 ,1
,1Genetic Analysis and Characterization of Hormone Response of Semi-Dwarf Mutant dw-1 in Brasscia napus L.
SONG Xi1, PU DingFu2, TIAN LuShen1, YU QingQing1, YANG YuHeng1, Dai BingBing1, ZHAO ChangBin1, HUANG ChengYun1, DENG WuMing,1通讯作者:
编委: 李莉
收稿日期:2019-01-21接受日期:2019-03-11网络出版日期:2019-05-16
| 基金资助: |
Received:2019-01-21Accepted:2019-03-11Online:2019-05-16

摘要
关键词:
Abstract
Keywords:
PDF (865KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
宋稀, 蒲定福, 田露申, 余青青, 杨玉恒, 代兵兵, 赵昌斌, 黄成云, 邓武明. 甘蓝型油菜半矮秆突变体dw-1的遗传分析与激素响应特性[J]. 中国农业科学, 2019, 52(10): 1667-1677 doi:10.3864/j.issn.0578-1752.2019.10.001
SONG Xi, PU DingFu, TIAN LuShen, YU QingQing, YANG YuHeng, Dai BingBing, ZHAO ChangBin, HUANG ChengYun, DENG WuMing.
0 引言
【研究意义】株高是影响作物抗倒性和丰产潜能的关键性状。生产上运用的甘蓝型油菜植株高大易倒伏,杂交油菜的普及则再次增大了倒伏风险[1]。倒伏可引起光能利用率下降、贪青迟熟、病害蔓延、经济性状恶化,最终引起产量下降。油菜结实期倒伏可减产15%—60%,花期倒伏甚至绝收,另外还可造成含油率等下降[2,3]。随着油菜全程机械化技术的大力推广,株高的限制则进一步凸显。植株高大,一方面结角层分散,成熟期一致性差,机收损失率大;另一方面分枝交织度高,易堵塞机具,影响机收效率;更重要的是倒伏田块机收困难。欧洲以此提出机收油菜的理想株高1.2—1.3 m,傅廷栋[4]认为中国机收油菜品种株高以小于1.5 m为宜。可见加强矮秆、半矮秆资源的创制、研究与应用对实现油菜矮化育种意义重大。【前人研究进展】油菜矮化资源主要由化学诱变和自然突变产生,其中少数为多基因控制,如甘蓝型油菜矮化自交系74-1002[5]为2对加性-显性-上位性主基因调控,突变体6-8808[6]为2对隐性重叠基因控制,99CDAM[7]为3对隐性基因控制且存在细胞质效应。大部分矮秆材料为单基因遗传,根据基因效应不同分为隐性、显性、部分显性或加性。ZENG等[8]通过EMS诱变获得甘蓝型突变体bnaC.dwf株高约95 cm,为1对隐性基因控制,且对赤霉素不敏感。澳大利亚引进的甘蓝型油菜矮源一号株高仅24 cm,为单显性基因控制,显性度0.66—0.80[9]。WANG等[10,11]报道了2个EMS诱变的甘蓝型矮化突变体Bndwf/dcl1和Bndwf1株高约50和80 cm,均由单个显性基因控制,分别定位于C5、A9染色体。WILLIAMS等[12]在快繁过程中发现了4个白菜型矮秆突变体,其中dwf2[13]株高约45 cm,受1对不完全显性基因控制,对赤霉素不敏感;MUANGPROM等[14,15]利用同源比较作图的方法将矮秆基因定位于A6染色体,并揭示了矮化机理,后续将其成功转入甘蓝型油菜。李云等[16]在杂交选育后代中发现的bndf-1株高约75 cm,由1对部分显性基因调控。石淑稳等[17]利用EMS诱变获得2株突变体ds-1和ds-2,株高分别为106和95 cm,其中ds-1[18]为1对不完全显性基因控制,对赤霉素不敏感,被认为是一个极具育种价值的矮源;LIU等[19]将其定位于A6染色体并进行了基因克隆与功能分析。ZHAO等[20]报道的ds-3株高与ds-1接近,为1对不完全显性基因控制,并将DS-3定位于C7,但其对株高的调控作用弱于DS-1。FOISSET等[1,21]由EMS诱变获得甘蓝型油菜矮化突变体B192受1对加性基因bzh控制,定位于A6染色体;BARRET等[22]开发了连锁的SCAR标记;bzh是第一个在生产上应用的油菜矮秆基因,近年来,研究发现导入该基因的半矮秆杂交种较基因型为Bzh/Bzh的正常秆杂交种具有更高的氮肥利用率[23]。NDF-1[24]是利用快中子照射与DES诱变获得甘蓝型矮化突变体,株高约70 cm,为1对加性效应基因控制;GAO等[25]发现其对赤霉素不敏感;LI等[26]对矮化相关基因进行了序列分析。此外,前人通过芥甘杂交、甘白杂交等远缘杂交与航空诱变的方式,也获得了100—170 cm的甘蓝型半矮秆油菜资源[27,28,29,30]。【本研究切入点】目前,仅少数矮源进行了基因定位、克隆、功能分析等深入研究或育种利用,国内还未有油菜矮秆基因在生产上成功应用的报道。优异矮秆种质资源匮乏是当前油菜矮化育种面临的主要问题。【拟解决的关键问题】本研究通过对甘蓝型油菜中发现的自然矮化突变体田间性状进行表型鉴定,采用主基因+多基因混合遗传模型进行株高遗传研究,同时就矮化与激素的关系进行形态学、生理学分析。明确该矮化突变体的遗传规律与致矮的激素类型,综合评估其利用潜能,为其在油菜矮化育种中的应用提供理论指导,同时为后续基因定位、克隆及分子标记辅助选择奠定基础。1 材料与方法
1.1 试验材料
将甘蓝型油菜高秆品系141492(由南充市农业科学院选育的低芥酸、低硫苷波里马细胞质雄性不育保持材料(南B7×SC94005)F5与高芥酸、高硫苷恢复系7R杂交并连续自交4代结合室内品质测试选获的“双低”保持材料)自交6代后发现的自然矮化突变体经游离小孢子培养获得DH系群体,选取1个半矮DH系,暂命名dw-1。其自交后代多年种植平均株高约95 cm(变幅83—105 cm),苗期叶色深绿,叶片匍匐,表面少许凹陷,分枝、茎秆等形态正常。所用半矮秆材料均为dw-1自交后代。外源GA3为sigma分装,购于成都康迪生物技术有限公司。
1.2 遗传群体构建与田间性状调查
2013年8月于贵州威宁夏繁配制高秆(P2)×半矮秆(P1)正反杂交组合F1,2014年正季按照随机区组设计种植正反交组合各330株,成熟期株高方差分析显示正反交F1差异不显著(F=1.81,F0.05=18.51)。同时,将高秆×半矮秆正交F1套袋自交获得F2群体,将正交F1作母本分别与P1、P2杂交获得回交群体B1、B2。2015年9月28日于南充市农业科学院潆溪试验基地(北纬30.52°,东经106.02°,海拔290 m;全生育期日均温度1.1—23.0℃,日照时数约547.6 h)播种6个世代群体。本田前作大豆,黏壤土,土壤基础养分状况为pH7.07,有机质含量9.7g·kg-1、碱解氮59.2 mg·kg-1、速效磷2.1 mg·kg-1、速效钾92.0 mg·kg-1。为防止倒伏,以风向垂直方向开厢,施复合肥25 kg·667m-2(N:P2O5:K2O=30:5:5,总养分≥40%),菜籽饼粕50 kg·667m-2(有机质含量≥75%,总养分N+P2O5+K2O≥8%)。11月6日移栽,株行距0.334 m×0.5 m,每行11株。完全随机区组设计,3次重复,各重复P1、P2、F1、B1、B2、F2分别种植3、3、5、15、15、28行,世代间不设保护行,四周设置2行保护行。成熟期测量各群体株高,其中,P1 99株、P2 96株、F1 167株、B1 492株、B2 490株、F2 920株。同时成熟期按小区对两亲本全生育期和菌核病病指进行田间观察记载,各重复再分别取主茎无病的连续5株考察株高、分枝部位高度、重心高度、主序有效长、主序角果数、一次分枝数、二次分枝数、单株角果数等主要农艺性状,整株风干后按单株脱粒考察单株产量与千粒重,每角粒数由单株产量/单株角果数/千粒重×1000换算。1.3 光暗处理形态学调查
参考王传富[6]的方法,选取正常饱满的高、矮种子10粒点播于培养皿,25℃光照培养间恒温培养。分别设置24 h黑暗(纸箱遮光)和16 h光照/8 h黑暗2个处理,3次重复,7 d后考察幼苗表型并测量下胚轴长度。1.4 赤霉素诱导试验
1.4.1 下胚轴对赤霉素敏感性测验 参考高勇等[31]方法,将草炭土与泥土4:1混合均匀,等量称取装于纸杯,选取正常饱满的高、矮种子12粒播种于纸杯,并放置25℃培养间16 h光照/8 h黑暗恒温培养。待子叶稍展开,每隔1天喷施1次GA3,露白25 d后测量下胚轴长度。GA3处理浓度为10、100、200、500和1 000 mg·L-1,清水为对照,每个处理3次重复。1.4.2 茎秆对赤霉素敏感性测验 参照GAO等[25]方法略作修改,选取土壤肥力均匀的防鸟网室,2016年11月2日穴播。株行距0.167 m×0.5 m,每行12株,每个处理3行。五叶期各行选取长势一致的植株7—9株挂牌标记,按行逐一喷施GA3直至叶片渐湿,每周处理1次至抽苔期。GA3处理浓度为10、50、100、200和500 mg·L-1,对照喷清水,成熟时测量标记单株株高。
1.5 数据处理
利用DPS 8.01软件对所有测量数据进行方差分析;采用最新SEA软件包(R语言编写)对株高进行植物数量性状混合遗传模型主基因+多基因多世代联合分离分析[32,33]。通过极大似然分析与IECM(iterated expectation and conditional maximization)算法,估算各世代成分分布参数及各种模型极大似然函数值。采用AIC值最小原则筛选候选模型,并进行一组适合性检验,包括均匀性检验(U12 U22 U32)、Smirnov检验(nW2)和Kolmogorov检验(Dn),以此确定最佳模型,并根据成分分布参数估算基因效应值及遗传方差。SEA软件包由华中农业大学植物科技学院章元明教授提供。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1dw-1及野生型苗期、成熟期田间表型
bar=10 cm
Fig. 1Phenotypes of seedling stage and mature stage of dw-1 and wild type in the field
2 结果
2.1 表型鉴定
田间种植4 000株/667 m2条件下,dw-1自交后代平均单株产量16.2 g,比野生型高秆降低37.2%(表1)。进一步剖析产量构成因子发现,其单株角果数比高秆增加12.3%,达极显著水平;千粒重变化不显著;每角粒数极显著降低,比高秆下降42.2%,是造成矮秆单株产量降低的主要因素。农艺性状中,除二次分枝数较高秆极显著增加外,其余性状均显著或极显著降低,其中分枝部位高度降幅最大,达72.5%,其次是株高与重心高度,分别降低38.2%和34.2%。另外,全生育期较高秆缩短约5.4 d,菌核病病指显著增加。Table 1
表1
表1dw-1与野生型农艺性状、经济性状及抗病性
Table 1
| 材料 Material | 株高 PH | 分枝部 位高度 BH | 重心 高度 HGC | 全生 育期 GD | 主序 有效长 LMI | 主序 角果数 SMI | 一次 分枝数 PB | 二次 分枝数 SB | 单株 角果数 SP | 千粒重 SW | 每角 粒数 SS | 单株 产量 YP | 菌核病 病指 DISSR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WT | 194.1±4.6 | 57.1±1.8 | 90.9±2.5 | 208.7±0.6 | 76.9±1.4 | 109.9±2.4 | 13.8±0.5 | 4.5±1.6 | 745.1±27.1 | 3.42±0.04 | 10.2±0.3 | 25.8±1.3 | 8.1±3.0 |
| dw-1 | 120.0±4.7 | 15.7±2.0 | 59.8±2.6 | 203.3±0.6 | 66.2±1.5 | 80.1±3.5 | 11.6±0.2 | 17.2±1.3 | 836.5±17.1 | 3.27±0.05 | 5.9±0.6 | 16.2±1.7 | 19.4±1.6 |
| ±WT(%) | -38.2** | -72.5** | -34.2** | -2.6** | -13.9* | -27.1** | -15.9** | 282.2** | 12.3** | -4.4 | -42.2** | -37.2** | 139.5* |
新窗口打开|下载CSV
2.2 遗传分析
2.2.1 株高表型数据分布 由表2可知,P1平均株高115.8 cm,P2平均株高201.0 cm,F1株高介于双亲之间,略低于双亲均值。B1、B2、F2群体均存在明显多峰现象,可见矮秆性状存在主效基因的控制。Table 2
表2
表26个世代株高次数分布及基本统计量
Table 2
| 群体 Population | 株高分布Distribution of plant height (cm) | 样本 容量 Sample size | 平均值± 标准差 Mean±SD | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤90 | 91-100 | 101-110 | 111-120 | 121-130 | 131-140 | 141-150 | 151-160 | 161-170 | 171-180 | 181-190 | 191-200 | 201-210 | 211-220 | >220 | |||
| P1 | 12 | 22 | 30 | 26 | 9 | 99 | 115.8±11.4 | ||||||||||
| F1 | 1 | 6 | 40 | 78 | 35 | 7 | 167 | 155.0±8.4 | |||||||||
| P2 | 2 | 5 | 6 | 29 | 35 | 15 | 4 | 96 | 201.0±11.6 | ||||||||
| B1 | 3 | 13 | 26 | 62 | 80 | 36 | 40 | 96 | 99 | 28 | 7 | 2 | 492 | 142.4±22.7 | |||
| B2 | 1 | 13 | 31 | 67 | 83 | 45 | 13 | 19 | 39 | 68 | 63 | 44 | 4 | 490 | 169.3±31.3 | ||
| F2 | 19 | 32 | 50 | 51 | 59 | 96 | 141 | 131 | 67 | 38 | 24 | 69 | 86 | 45 | 12 | 920 | 155.4±35.0 |
新窗口打开|下载CSV
2.2.2 候选遗传模型筛选及适合性检验 将6世代株高表型数据进行主基因+多基因联合分离分析,获得5类24种遗传模型的极大似然函数值和AIC值(表3)。根据AIC值最小原则,筛选出候选模型4个,分别为B-1(2对加性-显性-上位性主基因)、D-0(1对加性-显性主基因+加性-显性-上位性多基因)、E-0(2对加性-显性-上位性主基因+加性-显性-上位性多基因)、E-3(2对加性主基因+加性-显性多基因)。进一步对候选模型进行适合性检验,其中,E-3模型2个统计量达到显著水平,B-1、E-0模型均有1个统计量达到显著水平,D-0模型30个统计量均未达到显著水平,且其AIC值最低,因此,确定D-0为最佳遗传模型。
Table 3
表3
表3各模型的极大对数似然函数值、AIC值
Table 3
| 模型 Model | 模型含义 Implication of model | 极大似然值 MLV | AIC值 AIC value | 模型 Model | 模型含义 Implication of model | 极大似然值 MLV | AIC值 AIC value | |
|---|---|---|---|---|---|---|---|---|
| A-1 | 1MG-AD | -10417.3 | 20842.6 | D-0 | MX1-AD-ADI | -10267.8 | 20559.5 | |
| A-2 | 1MG-A | -10488.1 | 20982.2 | D-1 | MX1-AD-AD | -10339.9 | 20697.8 | |
| A-3 | 1MG-EAD | -10715.3 | 21436.7 | D-2 | MX1-A-AD | -10414.3 | 20844.6 | |
| A-4 | 1MG-NCD | -10981.0 | 21968.1 | D-3 | MX1-EAD-AD | -10450.5 | 20916.9 | |
| B-1 | 2MG-ADI | -10277.0 | 20574.1 | D-4 | MX1-NCD-AD | -10519.0 | 21053.9 | |
| B-2 | 2MG-AD | -10383.8 | 20779.7 | E-0 | MX2-ADI-ADI | -10267.0 | 20570.1 | |
| B-3 | 2MG-A | -10575.4 | 21158.8 | E-1 | MX2-ADI-AD | -10357.0 | 20744.1 | |
| B-4 | 2MG-EA | -10702.4 | 21410.8 | E-2 | MX2-AD-AD | -10334.8 | 20691.7 | |
| B-5 | 2MG-CD | -10715.4 | 21438.7 | E-3 | MX2-A-AD | -10280.7 | 20579.5 | |
| B-6 | 2MG-EAD | -10796.8 | 21599.5 | E-4 | MX2-EA-AD | -10517.1 | 21050.1 | |
| C-0 | PG-ADI | -10543.2 | 21106.4 | E-5 | MX2-CD-AD | -10450.4 | 20918.9 | |
| C-1 | PG-AD | -10579.2 | 21172.5 | E-6 | MX2-EAD-AD | -10525.1 | 21066.1 |
新窗口打开|下载CSV
2.2.3 遗传参数估计 根据IECM算法估算的最适遗传模型下6个世代各成分分布参数,利用最小二乘法估计株高的遗传参数。由表4可知,主基因加性效应为-47.5,与前面定义的P1为矮秆亲本相对应,该基因存在使株高降低。显性效应-7.7,表明矮秆基因显性效应作用大于高秆基因。显性度为0.2,说明主基因加性效应起主导作用,因此,高、矮杂交F1株高呈现中亲值略偏向于矮秆亲本。B1、B2、F2主基因遗传率分别为76.0%、84.0%和85.0%,多基因遗传率分别为4.1%、5.6%和6.7%,可见株高的遗传主要受到1对主基因的控制,多基因的遗传贡献率低。另外,3个世代环境变异分别为19.9%、10.4%和8.3%,故dw-1的育种利用可在早世代对株高进行选择。
Table 4
表4
表4D-0模型下株高遗传参数估计
Table 4
| 模型 Model | 一阶遗传参数 1st order genetic parameter | 估计值 Estimate | 二阶遗传参数 2nd order genetic parameter | 估计值 Estimate | ||
|---|---|---|---|---|---|---|
| B1 | B2 | F2 | ||||
| MX1-AD-ADI | m1 | 163.3 | σ2p | 513.4 | 980.7 | 1228.3 |
| m2 | 162.8 | σ2mg | 390.2 | 824.1 | 1043.8 | |
| m3 | 153.4 | σ2pg | 21.2 | 54.6 | 82.5 | |
| m4 | 167.8 | h2mg(%) | 76.0 | 84.0 | 85.0 | |
| m5 | 150.6 | h2pg(%) | 4.1 | 5.6 | 6.7 | |
| m6 | 156.2 | h2 mg +pg (%) | 80.1 | 89.6 | 91.7 | |
| d | -47.5 | |||||
| h | -7.7 | |||||
| h/d | 0.2 | |||||
新窗口打开|下载CSV
2.3 矮化生理分析
2.3.1 光暗处理形态构建 为弄清dw-1矮化是否与BR途径相关,进行了光暗处理幼苗形态学分析(图2)。光照处理下,dw-1子叶绿色、平展、有真叶;黑暗处理下,dw-1子叶黄化、呈闭合弯钩状、无真叶,光暗形态建成均表现正常。而前人在油菜近源物种拟南芥中研究发现,det2[34]、dwf4[35]、cpd[36]等BR合成突变体幼苗暗生长表现为下胚轴短、顶端弯钩消失、子叶展开真叶出现、脱黄化等光形态建成,故此推测dw-1致矮机制与BR途径无关。且无论光照或是黑暗处理,dw-1下胚轴长度均极显著小于野生型(图3)。图2
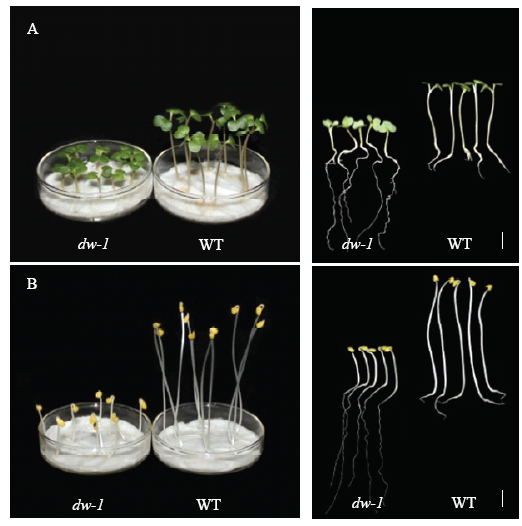 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2光照(A)与黑暗(B)处理dw-1及野生型幼苗表型
bar= 1cm
Fig. 2Phenotypes of dw-1 and wild type seedlings grown in light (A) and dark (B) condition
图3
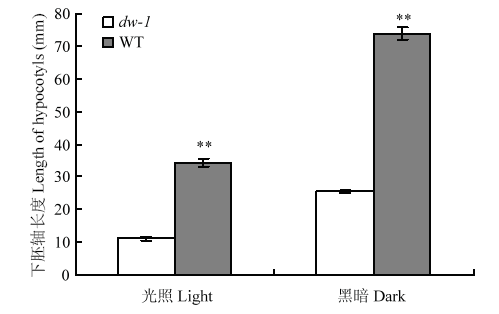 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3光暗处理下dw-1及原始高秆下胚轴长度
**表示在0.01水平差异显著
Fig. 3Hypocotyls length of dw-1 and wild type in light and dark condition
** indicates significant difference at 0.01 levels
2.3.2 赤霉素敏感性分析 下胚轴对GA3敏感性测验结果表明(图4-A),随着外源GA3处理浓度增加, 野生型高秆下胚轴逐渐伸长,当浓度达100 mg·L-1时下胚轴长度比野生型清水对照(CK1)显著伸长,浓度为1 000 mg·L-1时下胚轴伸长达最大值。dw-1在10—500 mg·L-1浓度GA3处理下,下胚轴长度随浓度增加伸长不明显,当浓度达1 000 mg·L-1时,下胚轴比其清水对照(CK2)显著伸长,伸长率为18.3%,但仍显著低于同浓度处理野生型下胚轴长度。为进一步明确dw-1株高对赤霉素的响应,进行了GA3诱导茎秆伸长试验,结果(图4-B)表明,当外施GA3浓度达50 mg·L-1时,野生型株高较CK1显著降低,200 mg·L-1后随浓度继续增大,不同浓度间株高变化不明显。而dw-1在外源GA3浓度10—100 mg·L-1时,株高相比CK2变化不明显;当处理浓度为200—500 mg·L-1时,株高显著增加,伸长率分别为9.5%和8.3%,但仍极显著低于同浓度处理野生型株高。综合dw-1下胚轴与茎秆对赤霉素的响应结果,发现具有相似的规律,均表现对低浓度GA3反应不明显,高浓度处理下有小部分恢复。说明dw-1相比野生型对赤霉素的敏感程度明显下降,且下胚轴相比茎秆对赤霉素的反应更为钝感。
图4
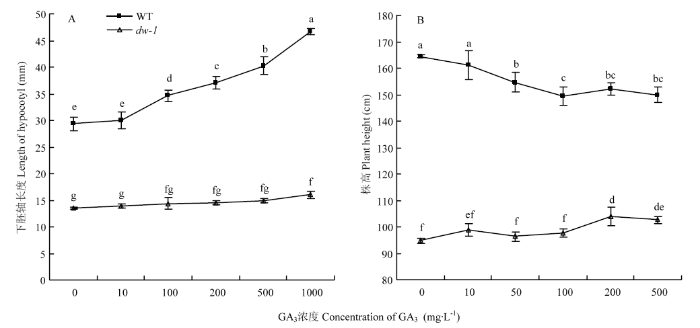 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4赤霉素对下胚轴(A)和茎秆(B)的伸长效应
不同字母表示处理间在0.05水平差异显著
Fig. 4Effects of GA3 on hypocotyl (A) and stem (B) elongation
Different letters represent significant difference between different treatments at 0.05 level
3 讨论
水稻、小麦等禾本科作物矮化育种已十分成熟,迄今登记的水稻矮秆基因已达100余个(http://shigen. nig.ac.jp/rice/oryzabase/),其中大部分为单隐性基因控制,少数为显性或部分显性,生产上主要利用的是隐性基因sd-1,其介导了赤霉素的生物合成[37]。小麦生产上主要利用的是Rht系列基因,目前已有20多个被报道[38],包括单隐性基因、单显性或部分显性基因,生产上广泛应用的是Rht1、Rht2和Rht8,其中Rht1和Rht2参与了赤霉素信号转导[39]。目前,油菜中发现的矮源仍十分有限,具有育种利用价值的矮化资源匮乏。本研究发现的矮秆材料dw-1株高遗传稳定,其自交后代多年种植成熟期平均株高约95 cm(文中稀植条件下平均株高115.8 cm),与突变体ds-1、bnaC.dwf株高接近,同属于甘蓝型半矮秆油菜。相比bndf-1、Bndwf1、NDF-1等矮秆突变体高出20 cm左右,比Bndwf/dcl1、矮源1号则高45—70 cm,株高的部分优势,使得dw-1单株生产潜力较好,产量下降程度明显较bndf-1[16]、NDF-1[24]缓慢。从产量构成的变化看,目前已报道的矮化突变体主要有以下三类,分别为角果数显著减少,如ds-1[18];角果数与角粒数显著减少,如NDF-1[24];以及角果数、角粒数、千粒重均显著减少,如bndf-1[16]。可见单株角果数减少在矮化突变体中发生较为普遍,但dw-1单株角果数较原始高秆却极显著增加,其主要源于二次分枝角果数的增加;dw-1角粒数极显著降低,也是造成单株产量下降的主要原因,千粒重则变化不大。此外,发现dw-1生育期提前,这与大多数矮化突变体表现正好相反,而与梅德胜等[7]报道的99CDAM相似,其早熟特性有利于机收品种的选育。菌核病抗性下降则是目前大多数矮秆材料普遍存在的问题,可能与株高、分枝部位高度大幅降低,田间通透性下降有关,且早熟品种相比中熟品种也更易感病,属于植物的时间避病机制。为此,笔者尝试将dw-1与不同来源中抗菌核病资源进行杂交选育,以期提高其自身菌核病抗性。同时近两年通过化学杀雄方式将dw-1与自育的系列优良高秆材料进行组配,发现直接配组仍然能够筛选到部分田间菌核病发病较轻的组合。因此,就田间综合性状表现来看,认为dw-1是一个优良的半矮秆材料,具有较好的利用前景。油菜矮化突变体的遗传,前人研究表明不同的矮源可受1—3对主基因控制,但以单基因遗传居多。本研究发现dw-1矮化表型以1对加性-显性主基因控制为主,同时存在多基因的作用。从单个主基因的性质看,已报道的矮源除B192[1]、bnaC.dwf[8]、NDF-1[24]、10D130[30]外,大都表现矮对高为显性,这点dw-1与大多数矮化突变体一致。但dw-1主基因显性度不高,相比而言加性效应作用更大,其主基因的遗传规律与突变体dwf1[13]、bndf-1[16]、ds-1[18]、ds-3[20]较为相似。目前理论与大量育种实践表明1.5—1.7 m是机收油菜较为理想的株高[4,40-41],能较好的兼顾抗倒性与产量提升,而普通油菜品种株高1.7—2.2 m。因此,加性基因控制为主的矮秆资源更能切合油菜杂种优势利用为主的育种现状,育种者可根据杂交种F1株高实际需求的不同,选择不同高度的高秆亲本与dw-1进行组配,达到对杂交种F1理想株高人为调控的目的。
作物矮化大多与GA、BR生物合成或信号转导有关,少数与生长素(IAA)有关[42]。KUMAR等[43]在水稻中最先提出利用GA3敏感性反应作为鉴定新矮源的方法。目前激素对油菜矮化突变体的影响研究并不多,已报道的突变体主要与赤霉素相关。其中,M176[44]及快繁过程发现的白菜型油菜矮秆突变体rosette和dormant[45]为赤霉素敏感型;ds-3[20]为赤霉素敏感性减弱应答类型;6-8808[6]、bnaC.dwf[8]、dwf2[13]、ds-1[18]、NDF-1[25]为赤霉素不敏感类型。本文发现dw-1也属于赤霉素敏感性降低类型,高浓度赤霉素处理对下胚轴和茎秆伸长虽有显著促进作用,但不能完全恢复至野生型对照表型,且诱导下胚轴伸长所需浓度高于茎秆伸长浓度。相比ds-3,下胚轴伸长虽不及同浓度赤霉素处理野生型表型,但在低浓度处理下即可显著超过野生型对照表型;同样ds-1在外源赤霉素浓度100 mg·L-1时下胚轴长度亦可显著超过正常秆对照表型。可见就下胚轴对赤霉素的反应而言,dw-1较ds-1、ds-3更为钝化。另外黑暗处理下ds-1与dw-1表现也存在差异,前者下胚轴长度与正常秆品种间并无差异,而dw-1下胚轴长度仍不及野生型1/2。但是,相比ds-1、NDF-1等赤霉素不敏感突变体,外施赤霉素对茎秆伸长无显著作用,而dw-1茎秆对赤霉素的敏感性明显增强。因此,初步判定dw-1可能为一个新的半矮秆资源,其矮化与赤霉素信号转导途径相关基因的突变有关,后续分子标记与定位研究仍在进行中。
4 结论
dw-1矮化表型受到1对加性-显性主基因+加性-显性-上位性多基因控制,主基因遗传率较高,适于早世代选择。其矮化机制与油菜素内酯途径无关,为赤霉素敏感性减弱应答类型。dw-1是一个优良的半矮秆材料,可在油菜株高改良中加以利用。(责任编辑 李莉)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1673-887X.2008.05.005URL [本文引用: 2]

正1.中国油菜生产20世纪五六十年代,我国油菜年均种植面积为170万~190万hm2,单产500kg/hm2,年总产85万~95万t。2000年以来,我国油菜年种植面积710万~740万hm2,单产1500~1700kg/hm2,年总产1065万~1258万t。面积、单产增加了3倍多,总产增加了10多倍。
DOI:10.3969/j.issn.1673-887X.2008.05.005URL [本文引用: 2]
正1.中国油菜生产20世纪五六十年代,我国油菜年均种植面积为170万~190万hm2,单产500kg/hm2,年总产85万~95万t。2000年以来,我国油菜年种植面积710万~740万hm2,单产1500~1700kg/hm2,年总产1065万~1258万t。面积、单产增加了3倍多,总产增加了10多倍。
URL [本文引用: 1]
URL [本文引用: 1]
[D].
[本文引用: 3]
[D].
[本文引用: 3]
DOI:10.3321/j.issn:0253-9772.2006.07.015URLMagsci [本文引用: 2]
随着杂种优势的利用,油菜株高增加了20cm以上,导致油菜生长后期倒伏的风险加大。通过利用特殊矮秆基因来控制株高将是解决倒伏问题的有效方法。在甘蓝型油菜自交多代品系中发现了一个株高85cm左右的矮秆突变株99CDAM,该突变株具有开花早、分枝多等优良特性,产量性状和品质性状较好,各性状都能稳定遗传,具有重要的育种价值。对99CDAM和高秆品系2091、7045和7350的正反交F1以及99CDAM和2091的F2、BC1及F2:3代的遗传分析结果表明: 99CDAM的矮秆性状受3对隐性基因控制,存在母体细胞质效应,与以往的甘蓝型矮秆油菜有明显的区别。
DOI:10.3321/j.issn:0253-9772.2006.07.015URLMagsci [本文引用: 2]
随着杂种优势的利用,油菜株高增加了20cm以上,导致油菜生长后期倒伏的风险加大。通过利用特殊矮秆基因来控制株高将是解决倒伏问题的有效方法。在甘蓝型油菜自交多代品系中发现了一个株高85cm左右的矮秆突变株99CDAM,该突变株具有开花早、分枝多等优良特性,产量性状和品质性状较好,各性状都能稳定遗传,具有重要的育种价值。对99CDAM和高秆品系2091、7045和7350的正反交F1以及99CDAM和2091的F2、BC1及F2:3代的遗传分析结果表明: 99CDAM的矮秆性状受3对隐性基因控制,存在母体细胞质效应,与以往的甘蓝型矮秆油菜有明显的区别。
[本文引用: 3]
URL [本文引用: 1]
甘蓝型矮秆油菜“矮源1号”鉴定研究初报浦惠明,戚存扣,傅寿仲(江苏省农业科学院经济作物所,南京210014)矮源1号是江苏省农科院经作所1990年从澳大利亚太平洋种子公司引进的一个甘蓝型矮秆油菜,国外原名Dwarf,现定名为矮源1号。经几年观察,矮源...
URL [本文引用: 1]
甘蓝型矮秆油菜“矮源1号”鉴定研究初报浦惠明,戚存扣,傅寿仲(江苏省农业科学院经济作物所,南京210014)矮源1号是江苏省农科院经作所1990年从澳大利亚太平洋种子公司引进的一个甘蓝型矮秆油菜,国外原名Dwarf,现定名为矮源1号。经几年观察,矮源...
DOI:10.1186/s12870-016-0865-6URLPMID:4991092 [本文引用: 1]
Key genes related to plant type traits have played very important roles in the “green revolution” by increasing lodging resistance and elevating the harvest indices of crop cultivars. Although there have been numerous achievements in the development of dwarfism and plant type inBrassica napusbreeding, exploring new materials conferring oilseed rape with efficient plant types that provide higher yields is still of significance in breeding, as well as in elucidating the mechanisms underlying plant development. Here, we report a new dwarf architecture with down-curved leaf mutant (Bndwf/dcl1) isolated from an ethyl methanesulphonate (EMS)-mutagenizedB. napusline, together with its inheritance and gene mapping, and pleiotropic effects of the mapped locus on plant-type traits. We constructed a high-density single-nucleotide polymorphism (SNP) map using a backcross population derived from theBndwf/dcl1mutant and the canola cultivar ‘zhongshuang11’ (‘ZS11’) and mapped the dwarf architecture with the down-curved leaf dominant locus,BnDWF/DCL1, in a 6.58-cM interval between SNP marker bins M46180 and M49962 on the linkage group (LG) C05 ofB. napus.Further mapping with other materials derived fromBndwf/dcl1narrowed the interval harbouringBnDWF/DCL1to 17502kb in length and this interval contained 16 annotated genes. Quantitative trait locus (QTL) mappings with the backcross population for plant type traits, including plant height, branching height, main raceme length and average branching interval, indicated that the mapped QTLs for plant type traits were located at the same position as theBnDWF/DCL1locus. This study suggests that theBnDWF/DCL1locus is a major pleiotropic locus/QTL inB. napus,which may reduce plant height, alter plant type traits and change leaf shape, and thus may lead to compact plant architecture. Accordingly, this locus may have substantial breeding potential for increasing planting density. The online version of this article (doi:10.1186/s12870-016-0865-6) contains supplementary material, which is available to authorized users.
DOI:10.1007/s00122-016-2718-yURLPMID:27147069 [本文引用: 1]
Abstract KEY MESSAGE: A saturated map was constructed using SNP markers to fine-map a Brassica napus dominant locus for dwarf mutant onto a 152-kb interval of chromosome A09 containing 14 genes. Major dwarf loci in crops may play important roles in crop improvement and developmental genetics. The present study investigated and fine-mapped a Brassica napus dwarf-dominant locus BnDWF1. Plants carrying the BnDWF1 locus in populations derived from 'zhongshuang11' and Bndwf1 have deep-green leaves and dwarf architecture that differ sharply from tall plants with normal green leaves. BnDWF1, as a major locus controlling plant height, showed a very high heritability (0.91-0.95). To map this locus, a high-density single-nucleotide polymorphism map was constructed, and the BnDWF1 locus was mapped at an interval between single-nucleotide polymorphism markers, M19704 and M19695, on linkage group A09 of B. napus, with five co-segregating single-nucleotide polymorphism markers. Furthermore, fine mapping narrowed the interval harboring BnDWF1 to 152 kb in length in B. napus. This interval contains 14 annotated or predicted genes, seven of which are candidates responsible for the dwarf trait. This study provides an effective foundation for the study of plant height regulation and plant type breeding in B. napus.
[本文引用: 1]
DOI:10.1007/s00122-003-1551-2URLPMID:14727029 [本文引用: 3]
Dwarf genes have been valuable for improving harvestable yield of several crop plants and may be useful in oilseed Brassica . We evaluated a dwarf gene, dwf2 , from Brassica rapa in order to determine its phenotypic effects and genetic characteristics. The dwf2 mutant was insensitive to exogenous GA 3 for both plant height and flowering time, suggesting that it is not a mutation in the gibberellin biosynthesis pathway. The dwarf phenotype was controlled by a semidominant allele at a single locus. Near-isogenic lines that were homozygous or heterozygous for dwf2 had 47.4% or 30.0% reduction in plant height, respectively, compared to the tall wild-type line, and the reduction was due to reduced internode length and number of nodes. The dwf2 homozygous and heterozygous lines had the same or significantly higher numbers of primary branches than the wild-type line, but did not differ in flowering time. The DWF2 gene was mapped to the bottom of linkage group R6, in a region having homology to the top of Arabidopsis thaliana chromosome 2. The map position of DWF2 in comparison to markers in A. thaliana suggests it is a homolog of RGA ( repressor of ga1-3 ), which is a homolog of the wheat 鈥淕reen Revolution鈥 gene. This dwarf gene could be used to gain more insight on the gibberellin pathway and to reduce lodging problems in hybrid oilseed Brassica cultivars.
DOI:10.1104/pp.104.057646URL [本文引用: 1]
Mutations in the biosynthesis or signaling pathways of gibberellin (GA) can cause dwarfing phenotypes in plants, and the use of such mutations in plant breeding was a major factor in the success of the Green Revolution. DELLA proteins are GA signaling repressors whose functions are conserved in different plant species. Recent studies show that GA promotes stem growth by causing degradation of DELLA proteins via the ubiquitin-proteasome pathway. The most widely utilized dwarfing alleles in wheat (Triticum aestivum; e.g. Rht-B1b and Rht-D1b) encode GA-resistant forms of a DELLA protein that function as dominant and constitutively active repressors of stem growth. All of the previously identified dominant DELLA repressors from several plant species contain N-terminal mutations. Here we report on a novel dwarf mutant from Brassica rapa (Brrga1-d) that is caused by substitution of a conserved amino acid in the C-terminal domain of a DELLA protein. Brrga1-d, like N-terminal DELLA mutants, retains its repressor function and accumulates to high levels, even in the presence of GA. However, unlike wild-type and N-terminal DELLA mutants, Brrga1-d does not interact with a protein component required for degradation, suggesting that the mutated amino acid causes dwarfism by preventing an interaction needed for its degradation. This novel mutation confers nondeleterious dwarf phenotypes when transferred to Arabidopsis (Arabidopsis thaliana) and oilseed rape (Brassica napus), indicating its potential usefulness in other crop species.
DOI:10.1007/s11032-005-3734-9URL [本文引用: 1]
Lodging can be a serious problem in oilseed Brassica napus . Dwarf genes have been used to reduce lodging in other crops, and their use in wheat and rice was a major factor in the success of the ‘ Green Revolution ’. We previously reported on a single, semi-dominant, gibberellin insensitive dwarf mutant of B. rapa ( Brrga1-d ), and provided evidence for homology of this gene to the wheat ‘ Green Revolution ’ dwarf gene. In this paper, we report using interspecific hybridization of B. rapa and B. oleracea and embryo rescue to resynthesize B. napus containing the Brrga1-d dwarf gene. The dwarf gene was backcrossed into two parents of a commercial hybrid combination and evaluated as inbred and hybrid lines in field experiments. The Brrga1-d gene reduced plant height and lodging in inbred and hybrid lines of B. napus , even when present as a single dose in heterozygous genotypes. Seed yields of inbred lines homozygous for Brrga1-d were reduced compared to near isogenic wild-type inbreds, presumably due to damage by insects caused by a slight delay in flowering time. However, yields of hybrids that were homozygous or heterozygous for Brrga1-d were similar to those of near isogenic wild-type hybrids. In addition, we report on a ‘perfect’ PCR marker for selection of the Brrga1-d gene that is based on detecting the nucleotide mutation causing the dwarf phenotype.
DOI:10.3969/j.issn.1000-6850.2013.13.032URL [本文引用: 4]
Dwarfism in oilseed rape is one of the main strategies to improve lodging resistance and be suitable for the mechanized operation. A dwarf mutation bndf-1with plant height of about 75cm discovered from hybrid offspring showed obviously decreased height and increased numbers of primary branches in comparison with the wild type. The mutant also showed other values such as compact construct, strong stem stalk and as well as much more primary branches than the wild type. Genetic analysis on F1 and F2 populations derived from reciprocal crosses between bndf-1 with high-stalk lines 0003 and 0826, respectively, indicated that the dwarfism was controlled by an incomplete dominant gene. In addition, the analysis of F1 yield traits indicated that the dwarfism of bndf-1 has great potential to be used in breeding.
DOI:10.3969/j.issn.1000-6850.2013.13.032URL [本文引用: 4]
Dwarfism in oilseed rape is one of the main strategies to improve lodging resistance and be suitable for the mechanized operation. A dwarf mutation bndf-1with plant height of about 75cm discovered from hybrid offspring showed obviously decreased height and increased numbers of primary branches in comparison with the wild type. The mutant also showed other values such as compact construct, strong stem stalk and as well as much more primary branches than the wild type. Genetic analysis on F1 and F2 populations derived from reciprocal crosses between bndf-1 with high-stalk lines 0003 and 0826, respectively, indicated that the dwarfism was controlled by an incomplete dominant gene. In addition, the analysis of F1 yield traits indicated that the dwarfism of bndf-1 has great potential to be used in breeding.
URL [本文引用: 1]
甘蓝型油菜矮秆突变体DS-1和DS-2甘蓝型油菜矮秆突变体DS-1和DS-2产生于离体诱变的甘蓝型油菜小孢子胚状体的胚性培养物。1993年3月,采用NLN液体培养基(Lichter,1982)对生长于大田的甘蓝型油菜品系92-B10植株的单核期小孢子...
URL [本文引用: 1]
甘蓝型油菜矮秆突变体DS-1和DS-2甘蓝型油菜矮秆突变体DS-1和DS-2产生于离体诱变的甘蓝型油菜小孢子胚状体的胚性培养物。1993年3月,采用NLN液体培养基(Lichter,1982)对生长于大田的甘蓝型油菜品系92-B10植株的单核期小孢子...
[D].
[本文引用: 4]
[D].
[本文引用: 4]
DOI:10.1007/s00122-010-1306-9URL [本文引用: 1]
http://link.springer.com/article/10.1007%2Fs00122-010-1306-9
DOI:10.1007/s00122-016-2846-4URLPMID:28093630 [本文引用: 3]
Abstract KEY MESSAGE: Identification and characterization of a semi-dwarfing gene ds-3 encoding a mutant DELLA protein regulating plant height through gibberellin signaling pathway. Lodging is one of the most important factors causing severe yield loss in oilseed rape. Utilization of semi-dwarf varieties has been proved the most effective way to increase lodging resistance and yield in many crops. To develop semi-dwarf germplasm in oilseed rape, we identified a semi-dwarf mutant ds-3 which showed a reduced response to phytohormones gibberellins (GAs). Genetic analysis indicated the dwarfism was controlled by a single semi-dominant gene, ds-3. The DS-3 gene was mapped to a genomic region on chromosome C07, which is syntenic to the region of a previously identified semi-dwarf gene ds-1 (BnaA06.RGA). In this region, DS-3 (BnaC07.RGA) gene was identified to encode a DELLA protein that functions as a repressor in GA signaling pathway. A substitution of proline to leucine was identified in ds-3 in the conserved VHYNP motif, which is essential for GA-dependent interaction between gibberellin receptor GID1 and DELLA proteins. Segregation analysis in the F 2 population derived from the cross between ds-1 and ds-3 demonstrated that BnaA06.RGA displayed a stronger effect on plant height than BnaC07.RGA, indicating that different RGA genes may play different roles in stem elongation. In addition to BnaA06.RGA and BnaC07.RGA, two more RGA genes (BnaA09.RGA and BnaC09.RGA) were identified in the Brassica napus (B. napus) genome. Reverse-transcription polymerase chain reaction (RT-PCR) and yeast two-hybrid (Y2H) assays suggest that both BnaA09.RGA and BnaC09.RGA are transcribed in leaves and stems and can mediate GA signaling in vivo. These genes represent potential targets for screening ideal semi-dwarfing alleles for oilseed rape breeding.
DOI:10.1007/BF00230119URLPMID:24162475 [本文引用: 1]
We have undertaken the construction of a Brassica napus genetic map with isozyme (4%), RFLP (26.5%) and RAPD (68%) markers on a 152 lines of a doubled-haploid population. The map covers 1765 cM and comprises 254 markers including three PCR-specific markers and a morphological marker. They are assembled into 19 linkage groups, covering approximatively 71% of the rapeseed genome. Thirty five percent of the studied markers did not segregate according to the expected Mendelian ratio and tended to cluster in eight specific linkage groups. In this paper, the structure of the genetic map is described and the existence of non-Mendelian segregations in linkage analysis as well as the origins of the observed distortions, are discussed. The mapped RFLP loci corresponded to the cDNAs already used to construct B. napus maps. The first results of intraspecific comparative mapping are presented.
[本文引用: 1]
DOI:10.2135/cropsci2015.09.0554URL [本文引用: 1]
[本文引用: 4]
DOI:10.1007/s10722-010-9541-0URL [本文引用: 3]
A novel dwarf mutant of Brassica napus L. em. Metzg., named NDF - 1 , was derived from a high doubled haploid line ‘3529’ of which seeds were jointly treated with chemical inducers and fast neutron bombardment. The germination results showed that the germination of NDF - 1 was insensitive in response to exogenous gibberellic acid 3 (GA 3 ). The studies on growth response to exogenous GA 3 showed that NDF - 1 seeding has at least 10-fold insensitivity than the wild-type. Moreover, no matter what concentrations of GA 3 were added to the seedlings and adult plants, the NDF - 1 could not restore the wild type phenotype. These results indicated that the B. napus dwarf mutant NDF - 1 was GA-insensitive mutant. The histological observations showed that the key reason of leading NDF - 1 to dwarf was the reduction of hypocotyls and stems cell numbers.
DOI:10.1007/s11033-010-0094-2URLPMID:20358292 [本文引用: 1]
A dwarf mutant from Brassica napus , namely NDF-1 , which was derived from a high doubled haploid (DH) line ‘3529’( Brassica napus L.) of which seeds were jointly treated with chemical inducers and fast neutron bombardment, was revealed that dwarfism is under the control of a major gene(designated as ndf1 ) with a mainly additive effect and non-significant dominance effect. The germination and hypocotyls elongation response of dwarf mutants after exogenous GA and uniconazol application showed NDF-1 was a gibberellin insensitive dwarf. We cloned the Brassica napus GID1 gene, named BnGID1 , and found it was the ortholog of AtGID1a . The sequence blasting of the BnGID1 genes from NDF-1 and wild type showed there was no mutant in the gene. But the quantitative RT-PCR analysis of GID1 EST pointed out the mutation was caused by the low-level expression of BnGID1 gene. After sequenced the BnGID1 gene’s upstream, we found three bases mutated in the pyrimidine box (P-box) of the BnGID1 promoter, which is linkage with the dwarf mutant.
DOI:10.3969/j.issn.1001-4829.2006.03.007URL [本文引用: 1]
通过对“神舟四号”飞船、第十 八颗返回式卫星搭载甘蓝型油菜种子及其后代的初步研究,结果表明:12个搭载品系种子的出苗率、存苗率、结实率和植株性状在当代(SP1)与对照相比均没 有明显变化;但在SP2代,通过田间观测、室内考种和品质测试,发现出长果、多果、早花、矮杆、大粒等有利变异株,多头茎、主茎短缩等畸形变异株,还有一 些芥酸、硫甙、含油率、蛋白质含量发生改变的变异群体。从后代中有可能选择出有利于育种目标的变异单株或群体。
DOI:10.3969/j.issn.1001-4829.2006.03.007URL [本文引用: 1]
通过对“神舟四号”飞船、第十 八颗返回式卫星搭载甘蓝型油菜种子及其后代的初步研究,结果表明:12个搭载品系种子的出苗率、存苗率、结实率和植株性状在当代(SP1)与对照相比均没 有明显变化;但在SP2代,通过田间观测、室内考种和品质测试,发现出长果、多果、早花、矮杆、大粒等有利变异株,多头茎、主茎短缩等畸形变异株,还有一 些芥酸、硫甙、含油率、蛋白质含量发生改变的变异群体。从后代中有可能选择出有利于育种目标的变异单株或群体。
DOI:10.3321/j.issn:1007-9084.2003.04.003URL [本文引用: 1]
为了拓宽甘蓝型油菜的遗传基础,从四川、重庆、云南和西藏部分地区采(收)集了一批白菜型油菜、白菜和白花芥蓝地方品种,包括一部分白菜型油菜野生种和半野生种,通过远缘杂交,人工合成了一批甘蓝型油菜新材料.这批新材料具有丰富的遗传变异,主要包括特长角、大粒、多分枝、矮秆、黄籽、白花、抗病等重要遗传类型,部分材料具有重要的育种及研究利用价值.
DOI:10.3321/j.issn:1007-9084.2003.04.003URL [本文引用: 1]
为了拓宽甘蓝型油菜的遗传基础,从四川、重庆、云南和西藏部分地区采(收)集了一批白菜型油菜、白菜和白花芥蓝地方品种,包括一部分白菜型油菜野生种和半野生种,通过远缘杂交,人工合成了一批甘蓝型油菜新材料.这批新材料具有丰富的遗传变异,主要包括特长角、大粒、多分枝、矮秆、黄籽、白花、抗病等重要遗传类型,部分材料具有重要的育种及研究利用价值.
DOI:10.3969/j.issn.1006-060X.2007.05.015URL [本文引用: 1]
介绍了油菜矮杆抗倒新品系的来源、特征特性、遗传组成和利用价值。该品系来自于油菜芥甘种间 杂交后代,具有矮杆抗倒、早熟高产高油分、超低硫苷、较高油酸含量等优良特性。随机扩增多态性DNA标记分析表明,其具有10%以上的芥菜型油菜遗传物 质。该品系有望直接作为新品种推广应用,也可作为杂交亲本转育成矮杆抗倒品种或杂种亲本,从而开创矮杆油菜新时代,推动油菜机械化生产向前发展。
DOI:10.3969/j.issn.1006-060X.2007.05.015URL [本文引用: 1]
介绍了油菜矮杆抗倒新品系的来源、特征特性、遗传组成和利用价值。该品系来自于油菜芥甘种间 杂交后代,具有矮杆抗倒、早熟高产高油分、超低硫苷、较高油酸含量等优良特性。随机扩增多态性DNA标记分析表明,其具有10%以上的芥菜型油菜遗传物 质。该品系有望直接作为新品种推广应用,也可作为杂交亲本转育成矮杆抗倒品种或杂种亲本,从而开创矮杆油菜新时代,推动油菜机械化生产向前发展。
DOI:10.3724/SP.J.1006.2013.00216URLMagsci [本文引用: 2]
<div ><span >株型改良是油菜高产、优质育种的主攻方向之一。矮杆及半矮杆株型有利于提高植株抗倒伏能力和经济系数、减少收获难度。</span><span >10D130</span><span >是一个半矮杆新品系</span><span >, </span><span >用</span><span >10D130</span><span >和常规优良品种中双</span><span >11</span><span >杂交</span><span >, </span><span >构建</span><span >6</span><span >世代遗传群体</span><span >(P<sub>1</sub></span><span >、</span><span >F<sub>1</sub></span><span >、</span><span >P<sub>2</sub></span><span >、</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub>), </span><span >以主基因</span><span >+</span><span >多基因混合遗传模型对该组合株高及其关联性状进行遗传分析。结果表明</span><span >, 10D130×</span><span >中双</span><span >11</span><span >组合株高、分枝部位、主花序长度的遗传均受到</span><span >1</span><span >对加性</span><span >-</span><span >显性</span><span >-</span><span >上位性主基因</span><span >+</span><span >加性</span><span >-</span><span >显性</span><span >-</span><span >上位性多基因控制</span><span >(D-0</span><span >模型</span><span >)</span><span >。其中</span><span >, </span><span >株高性状加性效应值为</span><span >–8.58, </span><span >显性效应值为</span><span >7.44, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >中分别为</span><span >23.52%</span><span >、</span><span >0.91%</span><span >和</span><span >17.81%</span><span >;一</span><span >次有效分枝起始部位的</span><span >1</span><span >对主基因加性效应值为</span><span >–22.11, </span><span >显性效应值为</span><span >3.13, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >中分别为</span><span >49.95%</span><span >、</span><span >40.85%</span><span >和</span><span >61.15%</span><span >;主花序长的主基因加性效应值为</span><span >–2.21, </span><span >显性效应值为</span><span >1.6, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >中分别为</span><span >0.68%</span><span >、</span><span >47.94%</span><span >和</span><span >40.07%</span><span >。一次有效分枝间距的最适宜遗传模型为</span><span >E-1</span><span >模型</span><span >(2</span><span >对加</span><span >-</span><span >显</span><span >-</span><span >上位性主基因</span><span >+</span><span >加</span><span >-</span><span >显</span><span >-</span><span >上位性多基因混合遗传模型</span><span >), </span><span >其中第</span><span >1</span><span >对主基因加性效应值为</span><span >–0.55</span><span >、显性效应值为</span><span >–1.66, </span><span >第</span><span >2</span><span >对主基因加性效应值为</span><span >0.74</span><span >、显性效应值为</span><span >–1.29, </span><span >均表现超显性遗传</span><span >, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >三个分离世代群体中分别为</span><span >10.99%</span><span >、</span><span >38.65%</span><span >和</span><span >44.10%</span><span >。一次有效分枝部位高度、主花序长、有效分枝节间距和有效分枝数与株高均呈显著正相关。</span></div>
DOI:10.3724/SP.J.1006.2013.00216URLMagsci [本文引用: 2]
<div ><span >株型改良是油菜高产、优质育种的主攻方向之一。矮杆及半矮杆株型有利于提高植株抗倒伏能力和经济系数、减少收获难度。</span><span >10D130</span><span >是一个半矮杆新品系</span><span >, </span><span >用</span><span >10D130</span><span >和常规优良品种中双</span><span >11</span><span >杂交</span><span >, </span><span >构建</span><span >6</span><span >世代遗传群体</span><span >(P<sub>1</sub></span><span >、</span><span >F<sub>1</sub></span><span >、</span><span >P<sub>2</sub></span><span >、</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub>), </span><span >以主基因</span><span >+</span><span >多基因混合遗传模型对该组合株高及其关联性状进行遗传分析。结果表明</span><span >, 10D130×</span><span >中双</span><span >11</span><span >组合株高、分枝部位、主花序长度的遗传均受到</span><span >1</span><span >对加性</span><span >-</span><span >显性</span><span >-</span><span >上位性主基因</span><span >+</span><span >加性</span><span >-</span><span >显性</span><span >-</span><span >上位性多基因控制</span><span >(D-0</span><span >模型</span><span >)</span><span >。其中</span><span >, </span><span >株高性状加性效应值为</span><span >–8.58, </span><span >显性效应值为</span><span >7.44, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >中分别为</span><span >23.52%</span><span >、</span><span >0.91%</span><span >和</span><span >17.81%</span><span >;一</span><span >次有效分枝起始部位的</span><span >1</span><span >对主基因加性效应值为</span><span >–22.11, </span><span >显性效应值为</span><span >3.13, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >中分别为</span><span >49.95%</span><span >、</span><span >40.85%</span><span >和</span><span >61.15%</span><span >;主花序长的主基因加性效应值为</span><span >–2.21, </span><span >显性效应值为</span><span >1.6, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >中分别为</span><span >0.68%</span><span >、</span><span >47.94%</span><span >和</span><span >40.07%</span><span >。一次有效分枝间距的最适宜遗传模型为</span><span >E-1</span><span >模型</span><span >(2</span><span >对加</span><span >-</span><span >显</span><span >-</span><span >上位性主基因</span><span >+</span><span >加</span><span >-</span><span >显</span><span >-</span><span >上位性多基因混合遗传模型</span><span >), </span><span >其中第</span><span >1</span><span >对主基因加性效应值为</span><span >–0.55</span><span >、显性效应值为</span><span >–1.66, </span><span >第</span><span >2</span><span >对主基因加性效应值为</span><span >0.74</span><span >、显性效应值为</span><span >–1.29, </span><span >均表现超显性遗传</span><span >, </span><span >主基因遗传率在</span><span >B<sub>1</sub></span><span >、</span><span >B<sub>2</sub></span><span >和</span><span >F<sub>2</sub></span><span >三个分离世代群体中分别为</span><span >10.99%</span><span >、</span><span >38.65%</span><span >和</span><span >44.10%</span><span >。一次有效分枝部位高度、主花序长、有效分枝节间距和有效分枝数与株高均呈显著正相关。</span></div>
DOI:10.3969/j.issn.0490-6756.2007.05.045URL [本文引用: 1]
以甘蓝型油菜矮化突变体"DDF-1"为材料,研究了赤霉素(GA3)、生长素(IAA)和油菜素内酯(BR)3种激素对突变体幼苗的影响,并用石蜡切片观察其细胞形态,用酶联免疫吸附技术(ELISA)检测下胚轴内源激素GA3含量.结果表明,外加一定浓度的GA3(7 mg/L)对矮化突变体"DDF-1"的下胚轴伸长作用显著,但不能使之恢复到野生型的长度;萌发早期,BR有明显的促进伸长作用,萌发后期效果不明显;突变体对IAA敏感性较弱.三种激素交互作用对矮秆下胚轴伸长的影响也不显著.施加外源GA3的矮化突变体"DDF-1"的细胞也有明显的伸长.矮化突变体"DDF-1"的内源激素GA3含量高于野生型高秆,但低于施加外源GA3的矮化突变体.矮化突变体"DDF-1"为赤霉素GA3的非应答型矮化突变体.
DOI:10.3969/j.issn.0490-6756.2007.05.045URL [本文引用: 1]
以甘蓝型油菜矮化突变体"DDF-1"为材料,研究了赤霉素(GA3)、生长素(IAA)和油菜素内酯(BR)3种激素对突变体幼苗的影响,并用石蜡切片观察其细胞形态,用酶联免疫吸附技术(ELISA)检测下胚轴内源激素GA3含量.结果表明,外加一定浓度的GA3(7 mg/L)对矮化突变体"DDF-1"的下胚轴伸长作用显著,但不能使之恢复到野生型的长度;萌发早期,BR有明显的促进伸长作用,萌发后期效果不明显;突变体对IAA敏感性较弱.三种激素交互作用对矮秆下胚轴伸长的影响也不显著.施加外源GA3的矮化突变体"DDF-1"的细胞也有明显的伸长.矮化突变体"DDF-1"的内源激素GA3含量高于野生型高秆,但低于施加外源GA3的矮化突变体.矮化突变体"DDF-1"为赤霉素GA3的非应答型矮化突变体.
[本文引用: 1]
[本文引用: 1]
DOI:10.7685/j.issn.1000-2030.2013.06.001 [本文引用: 1]
近十几年来,利用双亲分离群体数量性状表型观测值来鉴定数量性状主基因+多基因混合遗传模式的研究时有报道,其结果可指导作物育种实践。然而,随着计算机技术的进步,本课题组研制的软件操作性、算法和功能模块等方面都存在一定缺陷。在Microsoft Visual Studio 2010操作平台下,利用C++语言研制了Windows界面友好的植物数量性状分离分析(segregation analysis,SEA)软件包,其中采用了线性算法包Clapack V3.1.1解线性方程组并估计一阶遗传参数,更精确地计算适合性检验的概率,增加了结果输出模块和计算分离群体植株或家系的主基因型后验概率。新软件包计算更稳定,结果更全面,界面更友好。
DOI:10.7685/j.issn.1000-2030.2013.06.001 [本文引用: 1]
近十几年来,利用双亲分离群体数量性状表型观测值来鉴定数量性状主基因+多基因混合遗传模式的研究时有报道,其结果可指导作物育种实践。然而,随着计算机技术的进步,本课题组研制的软件操作性、算法和功能模块等方面都存在一定缺陷。在Microsoft Visual Studio 2010操作平台下,利用C++语言研制了Windows界面友好的植物数量性状分离分析(segregation analysis,SEA)软件包,其中采用了线性算法包Clapack V3.1.1解线性方程组并估计一阶遗传参数,更精确地计算适合性检验的概率,增加了结果输出模块和计算分离群体植株或家系的主基因型后验概率。新软件包计算更稳定,结果更全面,界面更友好。
DOI:10.2307/3869351URLPMID:12324600 [本文引用: 1]
The greening phenotypes produced by recessive mutations in a gene designated de-etiolated-2 (DET2) are described. Recessive mutations in the DET2 gene uncouple light signals from a number of light-dependent processes. det2 mutations result in dark-grown Arabidopsis thaliana seedlings with many characteristics of light-grown plants, including hypocotyl growth inhibition, cotyledon expansion, primary leaf initiation, anthocyanin accumulation, and derepression of light-regulated gene expression. In contrast to these morphological and gene expression changes, however, the chloroplast development program is not initiated in the dark in det2 mutants, suggesting that light-regulated gene expression precedes the differentiation of etioplasts to chloroplasts. det2 mutations thus reveal at least two classes of downstream light-regulated responses that differ in their timing and control mechanisms. Homozygous det2 mutations also affect photoperiodic responses in light-grown plants, including timing of flowering, dark adaptation of gene expression, and onset of leaf senescence. The phenotype of det1 det2 double mutants is additive, implying that DET1 and DET2 function in distinct pathways that affect downstream light-regulated genes. Furthermore, these pathways are not utilized solely during early seedling development but must also be required to regulate different aspects of the light developmental program during later stages of vegetative growth.
[本文引用: 1]
DOI:10.1016/S0092-8674(00)81094-6URLPMID:8612270 [本文引用: 1]
The cpd mutation localized by T-DNA tagging on Arabidopsis chromosome 5-14.3 inhibits cell elongation controlled by the ecdysone-like brassinosteroid hormone brassinolide. The cpd mutant displays de-etiolation and derepression of light-induced genes in the dark, as well as dwarfism, male sterility, and activation of stress-regulated genes in the light. The CPD gene encodes a cytochrome P450 (CYP90) sharing homologous domains with steroid hydroxylases. The phenotype of the cpd mutant is restored to wild type both by feeding with C23-hydroxylated brassinolide precursors and by ectopic overexpression of the CPD cDNA. Brassinosteroids also compensate for different cell elongation defects of Arabidopsis det, cop, fus, and axr2 mutants, indicating that these steroids play an essential role in the regulation of plant development.
DOI:10.1093/dnares/9.1.11URLPMID:11939564 [本文引用: 1]
A rice semidwarfing gene, sd-1, known as the green revolution gene, was isolated by positional cloning and revealed to encode gibberellin 20-oxidase, the key enzyme in the gibberellin biosynthesis pathway. Analysis of 3477 segregants using several PCR-based marker technologies, including cleaved amplified polymorphic sequence, derived-CAPS, and single nucleotide polymorphisms revealed 1 ORF in a 6-kb candidate interval. Normal-type rice cultivars have an identical sequence in this region, consisting of 3 exons (558, 318, and 291 bp) and 2 introns (105 and 1471 bp). Dee-Geo-Woo-Gen-type sd-1 mutants have a 383-bp deletion from the genome (278-bp deletion from the expressed sequence), from the middle of exon 1 to upstream of exon 2, including a 105-bp intron, resulting in a frame-shift that produces a termination codon after the deletion site. The radiation-induced sd-1 mutant Calrose 76 has a 1-bp substitution in exon 2, causing an amino acid substitution (Leu [CTC] to Phe [TTC]). Expression analysis suggests the existence of at least one more locus of gibberellin 20-oxidase which may prevent severe dwarfism from developing in sd-1 mutants.
DOI:10.1007/s10681-014-1275-1URL [本文引用: 1]
Plant height and spikelet density are important agronomic traits related to plant architecture and grain yield in wheat. The development of wheat mutants not only provided new genetic resources for wheat improvement, but also facilitated our understanding of the regulation of these traits at the molecular level. Here, we report the identification of a dwarf mutant with a compact spike. NAUH164, produced from ethyl methyl sulfonate treatment of wheat variety Sumai 3, has reduced plant height and shortened spike length. Microscopic observations showed that the mutant has an atypical vascular bundle arrangement and overall smaller cell volumes. Sensitivity of seedlings to exogenous gibberellin was not changed, and plant height was not restored to the level of the wildtype. Dwarfness and compact spike were controlled by a single dominant gene that was designated Rht23 , which was mapped 4.7 cM distal to SSR marker Gdm63 and 11.1 cM proximal to Barc110 on chromosome 5DL. Comparative sequence analysis of the co-linear regions in common wheat and Aegilops taushii indicated that Rht23 occurs in a physical interval of 8.5 Mb.
DOI:10.1104/pp.111.183657URL [本文引用: 1]
The introduction of the Reduced height (Rht)-B1b and Rht-D1b semidwarfing genes led to impressive increases in wheat (Triticum aestivum) yields during the Green Revolution. The reduction in stem elongation in varieties containing these alleles is caused by a limited response to the phytohormone gibberellin (GA), resulting in improved resistance to stem lodging and yield benefits through an increase in grain number. Rht-B1 and Rht-D1 encode DELLA proteins, which act to repress GA-responsive growth, and their mutant alleles Rht-B1b and Rht-D1b are thought to confer dwarfism by producing more active forms of these growth repressors. While no semidwarfing alleles of Rht-A1 have been identified, we show that this gene is expressed at comparable levels to the other homeologs and represents a potential target for producing novel dwarfing alleles. In this study, we have characterized additional dwarfing mutations in Rht-B1 and Rht-D1. We show that the severe dwarfism conferred by Rht-B1c is caused by an intragenic insertion, which results in an in-frame 90-bp insertion in the transcript and a predicted 30-amino acid insertion within the highly conserved amino-terminal DELLA domain. In contrast, the extreme dwarfism of Rht-D1c is due to overexpression of the semidwarfing Rht-D1b allele, caused by an increase in gene copy number. We show also that the semidwarfing alleles Rht-B1d and Rht-B1e introduce premature stop codons within the amino-terminal coding region. Yeast two-hybrid assays indicate that these newly characterized mutations in Rht-B1 and Rht-D1 confer "GA-insensitive" dwarfism by producing DELLA proteins that do not bind the GA receptor GA INSENSITIVE DWARF1, potentially compromising their targeted degradation.
DOI:10.3969/j.issn.1001-8581.2009.05.009URL [本文引用: 1]
总结了适合于全程机械化作业的镇油5号的育种实践和育成应用;结合镇油5号在生产中适合机械化作业的表现,探讨了适合全程机械化作业的油菜新品种育种的途径和技术以及在推广实践中面临的问题。
DOI:10.3969/j.issn.1001-8581.2009.05.009URL [本文引用: 1]
总结了适合于全程机械化作业的镇油5号的育种实践和育成应用;结合镇油5号在生产中适合机械化作业的表现,探讨了适合全程机械化作业的油菜新品种育种的途径和技术以及在推广实践中面临的问题。
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1104/pp.105.062968URLPMID:16040652 [本文引用: 1]
Molecular genetic studies of plant dwarf mutants have indicated that gibberellin (GA) and brassinosteroid (BR) are two major factors that determine plant height; dwarf mutants that are caused by other defects are relatively rare, especially in monocot species. Here, we report a rice (Oryza sativa) dwarf mutant, dwarf and gladius leaf1 (dgl1), which exhibits only minimal response to GA and BR. In addition to the dwarf phenotype, dgl1 produces leaves with abnormally rounded tip regions. Positional cloning of DGL1 revealed that it encodes a 60-kD microtubule-severing katanin-like protein. The protein was found to be important in cell elongation and division, based on the observed cell phenotypes. GA biosynthetic genes are up-regulated in dgl1, but the expression of BR biosynthetic genes is not enhanced. The enhanced expression of GA biosynthetic genes in dgl1 is not caused by inappropriate GA signaling because the expression of these genes was repressed by GA3treatment, and degradation of the rice DELLA protein SLR1 was triggered by GA3in this mutant. Instead, aberrant microtubule organization caused by the loss of the microtubule-severing function of DGL1 may result in enhanced expression of GA biosynthetic genes in that enhanced expression was also observed in a BR-deficient mutant with aberrant microtubule organization. These results suggest that the function of DGL1 is important for cell and organ elongation in rice, and aberrant DGL1-mediated microtubule organization causes up-regulation of gibberellin biosynthetic genes independently of gibberellin signaling.
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1007/BF02279323URL [本文引用: 1]
Eight rapid-cycling Brassica genotypes differing in height were treated with gibberellins (GAs) by syringe application to the shoot tip. The height of two genotypes of Brassica napus , Bn5-2 and Bn5-8, and B. rapa mutants, dwarf 1 (dwf1) and dwarf 2 (dwf2) , was unaffected by exogenous GA 3 at dosages up to 0.1 μg/plant, a level which increased shoot elongation of normal genotypes. Thus, these dwarf mutants are “GA-insensitive.” In contrast to the B. napus dwarfs, two B. rapa mutants, rosette (ros) , and dormant (dor) , elongated following GA 3 application. The dwarf ros was most sensitive, responding to applications as low as 1 ng GA 3 /plant. Furthermore, ros also responded to GA 1 and some of its precursors with decreasing efficacy: GA 3 > ent -kaurenoic acid ≥GA 1 >GA 20 ≥GA 19 =GA 44 ≥GA 53 . Endogenous GAs were measured by gas chromatography-selected ion monitoring using [ 2 H 2 ]GA internal standards for calibration, from shoots of the GA-insensitive genotypes Bn5-2, Bn5-8 which contained the B. napus mutant dwarf 1 , and from a normal genotype Bn5-1. Concentrations of GA 1 and GA 20 averaged 3.2- and 4.6-fold higher, respectively, and GA 19 levels also tended to be higher in the dwarfs than in the normal genotype.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}