 ,, 蔡泽江, 文石林, 吴红慧中国农业科学院农业资源与农业区划研究所/耕地培育技术国家工程实验室,北京 100081
,, 蔡泽江, 文石林, 吴红慧中国农业科学院农业资源与农业区划研究所/耕地培育技术国家工程实验室,北京 100081Quantitative Analysis of Driving-Factors of Soil Acidification in Qiyang County, Hunan Province
ZHOU HaiYan, XU MingGang,, CAI ZeJiang, WEN ShiLin, WU HongHuiInstitute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences/National Engineering Laboratory for Improving Quality of Arable Land, Beijing 100081通讯作者:
收稿日期:2018-09-29接受日期:2018-11-27网络出版日期:2019-04-16
| 基金资助: |
Received:2018-09-29Accepted:2018-11-27Online:2019-04-16
作者简介 About authors
周海燕,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (530KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
周海燕, 徐明岗, 蔡泽江, 文石林, 吴红慧. 湖南祁阳县土壤酸化主要驱动因素贡献解析[J]. 中国农业科学, 2019, 52(8): 1400-1412 doi:10.3864/j.issn.0578-1752.2019.08.010
ZHOU HaiYan, XU MingGang, CAI ZeJiang, WEN ShiLin, WU HongHui.
0 引言
【研究意义】土壤酸化是指土壤pH不断降低、土壤交换性酸不断增加的过程,主要受土壤植物体系中各种途径产生的氢离子(H+)驱动[1]。近年来,我国南方红壤大面积快速酸化,土壤pH下降了0.23—0.30个单位[2]。2017年公布的我国耕地质量监测报告也显示,我国的区域性土壤酸化问题日益显现,如:长江中游区土壤总体上呈强酸性(4.5<pH<5.5)和中度酸性(5.5<pH<6.5),两者占比高达77%[3]。这种“酸上加酸”的状况导致红壤pH已逼近铝、锰等毒性金属大量活化的临界阈值,严重威胁国家粮食安全和生态文明建设,引发了全社会的普遍关注。在区域性土壤酸化的大背景下,选择我国典型县进行整县域土壤酸化致酸因素解析和验证将对我国其他大区域尺度乃至全国的土壤酸化研究将具有重大的借鉴意义。【前人研究进展】由CO2溶解到土壤溶液中,其HCO3-淋洗出土壤,导致H+产生的碳循环过程,通常称之为自然酸化,自然酸化对总氢离子产生量贡献不足10%[4]。因此,无论是农田中的各种作物体系还是森林等自然生态系统,自然酸化的产酸量大致相同,碳循环导致的自然酸化的影响可以不予考虑。除自然酸化以外,土壤植物系统中H+来源途径包括:酸沉降(直接带入H+)、铵态氮投入和植物吸收阳离子等[5,6]。1 mol铵态氮完全硝化时,会产生2 mol的H+,因此,长期铵态氮投入会导致土壤发生明显酸化[7]。植物生长和收获会从土壤中吸收和移除盐基离子,根系每吸收1mol盐基离子,会向土壤中释放等当量H+[5]。酸雨集中分布在我国南方地区,主要包括江西、湖南、福建等长江以南的省份[8],研究上述区域的土壤酸化问题,酸沉降因素尤其重要。利用物质平衡和电荷平衡的原理,理论计算各个途径的H+产生量,定量化各个产酸途径对土壤酸化的贡献,已成为土壤酸化定量研究常用的重要方法[5,9]。ZHU等[10]采用此方法定量化了我国森林生态系统中大气沉降与森林收获和生长对土壤酸化贡献。GUO等[2]同样采用此方法定量比较了我国四大农作物系统:小麦-玉米、小麦-水稻、双季稻和保护地蔬菜的年度H+产生量。湖南省祁阳县地处我国南部土壤酸化中度敏感区的中心地带[11],于1990年建站的国家土壤肥力与肥效长期监测试验站之一的祁阳长期定位试验站就坐落在祁阳县域的北部,该试验站保存并积累了自建站到现在近30年的历史土壤样品和数据资料[12]。祁阳县总土地面积为25.2×104 hm2,其中,水田占17%,旱地农田占3%,果园占1%,森林占59%,农田和森林在祁阳县都有广泛分布,总面积占到祁阳县土地面积的80%。姬钢[13]对祁阳红壤试验站周边的旱地农田、自然林和人工林等11种土地利用方式的土壤采样检测分析结果表明,无论是施肥的农田还是森林表层土壤都酸化严重。周晓阳等[14]对湖南等南方六省的水田系统研究表明水田土壤也发生显著酸化,25年间(1988—2013年)土壤pH 下降0.59个单位。以上研究结果表明了祁阳县域的旱地农田、水田和森林的表层土壤均发生酸化,但对导致该区域土壤酸化的主要驱动因素却未能进一步定量化。【本研究切入点】前人众多的研究结果表明理论计算H+产生量是定量化研究土壤酸化致酸因素的可靠手段,但此前的许多研究都集中在某种自然生态系统或某些农作物系统,关于自然生态系统和农田生态系统同时广泛分布较为复杂的整个区域(比如祁阳县)土壤酸化驱动因素的定量研究则较少,并且也未能对所采用的理论H+产生量的计算方法的科学性进行验证。【拟解决的关键问题】以我国南方土壤显著酸化的典型县域—祁阳作为研究区域,通过搜集大量公开发表的文献、统计年鉴等,获取施肥量、主要农作物产量和林木生物量,以及地上部不同部位的养分含量等数据,基于经典的H+产生量的计算方法,解析氮循环过程、盐基离子吸收和酸沉降等3个关键过程的相对贡献大小,深化对区域土壤酸化致酸因素贡献率的认识,为我国其他区域的酸化土壤防治提供理论依据,为保障国家粮食安全和生态文明建设提供科技支撑。1 材料与方法
1.1 祁阳县的基本概况
祁阳县地处湖南省西南部,东经110o35' —112o14',北纬26o02' —26o51'。属于亚热带气候,年平均气温18.2℃,年平均降雨量1 276 mm[15]。主要土壤类型为红壤,土壤缓冲能力较低。祁阳县种植的农作物复杂多样,林木种类繁多,本文仅选择主要农作物和主要林木系统为研究对象。主要农作物定义为:根据最新的(2013年)祁阳县农业统计数据[16],常年播种面积大于2 500 hm2的农作物。在本研究中的主要农作物包括水稻、油菜、玉米、花生、大豆和甘薯6种。祁阳县的总体的林木种植面积从1980年开始到现在较为稳定,主要林木定义为:根据祁阳县农业统计数据,种植面积大于100 hm2的林木。按照此标准,本研究的主要林木包含马尾松、湿地松、杉树、竹林、油茶林、板栗和柑橘7种。1.2 主要农作物和柑橘的施肥量及其肥料形态和养分含量
本研究通过施肥量和肥料形态计算施肥带入的养分量。目前我国化肥市场氮肥的形态主要为尿素(尿素占我国氮肥产量的64%—68%)和铵态氮(碳酸氢铵占我国氮肥产量的4%,氯化铵占6%,复合肥中的氮占18%),两者占我国氮肥产品总用量的95% [17],因此,本研究假设尿素态氮和铵态氮在各种农作物的年度氮肥投入量中各占一半。磷肥形态主要有磷酸二铵、磷酸一铵和过磷酸钙,占我国磷肥组成类型的90%以上[17],因此,本研究假设施入的磷肥中磷酸二铵(N 19%,P2O5 46%)、磷酸一铵(N 12%,P2O5 51%)和过磷酸钙(P2O5 12%,CaO 12%)的比例分别为45%、35%和20%,因为过磷酸钙在磷肥施用总量中占比最低,且含有的游离酸的质量分数≤5.5%(GB/T 20413—2017),因此本研究不考虑由于施用过磷酸钙直接带入的游离酸。氯化钾占所有消费钾肥形态的95%以上[18],因此,为简化计算,本研究假设施用的钾肥均为氯化钾。祁阳县农户施用有机肥数量较少,本研究不考虑有机肥带入的养分量。在农田中,施入的氮肥是主要的氮源,因此,土壤中的有机氮矿化不予考虑。祁阳县主要农作物和柑橘通过施肥和大气沉降带入的氮磷钾钙镁等养分的年度输入量见表1。Table 1
表1
表1主要农作物和柑橘的年度养分输入量
Table 1
| 输入形式 Input | 作物类型 Crop | N (kg·hm-2·a-1) | P2O5 (kg·hm-2·a-1) | K2O (kg·hm-2·a-1) | Ca (kg·hm-2·a-1) | Mg (kg·hm-2·a-1) |
|---|---|---|---|---|---|---|
| 化肥带入 Chemical fertilizer input | 水稻 Rice | 180 | 59 | 78 | 1.4 | - |
| 玉米Corn | 216 | 65 | 42 | 1.6 | - | |
| 花生Peanut | 120 | 52 | 70 | 1.2 | - | |
| 油菜Rape | 160 | 51 | 74 | 1.2 | - | |
| 甘薯Sweet potato | 102 | 45 | 51 | 1.1 | - | |
| 大豆Soybean | 165 | 41 | 41 | 1.0 | - | |
| 柑橘Citrus | 239 | 83 | 93 | 2.0 | - | |
| 大气沉降 Atmospheric deposition | 47.6 | - | 2.8 | 15 | 2.4 |
新窗口打开|下载CSV
1.3 氢离子(H+)产生量的计算方法
本研究基于经典物质和电荷平衡定量产酸量的方法,将土壤的H+产生(消耗)量分为4个独立的部分,包括氮循环过程(N)、盐基阳离子吸收(BC)、酸沉降(H)和磷吸收(P)过程,分别进行计算。各种致酸因素的总产酸量等于N、BC和H这三个主要过程的H+产生量的总和。净H+产生量等于总H+产生量减去P吸收过程的消耗H+量。主要是因为植物根系每吸收1 mol的H2PO4-,则产生1 mol的OH-,即消耗1 mol的H+[2]。本研究未考虑SO42-和Cl-的被植物根系吸收产生的OH-,主要是因为主要农作物和林木对SO42-和Cl-吸收量较少。1.3.1 氮循环过程的H+产生量的计算 农田氮转化过程包括氨挥发、硝化作用、反硝化、淋洗和农作物根系吸收等。农作物的吸氮量则采用籽粒和秸秆的产量分别乘以籽粒和秸秆氮的养分浓度(表2)计算得到。氨挥发、反硝化和淋洗这3个氮转化过程是利用CUI等[19,20]建立旱地农田和水田的施氮量(Nrate)与氮损失途径间的经验模型计算得到祁阳主要农作物种植体系的氮输出通量(表3)。祁阳县氮沉降的总量为47.6 kg·hm-2·a-1 [9],铵态氮含量是硝态氮的3.5倍[28],因此,祁阳作为一个县级区域,无论是农田还是林地使用同一套沉降量参数,铵态氮沉降量为37.1 kg·hm-2·a-1,硝态氮沉降量为10.5 kg·hm-2·a-1。
Table 2
表2
表2祁阳县主要农作物籽粒秸秆的养分含量[21,22,23,24,25]
Table 2
| 作物 Crop | 面积 Area (hm2) | 籽粒产量 Grain yield (kg·hm-2) | 秸秆产量 Straw yield (kg·hm-2) | P(%) | K(%) | Ca(%) | Mg(%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 籽粒 Grain | 秸秆 Straw | 籽粒 Grain | 秸秆 Straw | 籽粒 Grain | 秸秆 Straw | 籽粒 Grain | 秸秆 Straw | ||||
| 水稻 Rice | 71133 | 6660 | 5997 | 0.35 | 0.176 | 0.19 | 1.99 | 0.03 | 0.54 | 0.12 | 0.212 |
| 玉米Corn | 3224 | 3420 | 4104 | 0.26 | 0.152 | 0.34 | 1.18 | 0.013 | 0.54 | 0.120 | 0.224 |
| 花生Peanut | 2863 | 2610 | 2093 | 0.5 | 0.163 | 0.85 | 1.09 | 0.074 | 1.76 | 0.255 | 0.56 |
| 油菜Rape | 6144 | 1470 | 2195 | 1.47 | 0.144 | 7.77 | 1.94 | - | 1.52 | 0.94 | 0.25 |
| 甘薯Sweet potato | 6023 | 4320 | 2159 | 0.17 | 0.283 | 0.73 | 3.05 | 0.14 | 2.11 | 0.073 | 0.46 |
| 大豆Soybean | 4401 | 3045 | 4880 | 0.85 | 0.196 | 2.17 | 1.17 | 0.26 | 1.71 | 0.24 | 0.48 |
新窗口打开|下载CSV
Table 3
表3
表3祁阳典型种植体系氮的输入-输出通量
Table 3
| 作物类型 Cropping systems | 施氮量 N rate (kg·hm-2·a-1) | 氮沉降 N deposition (kg·hm-2·a-1) | 秸秆吸氮量 Straw N uptake (%) | 籽粒吸氮量 Grain N uptake (%) | NH3挥发 NH3 volatilization (%) | NO3-N淋洗量 NO3-N leaching (%) |
|---|---|---|---|---|---|---|
| 水稻 Rice | 180 | 47.6 | 1.20 | 1.21 | 17.3 | 2.9 |
| 玉米Corn | 216 | 47.6 | 0.92 | 1.15 | 24.5 | 20.2 |
| 花生Peanut | 120 | 47.6 | 1.82 | 4.57 | 24.8 | 12.9 |
| 油菜Rape | 160 | 47.6 | 0.87 | 8.67 | 24.6 | 15.1 |
| 甘薯Sweet potato | 102 | 47.6 | 2.37 | 0.33 | 25.3 | 11.4 |
| 大豆Soybean | 165* | 47.6 | 1.81 | 7.82 | 25.0 | 11.7 |
| 柑橘Citrus | 239 | 47.6 | - | - | 24.6 | 17.7 |
新窗口打开|下载CSV
N过程是H+产生的关键途径,其H+产生量为硝态氮的净输出和铵态氮的净输入之和[7, 29](公式1)。
式中,NO3-in、NH4+in分别表示硝态氮和铵态氮的输入,NO3-out、 NH4+out分别表示硝态氮和铵态氮的输出。
农田和林地的氮输入和输出过程存在差异,需要分别进行计算。
农田:如果氮肥形态是铵态氮。则计算公式为2:
式中,NO3-淋洗、NO3-沉降分别表示投入的氮形态为铵态氮时,硝态氮的淋洗量和沉降量;NH4+投入表示铵态氮的输入量,包括来源于化肥和沉降两部分的总量,NH4+out表示铵态氮的输出。
农田:如果氮肥形态是尿素。假设尿素施入后全部迅速转化为铵态氮,仅以硝态氮形式淋洗。因此,尿素施入土壤后,1 mol尿素全部水解消耗1 mol的H+离子,水解产生的铵态氮发生氨挥发释放1 mol的H+离子,则氨挥发途径的净氢离子产生量为0;水解产生的铵态氮发生硝化作用产生2 mol的H+离子和1 mol 的NO3-,NO3-离子被植物吸收则消耗1 mol的H+离子,因此,被农作物吸收的N途径的净氢离子产生量为0,每1 mol的NO3-离子淋洗,则产生1 mol的H+离子,计算公式简化为公式3:
式中,NO3-淋洗、NO3-沉降分别表示投入的氮形态为尿素时,硝态氮的淋洗量和沉降量。
林地:一年中进入森林土壤中的总沉降氮是多次沉降的总和,而且森林土壤通常都处在氮不饱和的条件下,因此,假定大气沉降中的NH4+被迅速吸收或硝化,仅以NO3-淋洗,剩余的N全部被地上部吸收。基于周才平等[30]对我国森林生态系统的年有机氮矿化数量的量化研究结果,将祁阳区域林地的有机氮矿化量设定为110 kg·hm-2·a-1。因此,林木地上部吸收的总N主要来源于有机氮矿化和氮沉降,本研究中按照这两种氮源分别进行H+产生量的计算。根据ZHU等[10]的研究结果,将祁阳林地的氮淋洗系数设定为0.25,即N淋洗 / N总 = 0.25。来源于氮沉降的铵态氮,被林木每吸收1 mol N,则产生1 mol H+;1 mol N发生淋洗,则产生2 mol H+。来源于有机氮矿化的氮,被林木吸收1 mol N,则不产生H+;1 mol N发生淋洗,则产生1 mol H+。氮沉降和有机氮矿化来源的氮的H+产生量计算公式分别为(4)和(5):
式中,NO3-in、NH4+in分别表示沉降过程硝态氮和铵态氮的输入,NO3-out、 NH4+out分别表示沉降过程硝态氮和铵态氮的输出。
式中,NO3-淋洗表示有机氮矿化过程的硝态氮淋洗量。
将柑橘作为一种特殊的林地类型处理,常年施N量为239kg·hm-2(近5年施氮量的平均值),因此,柑橘园的氮源主要是施用的化肥和沉降带入的氮,两者的总和为286.6 kg·hm-2。根据巨晓棠等[31]对农田有机氮矿化的研究结果,农田土壤的有机氮矿化量随土壤施氮量的增加而显著降低,施氮量达到300 kg·hm-2时,有机氮的净矿化量几乎为零。基于以上原因,不考虑有机氮矿化产生的氮。柑橘的管理和施肥措施与旱地农田最为相近,因此,氮输出通量按照旱地农田[24,25]的计算方法进行,具体结果见表3,氮磷钾钙镁等各种养分的输入通量详见表1。
1.3.2 BC过程H+产生量的计算 根据电荷守恒,土壤溶液中的BC被植物吸收,植物体将释放H+,因此,BC过程的H+产生量为农作物籽粒和秸秆的收获的BC净移除量或林木的地上部(包括枝、干、果)BC吸收量。相比于Ca2+、Mg2+、K+等阳离子,植物对Na+的吸收量很少,因此,本研究不考虑Na+吸收产生的H+。主要农作物的籽粒秸秆的产量及养分含量见表2。林木的树叶通常认为归还土壤,不计算在内,主要林木的地上部生物量及养分含量见表4和表5。计算公式为6。
式中,BC = Ca2++Mg2++K+,BCupt表示森林生长吸收的盐基离子数量,等于生物量与盐基离子浓度的乘积。
Table 4
表4
表4祁阳县主要用材林木生物量及枝干皮养分含量[34,35,36]
Table 4
| 林木类型 Tree species | 面积 Area (hm2) | 生物量 Biomass (kg·hm-2) | 树干养分含量 Element content in stem wood (%) | 树枝养分含量 Element content in branch wood (%) | 树皮养分含量 Element content in bark wood (%) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 树干Stem | 树枝 Branch | 树皮Bark | N | P | K | Ca | Mg | N | P | K | Ca | Mg | N | P | K | Ca | Mg | ||
| 湿地松 Slash pine | 21080 | 34059 | 9874 | 7095 | 0.194 | 0.008 | 0.105 | 0.214 | 0.019 | 0.485 | 0.036 | 0.256 | 0.867 | 0.07 | 0.266 | 0.018 | 0.086 | 0.309 | 0.031 |
| 杉木 Fir | 20876 | 62406 | 9664 | 8386 | 0.075 | 0.007 | 0.031 | 0.067 | 0.015 | 0.443 | 0.032 | 0.289 | 0.543 | 0.207 | 0.283 | 0.024 | 0.231 | 0.432 | 0.045 |
| 马尾松Pine | 7905 | 67400 | 12100 | 5910 | 0.17 | 0.014 | 0.121 | 0.235 | 0.05 | 0.381 | 0.026 | 0.143 | 0.164 | 0.099 | 0.477 | 0.042 | 0.353 | 0.856 | 0.144 |
| 竹 Bamboo | 7108 | 42685 | 3488 | - | 0.247 | 0.016 | 0.225 | 0.013 | 0.029 | 0.386 | 0.017 | 0.108 | 0.015 | 0.02 | - | - | - | - | - |
新窗口打开|下载CSV
Table 5
表5
表5祁阳县主要经济林生物量及枝干和果实养分含量[34,35,36,37]
Table 5
| 林木类型 Tree species | 面积 Area (hm2) | 生物量 Biomass (kg·hm-2) | 枝干养分含量 Element content in stem wood (%) | 果实养分含量 Element content in fruits (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 枝干 Stems and branches | 果实 Fruits | N | P | K | Ca | Mg | N | P | K | Ca | Mg | ||
| 板栗Chestnut | 137 | 13725 | 3750 | 0.978 | 0.186 | 0.253 | 1.400 | 0.210 | 1.376 | 0.27 | 0.664 | 2.048 | 0.222 |
| 油茶林Camellia oleifera | 28513 | 7681 | 1798 | 0.65 | 0.045 | 0.31 | 0.129 | 0.131 | 0.831 | 0.093 | 0.971 | 0.437 | 0.129 |
| 柑橘Citrus | 2068 | 26190 | 40397 | 0.513 | 0.070 | 0.420 | 1.193 | 0.247 | 0.158 | 0.052 | 0.236 | 0.062 | 0.016 |
新窗口打开|下载CSV
1.3.3 酸沉降过程H+产生量的计算 祁阳地处我国南方酸雨较为严重的区域,因此,酸沉降不可忽略,主要由酸雨形式直接带入,基于湖南省环境质量状况公报(2002—2016)[32,33]公布的数据,将酸雨pH和酸雨频度分别取多年平均值,本研究假设整个祁阳县域的酸雨降雨量和频度是相同的,酸雨的pH为5.2,酸雨频度为34.7%。土壤中H+通常不与NO3-等阴离子伴淋洗,植物也很少吸收H+,因此,假定H+out 为零,H过程的产酸量为酸雨带来的氢离子数量(公式7)。
式中,H+in表示氢离子的直接输入。
1.3.4 磷吸收过程H+产生量的计算 因为H2PO4-是土壤中常见的阴离子,也是植物吸收的主要磷形态。1 mol的H2PO4-被植物吸收,产生1 mol的OH-,即产生-1 mol H+(公式8)。
式中,H2PO4-upt分别表示H2PO4-的吸收量,为生物量(或产量)与养分浓度的乘积。
1.4 土壤缓冲性能的测定
土壤酸碱缓冲性能采用酸碱滴定法测定[13]。加入的酸碱量与对应的土壤pH作图得到红壤近S型土壤缓冲曲线,其中,在土壤pH 4.0—6.0的区间范围内,土壤pH(y)与加酸量(x)之间呈现y = a - bx的直线关系(0<x<10 mmol·kg-1;a、b为数值型系数),利用此直线方程可以计算得到加入一定量酸后的土壤pH[38]。因此,如果要得到理论上土壤中加入一定酸后土壤pH变化情况,则直接把x(H+净产量,即加酸量)代入该直线方程计算得到y,即土壤pH的模拟值。1.5 理论H+产生量的计算方法的验证
土壤pH的降低受到两方面因素的影响,一是土壤的缓冲性能,缓冲能力越强的土壤,在外界酸输入量一定的情况下,其土壤pH下降的越缓慢;二是外界酸的输入量,已知土壤的缓冲性能和外界的酸输入量即可计算得到土壤pH的下降量。因此,将外界理论计算得到的酸输入量带入土壤缓冲曲线方程,通过比较理论土壤pH与实测土壤pH的吻合程度,即可以判断理论计算方法的准确性。本研究利用国家土壤肥力与肥效长期监测中的祁阳长期定位试验站的实际观测数据,对该H+产生量的计算方法进行校验。祁阳县农户常规施肥措施以配合施用化肥为主,因此,我们选择NPK处理作为研究对象,该处理与农民常规施肥最为接近。本长期定位试验从1990年开始,采用小麦-玉米一年两熟轮作制,初始耕层土壤pH 5.7。肥料用量为每年施用N(尿素,N46%)300 kg·hm-2、P2O5(过磷酸钙,P2O5 12%) 120 kg·hm-2、K2O(KCl,K2O 60%)120 kg·hm-2。小麦和玉米籽粒产量、秸秆产量,籽粒和秸秆中P、K、Ca、Mg等养分含量(表6)均来自专著和发表的文献[4, 12-13, 39-40]。1.6 数据统计分析方法
本研究采用Microsoft Excel 2010软件进行数据整理、统计和分析。采用国际上校验模型的均方根误差(RMSE)方法来评价模拟值与实测值之间的相对差异程度[41]。Table 6
表6
表6小麦-玉米长期试验籽粒秸秆产量及养分含量
Table 6
| 作物类型 Crop types | 平均产量Average yield (kg·hm-2·a-1) | 养分含量 Element content (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1990-1995 | 1996-2000 | 2001-2005 | 2006-2010 | 2011-2012 | P | K | Ca | Mg | |||
| 小麦 Wheat | 籽粒Grain | 1788 | 905 | 928 | 374 | 316 | 0.364 | 0.48 | 0.047 | 0.15 | |
| 秸秆 Straw | 1967 | 995 | 1020 | 411 | 347 | 0.80 | 1.05 | 0.52 | 0.17 | ||
| 玉米 Corn | 籽粒 Grain | 3369 | 3237 | 1858 | 1326 | 1188 | 0.26 | 0.39 | 0.007 | 0.12 | |
| 秸秆 Straw | 3032 | 2913 | 1672 | 1194 | 1088 | 0.15 | 1.18 | 0.54 | 0.22 | ||
新窗口打开|下载CSV
2 结果
2.1 不同土地利用方式致酸过程的H+产生量及贡献率
3种土地利用方式中,旱地农田单位面积的H+净产量(产酸量)最高,达到19.0 kmol·hm-2·a-1,其次为水田,林地的产酸量最低,旱地农田产酸量约为林地产酸量的6倍(表7)。对于整个祁阳县域,氮循环过程的产酸贡献率为66.5%,盐基吸收为33.0%,酸雨则仅为0.5%。氮循环过程产酸贡献率是盐基吸收过程的2倍,更是酸雨直接带入酸量的133倍,无论是旱地农田、水田还是林地,氮循环过程都是产生H+的主要来源,是导致土壤酸化的主控因素。Table 7
表7
表7不同土地利用类型的关键致酸过程的H+产生量
Table 7
| 利用类型 Land use pattern | 氮循环 N cycling process | 盐基吸收 BC uptake | 酸雨 Acid deposition | 总H+产量 Total H+ production (kmol·hm-2·a-1) | 磷吸收 P uptake (kmol·hm-2·a-1) | H+净产量 H+ net production (kmol·hm-2·a-1) | |||
|---|---|---|---|---|---|---|---|---|---|
| H+产量 H+ production (kmol·hm-2·a-1) | 贡献率 Contribution (%) | H+产量 H+ production (kmol·hm-2·a-1) | 贡献率 Contribution (%) | H+产量 H+ production (kmol·hm-2·a-1) | 贡献率 Contribution (%) | ||||
| 林地 Forest soil | 2.2 | 68.2 | 1.0 | 30.1 | 0.056 | 1.7 | 3.2 | -0.05 | 3.2 |
| 水田Paddy | 11.5 | 65.3 | 6.1 | 34.4 | 0.056 | 0.3 | 17.6 | -1.1 | 16.5 |
| 旱地Upland | 13.6 | 68.8 | 6.1 | 30.9 | 0.056 | 0.3 | 19.7 | -0.7 | 19.0 |
| 全县* Whole county | 7.3 | 66.5 | 3.6 | 33.0 | 0.056 | 0.5 | 10.9 | -0.5 | 10.4 |
新窗口打开|下载CSV
2.2 不同农作物体系致酸过程的H+产生量及贡献率
在祁阳县域的6种主要农作物种植体系中,不同的农作物体系产酸量存在差异,范围从10.1 kmol·hm-2·a-1到30.0 kmol·hm-2·a-1(表8)。产酸量最高作物体系是大豆,其余由多到少依次为油菜、花生、水稻、玉米和甘薯,经济作物(油菜、花生和大豆,也属于油料作物)的产酸量的平均值为23.5 kmol·hm-2·a-1,均明显高于粮食作物(水稻和玉米的平均值11.9 kmol·hm-2·a-1)。从不同致酸过程对比来看,水稻、玉米、花生、油菜、大豆的土壤酸化的关键致酸过程是氮循环过程,致酸贡献率从65.3%—78.3%。而甘薯却不同,盐基吸收与氮循环过程的致酸贡献率基本相当,甘薯属于收获块根的农作物,平均施氮量最低,仅为102 kmol·hm-2·a-1。从3种不同盐基离子吸收导致的产酸量来看,水稻、玉米和油菜这3种农作物,不同盐基吸收产酸量K>Ca>Mg,K是盐基吸收过程的主要致酸离子,而且水稻和油菜两种作物的K吸收的产酸量大于钙镁两者吸收产酸量之和。花生、甘薯和大豆的不同盐基吸收产酸量Ca>K>Mg,Ca是盐基吸收过程主要致酸因子。Table 8
表8
表8不同农作物体系的关键致酸过程的H+产生量
Table 8
| 作物类型 Crop system | 氮循环过程 N cycling process | 盐基吸收 BC uptake (kmol·hm-2·a-1) | 盐基吸收贡献率 BC uptake contribution (%) | 磷吸收 P uptake process (kmol·hm-2·a-1) | H+净产量 H+ net production (kmol·hm-2·a-1) | |||
|---|---|---|---|---|---|---|---|---|
| H+产量 H+ production (kmol·hm-2·a-1) | 贡献率 Contribution (%) | K | Ca | Mg | ||||
| 甘薯 Sweet potato | 4.8 | 45.3 | 2.49 | 2.57 | 0.65 | 54.2 | -0.439 | 10.1 |
| 玉米 Corn | 12.2 | 78.3 | 1.54 | 1.13 | 0.66 | 21.4 | -0.488 | 15.1 |
| 水稻 Rice | 11.5 | 65.3 | 3.35 | 1.67 | 1.04 | 34.4 | -1.096 | 16.5 |
| 花生 Peanut | 14.6 | 78.3 | 1.15 | 1.93 | 0.92 | 21.4 | -0.532 | 18.1 |
| 油菜 Rape | 16.5 | 71.2 | 4.00 | 1.66 | 0.96 | 28.6 | -0.795 | 22.4 |
| 大豆 Soybean | 21.8 | 70.1 | 3.15 | 4.56 | 1.53 | 29.7 | -1.147 | 30.0 |
新窗口打开|下载CSV
2.3 不同林地体系致酸过程的H+产生量及贡献率
在7种林地类型中,柑橘是唯一进行人为施肥的林地类型,产酸量最高,达27.8 kmol·hm-2·a-1,其余依次为板栗>油茶林>马尾松>杉木>竹>湿地松(表9),而且经济林(柑橘、板栗、油茶林)的平均产酸量13.8 kmol·hm-2·a-1 高于用材林(马尾松、杉木、竹、湿地松)的平均产酸量2.3 kmol·hm-2·a-1,经济林是用材林产酸量的6倍之多。比较不同林木体系的致酸因素贡献率可以看出,湿地松、杉木、马尾松、竹、油茶林和柑橘土壤酸化的关键致酸过程是氮循环过程,致酸贡献率从62.8%—80.8%。而板栗与其他林地类型不同,盐基吸收致酸贡献率为(53.3%)稍高于氮循环过程致酸贡献率(46.1%)。Ca是湿地松、杉木、马尾松、竹和板栗的盐基吸收过程的主要致酸因子,而柑橘的盐基吸收过程的主要致酸因子是K。Table 9
表9
表9不同林地的关键致酸过程的H+产生量
Table 9
| 林地类型 Forest system | 氮循环过程 N cycling process | 盐基吸收 BC uptake (kmol·hm-2·a-1) | 盐基吸收贡献率 BC uptake contribution (%) | 磷吸收 P uptake process (kmol·hm-2·a-1) | H+净产量 H+ net production (kmol·hm-2·a-1) | |||
|---|---|---|---|---|---|---|---|---|
| H+产量 H+ production (kmol·hm-2·a-1) | 贡献率 Contribution (%) | K | Ca | Mg | ||||
| 湿地松 Slash pine | 1.3 | 66.8 | 0.09 | 0.45 | 0.06 | 30.4 | -0.012 | 2.0 |
| 竹 Bamboo | 1.6 | 77.0 | 0.17 | 0.17 | 0.09 | 20.3 | -0.024 | 2.1 |
| 杉木 Fir | 1.4 | 62.8 | 0.11 | 0.50 | 0.18 | 34.7 | -0.019 | 2.3 |
| 马尾松 Pine | 1.9 | 69.3 | 0.15 | 0.33 | 0.32 | 28.7 | -0.033 | 2.8 |
| 油茶林 Camellia oleifera | 2.1 | 63.9 | 0.48 | 0.42 | 0.23 | 34.4 | -0.060 | 3.2 |
| 板栗 Chestnut | 5.0 | 46.1 | 0.68 | 4.31 | 0.80 | 53.3 | -0.368 | 10.5 |
| 柑橘 Citrus | 23.0 | 80.8 | 2.58 | 2.03 | 0.80 | 19.0 | -0.702 | 27.8 |
新窗口打开|下载CSV
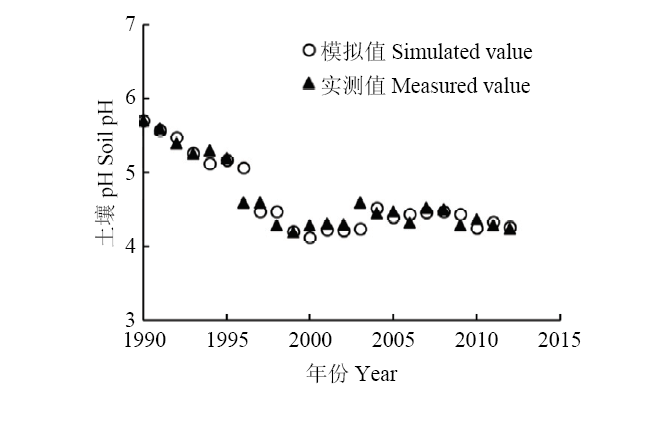
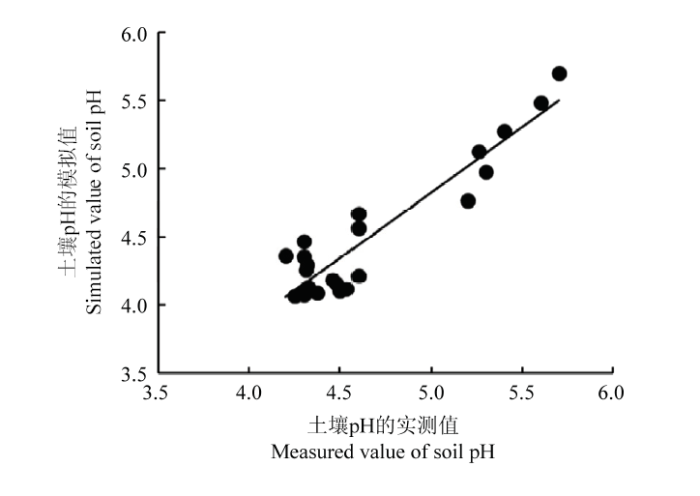
2.4 土壤酸化H+产生量计算方法的验证
根据本论文材料与方法中采用的理论H+产生量的计算方法得到祁阳小麦-玉米长期试验的NPK处理的H+净产量(表10),代入实测的土壤缓冲曲线直线段方程(表10)得到的土壤pH模拟值。将土壤pH实测值与模拟值做比较(图1),土壤pH的模拟值能较好的反映土壤pH实测值的变化,两者的吻合度较高,而且两者存在极显著的正相关(P<0.001),回归线的斜率为0.9782,R2为0.899(图2),均方根误差(RMSE)为0.15,说明本研究采用的计算氢离子产生量的方法科学准确。Table 10
表10
表10祁阳小麦-玉米长期试验田的土壤缓冲性能及年度理论氢离子产生量
Table 10
| 年份 Year | y = a - bx | H+净产量 H+ net production (kmol·hm-2·a-1) | ||
|---|---|---|---|---|
| a | b | R2 | ||
| 1991 | 5.70 | 0.0602 | 0.9698 | 5.02 |
| 1992 | 5.60 | 0.0631 | 0.9639 | 5.02 |
| 1993 | 5.40 | 0.0610 | 0.9855 | 5.02 |
| 1994 | 5.26 | 0.0650 | 0.9566 | 5.02 |
| 1995 | 5.30 | 0.0593 | 0.9551 | 5.02 |
| 1996 | 5.20 | 0.0701 | 0.9270 | 4.51 |
| 1997 | 4.60 | 0.0601 | 0.9359 | 4.51 |
| 1998 | 4.60 | 0.0603 | 0.9728 | 4.51 |
| 1999 | 4.30 | 0.0489 | 0.9639 | 4.51 |
| 2000 | 4.20 | 0.0400 | 0.9229 | 4.51 |
| 2001 | 4.30 | 0.0393 | 0.9566 | 3.76 |
| 2002 | 4.32 | 0.0593 | 0.9889 | 3.76 |
| 2003 | 4.31 | 0.0434 | 0.9387 | 3.76 |
| 2004 | 4.60 | 0.0439 | 0.9855 | 3.76 |
| 2005 | 4.46 | 0.0383 | 0.9591 | 3.76 |
| 2006 | 4.48 | 0.0292 | 0.9852 | 3.20 |
| 2007 | 4.33 | 0.0489 | 0.9607 | 3.20 |
| 2008 | 4.53 | 0.0373 | 0.9229 | 3.20 |
| 2009 | 4.50 | 0.0383 | 0.9579 | 3.20 |
| 2010 | 4.30 | 0.0312 | 09851 | 3.20 |
| 2011 | 4.38 | 0.0292 | 0.9889 | 3.11 |
| 2012 | 4.30 | 0.0219 | 0.9589 | 3.11 |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同年份的土壤pH实测值及模拟值的变化
Fig. 1The changes of simulated value and measured value of soil pH in different years
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2土壤pH的实测值与模拟值的对比
Fig. 2Comparison of simulated value and measured value of soil pH
3 讨论
3.1 区域土壤酸化的主要驱动因素贡献率
ZENG等[9]使用VSD+模型模拟了长期施肥(祁阳旱地、重庆、公主岭长期定位试验)农田土壤pH、盐基饱和度的变化,发现施用氮肥,硝酸根淋洗和铵态氮投入占质子产生量的80%,与本研究研究结果一致,氮循环过程是导致土壤酸化的主控因素。氮肥的累积施用量较少时,氮循环与BC吸收的H+产生量相当[9],本研究中甘薯(甘薯的施氮量仅为102 kg·hm-2,仅为主要农作物平均施氮量的一半)的研究结果印证了这一结论。我国四大耕作系统(双季稻、水稻-小麦、小麦-玉米和温室大棚蔬菜)中,GUO等[2]发现我国酸化农田三大粮食作物体系(小麦-玉米,水稻-小麦和双季稻)一年两季的氮循环过程的H+产生量20—33 kg·hm-2,换算为一年单季的H+产生量则为10—16 kmol·hm-2,与本研究的水稻、玉米一年单季的氮过程的H+产生量11.5 kmol·hm-2和12.2 kmol·hm-2基本相当。
盐基吸收产生的H+量存在偏好,K或Ca是盐基吸收过程产酸最多的元素,宋文峰等[42]的研究结果也表明小麦对K或Ca的吸收是盐基吸收过程产酸量最多的元素,因此,在实施土壤酸化改良调控时要注意补充K或Ca元素。
尽管酸雨是我国一个重要的区域性环境问题,出现酸雨(降雨pH达到4.0—5.6)的城市比例为38.8%[33],但该研究区域每年通过酸雨带入的酸量仅为0.056 kmol·hm-2,与氮循环和盐基离子吸收产生的酸化潜势相比,酸雨对农田土壤酸化的贡献相对较小,GUO等[2]对我国农田的研究结果也表明酸雨的贡献远低于氮循环过程和盐基吸收的贡献。
因此,减少氮循环过程的H+产生量(减少施用铵态氮肥,提高氮肥利用率)和维持土壤中盐基离子库的平衡是防治土壤酸化的有效措施。
3.2 不同土地利用方式下的植物系统产酸量
本研究中,不同农作物种植体系或者不同森林类型下,产酸量表现出很大的差异(表8,表9),这个结果和以前的研究结果[2, 9-10]相一致,经济作物体系的产酸量明显高于粮食作物体系,因此,种植经济作物的土壤pH下降幅度大,酸化程度要高于种植粮食作物的土壤[2]。本研究区域林地体系的平均盐基吸收贡献的产酸量(0.97 kmol·hm-2)较全国森林土壤酸化贡献中盐基吸收每年贡献的产酸量(0.52 kmol·hm-2)[10]相比有所提高,主要是因为全国尺度森林土壤酸化研究中包括了杨树、落叶松、桦木和栎树等多种落叶树种,与本研究的树种差异较大,而且未将树皮考虑在内,树皮的平均生物量占树体总生物量的10%(表4,表5),树皮同树木枝干一起生长或砍伐时会带走盐基离子导致土壤酸化。
3.3 理论计算的H+产生量与土壤实际酸化程度之间的关系
土壤pH的降低受到两方面因素的影响,一是土壤的缓冲性能,缓冲能力越强的土壤,在外界酸输入量一定的情况下,其土壤pH下降的越缓慢;二是外界酸的输入量,已知土壤的缓冲性能和外界的酸输入量即可计算得到土壤pH下降量。因此,将外界理论计算得到的酸输入量带入土壤缓冲曲线方程,通过比较理论土壤pH与实测土壤pH的吻合程度,即可以判断理论计算方法的准确性。祁阳小麦-玉米长期定位试验的土壤pH观测值与模拟值之间的均方根误差(RMSE)为0.15,模拟值与观测值之间的一致性较好,表明本研究采用的H+产生量的理论计算方法科学可靠,本研究提供了另外一种可行的方法验证理论计算的H+产生量与土壤实际酸化程度之间的匹配程度。之前,有许多文献都从我国农田[2]或者森林[10]等主要生态系统角度分别评估了土壤酸化的H+产生量,采用模型[9]或者H+产生量与土壤pH降低幅度之间的相关分析[10, 42]等方法验证了通过物质循环过程理论计算的H+能够准确同土壤实际酸化量相匹配。而像植物类型多样,旱地、水田和森林多种土地利用方式共存,既有人为干扰又有自然系统的这样的全区域尺度土壤酸化研究,以往的单独针对农田或者森林的定量经验模型是否适用于多种植被类型和多种土地利用方式县域尺度还不清楚。本研究很好的回答了这个问题,发现H+产生量的定量化的方法同样也适用于土地利用方式多样(3种)、农作物类型多样(6种)和林地植被类型多样(7种)等多种土地利用方式和多种植被类型都广泛分布的区域。
4 结论
祁阳县域3种土地利用方式下,产酸量的大小依次为:旱地农田>水田>林地;6种农作物体系的产酸量大小依次为:大豆>油菜>花生>水稻>玉米>甘薯;7种林地类型的产酸量大小依次为:柑橘>板栗>油茶>马尾松>杉木>竹>湿地松,不同的农作物和不同林地体系的产酸量存在很大差异,且主要农作物的产酸量普遍高于用材林(马尾松、杉木、竹、湿地松)。从3个关键致酸因素贡献率来看,氮循环过程是土壤酸化的主控因素,产酸贡献率高达66.5%,K或Ca吸收是盐基吸收过程的主要致酸离子。采用“长期定位试验+土壤缓冲性”的方法验证了经典的H+产生量的计算方法同样适用于多种植被类型和多种土地利用方式广泛分布,既有人为干扰又有自然系统的全区域土壤酸化研究。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.1126/science.1182570URL [本文引用: 8]
URL [本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
DOI:10.1016/j.atmosenv.2016.04.023URL [本文引用: 6]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[D].
[本文引用: 3]
[D].
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/gcb.12213URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1029/1999GB900015URL [本文引用: 2]
DOI:10.3864/j.issn.0578-1752.2011.20.009Magsci [本文引用: 2]

弄清中国不同区域农田养分投入、产出和平衡状况,对养分资源管理和肥料的科学分配与施用,提高肥料利用效率等具有重要意义。在参考现有资料的基础上,系统分析和论述了中国的养分资源及其利用现状,估算了各种有机肥资源及其可提供的氮磷钾养分数量,分析和评价了中国不同区域养分输入、输出特点及平衡状况。针对目前养分输入、输出存在的一系列问题,提出了养分资源管理的对策和建议。
DOI:10.3864/j.issn.0578-1752.2011.20.009Magsci [本文引用: 2]
弄清中国不同区域农田养分投入、产出和平衡状况,对养分资源管理和肥料的科学分配与施用,提高肥料利用效率等具有重要意义。在参考现有资料的基础上,系统分析和论述了中国的养分资源及其利用现状,估算了各种有机肥资源及其可提供的氮磷钾养分数量,分析和评价了中国不同区域养分输入、输出特点及平衡状况。针对目前养分输入、输出存在的一系列问题,提出了养分资源管理的对策和建议。
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
为了更好地了解温度和湿度对土壤氮矿化过程的影响,从而估计森林生态系统土壤有机氮的矿化速率,在中国7种典型森林生态系统中用PVC管采集森林土壤样品,通过在实验室控制土壤的温度与湿度,将不同湿度的土柱置于不同温度的生化培养箱中培养30 d,分析培养前后的NH+4-N和NO-3-N含量,确定土壤的净矿化速率和净硝化速率。结果表明,温度和湿度与土壤的矿化过程存在比较明显的相关关系(p<0.001)。同时建立了三元的方程来描述温度(t)和湿度(wfps)对土壤氮矿化速率(Rmin)的影响,Rmin=e-7.60+0.07×t+14.74×wfps-10.41×wfps2。利用这个实验模型估算了全国森林生态系统的年氮矿化量,估算结果与野外实测数据基本吻合。
Magsci [本文引用: 1]
为了更好地了解温度和湿度对土壤氮矿化过程的影响,从而估计森林生态系统土壤有机氮的矿化速率,在中国7种典型森林生态系统中用PVC管采集森林土壤样品,通过在实验室控制土壤的温度与湿度,将不同湿度的土柱置于不同温度的生化培养箱中培养30 d,分析培养前后的NH+4-N和NO-3-N含量,确定土壤的净矿化速率和净硝化速率。结果表明,温度和湿度与土壤的矿化过程存在比较明显的相关关系(p<0.001)。同时建立了三元的方程来描述温度(t)和湿度(wfps)对土壤氮矿化速率(Rmin)的影响,Rmin=e-7.60+0.07×t+14.74×wfps-10.41×wfps2。利用这个实验模型估算了全国森林生态系统的年氮矿化量,估算结果与野外实测数据基本吻合。
[本文引用: 1]
[本文引用: 1]
[R/OL]. (
URL [本文引用: 1]
[R/OL]. (
URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}