 ,1, ���,2
,1, ���,2Phenotypic Variation and Genome-wide Association Analysis of Root Architecture at Maize Flowering Stage
ZHANG XiaoQiong1, GUO Jian2, DAI ShuTao3, REN Yuan4, LI FengYan5, LIU JingBao3, LI YongXiang2, ZHANG DengFeng2, SHI YunSu2, SONG YanChun2, LI Yu2, WANG TianYu2, ZOU HuaWen,1, LI ChunHui,2ͨѶ����:
���α༭: ����
�ո�����:2019-03-20��������:2019-04-25�����������:2019-07-16
| ��������: |
Received:2019-03-20Accepted:2019-04-25Online:2019-07-16
����� About authors
����,E-mail: qiong2017@foxmail.com��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (5010KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
��С��, ����, ������, ��Ԫ, �����, ������, ������, �ŵǷ�, ʯ����, ���ഺ, ��ԣ, ������, ����, ���. �����ڸ�ϵ�ṹ�ı��ͱ�����ȫ�������������[J]. �й�ũҵ��ѧ, 2019, 52(14): 2391-2405 doi:10.3864/j.issn.0578-1752.2019.14.002
ZHANG XiaoQiong, GUO Jian, DAI ShuTao, REN Yuan, LI FengYan, LIU JingBao, LI YongXiang, ZHANG DengFeng, SHI YunSu, SONG YanChun, LI Yu, WANG TianYu, ZOU HuaWen, LI ChunHui.
0 ����
���о����塿��������������Ҫ����ʳ��������������ϵ��Ϊֲ��������ֱ�ӽӴ�������,��ˮ�ֺ����ֵ�����������Ҫ[1,2,3,4]�����,��������ϵ�ṹ�γɵ��Ŵ�����,�ھ�����ϵ�����״�������λ����,�Խ�һ��������ײ���������Ҫ���塣��ǰ���о���չ�������о�����,��ϵ�ij��Ⱥ�������С���������������,���ϵ���ֱ�Ϊ0.39��0.73[5,6];�ɺ�������,��ϵ��ˮƽ����ĽǶ���С�����������������,���ϵ����0.91[7],��ֱ�����������Լ���ϵ�������зֲ��ı仯������Ŀ������������[8,9];����ͨ�����ٵ��½ڸ����������Ӹ�ϵ������������ˮ��������,�������ٸɺ�в����ɵ��˺�[10]���о�����,����ϵ���ͱ���ḻ,��ϵ�����������ȡ�ֱ���ͽǶȹ�ͬ���������γ�һ�����ӡ���̬����ά��ϵ�ṹ[11],�����Խ�ϵ���ڵĸ����������ȡ�ֱ�����صı���ϵ���ֱ�Ϊ0.43��25.2��0.48��0.25��0.30��0.36[12,13]������ϵ�ṹ��Ϊ������ϵͳ���ߺ��ϵͳ,������ϵͳ��Ҫ���������Ӹ���1������������,���������ڷ�����Ҫ����,������ֲ�����������˻�,��ͬ����Ʒ�ֵ����Ӹ�����0��13������;�ߺ��ϵͳ�����ڸ��Ͳ��,�ڸ��Ӿ����ϳ���,�ӵ��¾��ڳ����ij�Ϊ���½ڸ�,�ӵ��Ͼ��ڳ����Ľڸ��ֳ�֧�ָ���������,�ߺ����Ҫ�̶�֧�ֺ�����ˮ�֡����ֵ�����,����Լ��6����½ڸ���2��3����Ͻڸ�,Լ70���ڸ�[14]����������ֲ�������������,��ϵ�������������ཬ������ﵽ���[15],���,����о���ѡ���ڿ����ڻ�����ڶ�����ϵ���ߺ��ϵͳ�����о�[5,16-17]��ǰ���о��������,��ͬ���������Ʒ���ڽڸ����������ȡ��Ƕȡ����ص���״�ϴ����������IJ���[5,7,18-19]�������о�֤ʵ,����������Ʒ�ָ��������,��ϵ���͵ı仯�Բ�������ֱ��Ӱ��[4]��Ŀǰ,�����������������Ѿ���λ��һЩ���ڸ������״��QTL��CAI��[5]����Ye478��Wu312������BC4F3Ⱥ��,�ֱ��ڰν��ڡ���˿�ڡ�������3��ʱ�ڶԵ��½ڸ������͵��½ڸ����Ƚ��ж�λ,����3������½ڸ�������ص�QTLλ��,8������½ڸ������йص�QTL,������2��QTL������2�������ڹ���λ���̺���[20]������һ��BC4F3Ⱥ������ںཬ�ڵĸ����ء��ܸ���������������½ڸ��������½ڸ����ȸ�ϵ��״,QTL�������������ڸ�ϵ�ṹ20��QTL�ཬ�ڸ�ϵ�ṹ14��QTL��ZHANG��[17]���ô�ۻ�ݺ�W22�����İ���866�ݲ��ϵ�BC2S3Ⱥ��,�ڿ����ڷֱ�Ժ��Ϻͱ�����9���ڸ��������״����QTL����,����λ��133��QTL������69��������64����,����62��QTL�뿪����QTL��ص�,15��QTL�����QTL��ص�,����5�����ƽڸ����������ں���ߵ�һ���ЧQTL��KU��[21]�Ի����ĺ�CML288������RILȺ�������F2Ⱥ��Ϊ����,���������ܲ�������Ч����������������QTL��λ,��RILȺ���й���10��QTL,������F2Ⱥ���й���λ��15��QTL��GU��[22]��F2:3Ⱥ����Ϊ�������,��������ֱ�������������ء����ص���״����QTL����,����93��QTL��GUO��[23]�ռ��������ѷ�����428������ϵ�����״QTL,����Ԫ��������,���53��Meta-QTL��ȫ���������������genome-wide association studies,GWAS�����о�������״����Ҫ����֮һ,���Ų������Ŀ��ٷ�չ,��ϸ��ܶȷ��ӱ��,����GWAS��������ֲ����Ҫ��״���Ŵ����Ƶ�Ӧ��Խ��Խ�ࡣPACE��[13]����Ames panel�е�384���Խ�ϵ,������22�����ڸ�ϵ�����״,���GBS�����õĻ�������������GWAS����������268�����ϵ��״������صı�ǡ�SANCHEZ��[24]������DHȺ������ڸ�ϵ�����״����GWAS����,����17����������SNP��ZAIDI��[16]����396���ȴ������Խ�ϵ��չ��ˮ��в�������³����ڸ�ϵ��״��GWAS����,����67�����������ء����������������������������ܶȵ���״����������SNPλ�㡣ZHANG��[17]����663�������Խ�ϵ�Խڸ�������GWAS����,��ϴ�ۻ�ݺ�W22������BC2S3Ⱥ�������������,����3����������SNP��ڸ����Ϳ�������ص�QTL����λ�������о�����㡿����ǰ�����÷���Ⱥ�����Ⱥ���һЩ����ϵ��״��ص�QTL,������ȫ������������������ھ��ڸ�ϵ�ṹ�Ŵ�������о�����,���о���ͬ����ʱ�ںͲ�ͬ����������Ⱥ���ϵĸ�ϵ�ṹ�仯���١��������Ĺؼ����⡿���о���111�������Խ�ϵΪ�������,��6�����ڵ��½ڸ������״���вⶨ,�Բ�ͬ��������Խ�ϵ��ϵ�ı仯����ͬ����������Ⱥ�����Խ�ϵ��ϵ�IJ�����з���,���������½ڸ������״����ȫ�������������,ּ���˽����ײ�ͬ����ʱ�ڸ�ϵ�ṹ�ı仯���ɺ�̽�������½ڸ������״���Ŵ����ơ�1 �����뷽��
1.1 ������ϼ��������
��2017���ڱ������������١�ɽ�����弰����ԭ��4����������������顣���Բ�����111�������Խ�ϵ��ɡ������Ʋ�����ȫ����������,2���ظ�,ÿ������˫������ֲ,�г�3 m,�о�0.6 m,���0.25 m,�ܶ�67 500��/hm2,�����ó��������1.2 ��״�ⶨ�뷽��
������ʱ,ѡȡ��С���ڳ���һ�¡�����һ�µ�3������,ͨ���˹��ڸ������ϵ��״��ÿ�������Գ�25 cm����25 cm�����Ϊ30 cm�ĸ�ϵ��Ҫ�ֲ�������Ϊȡ����λ,�ռ�������ϵ,ʹ�ø�ѹˮǹ����ϵ��ϴ�ɾ����ռ��ĸ�ϵ��4����λ����ͼƬ�ɼ�,����RESTroot��ϵ���������Ե��½ڸ��Ƕȣ�belowground node root angle,RA�������½ڸ������belowground node root shadow area,RS�����з���ȡƽ��ֵ,���½ڸ��Ƕ�Ϊ���½ڸ��ſ��ļн�,���½ڸ����Ϊ���½ڸ�����ͶӰ��������ͼƬ�ɼ��ĸ�ϵ��������ϵ�����IJ��ʹ�ù�������һ����,��¼���½ڸ�������belowground node root layer number,RLN���͵��½ڸ���������total number of belowground node root,TRN��,������ˮ���������½ڸ������belowground node root volume,RV��������ָ��ⶨ��ɺ�,����ϵװ��ţƤֽ����,105��ɱ��30 min,85����������,�ⶨ���½ڸ����أ�belowground node root dry weight,RDW����1.3 �������ݵ�ͳ�Ʒ���
����Excel2016��SPSS16.0�����Ե��½ڸ�������RLN�������½ڸ���������TRN�������½ڸ��Ƕȣ�RA�������½ڸ������RS�������½ڸ������RV���͵��½ڸ����أ�RDW����6����״����ͳ�Ʒ���,�ֱ�ȡ4�������¸���״��ƽ��ֵ,���������״�ı���ͱ���ϵ��������QTL IciMapping V4.1�����е�ANOVA�����Ŵ���,����SPSS16.0���������ݽ��е����ط������,����Bonferroni�������ж��رȽ�,ʹ��Minitab 16���Ƹ�ϵ��״��Ƶ���ֲ�ͼ,����R���Խ��и�ϵ��״�������Է�����1.4 ȫ����������������ѡ������ھ�
ʵ����ǰ�ڶ�111���Խ�ϵ�����˵���ȵ��ز���,�����152 352��������SNP��ǣ�ȱʧ��С��10%,Ч��λ����Ƶ�ʣ�minor allele frequency,MAF������0.05,λ���Ӻ϶�С��10%����������Щ�������ı��,����TASSEL5.0������111���Խ�ϵ����Ⱥ��ṹ����,��ϲ���ϵ��,����iTOL��������Neighbor-joining tree������Fixed and random model Circuitous Probability Unification��FarmCPU��ģ��[25]��4�������µ�6���ڸ������״��ƽ��ֵ����ȫ�������������,��P=1.0��10-5Ϊ��ֵ,�ж�SNP�����Ŀ����״�����������ԡ�ʹ��TASSEL5.0��������Ⱥ���������ƽ�⣨linkage disequilibrium,LD��ƽ��˥������,��������������SNP���������B73����������RefGen_v4�ϵ�����λ��,����LD��Χ���ھ������½ڸ������״�ĺ�ѡ����,��NCBI��https://blast.ncbi.nlm.nih. gov����MaizeGDB��https://www.maizegdb.org����TAIR��https://www.arabidopsis.org����վ�϶Ժ�ѡ������й���ע��,ʹ��agriGO���߷���������http://bioinfo. cau.edu.cn/agriGO/���Ժ�ѡ����Ĺ��ܽ��и���������
2 ���
2.1 ���½ڸ������״���ͷ���
ȡ4�������µĸ�ϵ�����״��ƽ��ֵ��Ϊ��������,��6�����½ڸ������״����ͳ�Ʒ���,111�������Խ�ϵ��6��������״RLN��TRN��RA��RS��RV��RDW�����ֽ�����̬�ֲ���ͼ1��,������Щ��״��Ϊ���͵�������״,��Ч�������ơ���111�������Խ�ϵ��,6��������״�ı��ͱ��ֳ��ϴ�ı��죨��1��������ϵ����������,RV��RDW�ı���ϵ�����,��RLN��TRN��RA��RS�ı�ֵ������1.99��4.00��,��6����״����ʾ�ϸߵ��Ŵ�����ͼ1
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1�����½ڸ������״����
A:����ϵ���Ͷ�����;B:���½ڸ������״Ƶ���ֲ�ͼ;RLN:���½ڸ�����;TRN:���½ڸ�������;RA:���½ڸ��Ƕȣ��㣩;RS:���½ڸ������cm2��;RV:���½ڸ������cm3��;RDW:���½ڸ����أ�g������ͬ
Fig. 1Phenotypic variation of belowground node root-related traits of the maize
A: Phenotypic diversity of maize root; B: Frequency distribution of belowground node root-related traits; RLN: Belowground node root layer number; TRN: Total number of belowground node root; RA: Belowground node root angle (��); RS: Belowground node root shadow area (cm2); RV: Belowground node root volume (cm3); RDW: Belowground node root dry weight (g). The same as below
Table 1
��1
��1���½ڸ������״ͳ�Ʒ���
Table 1
| ��״ Trait | ��ֵ Mean | ���ֵ Max | ��Сֵ Min | ���� SD | ����ϵ�� CV (%) | �����Ŵ��� H2 |
|---|---|---|---|---|---|---|
| ���½ڸ�����RLN | 6.67 | 8.67 | 5.38 | 0.62 | 9.29 | 0.67 |
| ���½ڸ�������TRN | 59.41 | 88.00 | 42.00 | 8.96 | 15.08 | 0.74 |
| ���½ڸ��Ƕ�RA | 58.67 | 80.70 | 35.82 | 8.51 | 14.50 | 0.80 |
| ���½ڸ����RS | 76.28 | 111.91 | 45.55 | 13.85 | 18.15 | 0.79 |
| ���½ڸ����RV | 68.75 | 144.17 | 19.63 | 24.84 | 36.13 | 0.75 |
| ���½ڸ�����RDW | 13.77 | 32.60 | 4.09 | 5.11 | 37.12 | 0.82 |
�´��ڴ�|����CSV
2.2 ���½ڸ������״������Է���
ͨ����4�������µ�6����ϵ��״��������Է�����ͼ2��,���½ڸ���������½ڸ����������Ƕȡ�����������������������ع�ϵ������,���½ڸ��������½ڸ����ص����ϵ���ﵽ0.88,����ԴK����ˮƽ;���½ڸ��Ƕ�����½ڸ���������ϵ��Ϊ0.72,Ҳ�ﵽ��������ˮƽ;���½ڸ��Ƕȡ��������½ڸ����ص����ϵ��Ϊ0.42��0.68,���,��ϵ��ռ�ռ�Խ��,��ϵ������Խ��,��ϵ�Ƕ�Խ��,��ϵ��ͶӰ�����Խ���½ڸ���������½ڸ������������������,���½ڸ�������������������½ڸ��ǶȺ�����ʸ����,˵�����½ڸ�����������Խ��,��ϵ�ṹ����Խ����,��ϵ�Ƕ�ԽС,��ϵ�����С��ͼ2
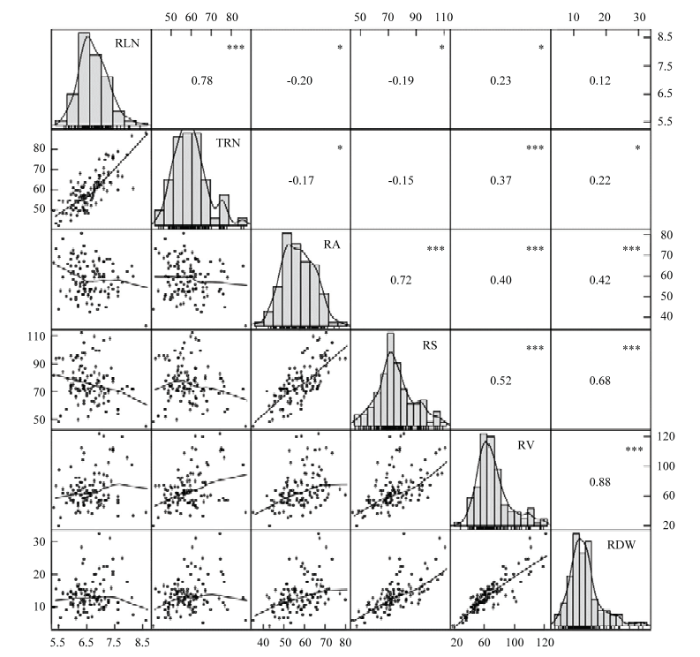 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2���½ڸ������״������Է���
*:��0.05ˮƽ���������;***:��0.001ˮƽ���������
Fig. 2Correlation analysis of belowground nodal root-related traits
*: Correlation is significant at the 0.05 level; ***: Correlation is significant at the 0.001 level
2.3 ��ͬ��������Խ�ϵ���½ڸ������״�ı��ͷ���
ͨ���������Թ��Բ��ϵ�ѡ��ʱ����б�ע����2��,111���Խ�ϵ����46���Խ�ϵ�ɸ�����ѡ��ʱ�仮�ֳ�4�����,�ֱ�Ϊ1970s��10�ݣ���1980s��17�ݣ���1990s��14�ݣ���2000s��5�ݣ�����������ĸ���,��ϵ����״�����ֳ���ͬ�̶ȵı仯,�����غ�����ڸ������IJ���δ�ﵽ����ˮƽ;���½ڸ����������������ǶȺ������2000sǰ�����������������,2000s�Խ�ϵ���½ڸ���������������������,���½ڸ��ǶȺ������������;���½ڸ�������������������ĸ������ֳ��½�������,���½ڸ��ǶȺ����������ĸ������ֳ����������ƣ�ͼ3����Table 2
��2
��2��ͬ��������Խ�ϵ
Table 2
| ��� Era | �������� Number of materials | �������� Name of materials |
|---|---|---|
| 1970s | 10 | C103��E28��H21��Mo17������02 Danhuang02��������Huangzaosi������322 Weifeng322��Ҵ8112 Ye8112��ԭ��02 Yuanwu02����63 Ji63 |
| 1980s | 17 | 853��5003��81162��4F1��B73��K12��Mo17Ht��X178����340 Dan340����344 He344����319 Qi319������4 Tianya4����7922 Tie7922��Ҵ478 Ye478��Ҵ488 Ye488��Ҵ52106 Ye52106��֣58 Zheng58 |
| 1990s | 14 | 444��5237��A801��C8605-2��Ki3��KL4��P138����7-2 Chang7-2����598 Dan598����229 Duo229����Ұ��3 Huangyesi3�����ƻ�17 Huotanghuang17����137 Shen137��֣22 Zheng22 |
| 2000s | 5 | ��3336 Shen3336����-144 Si-144��PH6JM����724 Jing724����725 Jing725 |
�´��ڴ�|����CSV
ͼ3
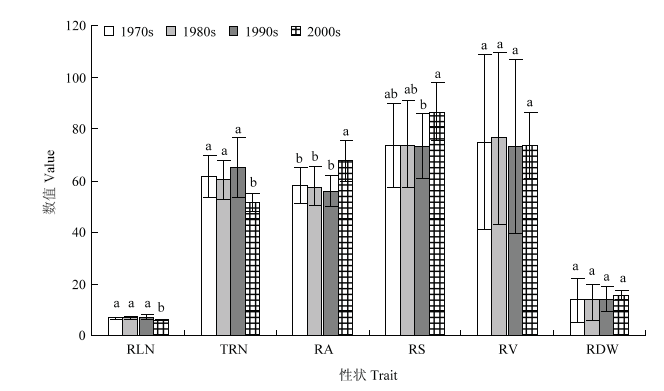 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3��ͬ����Խ�ϵ���½ڸ������״�IJ������
a��b:��0.05ˮƽ�ϲ�����������ͬ
Fig. 3Differences of belowground nodal root-related traits of inbred lines developed in different eras
a, b: Significant at the 0.05 level respectively. The same as below
2.4 ��ͬ��Ⱥ�����Խ�ϵ���½ڸ������״�ı��ͷ���
����111���Խ�ϵ������,����TASSEL5.0����������о��������ͼ4��,�������־ի��[26]��820�������Խ�ϵ��Ⱥ��ṹ�ķ������,111�ݲ����е�93��84%���ݲ��Ͽɻ���Ϊ���Ⱥ��Reid�����ô���Ⱥ��Lüda Red Cob,LRC��������˹��Ⱥ��Lancaster��������ƽͷȺ��Tang Si Ping Tou,TSPT����PȺ��P group��,����18�ݣ�16%�������γ���һ�����Ⱥ��mixed group������ͬ��Ⱥ��ϵ�����״��ͳ�Ʒ�������,��ͬ��Ⱥ��ĵ��½ڸ����������������ǶȺ����������Ƚ�С,���½ڸ����غ͵��½ڸ������5����Ⱥ�еı�����Ƚϴ���3������ͬ��Ⱥ��ϵ��״�����������,�ô�����Ⱥ��6�����½ڸ���״������������Ⱥ,��δ�ﵽ����ˮƽ;����˹����Ⱥ��6�����½ڸ���״ֵ�������ô�����Ⱥ;PȺ��6�����½ڸ���״ֵ�����ô�����Ⱥ����������;����ƽͷ��Ⱥ�ĵ��½ڸ��Ƕȡ������������������Ⱥ,���½ڸ���������������������ô�����Ⱥ����������;�����Ⱥ�ĵ��½ڸ����������������ô�����Ⱥ,����5�����½ڸ���״ֵ���ô�����Ⱥ���������죨ͼ5����ͼ4
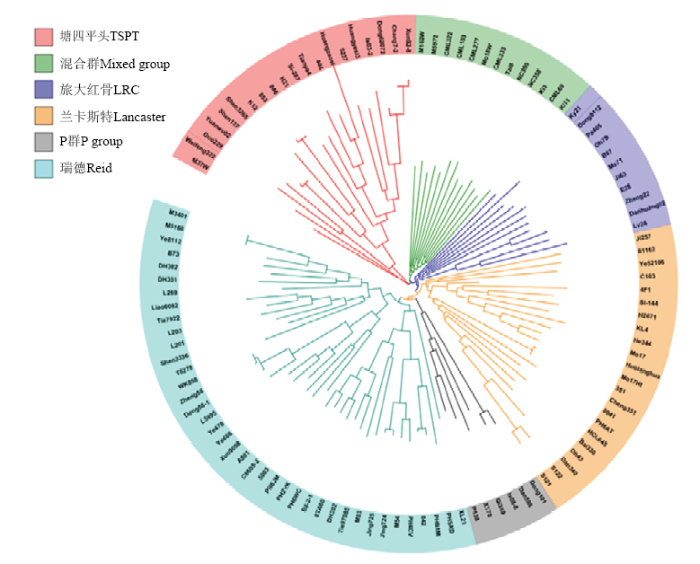 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4111�������Խ�ϵ�������
Fig. 4Cluster analysis of 111 maize inbred lines used in this study
Table 3
��3
��3��ͬ��Ⱥ���½ڸ������״��ͳ�Ʒ���
Table 3
| ��״/��Ⱥ Trait/Group | ��ֵ Mean | ���� SD | ��Сֵ Min | ����ֵ Max | ����ϵ�� CV (%) |
|---|---|---|---|---|---|
| RLN | |||||
| ���Reid | 6.47ab | 0.64 | 5.38 | 8.22 | 9.94 |
| ��LRC | 6.86a | 0.56 | 6.17 | 8.00 | 8.13 |
| ����˹��Lancaster | 6.44b | 0.43 | 5.56 | 7.33 | 6.73 |
| ����ƽͷTSPT | 7.02a | 0.64 | 6.17 | 8.17 | 9.12 |
| PȺP group | 6.54ab | 0.52 | 5.83 | 7.21 | 7.92 |
| ���ȺMixed group | 7.08a | 0.55 | 6.33 | 8.67 | 7.76 |
| TRN | |||||
| ���Reid | 56.86b | 7.33 | 43.13 | 75.83 | 12.90 |
| ��LRC | 63.72a | 10.53 | 49.39 | 77.33 | 16.52 |
| ����˹��Lancaster | 55.83b | 6.51 | 42.00 | 68.67 | 11.66 |
| ����ƽͷTSPT | 63.79a | 10.47 | 48.11 | 86.67 | 16.41 |
| PȺP group | 60.40ab | 9.26 | 47.92 | 74.80 | 15.33 |
| ���ȺMixed group | 63.35a | 9.77 | 51.67 | 88.00 | 15.42 |
| RA | |||||
| ���Reid | 61.32a | 7.13 | 48.65 | 80.70 | 11.62 |
| ��LRC | 62.66a | 4.81 | 53.71 | 67.65 | 7.67 |
| ����˹��Lancaster | 58.43ab | 9.82 | 43.89 | 78.03 | 16.81 |
| ����ƽͷTSPT | 54.82b | 6.94 | 44.87 | 66.00 | 12.66 |
| PȺP group | 57.52ab | 8.23 | 43.71 | 67.99 | 14.31 |
| ���ȺMixed group | 55.76ab | 9.94 | 35.82 | 71.87 | 17.82 |
| RS | |||||
| ���Reid | 82.07a | 11.62 | 60.58 | 109.61 | 14.16 |
| ��LRC | 82.68a | 12.71 | 68.47 | 106.65 | 15.37 |
| ����˹��Lancaster | 74.90a | 14.99 | 48.38 | 111.91 | 20.02 |
| ����ƽͷTSPT | 65.50b | 10.22 | 51.80 | 88.01 | 15.60 |
| PȺP group | 73.68ab | 13.88 | 57.50 | 97.73 | 18.84 |
| ���ȺMixed group | 74.34a | 13.85 | 45.55 | 107.94 | 18.63 |
| RV | |||||
| ���Reid | 69.72ab | 23.36 | 34.21 | 140.83 | 33.51 |
| ��LRC | 81.82a | 33.68 | 38.31 | 144.17 | 41.17 |
| ����˹��Lancaster | 66.95ab | 23.85 | 19.63 | 124.52 | 35.63 |
| ����ƽͷTSPT | 71.59ab | 29.86 | 33.33 | 139.17 | 41.71 |
| PȺP group | 68.97ab | 24.45 | 44.37 | 107.99 | 35.45 |
| ���ȺMixed group | 60.62b | 19.36 | 38.33 | 123.11 | 31.93 |
| RDW | |||||
| ���Reid | 14.25a | 4.34 | 6.49 | 24.86 | 30.44 |
| ��LRC | 16.21a | 7.84 | 7.55 | 32.60 | 48.38 |
| ����˹��Lancaster | 13.07a | 4.92 | 4.09 | 28.32 | 37.68 |
| ����ƽͷTSPT | 12.26a | 4.99 | 5.21 | 25.07 | 40.66 |
| PȺP group | 14.22a | 4.60 | 8.99 | 23.49 | 32.35 |
| ���ȺMixed group | 13.74a | 5.55 | 5.86 | 31.29 | 40.41 |
�´��ڴ�|����CSV
ͼ5
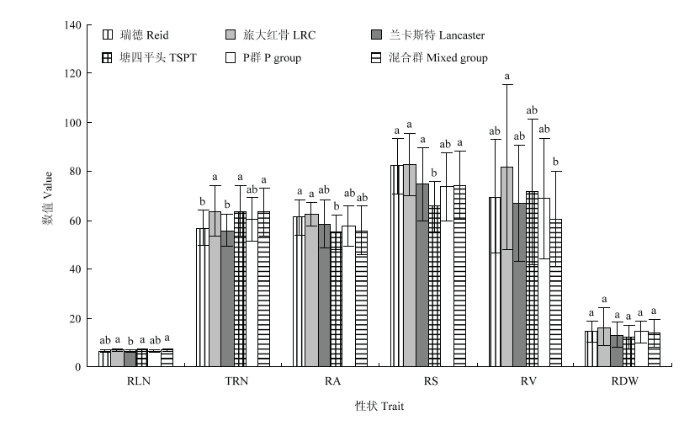 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5��ͬ��Ⱥ�Խ�ϵ���½ڸ������״�IJ������
Fig. 5Differences of belowground nodal root-related traits in inbred lines of different heterotic groups
2.5 ���½ڸ������״ȫ�������������
����FarmCPUģ�Ͷ�4�������µ�111�������Խ�ϵ���½ڸ������״��ƽ��ֵ����ȫ�������������,��P<0.00001ˮƽ��,�ж�SNP������о���״������������,����26������������SNPλ�㣨ͼ6����4��������,��6�����½ڸ�����SNPλ��,�ֱ�λ�ڵ�1��6��8Ⱦɫ����;4�����½ڸ�����SNPλ��,�ֱ�λ�ڵ�4��6Ⱦɫ����;10�����½ڸ����SNPλ��,�ֱ�λ�ڵ�3��4��6��8��9��10Ⱦɫ����;6�����½ڸ�����SNPλ��,�ֱ�λ�ڵ�3��4��6��7Ⱦɫ���ϡ�ͼ6
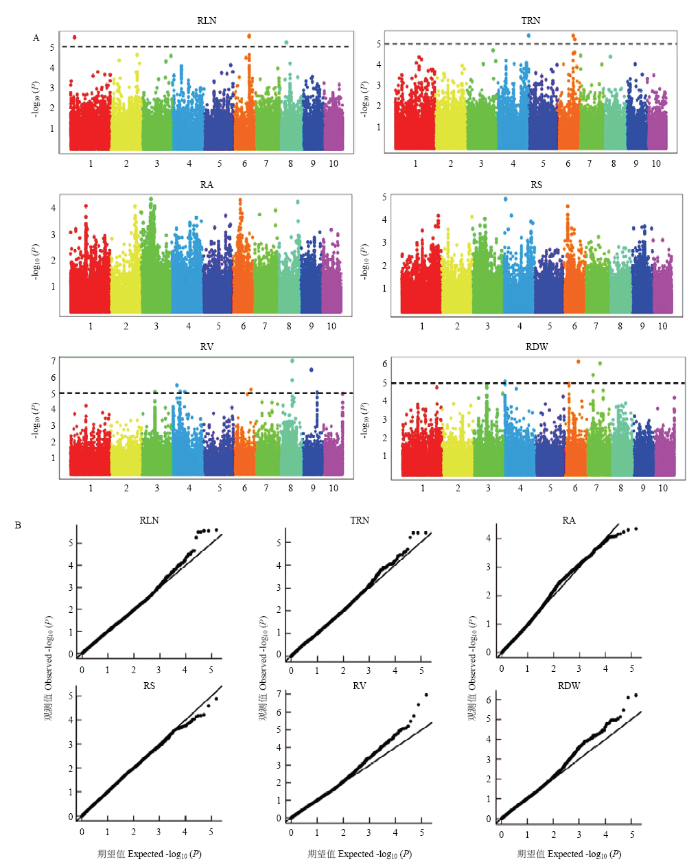 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6ȫ�������������������ͼ��A����QQͼ��B��
Fig. 6(A) Manhattan plot and (B) Quantile-quantile plot for genome-wide association study of maize root traits
Table 4
��4
��4Ӱ�������½ڸ������״��SNP������ܵĺ�ѡ�����书��ע��
Table 4
| ��״ Trait | SNP | Chr. | λ��a Position (bp) | Pֵ P value | ��ѡ����b Candidate gene | ������ע�� Gene annotation | �ο�����c Reference |
|---|---|---|---|---|---|---|---|
| RLN | Chr.1.S_31312069 | 1 | 31312069 | 3.07E-06 | Zm00001d028337 | ���������������������øPERK4 Proline-rich receptor-like protein kinase PERK4 | |
| RLN | Chr.1.S_31312087 | 1 | 31312087 | 3.07E-06 | Zm00001d028344 | �ƶ��Ľ������ܵ���C3 Putative metal tolerance protein C3 | |
| RDW | Chr.3.S_103120493 | 3 | 103120493 | 1.07E-05 | Zm00001d041183 | ����ø�����嵰����/�հ��ᵰ��øprpf4B-liken Protein kinase superfamily proteiserine/threonine-protein kinase prpf4B-liken | |
| RV | Chr.3.S_104251070 | 3 | 104251070 | 8.33E-06 | Zm00001d041192 | ����ת�˵���4 Sucrose transporter4 | |
| RDW | Chr.4.S_9975093 | 4 | 9975093 | 7.24E-06 | Zm00001d048943 | ���������׳����嵰�� Thioredoxin superfamily protein | [14] |
| RV | Chr.4.S_38214896 | 4 | 38214896 | 3.38E-06 | Zm00001d049623 | SNARE��ص��� SNARE-associated protein-related | |
| Zm00001d049627 | �����ᰱ��ø Methionine aminopeptidase | ||||||
| RV | Chr.4.S_63797065 | 4 | 63797065 | 7.79E-06 | Zm00001d050069 | ������-6-����ϳ�ø8 Trehalose-6-phosphate synthase8 | |
| RV | Chr.4.S_100804613 | 4 | 100804613 | 8.30E-06 | Zm00001d050558 | OSJNBa0043A12.20���� OSJNBa0043A12.20 protein | |
| TRN | Chr.4.S_246664102 | 4 | 246664102 | 3.96E-06 | Zm00001d054104 | E3���ص�������øUPL3 E3 ubiquitin-protein ligase UPL3 | |
| Zm00001d054105 | ���ռ�ø������3 Uridine kinase-like protein 3 | ||||||
| RDW | Chr.6.S_34950886 | 6 | 34950886 | 9.19E-06 | Zm00001d035587 | G��������S������˿����/�հ��ᵰ��øAt2g19130 G-type lectin S-receptor-like serine/threonine-protein kinase At2g19130 | [5,20] |
| Zm00001d035588 | G��������S������˿����/�հ��ᵰ��øAt2g19130 G-type lectin S-receptor-like serine/threonine-protein kinase At2g19130 | ||||||
| RDW | Chr.6.S_108781517 | 6 | 108781517 | 6.09E-07 | Zm00001d037004 | Ras��ص���RABA1d Ras-related protein RABA1d | [17] |
| Zm00001d037010 | ë����ص���øiota Shaggy-related protein kinase iota | ||||||
| RLN/TRN | Chr.6.S_123079904 | 6 | 123079904 | 4.00E-06 | Zm00001d037360 | �ƶ���bZIPת¼���ӳ����嵰�� Putative bZIP transcription factor superfamily protein | [20] |
| RLN/TRN | Chr.6.S_123079927 | 6 | 123079927 | 4.00E-06 | Zm00001d037368 | δ֪ Unknown | [20] |
| RLN | Chr.6.S_123845438 | 6 | 123845438 | 2.51E-06 | Zm00001d037384 | ������3-O-���ǻ�ת��ø Anthocyanidin 3-O-glucosyltransferase | [20] |
| TRN | Chr.6.S_135051799 | 6 | 135051799 | 6.13E-06 | Zm00001d037712 | MAR���˿״����1 MAR-binding filament-like protein 1 | |
| RV | Chr.6.S_136591446 | 6 | 136591446 | 6.19E-06 | Zm00001d037751 | ������̥�����ḻ��LEA�������Ǹ�������ǵ����� Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family | |
| RDW | Chr.7.S_48142656 | 7 | 48142656 | 3.42E-06 | Zm00001d019648 | �����ϵ���1 Nucleic acid binding protein 1 | [14] |
| RDW | Chr.7.S_101804639 | 7 | 101804639 | 7.71E-07 | Zm00001d020242 | пָCCCH�ṹ��ĵ�����64 Zinc finger CCCH domain-containing protein 64 | [14] |
| RLN | Chr.8.S_55681954 | 8 | 55681954 | 5.47E-06 | Zm00001d009324 | AMSH�����ص�������ø3 AMSH-like ubiquitin thioesterase 3 | |
| RV | Chr.8.S_100196991 | 8 | 100196991 | 1.68E-06 | Zm00001d010119 | ���ݿ˵���CS66 Cold shock protein CS66 | [20] |
| RV | Chr.8.S_100680141 | 8 | 100680141 | 1.07E-07 | Zm00001d010128 | ���Ժ����嵰��P2b��rpp2b�� Acidic ribosomal protein P2b (rpp2b) | [20] |
| RV | Chr.9.S_67485696 | 9 | 67485696 | 3.82E-07 | Zm00001d046152 | �ƶ���RING-H2ָ����ATL71 Putative RING-H2 finger protein ATL71 | |
| RV | Chr.9.S_111860806 | 9 | 111860806 | 9.44E-06 | Zm00001d046942 | ����-1,6-������ø Fructose-1,6-bisphosphatase | |
| RV | Chr.10.S_150110617 | 10 | 150110617 | 1.08E-05 | Zm00001d026687 | ��������Ӧ����17 Auxin response factor 17 | [14] |
| Zm00001d026694 | ��������Ӧ����13 Auxin response factor 13 |
�´��ڴ�|����CSV
2.6 ���½ڸ������״��ѡ�������
��Ⱥ���LD���������ʾ,r2˥����0.2ʱȺ���������ƽ��˥������Ϊ250 kb��ͼ7�����Թ��������õ�������������SNPλ�����������,����MaizeGDB��վ����������λ���LD��Χ�ڲ��������½ڸ������״�ĺ�ѡ����,���Ժ�ѡ�������ͳ�ơ�26�����ƽڸ���״�Ļ����������ڹ�������177����ѡ����,����,���й���ע�͵�135����ѡ����Ĺ��ܸ�����������,40����ѡ������Ҫ����ֲ�����ڵĴ�л����,13����ѡ������ֲ��Ӧ����Ӧ���漰������;�����,57����ѡ����Ĺ���δ���������,�����ѡ�������漰������ԡ������ԡ���ϵ���ϸ���ɷֵȣ�ͼ8��������ÿ����������SNP��LD��Χ�����к�ѡ����Ĺ���ע��,ѡ����Ŀ����״��صĺ�ѡ������ע��������4��ͼ7
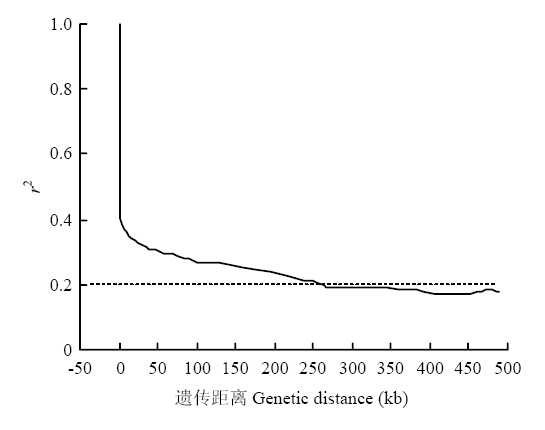 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ7Ⱥ��������ƽ��˥�����루LD��
Fig. 7Linkage disequilibrium attenuation distance of the population
ͼ8
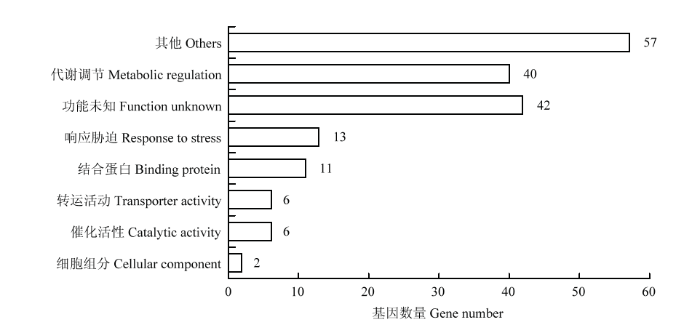 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ8��ѡ��������ѧ;��ע��ֱ��ͼ
Fig. 8Histogram of annotations for biological pathways of candidate genes
3 ����
3.1 ��ͬ��������Խ�ϵ���½ڸ������״�ı��ͷ���
����������Դ������Ʒ���Ŵ������������������õĻ���,���й����ײ�ҵ��չ�IJ�ͬ��,�����Խ�ϵ��ѡ��Ŀ�겻�ϸı䡣���Ų�ͬ����ʱ�������ϲ���״�ĸ���,���²���ϵ�ṹ���ܱ���ӵ�ѡ��,���ܵ��²�ͬ����������Խ�ϵ��ϵ��״������Ų��졣���о�ͨ���Բ�ͬ��������Խ�ϵ���½ڸ���״���з���,�������,�������ֽ��̵��ƽ�,��ͬ������������ʵ��¸�ϵ�ṹ���ֳ��˲�ͬ�̶ȵIJ���,���½ڸ����������������½�������,���½ڸ��ǶȺ����������������,�����غ�������������졣ǰ���о�����,��1980s��2000s����Ʒ�ֵĸ���,����ϵ��״�еĸ������������������ؾ���С,ֲ��ͨ�����ٸ�ϵ������Ӷ�����Ӫ�����ʵ�����[27,28]������ֲ��Ե��ص���������Ӱ���ϵ�Ĺ��ͼ��ֲ�[29],20����90������ŵ���ʩ����������,��ϵ�Ե��ʵ�����������,ֲ���ϵͨ�����ʹ�л���ĵijɱ������ֲ��������������[30],���ո�ϵ����Ϊ�������ࡣ�뱾�о��õ��ĵ��½ڸ������������������������ٵĽ��һ�¡�
3.2 ��ͬ��Ⱥ�����Խ�ϵ���½ڸ������״�ı��ͷ���
�й�����������Ⱥ��Ҫ����Ϊ����ƽͷȺ���ô���Ⱥ�����Ⱥ������˹��Ⱥ��PȺ��[31]�������о�����,��ͬ��Ⱥ�İ�Ҷ�����״�������ཬ����ˮ�����״�ȴ�����������[32,33]��Ȼ��,�Բ�ͬ��Ⱥ�������Խ�ϵ��ϵ�ṹ������о����б��������о����ֲ�ͬ��Ⱥ�ĵ��½ڸ������״���ڲ���,�ô�����Ⱥ���½ڸ��������������Ҹ�ϵ��������,���ڡ����ϵ��;�����Ⱥ���½ڸ���������;����˹����Ⱥ���½ڸ�������,�Ƕȡ����������������С;����ƽͷ��Ⱥ���½ڸ��ǶȺ������С,�ֲ�����;PȺ���½ڸ���������������صȾ������е�ˮƽ����ͬ��Ⱥ���½ڸ���״�IJ���,��Ϊ��ϵ�ṹ�Ŵ������ṩ�ο���Ϣ�����о����������ڵ��½ڸ���6����״֮���������������ػ��߸����,˵������״�ķ����������Эͬ�ٽ����á����,������ϵ���о���Ӧ�����ڼı��ͷ���,�ڽ����о���Ӧ������ϵ�ṹ���Ŵ�����������γ�֮��Ĺ�ϵ������Ҫλ��,��������ϵ�ṹ���Ŵ����ơ�3.3 ���½ڸ������״��ȫ�������������
�������컨�ڷ�����,��LD˥���Ͽ�[34],���������ù��������������и���������״���Ŵ����������о�����FarmCPUģ�ͽ���ȫ�������������,����26���������½ڸ������״�����½ڸ���������������������أ�����������SNPλ��,���½ڸ��ǶȺ͵��½ڸ����û�м�����������SNP������Щ���������ı����,����һ���Ч����,λ�ڵ�6Ⱦɫ���ϵ�Chr.6.S_123079904��Chr.6.S_123079927���ͬʱ����½ڸ�����������������������о�����26����������SNP,����11��λ���Ѷ�λ��QTL���������ڣ���4�������Chr.6.S_108781517����½ڸ�������������,λ�ڲ̺���[20]���Ŀ��Ƹ����غ�������QTL����umc1257-umc1857�ڡ����Chr.6.S_34950886����½ڸ����������,λ��CAI��[5]��λ���Ŀ��Ƴ����ڵ��½ڸ����ȵ�����bins 6.00��6.01�ڡ�λ�ڵ�4��7��10Ⱦɫ���������������Chr.4.S_9975093��Chr.7.S_48142656��Chr.7.S_101804639��Chr.10.S_ 150110617����λ��ZHANG��[17]����BC2S3Ⱥ����Ľڸ��������״QTL�����ڡ����о��й���6����������SNP����GUO��[23]ͨ����������ϵ�����״QTL��õ�Meta-QTL�����ڡ����о���õ�26����������SNP���漰177����ѡ����,����135���й���ע�͵Ļ����漰��ת�����ס���������ת�˵��ס����ڵ��ס�����ø��,�����漰������л���������䡢������ص�����ѧ;������ѡ����Zm00001d037004ΪRas��ص���RABA1d,���ë�����й�;Zm00001d037010Ϊ��ë����صĵ���ø;����ѡ������ͨ��MaizeGDB���ҵ�����֪������ϵ��صĻ�����бȶ�,����Zm00001d037751�Ĺ�����NCBI��վ����¼��һЩ����ϵ��ػ���Ĺ������,��֪��ϵ��صĻ���Zm00001d003807ΪLEA��ص�����,Zm00001d037751����LEA�����Ǹ�������ǵ�����;ֵ��ע�����,����½ڸ��������Ͳ�����ص���������λ��Chr.6.S_123079904��Chr.6.S_123079927������Zm00001d037368������������,Zm00001d037368�Ĺ���δ֪;ZmSUT2��Zm00001d041192���ٽ�����ֲ��������������������γ�,zmsut2ͻ����ֲ���Ұ����ֲ����������������ʹ����С��������[35]�����,ZmSUT2�������ϵ������ء�
4 ����
��ͬ����������Խ�ϵ���¸�ϵ���ڲ�ͬ�̶ȵIJ���,���½ڸ�������������������ĸ������ֳ��½�������,���½ڸ��ǶȺ����������ĸ������ֳ�����������,�����غ�����ڸ���������������졣�����Խ�ϵ�ĸ�ϵ�ṹ�ڲ�ͬ��Ⱥ����ڲ���,�ô���Ⱥ�ĵ��½ڸ��������������Ƕȡ������������ؾ�����������Ⱥ�����ø��ܶ�SNP��Ƕ�����ϵ��6����״����GWAS����,����26������������SNPλ��,�漰177����ѡ����,����135���й���ע�͡��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
DOI:10.1093/aob/mcs293URL [��������: 1]
DOI:10.1016/j.tplants.2014.02.002URL [��������: 1]
[��������: 1]
DOI:10.2135/cropsci2008.03.0152URL [��������: 2]
[��������: 5]
[��������: 1]
[��������: 1]
DOI:10.1007/s11104-015-2554-xURL [��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 1]
DOI:10.1016/S2095-3119(13)60243-9URL [��������: 1]
[��������: 2]
DOI:10.1111/tpj.2018.93.issue-6URL [��������: 4]
DOI:10.1007/s10681-015-1533-xURL [��������: 1]
[��������: 1]
DOI:10.11674/zwyf.2011.0179URLMagsci [��������: 2]

�����ԭλ�����£����ø�ϵ��̬�����������Խ�ϵҴ478����312Ϊ�ױ�������BC4F3Ⱥ�壬���øĽ���PLABQTL�����еĸ���������ͼ���Գ����ڣ�����ǰ10 d���ཬ���ڣ�������15 d������ϵ��״�ı仯�͵��ϲ�����������QTL��λ�����������Ŵ����ơ������������ǰ����Ը����ء��ܸ�������������������������ȸ�ϵ��״������27��QTL������QTL������Ϊ5��2%15��7%��������Ⱦɫ���6��02��10��04����ͬʱ�������ŵ��ϲ����������ܸ���������������������״��QTLs��������ͬ����ʱ�ڼ��Ĺ�ͬQTL����8��������ǰ������Ƹ�ϵ������QTL�����������β�ͬ�������������ԣ����ҶԵ��ϲ��������γ�����Ҫ���ף���Ϊ�˽���������¸�ϵ�����������ͽ�һ�������Ŵ������춨���Ŵ�������
DOI:10.11674/zwyf.2011.0179URLMagsci [��������: 2]
�����ԭλ�����£����ø�ϵ��̬�����������Խ�ϵҴ478����312Ϊ�ױ�������BC4F3Ⱥ�壬���øĽ���PLABQTL�����еĸ���������ͼ���Գ����ڣ�����ǰ10 d���ཬ���ڣ�������15 d������ϵ��״�ı仯�͵��ϲ�����������QTL��λ�����������Ŵ����ơ������������ǰ����Ը����ء��ܸ�������������������������ȸ�ϵ��״������27��QTL������QTL������Ϊ5��2%15��7%��������Ⱦɫ���6��02��10��04����ͬʱ�������ŵ��ϲ����������ܸ���������������������״��QTLs��������ͬ����ʱ�ڼ��Ĺ�ͬQTL����8��������ǰ������Ƹ�ϵ������QTL�����������β�ͬ�������������ԣ����ҶԵ��ϲ��������γ�����Ҫ���ף���Ϊ�˽���������¸�ϵ�����������ͽ�һ�������Ŵ������춨���Ŵ�������
DOI:10.1007/s11032-011-9655-xURL [��������: 1]
DOI:10.2135/cropsci2016.01.0031URL [��������: 1]
DOI:10.1007/s10681-018-2283-3 [��������: 2]
[��������: 1]
DOI:10.1371/journal.pgen.1005767URL [��������: 1]
DOI:10.3864/j.issn.0578-1752.2012.11.001URLMagsci [��������: 1]
��Ŀ�ġ�ѡ���д����ԵIJ��Ͻ����Ŵ���������Ⱥ��ṹ�Ľ����ǽ��е�λ��������״���� �������о�������������ʵ������Ҫ�����������������ø�������ȫ�������40 ������SSR ��ǣ����û��ڲ��� �Ļ����ͼ���������820 �ݴ����й�����������Դ���ʻ������Խ�ϵ����ȫ������ɨ�裬ͨ��PowerMarker V3.25 ��Structure V2.3.3 ��������ʾ������������Ⱥ��ṹ�����������820 ���Խ�ϵ�У�40 ��SSR ������� �ĵ�λ����Ϊ10—72 ����ƽ��36.87 ��/λ�㣻���������Ϊ0.46—0.9458��ƽ��0.8430��PIC Ϊ0.43—0.94�� ƽ��0.83���������������K=5 ʱ����K ֵ�����Щ�Խ�ϵ���Ի��ֳ�5 ����Ⱥ������Ϊ����˹�ء��ô�� �ǡ�����ƽͷ�������P Ⱥ������Ⱥ��ƽ����λ�������Ŵ������Էֱ�Ϊ24.23 ��/λ����0.8145��22 ��/λ���� 0.8398��11.8 ��/λ����0.7054��17.45 ��/λ����0.7686 �Լ�14.65 ��/λ����0.7495�������ۡ��й������Խ� ϵ������ʵ�����γ�����Զ�����������Ⱥ���̺��˱ȽϷḻ���Ŵ����죬��ʾ���˽ϸ�ˮƽ�Ļ�������ԡ��� ͬ��Ⱥ֮����Ŵ�������ˮƽ����һ���IJ��죬�ڻ��ֵ�5 ����Ⱥ�У�����˹�����ô�����Ⱥ���Ŵ�������� �Ϸḻ�������������Խϸߣ����Ϊ�����P Ⱥ������ƽͷ��Խϵ͡�
DOI:10.3864/j.issn.0578-1752.2012.11.001URLMagsci [��������: 1]
��Ŀ�ġ�ѡ���д����ԵIJ��Ͻ����Ŵ���������Ⱥ��ṹ�Ľ����ǽ��е�λ��������״���� �������о�������������ʵ������Ҫ�����������������ø�������ȫ�������40 ������SSR ��ǣ����û��ڲ��� �Ļ����ͼ���������820 �ݴ����й�����������Դ���ʻ������Խ�ϵ����ȫ������ɨ�裬ͨ��PowerMarker V3.25 ��Structure V2.3.3 ��������ʾ������������Ⱥ��ṹ�����������820 ���Խ�ϵ�У�40 ��SSR ������� �ĵ�λ����Ϊ10—72 ����ƽ��36.87 ��/λ�㣻���������Ϊ0.46—0.9458��ƽ��0.8430��PIC Ϊ0.43—0.94�� ƽ��0.83���������������K=5 ʱ����K ֵ�����Щ�Խ�ϵ���Ի��ֳ�5 ����Ⱥ������Ϊ����˹�ء��ô�� �ǡ�����ƽͷ�������P Ⱥ������Ⱥ��ƽ����λ�������Ŵ������Էֱ�Ϊ24.23 ��/λ����0.8145��22 ��/λ���� 0.8398��11.8 ��/λ����0.7054��17.45 ��/λ����0.7686 �Լ�14.65 ��/λ����0.7495�������ۡ��й������Խ� ϵ������ʵ�����γ�����Զ�����������Ⱥ���̺��˱ȽϷḻ���Ŵ����죬��ʾ���˽ϸ�ˮƽ�Ļ�������ԡ��� ͬ��Ⱥ֮����Ŵ�������ˮƽ����һ���IJ��죬�ڻ��ֵ�5 ����Ⱥ�У�����˹�����ô�����Ⱥ���Ŵ�������� �Ϸḻ�������������Խϸߣ����Ϊ�����P Ⱥ������ƽͷ��Խϵ͡�
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1104/pp.114.241711URL [��������: 1]
URL [��������: 1]
URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}