.93��0.00ab | 0.11��0.00a | 3.61��0.01a | 0.78��0.00a | 0.70��0.00a | 5.86��0.01a | |
| T3 | 0.93��0.04ab | 0.10��0.05a | 3.84��0.13a | 0.79��0.01a | 0.70��0.02ab | 5.84��0.20ab | |
| T4 | 0.90��0.01b | 0.04��0.02b | 3.80��0.30a | 0.79��0.01a | 0.67��0.02b | 5.58��0.14b |
�´��ڴ�|����CSV
2.4 ��в���²����ۺ�������������������ϲ�������Է���
ͼ2����,�����ۺ�����������ܸ�����Ҷ���غ�������������ʡ������Ⱥ��������ʳ���������ع�ϵ,���ڱ���Ҷ���غ�������������ʡ������Ⱥ��������ʳ���������ع�ϵ;Ҷ���غ�������������ʡ������ȡ�����CO2Ũ�Ⱥ�������������֮�����������ع�ϵ��ͼ2
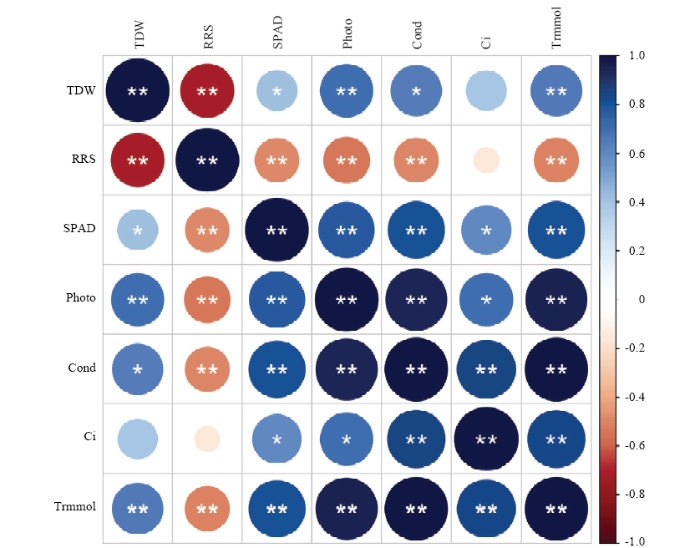 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2��в���²����ۺ�������������������ϲ�������Է���
TDW���ܸ���;RRS�����ڱ�;SPAD��Ҷ���غ���;Photo�����������;Cond��������;Ci������CO2Ũ��;Trmmol���������ʡ���*����ʾ��ͬ������IJ��������Լ����0.05ˮƽ;��**����ʾ��ͬ������IJ��������Լ����0.01ˮƽ����ɫ��ʾ�����,��ɫ��ʾ�����,Բ�εĴ�С˵������ԵĴ�С
Fig. 2Correlation between the biomass and photosynthetic parameters of jackfruit and breadfruit seedlings under salt stress
TDW: total dry weight; RRS: ratio of root/shoot; SPAD: chlorophyll content; Photo: net photosynthetic rate; Cond: stomatal conductance; Ci: Intercellular CO2 concentration; Trmmol: Transpiration rate. ��*�� indicates significant difference at 0.05 level; ��**�� indicates significant difference at 0.01 level. Blue indicates a positive correlation, red indicates a negative correlation. The size of the circle indicates the magnitude of the correlation
2.5 ��в�ȡ�Ҷ���غ�������ϲ����Բ����ۺ�����������������仯�Ĺ���
�Բ����ۺ����������������Ϊ����,̽����в�ȡ�Ҷ���غ�����ϲ�����������������Ӱ�졣ʹ��Bioenv��ѡ����������������ؼ������õĹ�ϲ���Ϊ����������ʺ�Ҷ���غ����������ֽ������VPA���Ľ����ͼ3��ʾ,��в�ȡ���������ʺ�Ҷ���غ����Լ�����֮�佻�����ý��������������仯�ı���Ϊ35.6%�����ڵ���������˵,��в�ȡ���������ʺ�Ҷ���غ������Խ��Ͳ��������������仯�ı����ֱ�Ϊ-2.1%��P=0.706����-1.3%��P=0.5����2.5%��P=0.173��;��в���뾻������ʹ����������������仯�ı���Ϊ6.3%��P=0.002��,�����������Ҷ���غ��������������������仯�ı���Ϊ10.3%��P=0.001��,��в����Ҷ���غ��������������������仯�ı���Ϊ-0.3%��P=0.001��,3�����ӹ����������������仯�ı���Ϊ20.2%��P=0.001����ͼ3
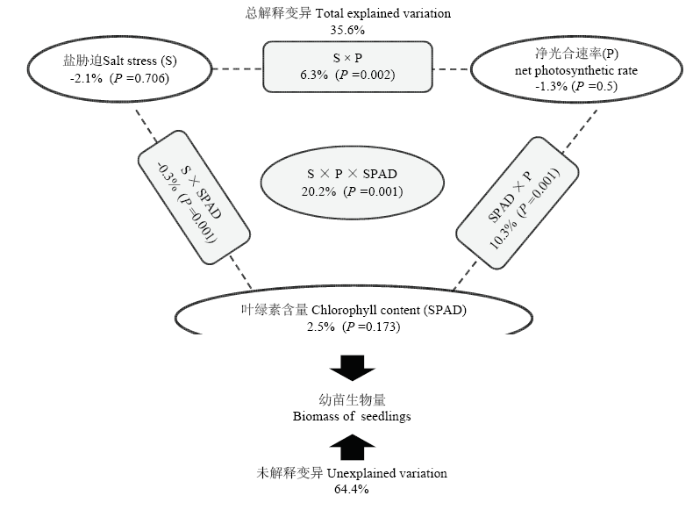 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3�����ۺ�����������������仯������������
ÿ������Ϊ�������ӵĽ�����,�������ϵ���ֵΪ���ӵĽ������ý�����,���ĵ�Ϊ���߽����Ľ�����,�ײ����ο�����ֵΪ���ܽ��͵���
Fig. 3Variation partitioning analysis (VPA) was used to quantify the effects of salt stress (S), net photosynthetic rate (P), and chlorophyll content (SPAD), and all of them on jackfruit and breadfruit seedlings biomass
Circles on the edges of the triangle show the percentage of variation explained by each factor alone. The percentage of variation explained by interactions between two or three of the factors is shown as rectangles on the sides and as a circle at the center of the triangle. The unexplained variation is depicted in the rectangle on the bottom
3 ����
�������в����Ӧ�����е�����������Ӱ������[19],���о����������в���������Ʋ����۵��ϲ�������,������������������ò�������л�����[20]������Ƽ��[19]�������о�������в�ȶ����������������������á���������ֲ����˵,�η�Ũ����û�г���������ֵʱ,ͬ�����ò��ᱻ����,�����ڽϵ�ʱ���ᱻ�ٽ�[21],��ľ���[22]������[23]�Ⱥܶ�����Ҳ���ֳ����Ƶ����ԡ������鷢��,�����۶���в������,���м��δ����IJ���������ָ�꼰���������������ڶ���,�������������ָ�������������½�����,�����첻��������������ϲ������������T1���������������������,��������Ũ�ȵ�������������,���²����ض���в�Ƚ�����,�����˵���ηֶԲ����ۺ�����������÷�ʽ��ͬ���ӱ�����Ľ������,������ϲ����������Ը�ǿ,T1��Ũ��Ϊ���������������Ũ��,��T4��Ũ�ȿ���Ϊ�����ۺ�����������������ֵ��Ҷ���غ�����ֲ��ҶƬ��������������,�Ǻ���ֲ����Ե���Ҫ����ָ��֮һ[24]����в������ǿҶ����ø����,�Ӿ�Ҷ����Ľ����������ϳ�,���Ҷ���غ����仯�ܷ�ӳ��в�ȶ�ֲ����˺��̶�[25,26,27]�����о��������,������в����ʹҶ���غ�����������,��������������ϴ�;������в�ȳ̶�����,�����۵�Ҷ���غ��������½�,�ý����ǰ���ڴ�[28]����[29]�ϵ��о�������ơ��������Ҷ���غ����仯����,����в�Ȳ����С����ڱȵ�ֵ��ӳ��ֲ�����澳�����¶��������ķ������,ֲ�ﲻͬ��λ���ηֵ����г̶Ȳ�ͬ,���¸��ڱ�ֵ�����仯[30]����1�������,��в����,�����ۺ�������ĸ��ڱȳ���������,��Ũ����в���µĸ��ڱ��������ڶ��ա��ɼ�����в�ȵ��¸�ϵ��������������ҶƬͬ������ķ������Ըı�[31]�����ڱ�Խ��,������ϵ���䵽��ͬ���������[32],�����ڸ�ϵ�Ŀ������š�������������,������Ũ������,���ڱȳ���������,�Ҳ����۵ĸ��ڱ�ֵ���������,������в����,�����۸�ϵ���ŵ��ٶȸ���,��˾����Ĺ�ϲ���ϵ��ϲ���,Ӱ���˵��ϲ��������ٶȡ����ַ���ģʽ���������Ӹ�ϵ��ˮ�ֺ�Ӫ���Ļ�ȡ,�Ӷ���ǿֲ�������������,ϡ��ϸ���ڵ��η�[33]��
��в���������Ʋ����ۺ������ҶƬ��������ʡ������Ⱥ���������,����ǰ�˶Դ���[34]����[29]���о����һ�¡�������в����,�������ϲ����½����ȵ��ڲ�����,˵�������������������Խ�ǿ����в�ȶ�ֲ�������ʹ�л��Ӱ���Ƕ���,���й�����õı仯�����ԡ�Ӱ��ֲ�������õ�������Ҫ��Ϊ�������Ӻͷ���������[35]�������о�����,��в�ȵ���ֲ��ҶƬϸ���������������䵼��֯���𡢹�����ʽ��͡�������������,��Ϊ������������в����ֲ���������½�����Ҫԭ��[36],����Щ�о�����,��������������в�Ⱥ��������½�����Ҫԭ��,��в����RuBP�Ȼ�øЧ�ʽ��ͺ�����ñ�������Ч���½����¹�����ʽ���[37]������������½�,������CO2Ũ�Ƚ���,��������������,����������ʽ�����Ҫԭ������������,����Ϊ����������[38]����˱�������,��в���²����ۺ������ҶƬ����������½�,�����Ƚ���,������CO2��Ҷ��������͡�����Ũ����T1��T2ʱ,ҶƬ����CO2Ũ��Ci��������Gs�ļ�С������,��������ֵLs����,���������Pn����,�������������в�ȴ̼�������в��,�������ر�,Ҷ��ϸ����϶��CO2���������ʼ���,�Ӷ�����Ci���͡��ɴ˱���,����Ũ��ΪT2ʱ,�����ۺ������ҶƬPn�½�����Ҫԭ��Ϊ��������,������ѧ�ڵ�[6]�о����ѵĽ��һ�¡�����ֲ���Gsֵ������Ũ�ȵ����ӳ��ֻ����½�������,��Ciȴ����������,Ls����,������Ǹ���в����ϸ���������ö��ݼ�ǿ,��ϸ���л����˴���������,�ƻ���Ҷ����ṹ��ҶƬ�������,���Ҷ���غ������͡�Ҷ��ϸ���Ĺ�ϻ����½�,˵����ʱ������ʽ��͵���Ҫԭ���Ƿ��������ơ����,�����ۺ������������Ũ�ȵ�����,������õ����ƾ��ɳ��ڵ�����������������ת��Ϊ��������������,���������߹�������������ӵ�ת����Ũ�Ȳ�ͬ,������ΪT2��Ũ��,���������ΪT3��Ũ��,������ٴ�֤ʵ�����ۺ���������ηֵ����̶ܳȲ�ͬ��
��������½���Ȼ��Ӱ��ֲ��Թ��ܵ����ա����ݺ�ת��,���Թ⻯ѧ�����½���Ϊͻ��[39]���о�����,Ҷ�����ϻ����ڸ�Ũ����в�����ױ��ƻ�,���¹�ϵͳ��ԭ������ת��Ч�ʡ�PS��DZ�ڻ��Խ���[19,29]��������Ҳ����,�ڵ�Ũ��в����,������ҶƬ�⻯ѧ���ϵ����qP������ϵͳ����ԣ�Fv/Fo����DZ��������ת��Ч�ʣ�Fv/Fm����ʵ�ʹ���ת��Ч�ʣ���PS����Ե��Ӵ���Ч�ʣ�rETR���ڴ�����Ͷ��վ��������Բ���,�ڸ�Ũ��в����,��NPQ����������������������,����ӫ������������½�,����������ҶƬ�����˹�����[40],˵������в��Ӱ���˲�����ҶƬ��ϵͳ��ļ����ܷ��䷽ʽ,ͨ������Ⱥ�ɢ���Ĺ��༤��������Ӧ��в�Ȼ��������������Fv/Fo��Fv/Fm�仯������,����ӫ������ڸ���в���¾������½�,˵�����������в�����ܹ���Ч���ֹ�ϵͳ����Ժ�DZ��������ת��Ч��,ʹ��ϻ��������ȶ�,��ֹ��������ʹ����½�,����������������в�Ȼ�����Ӧ��һ�ֱ������ơ�
Ҷ������Ӱ�����������ʵ���Ҫ����[41]�����о������ʾҶ���غ������ϲ�������������ع�ϵ,����������[41]��������ϵ��о���������Ǻϡ�������ö�ֲ������ʻ��۾�����Ҫ����[42]������������ʾ,�����ۺ�����������ܸ�����Ҷ���غ�������ϲ�������������ع�ϵ,Ҳ֤ʵ��ǿ�Ĺ�����ú�Ҷ���غ������ܴٽ������ܸ����ʵĻ��ۡ���в�ȶԲ����ۺ����������Ҷ���غ�������������ʺ�������������Ӱ�졣VPA��������,��в�ȡ�Ҷ���غ����;�������������估���߽������ý��������������仯�ı������������ԡ��������߽������ý��������������仯�ı���������,�����ۡ������������������в�ȳ̶ȵ����Ӷ�����,��ҶƬҶ���غ����ȹ�ϲ������½���������ϵ��
4 ����
��в���������Ʋ���������������Ҷ���غ���������������,����������������ڸ�Ũ����в���½����С���в�Ƚ����˲����ۡ������ҶƬ�Ĺ�ϲ���,�Ҳ����۵��½�����������������������¹�ϲ������͵�ԭ�������,�����ۺ������������Ũ�ȵ�����,������õ����ƾ��ɳ��ڵ�����������Ϊ�������Ӷ���ת��Ϊ�Է���������Ϊ�������ӡ���Ũ����в����,�����������qP��Fv/Fo��Fv/Fm����PS���rETR�������½�,��NPQ����������,˵������в�Ȼ����Ʋ����������ϵͳ����Ժ���ת��Ч��,���ӹ�ϵͳ��Ƿ��������ĺ�ɢ����Ӧ��в�Ȼ�������������Ĺ��ϵͳ����в�ȵķ��������������ڲ����ۡ���в�ȡ�Ҷ���غ����;���������Լ�����֮�佻�����ý��������������仯�ı���Ϊ35.6%�����ӫ�������ȷ�����ٵ��жϲ����ۺ������������ηֵ������ԡ��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
URLMagsci [��������: 1]

�������Է�������11������ʵ������Ϊ���ϣ��ò�ͬŨ��(0��3��5��8 g��kg-1��10 g��kg-1)NaCl��Һ����1���Խ��ദ��������в���¸����ֵ���̬���֡������������Խ������о������������(1)���κ����ﵽ8 g��kg-1ʱ��ŷ���o�������Ҷ��ֲ�����������������ӵ�10 g��kg-1ʱ�����Ͻ�����ֲ���������������������Ҳ�����ֲ�ͬ�̶ȵ��κ�֢״��(2)��в�Ⱥ����ֵ�����������½����������ۻ����٣������Ŵ���Ũ�ȵ����Ӿ����½����ƣ������ܽۡ������Ҷ��Ľ������(3)�δ��������ֵĸ��ڱ�ֵ����������в�ȶԹ�Ҷ�ᡢ��ˮţ�������Ͻ�����������Ӱ��(p<0.05)��(4)�ۺϷ��������ֵ���������̬���֣���Ϊ�ձ����㡢��ˮţ��������Ҷ��Ͷ�����и߶������ԣ����Ͻ���������ȸ����ӥצ�����ܽ۾����и߶������ԣ���ŷ���o�������Ҷ������ж������ԡ�
URLMagsci [��������: 1]
�������Է�������11������ʵ������Ϊ���ϣ��ò�ͬŨ��(0��3��5��8 g��kg-1��10 g��kg-1)NaCl��Һ����1���Խ��ദ��������в���¸����ֵ���̬���֡������������Խ������о������������(1)���κ����ﵽ8 g��kg-1ʱ��ŷ���o�������Ҷ��ֲ�����������������ӵ�10 g��kg-1ʱ�����Ͻ�����ֲ���������������������Ҳ�����ֲ�ͬ�̶ȵ��κ�֢״��(2)��в�Ⱥ����ֵ�����������½����������ۻ����٣������Ŵ���Ũ�ȵ����Ӿ����½����ƣ������ܽۡ������Ҷ��Ľ������(3)�δ��������ֵĸ��ڱ�ֵ����������в�ȶԹ�Ҷ�ᡢ��ˮţ�������Ͻ�����������Ӱ��(p<0.05)��(4)�ۺϷ��������ֵ���������̬���֣���Ϊ�ձ����㡢��ˮţ��������Ҷ��Ͷ�����и߶������ԣ����Ͻ���������ȸ����ӥצ�����ܽ۾����и߶������ԣ���ŷ���o�������Ҷ������ж������ԡ�
URLMagsci [��������: 1]
Ϊ��ʾ�����μ����������ָ�����̬������Ӧ���Ա仯����,�Ա����μ����̬�������ɵ��ִ�����Ϊ�о�����,����ǰ����Ϊ�հ��պ�ũ����ĵ�����Ϊ���̲���,�о��˱����μ����������ָ�����̬��������Ӧ�����ԡ���Ӧ���̺���Ӧ�ȡ������ʾ:�����μ���������������л��ʺ�ȫ��ָ����ۺ���̬������ʩ��Ӧ������ǿ,��ӦѸ��,������ǿ����Ӧ�ȴ�����ȫ����Ч��ָ����ۺ���̬������ʩ����Ӧ�������Բ�ǿ����������ø����ָ���������ڶ��ۺ���̬������ʩ��һ������Ӧ�ȡ�����pHֵ��ȫ�ء���Ч�غ�������øָ���������ڶ��ۺ���̬������ʩ��������Ӧ�����������μ����̬������ʩ�ڳ���(2a��)���Գ�Ч,�õ��������ĺ��������л��ʺ�ȫ��ָ��������̬����,���������ȵ��ؽ���,����ָ�����̬�����Ի���Ҫ��һ���о���
URLMagsci [��������: 1]
Ϊ��ʾ�����μ����������ָ�����̬������Ӧ���Ա仯����,�Ա����μ����̬�������ɵ��ִ�����Ϊ�о�����,����ǰ����Ϊ�հ��պ�ũ����ĵ�����Ϊ���̲���,�о��˱����μ����������ָ�����̬��������Ӧ�����ԡ���Ӧ���̺���Ӧ�ȡ������ʾ:�����μ���������������л��ʺ�ȫ��ָ����ۺ���̬������ʩ��Ӧ������ǿ,��ӦѸ��,������ǿ����Ӧ�ȴ�����ȫ����Ч��ָ����ۺ���̬������ʩ����Ӧ�������Բ�ǿ����������ø����ָ���������ڶ��ۺ���̬������ʩ��һ������Ӧ�ȡ�����pHֵ��ȫ�ء���Ч�غ�������øָ���������ڶ��ۺ���̬������ʩ��������Ӧ�����������μ����̬������ʩ�ڳ���(2a��)���Գ�Ч,�õ��������ĺ��������л��ʺ�ȫ��ָ��������̬����,���������ȵ��ؽ���,����ָ�����̬�����Ի���Ҫ��һ���о���
DOI:10.1126/science.1168572URL [��������: 1]
URL [��������: 1]
URL [��������: 1]
URL [��������: 1]
URL [��������: 1]
DOI:10.5846/stxb201409141821URLMagsci [��������: 2]
����ˮ�෨,�о��˲�ͬŨ��(0%��0.2%��0.4%��0.6%��0.8%)NaCl������1������۴�졯��������������������Լ�ҶƬϸ���ṹ��Ӱ��,Ϊ�μ�ء�۴�졯���ѵ������ṩ�ο����������:��Ũ���η�(NaCl��0.4%)������������Ҷ���غ�����ҶƬϸ���ṹ�����彻�������Լ�Ҷ����ӫ�����Ӱ�첻������������Ũ������,���������ܵ�����,ҶƬ��Ƥ�㡢դ����֯��������֯���,������֯��դ����֯ϸ����϶���,դ����֯ϸ��Ҷ��������,�ں�����������ﰿ������������;ͬʱ,Ҷ���غ��������������(<i>Pn</i>)��������(<i>Gs</i>)������CO<sub>2</sub>Ũ��(<i>Ci</i>)����������(<i>Tr</i>)��PS�����⻯ѧЧ��(<i>Fv/Fm</i>)��PS��DZ�ڻ���(<i>Fv/Fo</i>)���⻯ѧ���Ӵ���Ч��(<i>ETR</i>)���⻯ѧ���ϵ��<i>qP</i>�Լ�դ����֯/������֯�����½�,�ǹ⻯ѧ���ϵ��<i>qN</i>������;������0.8%NaClŨ��в����,����������Ҷ���غ�����ҶƬϸ���ṹ�����彻�������Լ�Ҷ����ӫ����������������Ա仯���ɴ˱���,��۴�졯�������ڽϵͺ�����(NaCl �� 0.4%)�Ļ���������������
DOI:10.5846/stxb201409141821URLMagsci [��������: 2]
����ˮ�෨,�о��˲�ͬŨ��(0%��0.2%��0.4%��0.6%��0.8%)NaCl������1������۴�졯��������������������Լ�ҶƬϸ���ṹ��Ӱ��,Ϊ�μ�ء�۴�졯���ѵ������ṩ�ο����������:��Ũ���η�(NaCl��0.4%)������������Ҷ���غ�����ҶƬϸ���ṹ�����彻�������Լ�Ҷ����ӫ�����Ӱ�첻������������Ũ������,���������ܵ�����,ҶƬ��Ƥ�㡢դ����֯��������֯���,������֯��դ����֯ϸ����϶���,դ����֯ϸ��Ҷ��������,�ں�����������ﰿ������������;ͬʱ,Ҷ���غ��������������(<i>Pn</i>)��������(<i>Gs</i>)������CO<sub>2</sub>Ũ��(<i>Ci</i>)����������(<i>Tr</i>)��PS�����⻯ѧЧ��(<i>Fv/Fm</i>)��PS��DZ�ڻ���(<i>Fv/Fo</i>)���⻯ѧ���Ӵ���Ч��(<i>ETR</i>)���⻯ѧ���ϵ��<i>qP</i>�Լ�դ����֯/������֯�����½�,�ǹ⻯ѧ���ϵ��<i>qN</i>������;������0.8%NaClŨ��в����,����������Ҷ���غ�����ҶƬϸ���ṹ�����彻�������Լ�Ҷ����ӫ����������������Ա仯���ɴ˱���,��۴�졯�������ڽϵͺ�����(NaCl �� 0.4%)�Ļ���������������
DOI:10.1146/annurev.arplant.59.032607.092911URL [��������: 1]
[��������: 1]
[��������: 1]
URL [��������: 1]
URL [��������: 1]
URLMagsci [��������: 1]
�о����μ������������ҶƬ��������ʣ�<EM>Pn</EM>�����������ʣ�<EM>Tr</EM>���������ȣ�<EM>Gs</EM>����ˮ������Ч�ʣ�<EM>WUE</EM>���Լ��������ص��ձ仯���������������ҶƬ<EM>Pn</EM>��<EM>Tr</EM>������˫�����ߣ����й��������������������ֵ�ԭ�������ڽϸߵ�̫����������ҶƬ�¶ȵ����ߣ�ҶƬ���Ȼ�Ч��(<EM>CE</EM>)�ͱ������Ӳ���(<EM>AQY</EM>)���½���ɵġ�
URLMagsci [��������: 1]
�о����μ������������ҶƬ��������ʣ�<EM>Pn</EM>�����������ʣ�<EM>Tr</EM>���������ȣ�<EM>Gs</EM>����ˮ������Ч�ʣ�<EM>WUE</EM>���Լ��������ص��ձ仯���������������ҶƬ<EM>Pn</EM>��<EM>Tr</EM>������˫�����ߣ����й��������������������ֵ�ԭ�������ڽϸߵ�̫����������ҶƬ�¶ȵ����ߣ�ҶƬ���Ȼ�Ч��(<EM>CE</EM>)�ͱ������Ӳ���(<EM>AQY</EM>)���½���ɵġ�
DOI:10.1146/annurev.arplant.51.1.463URL [��������: 1]
URLMagsci [��������: 1]
����в���»�����ijЩֲ����ڷ���һЩ�仯��������ѧ�����ﻯѧ����в�ȷ���ѧ���ƵĽǶȶ�ֲ�����в�ȵķ�Ӧ�о������˻عˣ����ṩ��һЩĿǰ֪ʶˮƽ��������ֲ�������Եķ���������������в����ֲ����������ա����������ʡ�������ø��ֲ�D�ء�������õȷ���ı仯���ɣ�����Ҳ������ֲ�﹦�ܵ��ڵ��о����������ڴӶ�ѧ���о��ĽǶ�������в�ȵ���̬��Ҫ�ԡ�
URLMagsci [��������: 1]
����в���»�����ijЩֲ����ڷ���һЩ�仯��������ѧ�����ﻯѧ����в�ȷ���ѧ���ƵĽǶȶ�ֲ�����в�ȵķ�Ӧ�о������˻عˣ����ṩ��һЩĿǰ֪ʶˮƽ��������ֲ�������Եķ���������������в����ֲ����������ա����������ʡ�������ø��ֲ�D�ء�������õȷ���ı仯���ɣ�����Ҳ������ֲ�﹦�ܵ��ڵ��о����������ڴӶ�ѧ���о��ĽǶ�������в�ȵ���̬��Ҫ�ԡ�
URLMagsci [��������: 1]
ֲ�����в�ȵ����ܷ�Ӧ�Ǹ����ӵĹ���,�ڷ���ˮƽ����������������źŵĸ�Ӧ�ʹ���,����ת¼���ӵļ�������ο�����������Ӧ���ЧӦ����ı���.������Ӧ����,�����������۸���ά�ֺ��ؽ�����ƽ���Ĥת�˵��ס�������������ϳɺ��ܼ�ˮ�ֿ���.��Щ��������Ӧ������ʹ��Һ��������Ũ�����ߺ��������ڰ����л���.������,ͨ���Ը�������ֲ���������ͻ������о�,������������Ӧ�������ת��;����ˮͨ���������������������л;��,��¡������ػ�������ת����ˮֲ���в������α���;��һ����,�����Ͻ�ͻ���弰���ý�ĸ������ͻ���깦�ܻ���ɸѡ�õ�һЩ�����źŴ��ݵ��Ļ���,��Щ�������ڲ���ֲ����в��Ӧ��ķ��ӻ��ơ�<br>Abstract:Plant responses to salt stress via a complex mechanism,including sensing and transducing the stress signal,activating the transcription factors and the corresponding metabolizing genes.Since the whole mechanism is still unclear,this review emphasize the biochemical events during the plant adaptation to salt stress referring to an index of importance:the homeostasis in cytoplasm,the biosynthesis of osmolytes and the transport of water.Most of these biochemical events were elucidated by study of halophyte and salt-sensitive mutations,also many important genes involved were cloned and used to generate stress-tolerance phenotypes in transgenic plants.On the other hand,about the molecular mechanism in signal transduction,the research of Arabidopsis mutations and yeast functional complementation provided helpful traces but not full pathway.<br>
URLMagsci [��������: 1]
ֲ�����в�ȵ����ܷ�Ӧ�Ǹ����ӵĹ���,�ڷ���ˮƽ����������������źŵĸ�Ӧ�ʹ���,����ת¼���ӵļ�������ο�����������Ӧ���ЧӦ����ı���.������Ӧ����,�����������۸���ά�ֺ��ؽ�����ƽ���Ĥת�˵��ס�������������ϳɺ��ܼ�ˮ�ֿ���.��Щ��������Ӧ������ʹ��Һ��������Ũ�����ߺ��������ڰ����л���.������,ͨ���Ը�������ֲ���������ͻ������о�,������������Ӧ�������ת��;����ˮͨ���������������������л;��,��¡������ػ�������ת����ˮֲ���в������α���;��һ����,�����Ͻ�ͻ���弰���ý�ĸ������ͻ���깦�ܻ���ɸѡ�õ�һЩ�����źŴ��ݵ��Ļ���,��Щ�������ڲ���ֲ����в��Ӧ��ķ��ӻ��ơ�<br>Abstract:Plant responses to salt stress via a complex mechanism,including sensing and transducing the stress signal,activating the transcription factors and the corresponding metabolizing genes.Since the whole mechanism is still unclear,this review emphasize the biochemical events during the plant adaptation to salt stress referring to an index of importance:the homeostasis in cytoplasm,the biosynthesis of osmolytes and the transport of water.Most of these biochemical events were elucidated by study of halophyte and salt-sensitive mutations,also many important genes involved were cloned and used to generate stress-tolerance phenotypes in transgenic plants.On the other hand,about the molecular mechanism in signal transduction,the research of Arabidopsis mutations and yeast functional complementation provided helpful traces but not full pathway.<br>
URL [��������: 1]
URL [��������: 1]
URL [��������: 1]
.
URL [��������: 1]
[A].
[��������: 1]
[��������: 1]
[��������: 1]
URLMagsci [��������: 3]
<P><FONT face=Verdana>��Ŀ�ġ��о���в�ȶԲ�ͬ�����ͺ�������Ҷ������ü�Ҷ����ӫ�������Ӱ��,Ϊ�������������������ɸѡ�ṩ���۲ο����������������ֺ���������¬���º�21�ţ��������ͣ����º�28�ţ������ͣ�Ϊ����,����ˮ�෨,��3Ҷ�ڿ�ʼ����NaCl������NaClŨ��Ϊ0��50��100��150��200��250 mmol<I>&#</I>8226;L-1��,�о���в���º�����������������ü�Ҷ����ӫ������ı仯����������о�����,��Ũ���η֣�NaCl��50 mmol<I>&#</I>8226;L-1���� �ٽ��������º�28�ŵ�����,�����ʸ���,���Ҷ����������Ҷ����a��b�ĺ���,ʹPS��⻯ѧ����Ч�� ����PS���⻯ѧ���ϵ����qP���ͷǹ⻯ѧ���ϵ����NPQ������,ʵ�ֵ�Ҷ��������ʣ�Pn������;��Ũ���η֣�NaCl��50 mmol<I>&#</I>8226;L-1�����������������͵�����,����ҶƬ��������ʣ�Pn���������ȣ�Gs����Fv/F0��Fv/Fm�� PS��⻯ѧ����Ч�ʣ���PS���⻯ѧ���ϵ����qP�����½�,�ǹ⻯ѧ���ϵ����NPQ���Ͱ���CO2Ũ�ȣ�Ci������ߡ������ۡ���в����,����Ũ������,������������½���ԭ��������������ת�����������,��ϲ�����Ҷ����ӫ������ı仯������Ϳ��Լ���Ũ��������ء�<BR></FONT></P>
URLMagsci [��������: 3]
<P><FONT face=Verdana>��Ŀ�ġ��о���в�ȶԲ�ͬ�����ͺ�������Ҷ������ü�Ҷ����ӫ�������Ӱ��,Ϊ�������������������ɸѡ�ṩ���۲ο����������������ֺ���������¬���º�21�ţ��������ͣ����º�28�ţ������ͣ�Ϊ����,����ˮ�෨,��3Ҷ�ڿ�ʼ����NaCl������NaClŨ��Ϊ0��50��100��150��200��250 mmol<I>&#</I>8226;L-1��,�о���в���º�����������������ü�Ҷ����ӫ������ı仯����������о�����,��Ũ���η֣�NaCl��50 mmol<I>&#</I>8226;L-1���� �ٽ��������º�28�ŵ�����,�����ʸ���,���Ҷ����������Ҷ����a��b�ĺ���,ʹPS��⻯ѧ����Ч�� ����PS���⻯ѧ���ϵ����qP���ͷǹ⻯ѧ���ϵ����NPQ������,ʵ�ֵ�Ҷ��������ʣ�Pn������;��Ũ���η֣�NaCl��50 mmol<I>&#</I>8226;L-1�����������������͵�����,����ҶƬ��������ʣ�Pn���������ȣ�Gs����Fv/F0��Fv/Fm�� PS��⻯ѧ����Ч�ʣ���PS���⻯ѧ���ϵ����qP�����½�,�ǹ⻯ѧ���ϵ����NPQ���Ͱ���CO2Ũ�ȣ�Ci������ߡ������ۡ���в����,����Ũ������,������������½���ԭ��������������ת�����������,��ϲ�����Ҷ����ӫ������ı仯������Ϳ��Լ���Ũ��������ء�<BR></FONT></P>
URL [��������: 1]
URL [��������: 1]
[��������: 1]
[��������: 1]
URL [��������: 1]
URL [��������: 1]
DOI:10.1016/j.agwat.2006.01.002URL [��������: 1]
DOI:10.1023/A:1006408712416URL [��������: 1]
URLMagsci [��������: 1]
�Բ�ͬ������ֲ���Ҷʯ��ľ(Raphiolepis umbellata)��ɣ(Morus alba)���¼�(Rosa chinensis)����в���µ����ҡ��¸�����������ָ̬��,�Լ�Na+��K+��Na+/K+,���븬���ᡢҶ���غ�����������ʡ������ȡ��������ʵ�����ָ��ıȽ��о�����:�ζ�3��ֲ��������Ӱ����Ũ�ȵ����ߺ�в��ʱ����ӳ�������;Na+��K+���������Ǻ�Ҷʯ��ľ��Ҫ������������,����ɣ���¼�����Ϊ���븬����;Ҷ���غ�����������ʵ�����ָ������Ũ������������,�仯����:�¼���ɣ����Ҷʯ��ľ;3��ֲ������������ǿ��������Ϊ��Ҷʯ��ľ��ɣ���¼�.
URLMagsci [��������: 1]
�Բ�ͬ������ֲ���Ҷʯ��ľ(Raphiolepis umbellata)��ɣ(Morus alba)���¼�(Rosa chinensis)����в���µ����ҡ��¸�����������ָ̬��,�Լ�Na+��K+��Na+/K+,���븬���ᡢҶ���غ�����������ʡ������ȡ��������ʵ�����ָ��ıȽ��о�����:�ζ�3��ֲ��������Ӱ����Ũ�ȵ����ߺ�в��ʱ����ӳ�������;Na+��K+���������Ǻ�Ҷʯ��ľ��Ҫ������������,����ɣ���¼�����Ϊ���븬����;Ҷ���غ�����������ʵ�����ָ������Ũ������������,�仯����:�¼���ɣ����Ҷʯ��ľ;3��ֲ������������ǿ��������Ϊ��Ҷʯ��ľ��ɣ���¼�.
URLMagsci [��������: 1]
<p>��1��������ľΪ�Բ�,����NaClŨ�ȷֱ�Ϊ0(CK)��0.15%��0.3%��0.45%��0.6%5������,���ÿ���Ҷ����ӫ���յ�����ѧ���߷�������(JIP-test),�о���NaClв�ȶԻ���ľҶƬ������ԺͿ���Ҷ����ӫ���յ�����ѧ������Ӱ��.�������: ����NaClŨ�ȵ�����, ����ľҶƬ�е�Ҷ����a��Ҷ����b����Ҷ���غ�������,Ҷ����a/b��ֵ�����ߺ��½�,����ܲ��غ���������; ҶƬ�ľ��������(<em>P</em><sub>n</sub>)��������(<em>g</em><sub>s</sub>)����,����NaClŨ�ȣ�0.3%ʱ,ҶƬ<em>P</em><sub>n</sub>�½���Ҫ����������;��NaClŨ�ȣ�0.3%ʱ,<em> P</em><sub>n</sub>�½���Ҫ�ɷ�������������;��λ�������Ĺ���(TR<sub>o</sub>/CS<sub>o</sub>)�����Ӵ��ݵ����Ӳ���(ET<sub>o</sub>/CS<sub>o</sub>)����λ����ķ�Ӧ��������(RC/CS<sub>o</sub>)�����Ӳ���������������(<em>ψ</em><sub>o</sub>��<em>φ</em><sub>Eo</sub>)����,����λ������յĹ���(ABS/CS<sub>o</sub>)��ӫ���յ�������K��(<em>W</em><sub>k</sub>)��J��(<em>V</em><sub>j</sub>)��������,˵����в�ȶԻ���ľҶƬ����������(OEC)��������PS��Ӧ����������˺�.��NaClŨ��Ϊ0.3%ʱ,PS�����⻯ѧЧ��(<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>)�⻯ѧ����ָ��(<em>PI</em><sub>ABS</sub>)�ֱ�ȶ����½� 17.7%��36.6%.</p>
URLMagsci [��������: 1]
<p>��1��������ľΪ�Բ�,����NaClŨ�ȷֱ�Ϊ0(CK)��0.15%��0.3%��0.45%��0.6%5������,���ÿ���Ҷ����ӫ���յ�����ѧ���߷�������(JIP-test),�о���NaClв�ȶԻ���ľҶƬ������ԺͿ���Ҷ����ӫ���յ�����ѧ������Ӱ��.�������: ����NaClŨ�ȵ�����, ����ľҶƬ�е�Ҷ����a��Ҷ����b����Ҷ���غ�������,Ҷ����a/b��ֵ�����ߺ��½�,����ܲ��غ���������; ҶƬ�ľ��������(<em>P</em><sub>n</sub>)��������(<em>g</em><sub>s</sub>)����,����NaClŨ�ȣ�0.3%ʱ,ҶƬ<em>P</em><sub>n</sub>�½���Ҫ����������;��NaClŨ�ȣ�0.3%ʱ,<em> P</em><sub>n</sub>�½���Ҫ�ɷ�������������;��λ�������Ĺ���(TR<sub>o</sub>/CS<sub>o</sub>)�����Ӵ��ݵ����Ӳ���(ET<sub>o</sub>/CS<sub>o</sub>)����λ����ķ�Ӧ��������(RC/CS<sub>o</sub>)�����Ӳ���������������(<em>ψ</em><sub>o</sub>��<em>φ</em><sub>Eo</sub>)����,����λ������յĹ���(ABS/CS<sub>o</sub>)��ӫ���յ�������K��(<em>W</em><sub>k</sub>)��J��(<em>V</em><sub>j</sub>)��������,˵����в�ȶԻ���ľҶƬ����������(OEC)��������PS��Ӧ����������˺�.��NaClŨ��Ϊ0.3%ʱ,PS�����⻯ѧЧ��(<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>)�⻯ѧ����ָ��(<em>PI</em><sub>ABS</sub>)�ֱ�ȶ����½� 17.7%��36.6%.</p>
URLMagsci [��������: 1]
�Զ�ӪҰ����(<em>Glycine soja </em>Sieb. et Zucc. ZYD 03262)��ɽ�������(<em>Glycine max</em> (L.) Merr. ɽ��11��)Ϊʵ�����,ͨ���о�2�ִ�ֲ�������ҶƬ�Բ�ͬŨ��NaCl(0,100��200 mmol/L)��������Ӧ,̽��2�ִ�ϻ�����NaClв����Ӧ�IJ���ͻ������������:NaCl��������ֲ���,2�ִ�ֲ��ҶƬ�Ĺ������(<em>Pn</em>)��PS�����⻯ѧЧ��(<em>Fv/Fm</em>)��PS��ʵ�ʹ⻯ѧЧ��(<em>��</em><sub>PS��</sub>)��Ҷ���غ��������Խ���,��������Ҳ���ܵ����ơ�����,NaClв�ȶ��������������ƾ���������Ұ����;Ұ����ҶƬ�е�Na<sup>+</sup>������Na<sup>+</sup>/K<sup>+</sup>ֵ���������������,��Ұ�����е�Na<sup>+</sup>����ȴ���Ը������������100��200 mmol/L NaCl����2�ִ�����ҶƬʱ,Ұ����<em>Fv/Fm</em>��<em>��</em><sub>PS��</sub>����λ����л��Է�Ӧ���ĵ���Ŀ(<em>RC/CS</em>)�⻯ѧ����ָ��(<em>PI</em>)���½�����ȴ�������������;ҶƬ�е�Na<sup>+</sup>����Ҳ���������������Щ�������,ʵ�����õ�Ұ���Ĺ�ϻ����������Ρ�����,����в��������,Ұ����ֲ��ȴ�ܹ���Ч�ر������Na<sup>+</sup>����ҶƬ�����֯,��ά�ֹ�ϻ����ϸߵĹ�ϻ���,����Ұ������������ε�ԭ��֮һ��
URLMagsci [��������: 1]
�Զ�ӪҰ����(<em>Glycine soja </em>Sieb. et Zucc. ZYD 03262)��ɽ�������(<em>Glycine max</em> (L.) Merr. ɽ��11��)Ϊʵ�����,ͨ���о�2�ִ�ֲ�������ҶƬ�Բ�ͬŨ��NaCl(0,100��200 mmol/L)��������Ӧ,̽��2�ִ�ϻ�����NaClв����Ӧ�IJ���ͻ������������:NaCl��������ֲ���,2�ִ�ֲ��ҶƬ�Ĺ������(<em>Pn</em>)��PS�����⻯ѧЧ��(<em>Fv/Fm</em>)��PS��ʵ�ʹ⻯ѧЧ��(<em>��</em><sub>PS��</sub>)��Ҷ���غ��������Խ���,��������Ҳ���ܵ����ơ�����,NaClв�ȶ��������������ƾ���������Ұ����;Ұ����ҶƬ�е�Na<sup>+</sup>������Na<sup>+</sup>/K<sup>+</sup>ֵ���������������,��Ұ�����е�Na<sup>+</sup>����ȴ���Ը������������100��200 mmol/L NaCl����2�ִ�����ҶƬʱ,Ұ����<em>Fv/Fm</em>��<em>��</em><sub>PS��</sub>����λ����л��Է�Ӧ���ĵ���Ŀ(<em>RC/CS</em>)�⻯ѧ����ָ��(<em>PI</em>)���½�����ȴ�������������;ҶƬ�е�Na<sup>+</sup>����Ҳ���������������Щ�������,ʵ�����õ�Ұ���Ĺ�ϻ����������Ρ�����,����в��������,Ұ����ֲ��ȴ�ܹ���Ч�ر������Na<sup>+</sup>����ҶƬ�����֯,��ά�ֹ�ϻ����ϸߵĹ�ϻ���,����Ұ������������ε�ԭ��֮һ��
DOI:10.3864/j.issn.0578-1752.2012.16.005URLMagsci [��������: 3]
��Ŀ�ġ��о���в�ȶԲ�ͬ����Ʒ�����������ü�Ҷ����ӫ�������Ӱ�죬Ϊ�����������������Ʒ�ֵ�ѡ��������в���˹������ṩ�������ݡ����������Ը�������Ʒ�֣�����15�ţ���������Ʒ�֣�����11�ţ�Ϊ���ϣ��˹���������Ӫ��Һ������ʪ��60%������/�ڰ�Ϊ12 h/12 h������ǿ��Ϊ134 μmol•m-2•s-1����/ҹ�¶�Ϊ28��/25�档��3Ҷ�ڿ�ʼ����NaCl������NaClŨ��Ϊ0��50��100��150��200 mmol•L-1�����о���в���¸������������ú�Ҷ����ӫ������ı仯�����������Ũ��NaCl��50 mmol•L-1��в�ȿ�������Ҷ���غ��������Ǹ�Ũ��NaCl��100—200 mmol•L-1��в�����Խ���Ҷ���غ�������в��ʹ��������ʣ�Pn���������ȣ�Gs�����������ʣ�Tr�������ӫ�⣨Fm����Fv/Fo��Fv/Fm��Fv′/Fm′���⻯ѧ���ϵ����qP���½�����ʼӫ�⣨Fo���ͷǹ⻯ѧ���ϵ����NPQ����ߣ���Ũ����в�ȣ�50 mmol•L-1 NaCl��ʹ����CO2Ũ�ȣ�Ci�����ͣ���Ũ�����෴������15������в�ȵ�Ӱ��̶�С������11�ţ����ֳ��Ϻõ������ԡ������ۡ�50 mmol•L-1Ũ�ȵĵ���в�ȶԸ��������Ӱ�첻���ԣ���������½�����Ҫԭ�����������ƣ�100—200 mmol•L-1Ũ�ȵĸ���в�ȶԸ��������кܴ�Ӱ�죬�����������½�����Ҫԭ���Ƿ��������ơ���в���£�����Ʒ������Ч�����ڲ��Ĺ�ϻ�������ǿ����в�ȵ���Ӧ�ԡ�
DOI:10.3864/j.issn.0578-1752.2012.16.005URLMagsci [��������: 3]
��Ŀ�ġ��о���в�ȶԲ�ͬ����Ʒ�����������ü�Ҷ����ӫ�������Ӱ�죬Ϊ�����������������Ʒ�ֵ�ѡ��������в���˹������ṩ�������ݡ����������Ը�������Ʒ�֣�����15�ţ���������Ʒ�֣�����11�ţ�Ϊ���ϣ��˹���������Ӫ��Һ������ʪ��60%������/�ڰ�Ϊ12 h/12 h������ǿ��Ϊ134 μmol•m-2•s-1����/ҹ�¶�Ϊ28��/25�档��3Ҷ�ڿ�ʼ����NaCl������NaClŨ��Ϊ0��50��100��150��200 mmol•L-1�����о���в���¸������������ú�Ҷ����ӫ������ı仯�����������Ũ��NaCl��50 mmol•L-1��в�ȿ�������Ҷ���غ��������Ǹ�Ũ��NaCl��100—200 mmol•L-1��в�����Խ���Ҷ���غ�������в��ʹ��������ʣ�Pn���������ȣ�Gs�����������ʣ�Tr�������ӫ�⣨Fm����Fv/Fo��Fv/Fm��Fv′/Fm′���⻯ѧ���ϵ����qP���½�����ʼӫ�⣨Fo���ͷǹ⻯ѧ���ϵ����NPQ����ߣ���Ũ����в�ȣ�50 mmol•L-1 NaCl��ʹ����CO2Ũ�ȣ�Ci�����ͣ���Ũ�����෴������15������в�ȵ�Ӱ��̶�С������11�ţ����ֳ��Ϻõ������ԡ������ۡ�50 mmol•L-1Ũ�ȵĵ���в�ȶԸ��������Ӱ�첻���ԣ���������½�����Ҫԭ�����������ƣ�100—200 mmol•L-1Ũ�ȵĸ���в�ȶԸ��������кܴ�Ӱ�죬�����������½�����Ҫԭ���Ƿ��������ơ���в���£�����Ʒ������Ч�����ڲ��Ĺ�ϻ�������ǿ����в�ȵ���Ӧ�ԡ�
URLMagsci [��������: 1]
ȫ������仯�������������Լ���Ϊ���ŵ����ؾ���ʹʪ��ֲ���ܵ���Ъ�������Ե���в�ȣ�����Ӱ�쵽ֲ��Ĵ��������ֲ��ͷ�ֳ���ڳ��ڵ���Ӧ���������У�ʪ��ֲ���γ��˶�����Ӧ��в�ȵIJ��ԣ���Ҫ�У�1)����ʷ���棬ֲ���ͨ�������ȷ�ʱ��ĵ������������ߡ�̥������ֳ��ʽ�ĸı���ӱ��ζȵ�ֱ���˺�;2)��̬ѧ���棬ֲ���ͨ������������ģʽ�ĵ����������ϻ�����Ҷ��Ӫ�����ٵ����ʻ��Ƚ������Na��+���뵽��л����Ծ�ľ��л����ų�����;3)����ѧ���棬ֲ���ͨ���������ݡ������ͨ����֯������ϸ��ľ˨�㡢���ʲ㼰դ����֯�ĺ�ȵ���ά��ֲ�������Ĺ�����úͺ�������;4)�����������棬ֲ���ͨ�����������������Ρ����Ρ�ѡ�������ա������ڡ����ص��ڼ���������ø���յ�����ά��ϸ������������ѹ��������ڻ��������ӣ�ROS��;5)����ˮƽ���棬ֲ���ͨ����������в����صĻ���������ϸ���ڵĶ��ִ�л��Ӧ���ڽ����о��У�Ca<SUP>2+</SUP>�Ը�����ϳɵĵ��ء��仯�ζ������µ���Ӧ���Լ���ϵ���ܵ�ά�ֵȷ��������һ����ǿ��
URLMagsci [��������: 1]
ȫ������仯�������������Լ���Ϊ���ŵ����ؾ���ʹʪ��ֲ���ܵ���Ъ�������Ե���в�ȣ�����Ӱ�쵽ֲ��Ĵ��������ֲ��ͷ�ֳ���ڳ��ڵ���Ӧ���������У�ʪ��ֲ���γ��˶�����Ӧ��в�ȵIJ��ԣ���Ҫ�У�1)����ʷ���棬ֲ���ͨ�������ȷ�ʱ��ĵ������������ߡ�̥������ֳ��ʽ�ĸı���ӱ��ζȵ�ֱ���˺�;2)��̬ѧ���棬ֲ���ͨ������������ģʽ�ĵ����������ϻ�����Ҷ��Ӫ�����ٵ����ʻ��Ƚ������Na��+���뵽��л����Ծ�ľ��л����ų�����;3)����ѧ���棬ֲ���ͨ���������ݡ������ͨ����֯������ϸ��ľ˨�㡢���ʲ㼰դ����֯�ĺ�ȵ���ά��ֲ�������Ĺ�����úͺ�������;4)�����������棬ֲ���ͨ�����������������Ρ����Ρ�ѡ�������ա������ڡ����ص��ڼ���������ø���յ�����ά��ϸ������������ѹ��������ڻ��������ӣ�ROS��;5)����ˮƽ���棬ֲ���ͨ����������в����صĻ���������ϸ���ڵĶ��ִ�л��Ӧ���ڽ����о��У�Ca<SUP>2+</SUP>�Ը�����ϳɵĵ��ء��仯�ζ������µ���Ӧ���Լ���ϵ���ܵ�ά�ֵȷ��������һ����ǿ��
[��������: 1]
URL [��������: 1]
URL [��������: 1]
DOI:10.5846/stxb201209291363URLMagsci [��������: 1]
�Ե�Ũ�ȣ�50 mmol/L����Ũ�ȣ�150 mmol/L��NaCl�������������ݣ�<em>Quercus virginiana</em>�������ݣ�<em>Quercus acutissima</em>��1�������磬�о���2����������в���µ����������ηֵ������Ժ������Լ����ϵ��̬ѧ�����仯�Լ���ϵ�������ӵ���������ۡ������������Ũ����в������������2���������ϲ��������Ļ��ۣ�<em>P</em>< 0.05��������Ũ����в�ȶԸ��������ݵ��ϲ����ص�Ӱ�첻���ԣ����������������ݵ��ϲ����أ�<em>P</em>< 0.05����2�������ĸ��ڱ�����в���³��������ƣ��ر����ڸ�Ũ����в���£�2�������ĸ��ڱ��������ӣ�<em>P</em>< 0.05������в���������������ڸ����ķ�����ֲ��Ӧ����в�ȵķ�ʽ֮һ��2������������������������в���±仯�����ԣ���2��������ϵ��̬ѧ��������в���µ���Ӧ��ͬ�����������ݸ�ϵ�ܳ��ȡ��ܱ���������������в���¾��в�ͬ�̶����ӣ��ر����ڵ�Ũ����в���£���ϵ��̬ѧ�����������ӣ�<em>P</em>< 0.05���������ݸ�ϵ��̬ѧ�������½����ƣ����������ȱ仯�����ԣ�ͨ���Բ�ͬ������ϵ�ܳ��ķ������֣����������ݸ�ϵ�ܳ��ȵ�������Ҫ������ֱ��С��2 mm��ϸ���ܳ������ӣ�ϸ�����ȵ����Ӷ���ֲ������ˮ�ֺ�Ӫ�����ʾ�����Ҫ���壻ͨ����Na<sup>+</sup>��Cl<sup>-</sup>�ڸ�ϵ�ĺ���������������в����2��������ϵ�����ӵĻ��۾����������ӣ������������ݸ�ϵ�����ӵĺ����ڵ�Ũ�Ⱥ�Ũ����в���µIJ��첻���ԣ��������ڸ�Ũ����в���¸�ϵ�����ӵĺ������Ը��ڸ��������ݡ��ۺ�2��������в���µ�������������Ժ�ϵ��̬ѧ��Ӧ�Լ������ӵĻ��۹��ɣ�֤��2������������������������Է��������ͬ���ص㣬����ϵ����Ӧ���Խ�Ȼ��ͬ����������������в�����ܹ������ϵ���շ�Χ��ά�ֽϸߵ�K<sup>+</sup>/Na<sup>+</sup>��ֵ������������в���¸�ϵ�������չ���������ӣ����¸�ϵ�����������ܵ����ƣ�Ӱ���˸�ϵ���澳�еķֲ���Χ���Ӷ���һ���̶��ϱ����˽�һ�����κ���
DOI:10.5846/stxb201209291363URLMagsci [��������: 1]
�Ե�Ũ�ȣ�50 mmol/L����Ũ�ȣ�150 mmol/L��NaCl�������������ݣ�<em>Quercus virginiana</em>�������ݣ�<em>Quercus acutissima</em>��1�������磬�о���2����������в���µ����������ηֵ������Ժ������Լ����ϵ��̬ѧ�����仯�Լ���ϵ�������ӵ���������ۡ������������Ũ����в������������2���������ϲ��������Ļ��ۣ�<em>P</em>< 0.05��������Ũ����в�ȶԸ��������ݵ��ϲ����ص�Ӱ�첻���ԣ����������������ݵ��ϲ����أ�<em>P</em>< 0.05����2�������ĸ��ڱ�����в���³��������ƣ��ر����ڸ�Ũ����в���£�2�������ĸ��ڱ��������ӣ�<em>P</em>< 0.05������в���������������ڸ����ķ�����ֲ��Ӧ����в�ȵķ�ʽ֮һ��2������������������������в���±仯�����ԣ���2��������ϵ��̬ѧ��������в���µ���Ӧ��ͬ�����������ݸ�ϵ�ܳ��ȡ��ܱ���������������в���¾��в�ͬ�̶����ӣ��ر����ڵ�Ũ����в���£���ϵ��̬ѧ�����������ӣ�<em>P</em>< 0.05���������ݸ�ϵ��̬ѧ�������½����ƣ����������ȱ仯�����ԣ�ͨ���Բ�ͬ������ϵ�ܳ��ķ������֣����������ݸ�ϵ�ܳ��ȵ�������Ҫ������ֱ��С��2 mm��ϸ���ܳ������ӣ�ϸ�����ȵ����Ӷ���ֲ������ˮ�ֺ�Ӫ�����ʾ�����Ҫ���壻ͨ����Na<sup>+</sup>��Cl<sup>-</sup>�ڸ�ϵ�ĺ���������������в����2��������ϵ�����ӵĻ��۾����������ӣ������������ݸ�ϵ�����ӵĺ����ڵ�Ũ�Ⱥ�Ũ����в���µIJ��첻���ԣ��������ڸ�Ũ����в���¸�ϵ�����ӵĺ������Ը��ڸ��������ݡ��ۺ�2��������в���µ�������������Ժ�ϵ��̬ѧ��Ӧ�Լ������ӵĻ��۹��ɣ�֤��2������������������������Է��������ͬ���ص㣬����ϵ����Ӧ���Խ�Ȼ��ͬ����������������в�����ܹ������ϵ���շ�Χ��ά�ֽϸߵ�K<sup>+</sup>/Na<sup>+</sup>��ֵ������������в���¸�ϵ�������չ���������ӣ����¸�ϵ�����������ܵ����ƣ�Ӱ���˸�ϵ���澳�еķֲ���Χ���Ӷ���һ���̶��ϱ����˽�һ�����κ���
DOI:10.1111/ppl.1992.86.issue-3URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1146/annurev.pp.33.060182.001533URL [��������: 1]
DOI:10.1046/j.1469-8137.2000.00667.xURL [��������: 1]
[��������: 1]
URL [��������: 2]
URL [��������: 2]
URL [��������: 1]
URL [��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}