CEBPα and p53 Regulate Kiss1 Gene Expression in Porcine Ovary Granulosa Cells
XIN XiaoPing1, WANG JiaYing1, ZHANG AiLing2, ZHONG YuYi1, HE YingTing1, CHEN ZanMou1, ZHANG Zhe1, ZHANG Hao1, LI JiaQi1, YUAN XiaoLong,11 Guangdong Provincial Key Lab of Agro-animal Genomics and Molecular Breeding/National Engineering Research Center for Breeding Swine Industry/College of Animal Science, South China Agricultural University, Guangzhou 510642 2 Development Center of Applied Ecology and Ecological Engineering in Universities/Biology and Food Engineering Institute, Guangdong University of Education, Guangzhou 510310
Received:2018-04-18Accepted:2019-03-18Online:2019-05-01 作者简介 About authors 辛晓萍,Tel:13602262891;E-mail: xiaopingxin1991@163.com。
摘要 【目的】预测母猪Kiss1(GenBank Gene ID: 100145896)上游区域潜在的转录因子结合位点,并验证部分转录因子在猪卵巢颗粒细胞中对Kiss1基因的调控作用,为研究Kiss1基因在母猪卵巢颗粒细胞中的分子调控机制提供理论基础。【方法】参考NCBI数据库中Kiss1基因上游区域的序列,通过生物信息学网站预测的Kiss1基因上游区域潜在转录因子结合的位点,结合文献阅读与资料查询,在转录因子CEBPα和p53与Kiss1基因上游区域潜在结合位点附近设计引物,利用染色质免疫共沉淀(ChIP)技术,验证转录因子CEBPα和p53与Kiss1基因上游区域的结合情况;参照NCBI数据库相关转录因子CEBPα和p53的mRNA序列,使用软件Primer Premier 5设计引物,PCR分别扩增转录因子CEBPα(带有Kpn I和Xho I限制性内切酶的酶切位点)和p53(带有Kpn I和Hind III限制性内切酶的酶切位点)的CDS区序列,并进行测序鉴定,连接到真核表达载体pcDNA3.1(+)上,构建含有潜在转录因子CEBPα和p53 CDS区序列的真核表达载体,并获得无内毒素质粒,分别命名为pcDNA3.1-CEBPα和pcDNA3.1-p53;化学合成目标转录因子CEBPα和p53的干扰siRNA片段。屠宰场采集猪的卵巢,快速分离并培养原代猪的卵巢颗粒细胞,阳离子脂质体转染法分别将转录因子CEBPα和p53真核表达载体(pcDNA3.1-CEBPα和pcDNA3.1-p53)或siRNA片段(CEBPα-siRNA和p53-siRNA)转染进母猪卵巢颗粒细胞,使用实时荧光定量PCR和Western Blot技术分别验证预测的转录因子CEBPα和p53对Kiss1基因mRNA水平和蛋白水平的影响。【结果】生物信息学预测结果表明,Kiss1基因上游区域(-850—+221)存在p53(肿瘤抑制蛋白,tumor protein p53,p53)、CCAAT增强子结合蛋白(CCAAT/enhancer binding protein, CEBP)、信号传导及转录活化家族Stat4(signal transducer and activator of transcription 4, Stat4)等转录因子的潜在结合位点,其中转录因子p53(GenBank Gene ID:397276,NM_213824.3)可能结合在Kiss1基因上游区域-533—-523 bp处,转录因子CEBPα(GenBank Gene ID:397307,XM_003127015.4)可能结合在Kiss1基因上游区域-744—-733 bp处;ChIP结果表明,转录因子p53和CEBPα可以特异性的结合在Kiss1基因上游区域-533—-523 bp和-744—-733 bp处;在母猪卵巢颗粒细胞中超表达转录因子p53或CEBPα后,Kiss1基因的mRNA的表达水平显著下降(P<0.05),蛋白水平显著下降(P<0.05);在母猪卵巢颗粒细胞中,干扰转录因子p53或CEBPα后,Kiss1基因的mRNA的表达水平显著升高(P<0.05),蛋白水平显著升高(P<0.05)。【结论】在猪卵巢颗粒细胞里,转录因子p53和CEBPα能结合到Kiss1基因的上游区域降低其启动子转录活性,进而降低其mRNA和蛋白表达水平。 关键词:猪;卵巢颗粒细胞;Kiss1基因;CEBPα;p53
Abstract 【Objective】The objectives of this study were to predict the potential transcription factors in the upstream region of Kiss1, and then to verify the regulatory role of these transcription factors on the expression of Kiss1 in the ovarian granulosa cells of pigs. This study could provide the basic data for the molecular mechanism of Kiss1 in granulosa cells of pigs. 【Method】 By using the bioinformatic software, we predicted the potential binding site of transcription factors in the upstream region of Kiss1 in NCBI database. In addition, we reviewed a lot of relevant papers and references, and then CEBPα and p53 were selected as potential binding transcriptional factors in the upstream region of Kiss1 gene. The primers were designed near the potential binding sites in the upstream region of Kiss1 genes, and verified the binding of transcriptional factors CEBPα and p53 to the upstream region of Kiss1 gene by ChIP. According to the mRNA sequences of CEBPα and p53 in NCBI database, Primer Premier 5 software was used to design primers, and CDS regions of CEBPα (containing KpnI and XhoI sites) and p53 (containing KpnI and Hind III sites) were amplified by PCR and identified by sequencing. Then the CDS regions were connected to the eukaryotic expression vector pcDNA3.1, and the constructed plasmids were extracted with endotoxin-freely, named pcDNA3.1-CEBPα and pcDNA3.1-p53. The interfering siRNA fragments of CEBPα and p53 were synthesized through the chemical method. Pig ovaries were collected from the slaughterhouse, and the ovarian granulosa cells were isolated and cultured. The eukaryotic expression vector or siRNA fragment was transfected into ovarian granulosa cells by cationic liposome, and the effects of p53 and CEBPα on the expression of Kiss1 were verified by the real-time fluorescence quantitative PCR and Western Blotting, respectively. 【Result】 The bioinformatic prediction indicated that there were putative binding sites of p53 (tumor protein p53, p53), CEBP (CCAAT/enhancer binding protein, CEBP), Stat4 (signal transducer and activator of transcription 4, Stat4) and other potential of transcription factor binding sites in the upstream (-850 -+221) of Kiss1. And the putative binding site of CEBPα (GenBank Gene ID: 397307, XM_003127015.4) were found in -744—-733 bp region and the putative binding site of p53 (GenBank Gene ID: 397276, NM_213824.3) were found in -533—-523 bp region. The results of ChIP showed that p53 and CEBPα were bounded at -533—-523 and -744—-733 of Kiss1, respectively. After overexpression of p53 or CEBPα, both mRNA and protein expression level of Kiss1 significantly decreased (P<0.05). Furthermore, both mRNA and the expression level of Kiss1 increased significantly (P<0.05) by interfering p53 or CEBPα. 【Conclusion】 In pigs, p53 and CEBPα could bind at the upstream region of Kiss1 to inhibit its expression in ovarian granulosa cells. Keywords:pig;ovarian granulosa cells;Kiss1;CEBPα;p53
PDF (644KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 辛晓萍, 王家迎, 张爱玲, 钟玉宜, 何颖婷, 陈赞谋, 张哲, 张豪, 李加琪, 袁晓龙. 转录因子CEBPα和p53在猪卵巢颗粒细胞中 对Kiss1基因表达的调控[J]. 中国农业科学, 2019, 52(9): 1624-1634 doi:10.3864/j.issn.0578-1752.2019.09.013 XIN XiaoPing, WANG JiaYing, ZHANG AiLing, ZHONG YuYi, HE YingTing, CHEN ZanMou, ZHANG Zhe, ZHANG Hao, LI JiaQi, YUAN XiaoLong. CEBPα and p53 Regulate Kiss1 Gene Expression in Porcine Ovary Granulosa Cells[J]. Scientia Acricultura Sinica, 2019, 52(9): 1624-1634 doi:10.3864/j.issn.0578-1752.2019.09.013
A. pcDNA3.1-p53真核表达载体的酶切鉴定;B. pcDNA3.1-CEBPα真核表达载体的酶切鉴定 Fig. 1Identified the eukaryotic expression vector of p53 and CEBPα
A. The enzyme digestion of eukaryotic expression vector pcDNA3.1-p53; B. The enzyme digestion of eukaryotic expression vector digestion pcDNA3.1- CEBPα
通过在线生物学软件分析猪Kiss1基因的-850—+221 区域序列,发现该区域存在p53(肿瘤抑制蛋白,tumor protein p53,p53)、CCAAT增强子结合蛋白(CCAAT/enhancer binding protein, CEBP)、信号传导及转录活化家族Stat4(signal transducer and activator of transcription 4, Stat4)等转录因子的潜在结合位点,其中,在-744—-733处存在CEBPα因子潜在结合位点,在-533—-523处存在的p53潜在结合位点(图2-A)。
M:DNA分子量标准(DL5 000 Marker),从上到下依次为:5000,3000,2000,1500,1000,750,500,250和100 bp. IgG:阴性对照;Input and H3:阳性对照。A:猪Kiss1基因上游区域区结合潜在转录因子生物信息学预测结果;B:转录因子p53结合猪Kiss1基因上游区域的ChIP鉴定;C:转录因子CEBPα结合猪Kiss1基因上游区域的ChIP鉴定。其中,IgG:阴性对照;Input和H3:阳性对照 Fig. 2Bioinformatics prediction for the potential binding site of transcription factors in the upstream region of porcine Kiss1, and ChIP results of transcription factors binding to the upstream region of porcine Kiss1 gene
M:DL5 000 DNA marker,from up to bottom:5000, 3000, 2000, 1500, 1000, 750, 500, 250, and 100 bp. IgG: Negative control; Input and H3: Positive control. A: The analysis of the binding sites for the transcription factor of the upstream region of porcine Kiss1 gene. B: The ChIP results for the binding of p53 with the upstream region of porcine Kiss1 gene. C: The ChIP results for the binding of CEBPα with the upstream region of porcine Kiss1 gene
A:超表达转录因子p53后p53的mRNA表达水平;B:超表达转录因子p53对Kiss1基因mRNA表达水平的影响;C、D:超表达转录因子p53对Kiss1基因蛋白表达水平的影响 Fig. 3The effects of over-expressed p53 on the expression of Kiss1
A: Over-expression of p53 on the mRNA level of p53; B: Over-expression of p53 on the mRNA level of Kiss1; C,D: Over-expression of p53 on the protein level of Kiss1
A:干扰p53的表达后p53的mRNA表达水平;B:干扰p53的表达后对Kiss1基因mRNA表达水平的影响;C、D:干扰p53表达后对Kiss1蛋白表达水平的影响 Fig. 4The effects of interfering p53 on the expression of Kiss1
A: siRNAs interference of p53 on the mRNA level of p53; B: siRNAs interference of p53 on the mRNA level of Kiss1; C, D: siRNAs interference of p53 on the protein level of Kiss1
A:超表达转录因子CEBPα后CEBPα的mRNA表达水平;B:超表达转录因子CEBPα对Kiss1基因mRNA表达水平的影响;C、D:超表达转录因子CEBPα对Kiss1基因蛋白表达水平的影响 Fig. 5The effects of over-expressed CEBPα on the expression of Kiss1
A: Over-expression of CEBPα on the mRNA level of CEBPα; B: Over-expression of CEBPα on the mRNA level of Kiss1; C,D: Over-expression of CEBPα on the protein level of Kiss1
A:干扰转录因子CEBPα后CEBPα的mRNA表达水平;B:干扰转录因子CEBPα对Kiss1基因mRNA表达水平的影响;C、D:干扰转录因子CEBPα对Kiss1基因蛋白表达水平的影响 Fig. 6The effects of interfering p53 on the expression of Kiss1
A: siRNAs interference of CEBPα on the mRNA level of CEBPα; B: siRNAs interference of CEBPα on the mRNA level of Kiss1; C, D: siRNAs interference of CEBPα on the protein level Kiss1
LEEJ H, MIELEM E, HICKSD J, PHILILIPSK K, TRENTJ M, WEISSMANB E, WELCHD R . Kiss-1, a novel human malignant melanoma metastasis-suppressor gene Journal of the National Cancer Institute, 1996,88(23):1731-1737. DOI:10.1093/jnci/88.23.1731URL [本文引用: 1]
PAPAOICONOMOUE, MSAOUELP, MAKRIA, DIAMANTI-KANDARAKIS E, KOUTSILIERISM . The role of kisspeptin/Gpr54 in the reproductive system In Vivo, 2011,25(3):343-354. [本文引用: 1]
GAYTANF, GAYTANM, CASTELLANOJ M, ROMEROM, ROAJ, APARICIOB, GARRIDON, SANCHEZ-CRIADOJ E, PELLICERA, FRASERH M, TENA-SEMPEREM . Kiss-1 in the mammalian ovary: Distribution of kisspeptin in human and marmoset and alterations in Kiss-1 Mrna levels in a rat model of ovulatory dysfunction American Journal of Physiology-Endocrinology and Metabolism, 2009,296(3):E520-E531. DOI:10.1152/ajpendo.90895.2008URL [本文引用: 1]
KAILUNH, HONGCUIZ, HSUNMINGC, YANGY, JIEQ . Kisspeptin/Kisspeptin receptor system in the ovary Frontiers in Endocrinology, 2018, doi.org/ 10.3389/fendo.2017.00365. [本文引用: 1]
GAYTANF, GAYTANM, CASTELLANOJM, ROMEROM, ROAJ, APARICIOB, GARRIDON, SANCHEZ-CRIADOJE, MILLARRP, PELLICERA, FRASERHM, TENA-SEMPEREM . Kiss-1 in the mammalian ovary: Distribution of kisspeptin in human and marmoset and alterations in Kiss-1 Mrna levels in a rat model of ovulatory dysfunction American Journal of Physiology-Endocrinology and Metabolism, 2009,296(3):E520-E531. DOI:10.1152/ajpendo.90895.2008URL [本文引用: 2]
GUOM J . Hyperprolactinemia affects the follicles development and ovulation by Kiss1/GPR54 system in mice [D]. Nanchang: Nanchang University, 2016. ( in Chinese) [本文引用: 3]
TOMIKAWAJ, HOMMAT, TAJIMAS, SHIBATAT, INAMOTOY, TAKASEK, INOUEN, OHKURAS, UENOYAMAY, MAEDAK, TSUKAMURAH . Molecular characterization and estrogen regulation of hypothalamic kiss1 gene in the pig Biology of Reproduction, 2010,82(2):313-319. DOI:10.1095/biolreprod.109.079863URL [本文引用: 1]
ZANGM, CHAOL G, ZHANGZ Q, TAIY L, WANGJ, WUY X, YANGG Y . Molecular clone and sequence analysis of porcine kiss1 gene Acta Agriculturae Jiangxi, 2008(11):5-7.(in Chinese) [本文引用: 2]
NAVARROV M, CASTELLANOJ M, FERNANDEZ-FERNANDEZ R, BARREIROM L, ROAJ, SANCHEZ-CRIADOJ E, AGUILAR E, DIEGUEZC, PINILLAL, TENA-SEMPEREM . Developmental and hormonally regulated messenger ribonucleic acid expression of Kiss-1 and its putative receptor, Gpr54, in rat hypothalamus and potent luteinizing hormone-releasing activity of Kiss-1 peptide Endocrinology, 2004,145(10):4565-4574. DOI:10.1210/en.2004-0413URL [本文引用: 1]
LIUP, WANGH F, WANGJ N, LUY F, ZHAOX B, DINGJ T . Localization and abundance of KISS-1 in the sow hypothalamus- pituitary-ovary axis .Journal of Anhui Agricultural Sciences, 2008(15):6324-6327. (in Chinese) [本文引用: 1]
REYNOLDSR M, LOGIEJ J, ROSEWEIRA K, MCKNIGHTA J, MILLARR P . A role for kisspeptins in pregnancy: Facts and speculations Reproduction, 2009,138(1):1-7. DOI:10.1530/REP-09-0026URL [本文引用: 2]
TOPALOGLUA K, TELLOJ A, KOTANL D, YILMAZM B, ERDOGANS, GURBUZF, TEMIZF, MILLARR P, YUKSELB . Inactivating Kiss1 mutation and hypogonadotropic hypogonadism The New England Journal of Medicine, 2012,366(7):629-635. DOI:10.1056/NEJMoa1111184URL [本文引用: 1]
D'ANGLEMONTD T X, FAGGL A, DIXONJ P, DAYK, LEITCHH G, HENDRICKA G, ZAHND, FRANCESCHINII, CARATYA, CARLTONM B, APARICIOS A, COLLEDGEW H . Hypogonadotropic hypogonadism in mice lacking a functional Kiss1 gene Proceedings of the National Academy of Sciences of the United States of America, 2007,104(25):10714-10719. DOI:10.1073/pnas.0704114104URL [本文引用: 1]
YUANX U, ZHANGA L, XIAOG, ZHANGZ, CHENZ M, ZHANGH, LIJ Q . P53 and Nfkb regulate microrna-34C expression in porcine ovarian granulosa cells Journal of Integrative Agriculture, 2016,15(8):1816-1824. DOI:10.1016/S2095-3119(15)61178-9URL [本文引用: 1]
XINX P, YUANX L, CHENX, ZHANGA L, CHENZ M, ZHANGZ, ZHANGH, LIJ Q . The expression patterns of key genes of PI3K signaling pathway in the development of the porcine ovary Journal of South China Agricultral University.2018(02):1-6.(in Chinese) [本文引用: 1]
BILBANM, GHAFFARI-TABRIZIN, HINTERMANNE, BAUERS, MOLZERS, ZORATTIC, MALLIR, SHARABIA, HIDENU, GRAIERW, KN?FLERM, ANDREAEF, WAGNERO, QUARANTAV, DESOYEG . Kisspeptin-10, a Kiss-1/Metastin-Derived decapeptide, is a physiological invasion inhibitor of primary human trophoblasts Journal of Cell Science, 2004,117(8):1319-1328. DOI:10.1242/jcs.00971URL [本文引用: 1]
LAIL D . The effect and mechanism of hyperprolactinemia on endocrine function of rat luteal cells by Kiss1/CPR54 system [D]. Nanchang :Nanchang University, 2016. ( in Chinese) [本文引用: 1]
NAGATAM, TAKENAKAH, SHIBAGAKIR, KISHIMOTOS . Apoptosis and P53 protein expression increase in the process of burn wound healing in Guinea-Pig skin British Journal of Dermatology, 1999,140(5):829-838. DOI:10.1046/j.1365-2133.1999.02811.xURL [本文引用: 1]
ZHANGX, HEY, LEEK, DUBOISW, LIZ Q, WUX L, KOVALCHUK A, ZHANGW M, HUANGJ . Rap2B, a novel P53 target, regulates P53-mediated pro-survival function Cell Cycle, 2013,12(8):1279-1291. DOI:10.4161/cc.24364URL [本文引用: 1]
WANGY, WENGH, ZHANGY, LONGY, LIY, NIUY, SONGF, BUY . The Prr11-Ska2 bidirectional transcription unit is negatively regulated by P53 through Nf-Y in lung cancer cells International Journal of Molecular Sciences, 2017, doi: 10.3390/ijms18030534. [本文引用: 1]
WONGM Y, YUY, WALSHW R, YANGJ L . Microrna-34 family and treatment of cancers with mutant or wild-type P53 (Review) International Journal of Oncology, 2011,38(5):1189-1195. [本文引用: 1]
ZHANGR T, SHIH R, RENF, CAOY, JIP C, WANGW W . Roles of P53 /CDK1 pathway of G2 /M checkpoint on proliferation and apoptosis of ovarian cancer cells .Acta Universitatis Medicinalis Anhui, 2018(04):502-508.(in Chinese) [本文引用: 1]
MAKRIGIANNAKISA, AMINK, COUKOSG, TILLYJ L, COUTIFARISC . Regulated expression and potential roles of P53 and Wilms' Tumor suppressor gene (Wt1) during follicular sevelopment in the human ovary Journal of Clinical Endocrinology and Metabolism, 2000,85(1):449-459. [本文引用: 1]
SIROTKINA V, BENCOA, TANDLMAJEROVAA, VASICEKD, KOTWICAJ, DARLAKK, VALENZUELAF . Transcription factor P53 can regulate proliferation, apoptosis and secretory activity of luteinizing porcine ovarian granulosa cell cultured with and without ghrelin and Fsh Reproduction, 2008,136(5):611-618. DOI:10.1530/REP-08-0229URL [本文引用: 1]
RAMJID P, FOKAP . Ccaat/Enhancer-binding proteins: Structure, function and regulation Biochemical Journal, 2002,365(3):561-575. DOI:10.1042/bj20020508URL [本文引用: 1]
FANH Y, LIUZ, JOHNSONP F, RICHARDSJ S . CCAAT/ enhancer-binding proteins (C/EBP)-α and -β are essential for ovulation, luteinization, and the expression of key target genes Molecular Endocrinology, 2011,25(2):253-268. DOI:10.1210/me.2010-0318URL [本文引用: 1]
PIONTKEWITZY, ENERBACKS, HEDINL . Expression of ccaat enhancer binding protein-alpha (C/Ebp Alpha) in the rat ovary: Implications for follicular development and ovulation Developmental Biology, 1996,179(1):288-296. DOI:10.1006/dbio.1996.0258URL [本文引用: 1]
FANH Y, LIUZ, JOHNSONP F, RICHARDJ S . CCAAT/enhancer- binding proteins (C/EBP)-α and -β are essential for ovulation, luteinization, and the expression of key target genes Molecular Endocrinology, 2011,25(2):253-268. DOI:10.1210/me.2010-0318URL [本文引用: 1]
 ,1
,1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT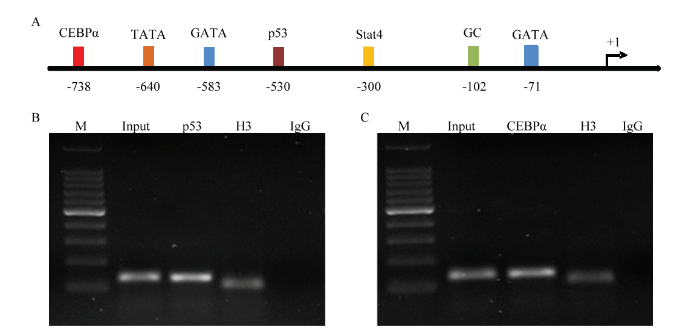 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT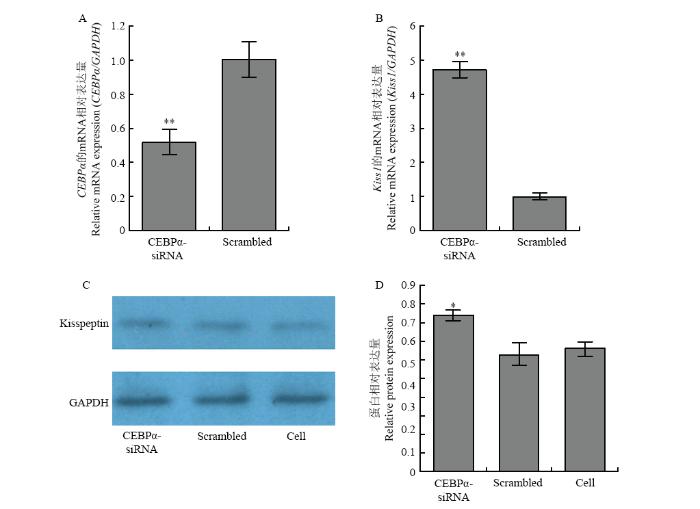 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}