 ,1
,1The Accumulation of Monoterpenes and the Expression of Its Biosynthesis Related Genes in ‘Aishen Meigui’ Grape Berries Cultivated in Different Trellis Systems During Ripening Stage
WANG HuiLing1, WANG XiaoYue2, YAN AiLing3, SUN Lei1, ZHANG GuoJun1, REN JianCheng1, XU HaiYing,1通讯作者:
收稿日期:2018-09-14接受日期:2019-01-11网络出版日期:2019-04-01
| 基金资助: |
Received:2018-09-14Accepted:2019-01-11Online:2019-04-01
作者简介 About authors
王慧玲,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (2177KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王慧玲, 王晓玥, 闫爱玲, 孙磊, 张国军, 任建成, 徐海英. 不同架式‘爱神玫瑰’葡萄果实成熟期间单萜积累及相关基因的表达[J]. 中国农业科学, 2019, 52(7): 1136-1149 doi:10.3864/j.issn.0578-1752.2019.07.002
WANG HuiLing, WANG XiaoYue, YAN AiLing, SUN Lei, ZHANG GuoJun, REN JianCheng, XU HaiYing.
0 引言
【研究意义】香气是衡量葡萄及葡萄酒品质的重要指标,直接影响消费者的认可程度。特别是玫瑰香型的鲜食葡萄备受消费者喜爱,玫瑰香味育种也一直是葡萄品质研究和育种的重点。萜烯类化合物因具浓郁香味,感官阈值低,是玫瑰香味葡萄的典型香气成分[1]。不同栽培架式结构影响果际微生态条件和叶片的光合作用最终影响葡萄香气积累[2]。‘爱神玫瑰’是极早熟无核鲜食葡萄新品种,具有典型的玫瑰香味[3]。探究不同栽培架式‘爱神玫瑰’葡萄果实发育中单萜化合物的积累模式,结合关键基因表达分析,可为阐明萜类物质的生物合成及调控机制,进而为葡萄香味品质改善和栽培架式的选择提供理论支撑。【前人研究进展】单萜的合成途径已经较为清晰,主要是通过位于质体中的脱氧-木酮糖-5-磷酸/甲基磷酸赤藓糖途径(DXP/MEP)合成[4]。影响果实萜类香气合成积累的因素很多,除了品种基因组成等内因外,还受生长环境及栽培技术措施等外因的影响[5,6,7]。在各种栽培措施中,栽培架式结构对叶幕类型可以产生最重要的和直接的影响,叶幕结构通过影响光能截留量影响叶际、果际微生态条件和叶片的光合作用,最终影响葡萄的产量和品质[8]。关于架式对葡萄果实品质的影响已有较多的报道,但大多关注可溶性固形物、花色苷、总酚等不可挥发性物质[9,10,11,12],对于香气成分研究较少。国内仅有的2篇报道主要研究了‘玫瑰香’葡萄在不同架式下芳香物质含量的变化。赵新节等[13]以7年生棚架和篱架栽培模式下的‘玫瑰香’葡萄为试材,研究发现篱架葡萄成熟后期酯类物质相对含量高于棚架, 而酮醛类物质和萜类物质相对含量低于棚架;商佳胤等[14]研究发现Y型架栽培的‘玫瑰香’葡萄果实中芳香化合物种类和含量都较高,葡萄果实风味优于篱架。而国外研究者主要关注架式对酿酒葡萄及其酿造葡萄酒中香气的影响。JI和DAMI[15]研究发现直立架式栽培的酿酒葡萄‘塔名内’(Traminette)中积累的单萜类化合物特别是香叶醇含量更高。FRAGASSO等[16]研究了不同整形方式下‘普里米蒂沃’(Primitivo)葡萄及其酿造葡萄酒中挥发性成分的变化。【本研究切入点】目前的研究主要是针对篱架和棚架,且主要在香气物质成分分析层面,其合成调控机制还不清晰。关于不同架式(V型和T型)对鲜食葡萄果实成熟过程中玫瑰香味化合物合成的影响及可能调控机制的研究鲜有报道。【拟解决的关键问题】本研究以T型和V型架式栽培下的‘爱神玫瑰’葡萄果实为试材,探讨两种架式下葡萄成熟期间单萜类化合物积累及其合成途径关键酶基因(VvDXS1、VvDXS3、VvDXR、VvDHR、VvLiner syn、Vvα-Terp syn)的表达模式,以期为适宜架式选择,为进一步提高果实中萜类物质的含量,改善果实玫瑰香味及阐明萜类物质的生物合成和调控机制提供一定的理论依据。1 材料与方法
试验于2016—2017年在北京市林业果树科学研究院进行。1.1 试验材料及地点
试验材料为7年生早熟玫瑰香型葡萄‘爱神玫瑰’(Vitis vinifera cv. Aishen Meigui),定植于北京市平谷区马昌营镇(北纬40°13′,东经117°12′),试验地土壤肥力中等,pH 6.9,有机质含量12.50 g?kg-1,全氮0.86 g?kg-1,全磷0.87 g?kg-1,全钾22.2 g?kg-1,可溶性盐1.03 g?kg-1。试验进行年份秋季沟施有机肥(1 t/667 m2),果实膨大期滴灌施肥(5 kg/667 m2)。栽培架式分为T型(顺行平棚架)和V型,两种架式的树形均为顺行水平龙干形,株行距1.5 m×3 m。两种架式均种植两行,各45株,2芽短枝修剪,每20 cm留一结果母枝,每个结果枝留2个果穗,豆果期疏去副穗和穗尖,保持单株产量一致。采用简易避雨、地表园艺地布覆盖、滴灌供水和常规病虫害等管理模式,生长期内修剪及肥水管理一致。1.2 试验方法
1.2.1 取样 2016、2017年每年6—8月取样。每种架式类型分为3个小区(30株/小区),葡萄果实从转色初期(花后30 d)开始取样,直至果实完全成熟。2016年分别于6月26日(花后30 d)、7月5日(花后39 d)、7月14日(花后48 d)、7月21日(花后57 d)、8月2日(花后67 d)和8月11日(花后76 d)取样6次。基于2016年试验数据,2017年于6月29日开始取样(花后30 d),每隔9 d取样一次,至8月4日(花后67 d)结束,共取样5次。取样时每个小区采集20穗(栽培行阴、阳面各取10穗),兼顾叶幕层内、外和上、中、下果穗,采样后立即运回实验室,液氮速冻后置于-80℃超低温冰箱保存备用。1.2.2 可溶性固形物和可滴定酸测定 从采集的新鲜果实样品中随机取出50粒,挤出葡萄汁,分别测定可溶性固形物和可滴定酸。可溶性固形物采用手持式折光仪PAL-1(Atago Co,Tokyo,Japan)测定;可滴定酸采用0.1 mol?L-1 NaOH进行测定,含量以酒石酸表示,3次重复。
1.2.3 香气化合物的提取和测定 香气物质的提取和测定参考WEN等[17]的方法略有改动。取50 g葡萄果实,液氮保护下打碎去籽,分别加入0.5 g D-葡萄糖酸内酯和2 g聚乙烯基聚吡咯烷酮,利用研磨机(IKA A11)打磨成粉末,室温静置浸提,然后于4℃下8 500 r/min离心10 min,得到澄清葡萄汁。每个处理进行3 次独立的提取。取5 mL葡萄汁加入1.0 g NaCl和10 μL 4-甲基-2-戊醇,40℃加热搅拌30 min后,用已活化或热解析过的聚二甲基硅氧烷/碳筛/二乙烯苯(PDMS/CAR/DVB)萃取头相同温度下吸附30 min,然后将萃取头插入气相色谱的进样口,250℃下热解析8 min。
试验所用气相色谱为安捷伦7890B,质谱为5977A(Agilent,USA),色谱柱为HP-INNOWAX(长60 m,内径0.25 mm,液膜厚度0.25 μm)。载气为高纯氦气,流速为1 mL?min-1。固相微萃取采用自动进样,不分流模式。柱温箱的升温程序为:50℃保持1 min,然后以3℃?min-1的速度升温至220℃,保持5 min。质谱接口温度为280℃,离子源温度为230℃,电离方式EI,离子能量70 ev,质量扫描范围为20—350 u。
根据已有化合物标样的色谱保留时间和质谱信息,参照NIST 05标准谱库及相关文献对样品进行定性分析;利用已有的化合物制备标准曲线进行定量分析,没有标准品的物质利用化学结构、官能团相似、碳原子数相近的标准品的标准曲线进行定量。
1.2.4 RNA提取和qPCR分析 采用植物RNA快速提取试剂盒(北京华越洋生物科技有限公司)提取葡萄果实的总RNA。然后使用反转录试剂盒(Promega,A3500),参照说明书反转录成cDNA,置于-80℃保存备用。荧光实时定量qPCR反应体系参照WEN等[17]的方法。在萜类生物合成途径中的6个关键基因的引物序列参照孙磊等[18]的合成,使用VvUbiquitin、VvEF1-α和VvGAPDH 3个内参基因做为对照。
1.3 数据处理与统计分析
数据统计分析利用分析软件SPSS 13.0,采用Duncan多重比较进行显著性方差分析,最低显著水平P<0.05;主成分分析和聚类分析采用MetaboAnalyst 4.0;绘图采用Excel和Sigma Plot 10.0。2 结果
2.1 不同架式葡萄果实成熟期间可滴定酸及可溶性固形物的变化
葡萄果实可溶性固形物含量在两种栽培架式下均随着果实成熟逐渐升高,可滴定酸含量逐渐降低(表1)。V型架栽培‘爱神玫瑰’葡萄在花后48 d可溶性固形物含量分别升至15.72%(2016年)和15.81%(2017年),可滴定酸含量分别降至4.06 g?L-1(2016年)和4.16 g?L-1(2017年),花后57 d可溶性固形物及可滴定酸含量均趋于稳定,确定此时葡萄进入成熟期。而T型架式栽培葡萄可溶性固形物含量在花后57 d分别上升至15.38%(2016年)和16.01%(2017年),在花后67 d含量升至最高,而可滴定酸降至较低水平。果实成熟后,T型架式栽培的葡萄果实可溶性固形物含量高于V型架,含酸量无显著差异。Table 1
表1
表1不同架式果实成熟期间可滴定酸及可溶性固形物分析
Table 1
| 盛花后天数 Days after full blossom (d) | 2016年 | 2017年 | ||||||
|---|---|---|---|---|---|---|---|---|
| 可溶性固形物 Soluble solids (%) | 可滴定酸 Titratable acid (g·L-1) | 可溶性固形物 Soluble solids (%) | 可滴定酸 Titratable acid (g·L-1) | |||||
| T | V | T | V | T | V | T | V | |
| 30 | 10.70±1.20a | 10.54±0.87a | 6.90±0.23a | 6.35±0.32b | 10.31±0.61a | 10.42±0.53a | 6.79±0.12a | 6.45±0.32a |
| 39 | 14.01±0.21b | 14.69±0.86a | 6.18±0.10a | 6.20±0.15a | 13.71±0.12b | 14.03±0.21a | 6.05±0.15a | 6.13±0.04a |
| 48 | 14.78±0.34b | 15.72±0.53a | 5.31±0.41a | 4.06±0.02b | 15.22±0.48b | 15.81±0.57a | 4.34±0.09a | 4.16±0.14b |
| 57 | 15.38±0.38b | 16.01±0.65a | 3.39±0.13a | 3.26±0.17a | 16.03±0.86b | 16.14±0.58a | 3.34±0.21a | 3.48±0.04a |
| 67 | 16.12±1.02a | 16.03±0.21b | 3.20±0.20a | 3.23±0.45a | 16.24±0.49a | 16.03±0.12b | 3.09±0.13a | 3.15±0.11a |
| 76 | 16.67±0.36a | 16.05±0.12b | 3.24±0.04a | 3.20±0.25a | - | - | - | |
新窗口打开|下载CSV
2.2 不同架式对葡萄果实中单萜类化合物组成的影响
利用SPEME-GC-MS方法,本研究2016年对28种游离态单萜类化合物进行了定性定量分析(表2)。在两种架式栽培‘爱神玫瑰’葡萄果实成熟过程中,异香叶醇在V型架葡萄果实成熟后期未检测到,其余化合物均检测到。在2017年对27种游离态单萜类化合物进行了定性定量分析,所有化合物均检测到(表2)。Table 2
表2
表2不同架式葡萄果实成熟期间单萜含量分析
Table 2
| 化合物 Compound | 年份 Year | 架式Trellis | 盛花后天数 Days after full blossom (d) | |||||
|---|---|---|---|---|---|---|---|---|
| 30 | 39 | 48 | 57 | 67 | 76 | |||
| β-月桂烯 β-Myrcene | 2016 | T V | 0.25±0.02a 0.24±0.03a | 0.86±0.06b 4.71±0.06a | 0.22±0.01b 0.34±0.02a | 1.28±0.09a 0.26±0.02b | 1.32±0.12a 0.44±0.07b | 0.45±0.04a 0.16±0.00b |
| 2017 | T V | 0.95±0.01b | 1.02±0.01b | 1.99±0.10b | 10.30±0.31a | 7.21±0.06a | - | |
| 1.17±0.40a | 1.24±0.34a | 3.35±0.11a | 1.56±0.09b | 1.17±0.02b | - | |||
| 柠檬烯 Limonene | 2016 | T V | 8.30±0.04a 3.69±0.11b | 7.31±0.00a 3.71±0.11b | 1.86±0.01b 5.86±1.04a | 9.08±0.15a 2.33±0.12b | 11.03±0.19a 1.68±0.08b | 7.73±0.14a 2.56±0.07b |
| 2017 | T V | 5.16±0.10a | 5.08±0.12a | 5.42±0.04b | 17.57±0.31a | 16.98±0.32a | - | |
| 3.43±0.12b | 5.20±0.09a | 9.18±0.51a | 6.50±0.20b | 3.35±0.07b | - | |||
| 水芹烯 Phellandrene | 2016 | T V | 1.95±0.06a 0.87±0.02b | 2.07±0.01a 0.77±0.02b | 0.56±0.10b 1.11±0.07a | 1.08±0.11a 0.53±0.01b | 1.91±0.07a 0.47±0.08b | 1.49±0.05a 0.56±0.03b |
| 2017 | T V | 1.72±0.01a | 3.40±0.01a | 0.74±0.02b | 1.56±0.05a | 1.82±0.02a | - | |
| 0.79±0.01b | 0.73±0.08b | 1.10±0.06a | 0.73±0.01b | 0.56±0.02b | - | |||
| β-trans-罗勒烯 β-trans-Ocimene | 2016 | T V | 3.08±0.00a 1.17±0.08b | 2.61±0.03a 1.21±0.01b | 0.71±0.00b 1.96±0.34a | 3.68±0.02a 0.83±0.10b | 4.41±0.03a 0.57±0.05b | 3.30±0.03a 1.00±0.03b |
| 2017 | T V | 3.07±0.01a | 2.96±0.03a | 3.99±0.22b | 9.24±0.26a | 8.89±0.06a | - | |
| 2.73±0.04b | 2.97±0.41a | 6.15±0.35a | 3.07±0.06b | 2.64±0.12b | - | |||
| γ-松油烯 γ-Terpinen | 2016 | T V | 5.26±0.02a 3.10±0.16b | 3.84±0.10a 1.41±0.08b | 0.91±0.03a 0.87±0.15b | 1.34±0.03a 0.30±0.01b | 1.93±0.05a 0.39±0.18b | 0.87±0.03a 0.32±0.01b |
| 2017 | T V | 5.79±0.61a | 3.74±0.04a | 2.17±0.07a | 2.06±0.40a | 1.56±0.07a | - | |
| 2.59±0.03b | 2.19±0.21b | 1.53±0.06b | 1.14±0.07b | 1.08±0.12b | - | |||
| β-cis-罗勒烯 β-cis-Ocimene | 2016 | T V | 4.22±0.00a 2.52±0.17b | 3.95±0.04a 2.60±0.04b | 1.50±0.01b 3.60±0.34a | 6.51±0.05a 1.86±0.12b | 8.44±0.16a 1.14±0.09b | 5.44±0.14a 2.18±0.00b |
| 2017 | T V | 6.36±0.21a | 5.07±0.11a | 3.54±0.01b | 18.94±0.03a | 19.06±0.15a | - | |
| 3.81±0.02b | 4.01±0.25b | 8.60±0.31a | 3.92±0.20b | 0.98±0.31b | - | |||
| 异松油烯 Terpinolen | 2016 | T V | 5.15±0.00a 2.71±0.13b | 4.57±0.00a 2.32±0.10b | 1.06±0.04b 3.47±0.96a | 4.94±0.05a 0.98±0.06b | 5.50±0.06a 0.73±0.18b | 4.50±0.04a 1.20±0.10b |
| 2017 | T V | 7.32±0.02a | 6.67±0.30a | 6.12±0.03b | 7.31±0.41a | 9.94±0.08a | - | |
| 4.32±0.40b | 4.33±0.13b | 7.79±0.16a | 3.12±0.04b | 1.83±0.17b | - | |||
| cis-氧化玫瑰 cis Rose oxide | 2016 | T V | 0.35±0.01b 0.80±0.02a | tr 0.25±0.01 | tr 0.21±0.02 | 0.30±0.01b 0.46±0.00a | 0.23±0.00 tr | tr tr |
| 2017 | T V | 0.64±0.01a | 0.68±0.01b | 0.73±0.13b | 0.85±0.02a | 0.59±0.12a | - | |
| 0.68±0.01a | 0.82±0.01a | 0.82±0.03a | 0.70±0.03b | 0.36±0.04b | - | |||
| trans-氧化玫瑰 trans-Rose oxide | 2016 | T V | tr tr | tr tr | tr tr | tr tr | tr tr | tr tr |
| 2017 | T V | tr | tr | 0.62±0.02a | 2.93±0.07a | 0.61±0.01 | - | |
| tr | tr | 0.64±0.01a | 0.56±0.04b | tr | - | |||
| 别罗勒烯 Allo-Ocimene | 2016 | T V | 3.16±0.01a 1.12±0.06b | 2.54±0.04a 1.10±0.02b | 0.70±0.03b 1.85±0.33a | 3.79±0.02a 0.87±0.01b | 4.48±0.07a 0.55±0.03b | 3.24±0.04a 0.89±0.01b |
| 2017 | T V | 2.98±0.01a | 2.86±0.30a | 2.86±0.06b | 5.36±0.06a | 6.12±0.09a | - | |
| 2.62±0.04b | 2.85±0.03a | 3.66±0.16a | 1.37±0.02b | 0.73±0.02b | - | |||
| 化合物 Compound | 年份 Year | 架式Trellis | 盛花后天数 Days after full blossom (d) | |||||
| 30 | 39 | 48 | 57 | 67 | 76 | |||
| (E,Z)-别罗勒烯 (E,Z)-Allo- Ocimene | 2016 | T V | 0.55±0.02a 0.37±0.01b | 0.57±0.02a 0.36±0.01b | 0.35±0.03b 0.58±0.08a | 1.27±0.03a 0.48±0.12b | 1.47±0.07a 0.26±0.03b | 0.82±0.01a 0.32±0.01b |
| 2017 | T V | 0.62±0.10b | 0.63±0.03a | 0.74±0.04b | 1.42±0.02a | 1.82±0.08a | - | |
| 0.69±0.01a | 0.64±0.01a | 0.81±0.09a | 0.64±0.04b | 0.41±0.02b | - | |||
| cis-呋喃型氧化里那醇 cis-furan linalool oxide | 2016 | T V | 9.31±0.01a 2.50±0.04b | 5.03±0.13a 2.15±0.14b | 1.46±0.05b 2.66±0.00a | 2.66±0.10a 1.32±0.05b | 3.37±0.16a 1.09±0.03b | 1.81±0.07a 1.18±0.00a |
| 2017 | T V | 4.92±0.07a | 2.65±0.01a | 1.23±0.31b | 3.19±0.02a | 4.00±0.22a | - | |
| 1.43±0.28b | 1.80±0.17b | 2.71±0.50a | 1.32±0.15b | 0.30±0.07b | - | |||
| trans-呋喃型氧化里那醇 trans-furan linalool oxide | 2016 | T V | 3.49±0.11a 1.73±0.03b | 2.32±0.05a 1.52±0.03b | 1.14±0.01b 1.47±0.03a | 1.68±0.04a 1.19±0.02b | 1.84±0.04a 1.05±0.01b | 1.23±0.02a 1.01±0.00b |
| 2017 | T V | 1.76±0.04a | 0.79±0.09b | 0.35±0.08b | 1.09±0.49a | 1.83±0.07a | - | |
| 1.41±0.05b | 1.14±0.31a | 0.58±0.12a | 0.53±0.01b | 0.23±0.04b | - | |||
| 橙花醚 Nerol oxide | 2016 | T V | 10.02±0.17a 5.92±0.05b | 10.18±0.30a 8.98±0.55b | 8.86±0.14a 3.09±2.40b | 7.05±0.94b 11.79±0.26a | 3.53±1.13a 3.38±0.60a | 3.65±0.02a 2.31±0.07b |
| 2017 | T V | 25.16±0.09a | 23.70±1.74a | 17.78±0.62b | 16.86±0.73a | 10.41±0.16a | - | |
| 14.15±0.53b | 17.16±0.73b | 21.77±0.22a | 15.12±0.12b | 8.97±0.21b | - | |||
| 香茅醛 Citronellal | 2016 | T V | 0.81±0.07a 0.42±0.05b | 0.59±0.06a 0.50±0.01a | 0.34±0.01b 0.65±0.04a | 0.77±0.04a 0.24±0.10b | 0.99±0.04a 0.10±0.01b | 0.60±0.06a 0.28±0.04b |
| 2017 | T V | - | - | - | - | - | - | |
| - | - | - | - | - | - | |||
| 里那醇 Linalool | 2016 | T V | 3.63±0.04b 5.58±0.04a | 8.33±0.12a 6.79±0.35b | 4.44±0.13b 14.62±0.77a | 41.67±0.94a 10.48±0.32b | 19.29±1.00a 4.23±0.55b | 33.60±0.70a 11.12±0.26b |
| 2017 | T V | 8.38±1.12a | 15.22±0.15b | 82.89±3.07a | 96.64±2.25a | 43.56±0.75a | - | |
| 7.01±0.05b | 15.26±0.37a | 71.08±1.03b | 67.56±0.54b | 8.90±0.43b | - | |||
| 4-松油烯醇 4-Terpineol | 2016 | T V | 2.95±0.01a 0.88±0.03b | 2.72±0.00a 0.52±0.05b | 0.39±0.03b 0.47±0.01a | 0.81±0.06a 0.22±0.01b | 1.24±0.02b 3.08±0.23a | 0.51±0.01a 0.16±0.01b |
| 2017 | T V | 2.91±0.09a | 3.23±0.04a | 2.89±0.07a | 2.56±0.01a | 1.98±0.02a | - | |
| 0.90±0.02b | 0.87±0.01b | 0.83±0.02b | 0.72±0.02b | 0.70±0.02b | - | |||
| 橙花醛 Neral | 2016 | T V | 0.67±0.01a 0.27±0.01b | 0.34±0.02a 0.33±0.05b | 0.13±0.01b 0.60±0.05a | 0.49±0.06b 0.59±0.04a | 0.67±0.08a 0.14±0.03b | 0.40±0.03a 0.15±0.08b |
| 2017 | T V | 0.87±0.03a | 0.75±0.01a | 0.63±0.01b | 0.65±0.05b | 0.84±0.01a | - | |
| 0.53±0.01b | 0.61±0.02b | 1.01±0.10a | 0.83±0.01a | 0.62±0.03b | - | |||
| α-衣兰油烯 α-Muurolene | 2016 | T V | 0.22±0.00a 0.10±0.00b | 0.09±0.00a 0.07±0.00b | 0.07±0.00a 0.07±0.02a | 0.08±0.01b 0.09±0.01a | 0.09±0.00a 0.08±0.00b | 0.07±0.02b 0.08±0.01a |
| 2017 | T | 0.60±0.01a | 0.54±0.01b | 0.55±0.01b | 0.58±0.02a | 0.52±0.05b | - | |
| V | 0.59±0.01a | 0.59±0.01a | 0.60±0.01a | 0.58±0.02a | 0.60±0.01a | - | ||
| α-萜品醇 α-Terpineol | 2016 | T V | 5.55±0.11a 4.49±0.02b | 5.28±0.01a 4.59±0.05b | 4.34±0.01b 4.99±0.11a | 6.53±0.19a 4.36±0.09b | 7.96±0.15a 2.80±0.97b | 5.78±0.13a 4.42±0.02b |
| 2017 | T | 5.40±0.04a | 9.23±1.07a | 6.65±0.57b | 22.29±0.11a | 19.89±0.62a | - | |
| V | 3.24±0.23b | 6.41±0.78b | 14.57±0.57a | 3.03±0.03b | 1.81±0.14b | - | ||
| 化合物 Compound | 年份 Year | 架式Trellis | 盛花后天数 Days after full blossom (d) | |||||
| 30 | 39 | 48 | 57 | 67 | 76 | |||
| 香叶醛 Geranial | 2016 | T V | 0.22±0.06b 0.31±0.09a | 0.44±0.02b 0.53±0.03a | 0.92±0.04b 1.08±0.11a | 2.03±0.85a 1.20±0.21b | 2.68±0.05a 0.53±0.09b | 0.73±0.02a 0.35±0.02b |
| 2017 | T | 0.63±0.01a | 0.74±0.01a | 0.87±0.02b | 1.68±0.01a | 2.13±0.13a | - | |
| V | 0.62±0.02a | 0.65±0.01b | 0.89±0.01a | 1.20±0.10b | 0.63±0.02b | - | ||
| β-香茅醇 β-Citronellol | 2016 | T | 1.00±0.05b 3.58±0.03a | 3.61±0.02a 3.65±0.04a | 3.59±0.03a 2.59±0.78b | 3.60±0.05a 0.53±0.09b | 2.54±0.76a 0.05±0.01b | 0.61±0.03a 0.23±0.01b |
| V | ||||||||
| 2017 | T | 2.59±0.15a | 4.54±0.06a | 5.56±0.06a | 7.58±0.01a | 3.70±0.11a | - | |
| V | 2.49±0.04b | 3.51±0.02b | 4.61±0.04b | 2.40±0.01b | 0.64±0.01b | - | ||
| γ-香叶醇 γ-Geraniol | 2016 | T V | 0.15±0.00a 0.06±0.01b | 0.14±0.03a 0.08±0.01b | 0.10±0.02a 0.08±0.02b | tr tr | tr tr | tr tr |
| 2017 | T | 0.54±0.01a | 0.52±0.03a | 0.53±0.04 | tr | tr | - | |
| V | 0.20±0.03b | 0.21±0.01b | tr | tr | tr | - | ||
| 橙花醇 Nerol | 2016 | T V | 0.15±0.01b 0.24±0.00a | 0.32±0.02a 0.29±0.04b | 0.27±0.01b 0.77±0.07a | 1.77±0.13a 0.45±0.07b | 3.58±0.01a 0.25±0.05b | 1.06±0.08a 0.25±0.04b |
| 2017 | T | 2.09±0.14b | 3.43±0.07b | 21.47±2.78b | 82.82±8.42a | 65.23±0.75a | - | |
| V | 2.89±1.17a | 4.62±0.35a | 38.38±3.60a | 19.56±0.54b | 8.99±2.07b | - | ||
| cis-异香叶醇 cis-Isogeraniol | 2016 | T V | 0.17±0.16 tr | tr tr | tr tr | tr tr | tr tr | tr nd |
| 2017 | T | tr | 0.25±0.08 | tr | tr | tr | - | |
| V | tr | tr | tr | tr | tr | - | ||
| trans-异香叶醇 trans-Isogeraniol | 2016 | T V | tr tr | tr tr | tr tr | tr tr | tr tr | nd nd |
| 2017 | T | 0.13±0.03 | tr | tr | tr | tr | tr | |
| V | tr | tr | tr | tr | tr | tr | ||
| 香叶醇 Geraniol | 2016 | T V | 0.62±0.05b 1.06±0.01a | 3.25±1.13b 4.76±0.01a | 4.91±0.02a 4.92±0.02a | 5.22±0.03a 1.33±0.20b | 5.62±0.02a 0.67±0.10b | 5.01±0.03a 1.07±0.04b |
| 2017 | T | 3.89±0.19b | 12.56±0.04b | 58.97±3.42a | 81.60±4.67a | 82.14±0.86a | - | |
| V | 7.31±0.10a | 20.40±0.55a | 54.27±6.01b | 21.46±3.07b | 17.59±2.26b | - | ||
| 香叶酸 Geranic acid | 2016 | T V | 0.29±0.02a 0.17±0.02b | 0.51±0.20a 0.33±0.04b | 0.81±0.30a 0.51±0.16b | 5.22±0.03a 1.33±0.20b | 0.39±0.14 tr | 0.24±0.01a 0.18±0.03b |
| 2017 | T | 0.12±0.01b | 2.98±0.12b | 6.12±0.22a | 8.12±0.56a | 1.99±0.12a | - | |
| V | 0.23±0.02a | 3.53±0.10a | 5.96±0.16b | 6.32±0.08b | 1.49±0.04b | - | ||
| 总量 Total | 2016 | T V | 71.50±0.09a 43.87±0.89b | 71.40±2.69a 53.52±1.32b | 39.66±0.23b 58.38±8.38a | 108.18±3.15a 43.12±1.11b | 94.52±4.29a 23.61±3.55b | 83.08±0.46a 31.97±0.63b |
| 2017 | T | 94.63±1.23a | 113.21±3.42a | 235.43±0.41b | 403.24±2.54a | 312.82±4.12a | - | |
| V | 65.86±3.11b | 100.46±4.10b | 260.91±0.91a | 163.97±0.87b | 64.61±5.16b | - | ||
新窗口打开|下载CSV
通过两个年份化合物数据分析发现,在葡萄果实成熟过程中,不同架式主要单萜组分差异显著。成熟初期(花后30 d),T型架葡萄果实中含量最高的5个化合物依次为橙花醚(2016—2017年)、里那醇(2017年)、异松油烯(2017年)、β-cis-罗勒烯(2017年)、cis-呋喃型氧化里那醇(2016年)、柠檬烯(2016年)、α-萜品醇(2016年)和γ-松油烯(2016—2017年)等,5个化合物含量占单萜总含量的60%以上。而V型架葡萄果实中主要单萜类化合物包括橙花醚(2016—2017年)、里那醇(2016—2017年)、异松油烯(2017年)、α-萜品醇(2016年)、柠檬烯(2016—2017年)和β-香茅醇(2016年)。花后39 d,T型架栽培果实中里那醇含量迅速升高成为第二主要成分,V型架果实中香叶醇成为主要成分,橙花醚在两种架式果实中含量仍然较高。花后48 d,两年数据均显示橙花醚、香叶醇、里那醇、α-萜品醇成为T型架果实中主含量萜类物质;而V型架葡萄中里那醇含量最高,此外含量较高的还有香叶醇(2016—2017年)、柠檬烯(2016年)、α-萜品醇(2016—2017年)和β-cis-罗勒烯(2016年)。T型架果实在花后57 d主要单萜组成包括里那醇(2016—2017年)、橙花醇(2017年)、香叶醇(2017年)、柠檬烯(2016年)、橙花醚(2016年)、α-萜品醇(2016—2017年)和β-cis-罗勒烯(2016年),其中里那醇含量分别达到41.67 μg?L-1(2016年)和96.64 μg?L-1(2017年);V型架果实主要单萜成分有橙花醚(2016—2017年)、里那醇(2016—2017年)、橙花醇(2017年)、香叶醇(2017年)、α-萜品醇(2016年)、柠檬烯(2016—2017年)和β-cis-罗勒烯。花后67 d,两种架式果实中主要单萜类化合物组分变化较小,其中T型果实中主要是里那醇、柠檬烯、α-萜品醇、β-cis-罗勒烯和香叶醇,V型架栽培果实中主要有里那醇、香叶醇、柠檬烯、橙花醚等,2016年里那醇在两种架式果实中含量最高,而在2017年香叶醇含量最高。
2.3 不同架式对葡萄果实成熟期间单萜化合物含量的影响
2年化合物数据显示不同架式单萜化合物总量差异显著(表2)。转色初期(花后30 d)到花后39 d,T型架栽培葡萄果实中单萜总量显著高于V型架。花后48 d,V型架葡萄果实单萜总量分别升高达到58.4 μg·L-1(2016年)及260.9 μg·L-1(2017年),显著高于T型葡萄果实单萜总量。花后57 d,T型架葡萄果实单萜含量迅速上升,远高于V型架。显著性差异分析显示,成熟期大多数单萜类化合物在T型架果实含量显著高于V型架(表2)。根据两年化合物含量进行聚类分析(图1),不同架式‘爱神玫瑰’果实成熟期间游离态单萜含量变化主要表现两种模式(红色数字表示)。第一种模式包含(E,Z)-别罗勒烯(M11)、β-cis-罗勒烯(M6)、柠檬烯(M2)、β-trans-罗勒烯(M4)、别罗勒烯(M10)、异松油烯(M7)、α-萜品醇(M20)、香叶醇(M27)、里那醇(M16)、橙花醇(M24)、香叶醛(M21)、香叶酸(M28)和β-月桂烯(M1)等化合物,这一大类化合物大多数在果实成熟时含量达到最高。但是在不同架式表现又略微不同。(E,Z)-别罗勒烯(M11)、β-cis-罗勒烯(M6)、柠檬烯(M2)、β-trans-罗勒烯(M4)、别罗勒烯(M10)、异松油烯(M7)和α-萜品醇(M20)这7种化合物在T型架果实中表现为先下降,花后57 d急剧升高,成熟后期(花后76 d)又下降的趋势。而在V型架果实中这7种化合物含量随着果实成熟逐渐上升,花后48 d达到积累高峰,之后又逐渐下降至最低含量。香叶醇(M27)、里那醇(M16)、橙花醇(M24)、香叶醛(M21)、香叶酸(M28)和β-月桂烯(M1)等化合物则随着果实成熟含量逐渐升高,到果实成熟期达到最高,到果实成熟后期(花后76 d)又有所下降。在第二种模式中,大多数化合物在成熟初期(花后30 d)含量最高,随着果实成熟含量降低。在T型架果实中,这类化合物在花后48 d含量降至最低,随着果实成熟又有所升高。而在V型架果实中化合物含量随着成熟过程持续降低。这类化合物包括γ-松油烯(M5)、cis-呋喃型氧化里那醇(M12)、trans-呋喃型氧化里那醇(M13)、4-松油烯醇(M17)、水芹烯(M3)、β-香茅醇(M22)和香茅醛(M15)等。
图1
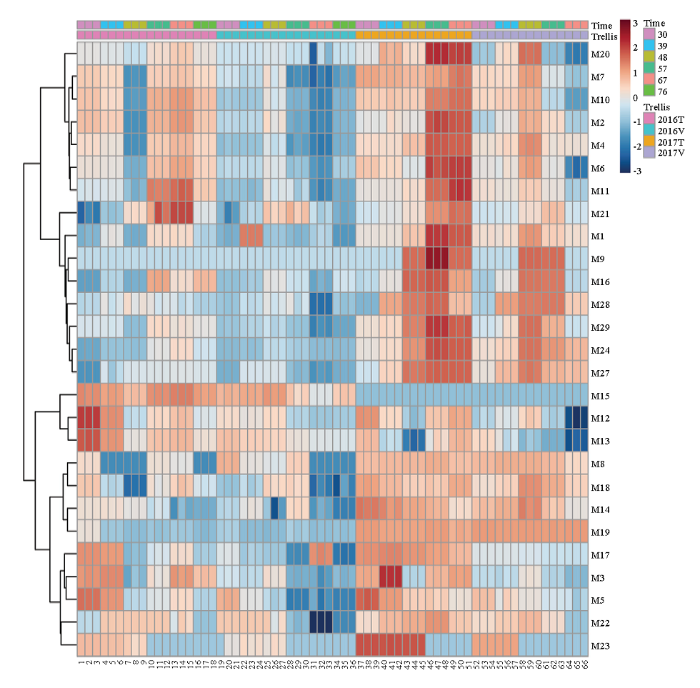 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同架式葡萄果实成熟期间单萜含量聚类分析
Fig. 1Hierarchical cluster analysis of monoterpenes in grape berries cultivated in different trellis systems during maturity
此外,对两年单萜化合物含量数据进行了主成分和最小二乘判别分析,由结果可见(图2,图3),2016年前两个主成分的累计方差贡献率为76.8%,而2017年前两个主成分的累计方差贡献率为84.1%,均反映了所有化合物的绝大部分信息。根据两个主成分可以把不同架式处理进行很好的区分。最小二乘判别分析结果显示VIP值大于1.0被认为是主要贡献差异化合物,由图3可知,2016年VIP值较大的5个化合物依次为β-香茅醇(M22)、香叶醇(M27)、香叶酸(M28)、γ-松油烯(M5)、橙花醇(M24)和里那醇(M16)。2017年VIP值较大的5个化合物依次为4-松油烯醇(M17)、trans-氧化玫瑰(M9)、香叶醇(M27)、γ-香叶醇(M23)和β-香茅醇(M22)。
图2
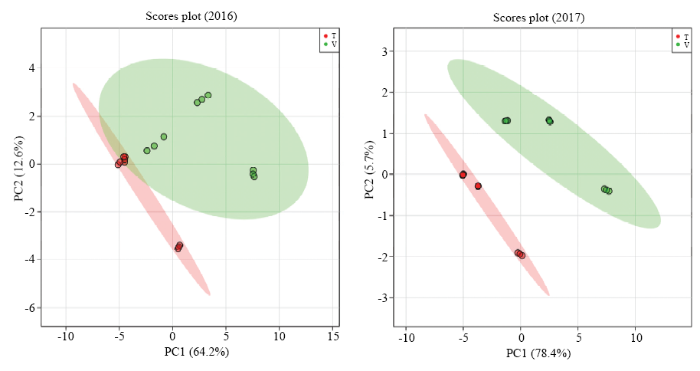 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同架式葡萄果实成熟期间单萜含量主成分分析
Fig. 2Principal analysis of monoterpenes in grape berries cultivated in different trellis systems during maturity
图3
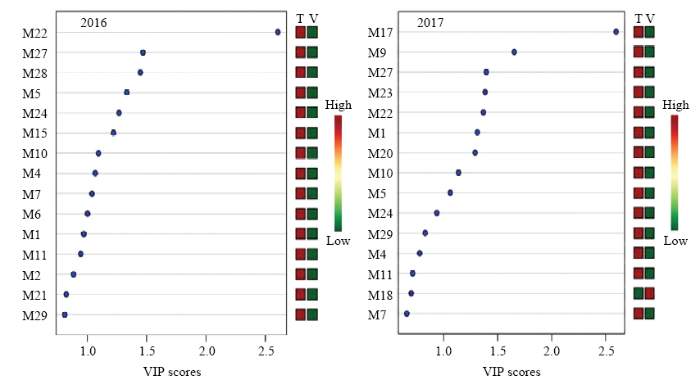 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同架式葡萄果实成熟期间基于VIP值的主要化合物
Fig. 3The selected compound based on VIP score in grape berries cultivated in different trellis systems during maturity
2.4 不同架式栽培葡萄果实成熟期间单萜合成途径基因转录水平变化
DXS1表达量在两种架式葡萄果实中均从成熟初期(花后30 d)开始上升,T型架果实中DXS1在花后48 d表达量升至最高,随后逐渐降低;而V型架DXS1在花后39 d达到表达高峰,随后降低。T型架DXS1最高表达量显著高于V型架,是V型架DXS1表达量的1.40倍。DXS3在两种架式不同成熟时期葡萄果实中表达量呈现不同的变化趋势,在T型架葡萄果实中,DXS3随着果实成熟略有下降之后逐渐上升,至花后57 d达到表达高峰,之后又迅速下降。而对于V型架,随着果实成熟,DXS3表达量逐渐上升,到花后48 d又有所下降,之后又逐渐上升。成熟期DXS3在T型架栽培果实中表达量高于V型架。DXR在两种架式葡萄果实成熟期间表达模式相似,都是先上升再下降,随着果实成熟再次上升,至成熟后期又下降。在成熟期T型架葡萄果实DXR表达量高于V型架。HDR在两种架式葡萄果实成熟期间表达模式不同,在T型架果实成熟过程中,HDR表达量逐渐上升,至花后48 d表达量最高,之后逐渐降低;在V型架葡萄果实成熟过程中HDR并没有明显的表达高峰。Liner syn在T型架葡萄果实中表达量明显高于V型架,转色后表达量下降,随后在花后57 d急剧上升至最高表达量。V型架葡萄果实中Liner syn转色后表达量同样略有上升,之后有所下降,整体变化趋势平缓。Terp syn在两种架式葡萄果实成熟过程中表达模式差异明显,在T型架葡萄果实中,成熟初期(花后30 d)表达量最高,之后下降,至48 d后又略有上升;而对于V型架栽培葡萄果实,Terp syn在花后48 d表达上升之后下降。
3 讨论
栽培技术和栽培措施都与葡萄浆果品质有着密切关系。适宜架式的选择、架形改造和新式架型的应用是提高葡萄果实品质的重要举施。研究表明不同架式对葡萄香气物质产生重要的影响[13,14,15,16]。本研究发现T型架栽培葡萄果实中萜类物质含量更高,这可能是因为T型架栽培的相对光照强度高于V型架 [19],而光照有利于香气物质积累[1]。此外,两种架式萜类化合物积累模式有差异。V型架葡萄果实萜类化合物积累高峰出现更早一些(花后48 d),T型架葡萄果实在花后57—67 d(图1)。这可能与V型架葡萄果实更早进入成熟期有关(表1)。陆贵峰等[19]也研究发现T型篱架栽培的‘凌丰’葡萄果实成熟期比V型篱架栽培晚3—5 d。随着果实成熟,大部分萜类化合物含量逐渐升高。但是在果实成熟后期,两种架式‘爱神玫瑰’葡萄果实中大部分萜类物质均呈下降趋势。有研究结果表明,‘瑞都香玉’葡萄主要呈香型萜烯成分里那醇、萜品醇含量在成熟期仍有增加,果实香气成熟期稍晚于生理成熟期,适当延迟采收有助于其特征香型的形成[20],但是对于‘爱神玫瑰’葡萄延迟采收会导致玫瑰香味品质降低,可能是由品种不同造成的差异。一般认为玫瑰香型葡萄中主要呈香单萜类物质为里那醇、香茅醇、橙花醇、萜品醇和香叶醇[21]。本研究发现两种架式葡萄果实中主要萜类化合物组分随着果实发育发生变化(表2)。2016年在成熟期‘爱神玫瑰’葡萄果实中,主要单萜类成分有里那醇、α-萜品醇、香叶醇、柠檬烯、β-cis-罗勒烯、橙花醚和异松油烯,2017年主要有里那醇、α-萜品醇、香叶醇、橙花醇和-cis-罗勒烯,而香茅醇含量并不高。但是最小二乘判别分析结果显示不同架式主要差异萜类化合物里包含β-香茅醇(M22)、香叶醇(M27)及氧化玫瑰等化合物,被认为是主要贡献萜类成分,可见化合物含量高低并不是玫瑰香味的主要决定因素。RUIZ-GARCíA等[22]指出氧化玫瑰存在与否可以作为鲜食葡萄玫瑰香味有无的评判标准,而氧化玫瑰含量很低。本研究结果表明两个年份两种架式成熟葡萄果实中里那醇含量均比较高,且高于其香气阈值[17],可能是‘爱神玫瑰’葡萄的主要呈香成分。
葡萄中单萜主要通过DXP/MEP途径合成。本研究跟踪了单萜合成途径中6个重要基因的表达变化,各个基因表达量随着果实成熟都有一个上升的过程(图4)。不同架式葡萄果实成熟期间单萜总量积累规律与DXS3、HDR、Liner syn和Terp syn表达规律相似。王继源等[23,24]研究发现‘阳光玫瑰’葡萄果实成熟过程中DXS3表达量上调促进单萜物质的生成。孙磊等[18]也发现在‘亚历山大’葡萄果实发育过程中,单萜总量积累与DXS3和Terp syn的转录情况具有显著相关性,而与DXS1表达相关性较低,认为果实后期萜类积累有可能是由于DXS3表达造成的。研究认为葡萄DXS1对于玫瑰香味产生具有重要的作用,其基因序列的SNP突变是产生玫瑰香味的重要原因[25,26,27],但是对于其表达关注比较少。本研究发现DXS1随着果实成熟表达上调,而表达积累高峰早于DXS3,可能果实成熟早期单萜积累主要与DXS1表达有关。成熟期T型架果实中各个基因表达量明显高于V型架(图4),与单萜类化合物积累模式相一致。此外MARTIN等[28]用琼瑶浆为材料的试验结果也证明HDR的表达与单萜积累规律相一致。Liner syn和Terp syn的表达高峰与里那醇和萜品醇等产物积累略有差异,T型架和V型架果实中Liner syn尽管随着果实成熟表达上调,但是均在成熟初期(花后30 d)表达较高,而里那醇和萜品醇含量此时并不高,这可能与里那醇合成酶和萜品醇合酶还参与其他单萜化合物月桂烯、β-cis-罗勒烯和柠檬烯等合成有关[29,30,31]。T型架果实中Liner syn表达量远高于V型架,这与里那醇等化合物积累相一致。除了花后48 d的果实,T型架果实中Terp syn的表达也高于V型架,与化合物含量变化一致。
图4
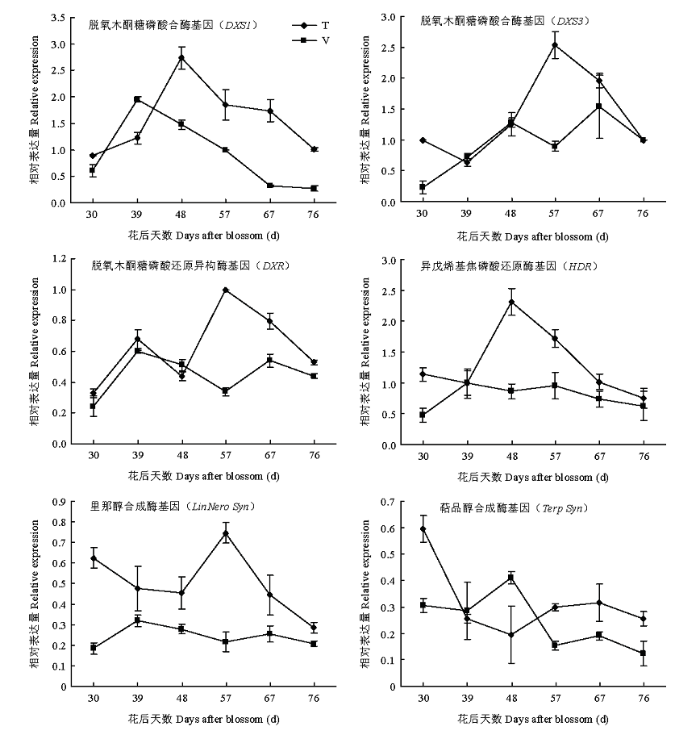 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4同架式葡萄果实成熟期间单萜合成途径基因表达变化
Fig. 4Changes of monoterpene biosynthesis pathway genes expression in grape berries cultivated in different trellis systems during maturity
4 结论
单萜积累与其代谢途径中多个关键酶基因表达具有紧密相关性,其合成受到各个基因转录水平的调控。T型架式栽培‘爱神玫瑰’葡萄果实单萜类化合物积累更多。(责任编辑 赵伶俐)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLMagsci [本文引用: 2]

单萜化合物是葡萄与葡萄酒中一类典型香气成分,以游离态和结合态形式存在。重点阐述了葡萄果实单萜化合物的生物合成途径及其关键酶——单萜合成酶的研究进展,葡萄果实与葡萄酒中的单萜化合物及其影响因素,糖苷结合态单萜化合物的结构、含量及其分析方法,并对单萜化合物的研究前景进行了展望。 <BR>
URLMagsci [本文引用: 2]
单萜化合物是葡萄与葡萄酒中一类典型香气成分,以游离态和结合态形式存在。重点阐述了葡萄果实单萜化合物的生物合成途径及其关键酶——单萜合成酶的研究进展,葡萄果实与葡萄酒中的单萜化合物及其影响因素,糖苷结合态单萜化合物的结构、含量及其分析方法,并对单萜化合物的研究前景进行了展望。 <BR>
DOI:10.1016/j.foodchem.2015.02.082URLPMID:25794740 [本文引用: 1]
C6 and C9 volatiles, originated from fatty acids, are important volatiles for ‘Cabernet Sauvignon’ grapes and wines. This study evaluated the influence of different training systems including Modified Vertical Shoot Positioned, (M-VSP); Fan training system with two trunks (F-TT); Fan training system with multiple trunks (F-MT) on these volatiles and the long-chain fatty acids (>C12) of grape berries and wines in the northwest of China. The expression profiles of genes from associated metabolic pathway were also analyzed. F-MT training resulted in lower vine vigor, larger yield, higher content of unsaturated fatty acids in grapes and lower C6 esters in wines in comparison with M-VSP and F-TT. M-VSP and F-TT enhanced C6 volatiles in grape berries. The concentrations of C6 volatiles were positively correlated with the expression of VvLOXA and VvHPL1. The results expanded the knowledge of the influence of training systems on fatty acids and their derived volatiles of grapes and wines.
[本文引用: 1]
DOI:10.1007/s00425-012-1704-0URLPMID:22824963 [本文引用: 1]
AbstractIn developing grapevine (Vitis vinifera L.) berries, precursor volatile organic compounds (PVOCs) are largely stored as glycosides which may be hydrolyzed to release VOCs during fruit ripening, wine making, or aging. VOCs can be further transformed by yeast metabolism. Together, these processes contribute to complexity of wine aromas. Floral and citrus odors of many white wine varietals are attributed to monoterpenes and monoterpene alcohols, while phenolic compounds, norisoprenoids, and other volatiles also play important roles in determining aroma. We present an analysis of PVOCs stored as glycosides in developing Gew眉rztraminer berries during the growing season. We optimized a method for PVOC analysis suitable for small amounts of Muscat grapevine berries and showed that the amount of PVOCs dramatically increased during and after v茅raison. Transcript profiling of the same berry samples underscored the involvement of terpenoid pathway genes in the accumulation of PVOCs. The onset of monoterpenol PVOC accumulation in developing grapes was correlated with an increase of transcript abundances of early terpenoid pathway enzymes. Transcripts encoding the methylerythritol phosphate pathway gene 4-hydroxy-3-methylbut-2-enyl diphosphate reductase, as well as geraniol diphosphate synthase, were up-regulated preceding and during the increase in monoterpenol PVOCs. Transcripts for linalool/nerolidol synthase increased in later v茅raison stages.
DOI:10.3390/molecules18078200URL [本文引用: 1]
DOI:10.1016/j.foodchem.2017.05.127URLPMID:28764010 [本文引用: 1]
Abstract Terpenes are important aroma compounds in table Muscat grape and wine, and their content in the berry can be affected by sunlight. The effects of sunlight exclusion on monoterpene profiles and relevant gene expression profiles in the exocarp and mesocarp of table Muscat grape 'Jingxiangyu' at different development stages were thoroughly surveyed by bagging pre-veraison clusters in special opaque boxes. The responses of monoterpenes to sunlight treatments varied in three types, representatively linalool, ocimene and geraniol. Linalool was the most sensitive compound to sunlight, whose biosynthesis was severely inhibited by sunlight exclusion and then was elevated by re-exposure. Ocimene and glycosylated geraniol showed a certain suppressive and stimulative responses to sunlight exclusion respectively. Further transcription analysis revealed that VvPNLinNer1, VvCSbOci, VvGT7 and VvGT14 genes were mainly responsible for monoterpene accumulation and sensitivity to sunlight. VvDXS2 and VvDXR genes were partially related to the differential accumulation of total terpenes under different sunlight treatments. Copyright 2017 Elsevier Ltd. All rights reserved.
DOI:10.1111/ajgw.12229URL [本文引用: 1]
Abstract Background and AimsInsolation of grape bunches has long been postulated to enhance the flavour of aromatic grape cultivars. This hypothesis was tested by combining gene expression and metabolic analysis of the monoterpene and flavonol synthesis pathways. Methods and ResultsGrape bunches were shaded or shaded and reilluminated to investigate the influence of light on the monoterpene and flavonol biosynthetic pathways. The expression of terpenoid and flavonol metabolic genes was measured by quantitative polymerase chain reaction under light and shade conditions during the ripening phase and compared to the accumulation of their respective metabolic products. Expression of flavonol synthase and flavonol glycosyltransferase genes was virtually absent in shaded bunches, but expression increased strongly upon reillumination, as did the flavonol content of the berries. The expression of the terpene synthase genes as well as the monoterpene content were greatly reduced in shaded bunches, and then increased upon reillumination. The expression of terpene glycosyltransferases was affected only slightly by light. ConclusionsThe results of this study show the positive influence of light on monoterpene and flavonol biosynthesis. Significance of the StudyOptimising the light exposure of grape berries enables the viticulturist to manipulate grape berry aroma and consequently the composition of wine.
[本文引用: 1]
URLMagsci [本文引用: 1]
<p>摘要:以三年生晚红葡萄为试验材料,研究了不同架式及负载量对果实品质的影响。结果表明:立体棚架栽培的果实单粒重、可溶性固形物含量、固酸比等品质指标均高于平棚架和篱架,总酸含量低于二者;立体棚架叶片净光合速率(Pn)极显著高于棚架和篱架(P<0.01),并且叶片Pn与果实单粒重、总糖含量呈显著正相关。单粒重与总糖、固酸比呈极显著正相关,与总酸呈显著负相关。总糖与固酸比呈极显著正相关。总酸与固酸比呈极显著负相关。立体棚架保留8-10个/m2新梢对其光能利用和果实品质较适宜。</p>
URLMagsci [本文引用: 1]
<p>摘要:以三年生晚红葡萄为试验材料,研究了不同架式及负载量对果实品质的影响。结果表明:立体棚架栽培的果实单粒重、可溶性固形物含量、固酸比等品质指标均高于平棚架和篱架,总酸含量低于二者;立体棚架叶片净光合速率(Pn)极显著高于棚架和篱架(P<0.01),并且叶片Pn与果实单粒重、总糖含量呈显著正相关。单粒重与总糖、固酸比呈极显著正相关,与总酸呈显著负相关。总糖与固酸比呈极显著正相关。总酸与固酸比呈极显著负相关。立体棚架保留8-10个/m2新梢对其光能利用和果实品质较适宜。</p>
URL [本文引用: 1]
[目的]探索适宜当地"赤霞珠"葡萄优质、高效生产的架式与树形.[方法]以2年生"赤霞珠"葡萄品种为试材,研究了传统"篱架"、"V"型架对伊犁67团"赤霞珠"葡萄节间生长情况、果实相关品质的影响.[结果]采用"V"型架的整形方式可显著增加结果枝的节间长度,降低节间粗度;增加可溶性固形物、总酚、总花色素含量,降低可滴定酸和单宁含量;显著地增加单穗重、单粒重、果实纵横径、每棵树结果数.[结论]"V"型架在品质、产量均优于篱架,对当地酿酒葡萄的发展具有一定的指导和理论意义.
URL [本文引用: 1]
[目的]探索适宜当地"赤霞珠"葡萄优质、高效生产的架式与树形.[方法]以2年生"赤霞珠"葡萄品种为试材,研究了传统"篱架"、"V"型架对伊犁67团"赤霞珠"葡萄节间生长情况、果实相关品质的影响.[结果]采用"V"型架的整形方式可显著增加结果枝的节间长度,降低节间粗度;增加可溶性固形物、总酚、总花色素含量,降低可滴定酸和单宁含量;显著地增加单穗重、单粒重、果实纵横径、每棵树结果数.[结论]"V"型架在品质、产量均优于篱架,对当地酿酒葡萄的发展具有一定的指导和理论意义.
DOI:10.3864/j.issn.0578-1752.2016.21.019URL [本文引用: 1]
[目的]探讨直立叶幕和水平叶幕对‘摩尔多瓦’葡萄果穗微域环境及果实品质的影响,为建立合理的葡萄栽培架式提供参考.[方法]2013年对部分篱架栽培的‘摩尔多瓦’进行架式改造,培养为棚架水平叶幕,与篱架直立叶幕相比较进行研究,其他栽培管理技术一致.2 01 5年6月1日开始,通过温湿度记录仪实时监控直立叶幕和水平叶幕葡萄果穗周围微域环境的温度和湿度,并用光合辐射检测仪实时记录水平叶幕上下及直立叶幕两侧所接受的光合有效辐射.于花后9周开始至果实成熟,每两周采摘两种叶幕类型下的果实,测定发育过程中果实的各项品质指标,分析两种叶幕类型对果穗微域环境和果实品质的影响.[结果]与直立叶幕相比,水平叶幕显著降低了6-9月的果实表面温度、35℃以上高温比例,其中7月份水平叶幕果穗周围35℃以上高温比例比直立叶幕的降低68.97%,8月份水平叶幕未出现35℃以上高温,而直立叶幕高温达41.6℃;同时,水平叶幕增大了果穗微域环境的湿度,减小了其波动,其上部10:00光合有效辐射比直立叶幕东侧低25.8%,而下午14:00比直立叶幕西侧低5.95%,仅中午12:00所受光合有效辐射高于直立叶幕,但水平叶幕下部全天接受光合有效辐射均较低.在果实品质方面,水平叶幕使果实第二次生长量和膨大速率显著增加,其成熟果实纵横径、果形指数及粒重均高于直立叶幕;且水平叶幕提高了成熟果实的糖、酸、总酚及花色苷含量,2015年果实成熟时水平叶幕下果实的还原糖、总酸、总酚及果皮花色苷含量分别比直立叶幕提高了2.40%、4.35%、5.55%及7.04%,同时,水平叶幕使果皮亮度显著提高,红蓝色度加深,即水平叶幕改善了‘摩尔多瓦’的果实色泽.[结论]与直立叶幕相比,水平叶幕改善了‘摩尔多瓦’果实周围的微环境,提高了果实品质.
DOI:10.3864/j.issn.0578-1752.2016.21.019URL [本文引用: 1]
[目的]探讨直立叶幕和水平叶幕对‘摩尔多瓦’葡萄果穗微域环境及果实品质的影响,为建立合理的葡萄栽培架式提供参考.[方法]2013年对部分篱架栽培的‘摩尔多瓦’进行架式改造,培养为棚架水平叶幕,与篱架直立叶幕相比较进行研究,其他栽培管理技术一致.2 01 5年6月1日开始,通过温湿度记录仪实时监控直立叶幕和水平叶幕葡萄果穗周围微域环境的温度和湿度,并用光合辐射检测仪实时记录水平叶幕上下及直立叶幕两侧所接受的光合有效辐射.于花后9周开始至果实成熟,每两周采摘两种叶幕类型下的果实,测定发育过程中果实的各项品质指标,分析两种叶幕类型对果穗微域环境和果实品质的影响.[结果]与直立叶幕相比,水平叶幕显著降低了6-9月的果实表面温度、35℃以上高温比例,其中7月份水平叶幕果穗周围35℃以上高温比例比直立叶幕的降低68.97%,8月份水平叶幕未出现35℃以上高温,而直立叶幕高温达41.6℃;同时,水平叶幕增大了果穗微域环境的湿度,减小了其波动,其上部10:00光合有效辐射比直立叶幕东侧低25.8%,而下午14:00比直立叶幕西侧低5.95%,仅中午12:00所受光合有效辐射高于直立叶幕,但水平叶幕下部全天接受光合有效辐射均较低.在果实品质方面,水平叶幕使果实第二次生长量和膨大速率显著增加,其成熟果实纵横径、果形指数及粒重均高于直立叶幕;且水平叶幕提高了成熟果实的糖、酸、总酚及花色苷含量,2015年果实成熟时水平叶幕下果实的还原糖、总酸、总酚及果皮花色苷含量分别比直立叶幕提高了2.40%、4.35%、5.55%及7.04%,同时,水平叶幕使果皮亮度显著提高,红蓝色度加深,即水平叶幕改善了‘摩尔多瓦’的果实色泽.[结论]与直立叶幕相比,水平叶幕改善了‘摩尔多瓦’果实周围的微环境,提高了果实品质.
URL [本文引用: 1]
为了研究水平叶幕和直立叶幕所构成的微域环境对套袋葡萄果实花色苷代谢的影响,分别以棚架、篱架代表水平叶幕和直立叶幕,连续两年对棚架和篱架的‘摩尔多瓦’葡萄进行套袋,于果实膨大期开始实时监控果穗袋内微环境的温度和湿度;从转色期至果实成熟,测定不同发育阶段浆果果皮花色苷单体组分与含量,以及果皮花色苷代谢途径相关基因表达量与酶活性,分析果实品质差异。结果表明,2015年、2016年棚架水平叶幕下套袋果实袋内的高温极值及高温比例和湿度波动幅度与篱架直立叶幕套袋相比均有明显降低;连续两年成熟果实还原糖含量分别比篱架套袋的高18.33%和15.41%,且棚架的果实酸度较低;2015年测定棚架套袋果实的单宁及花色苷含量分别比篱架套袋的提高22.32%和35.29%,果实果皮红色度显著提高,而2016年夏季较冷凉,棚架套袋果实的总酚、黄烷醇、类黄酮含量分别比篱架套袋降低10.72%、10.97%和45.04%。2015年花后77 d,棚架套袋果实的花色苷单体含量较高,但到花后91 d时,篱架套袋果实花色苷单体含量补偿性增加。成熟果实果皮中均检测到21种花色苷单体,棚架套袋葡萄花色苷双糖苷化、甲基化及总修饰度均高于篱架套袋果实,但有19种花色苷单体含量低于篱架套袋果实。花后63~105 d,除Vv OMT外,棚架与篱架套袋葡萄花色苷代谢途径中Vv UFGT、Vv LDOX、Vv5GT和Vv PPO基因表达变化趋势一致,但相关酶活性差异较大。由此可见,水平叶幕可降低果实微域环境的温湿度,增加套袋果实品质及花色苷单体修饰程度,但降低花色苷单体含量,并影响花色苷代谢途径中相关酶基因的表达量及活性。
URL [本文引用: 1]
为了研究水平叶幕和直立叶幕所构成的微域环境对套袋葡萄果实花色苷代谢的影响,分别以棚架、篱架代表水平叶幕和直立叶幕,连续两年对棚架和篱架的‘摩尔多瓦’葡萄进行套袋,于果实膨大期开始实时监控果穗袋内微环境的温度和湿度;从转色期至果实成熟,测定不同发育阶段浆果果皮花色苷单体组分与含量,以及果皮花色苷代谢途径相关基因表达量与酶活性,分析果实品质差异。结果表明,2015年、2016年棚架水平叶幕下套袋果实袋内的高温极值及高温比例和湿度波动幅度与篱架直立叶幕套袋相比均有明显降低;连续两年成熟果实还原糖含量分别比篱架套袋的高18.33%和15.41%,且棚架的果实酸度较低;2015年测定棚架套袋果实的单宁及花色苷含量分别比篱架套袋的提高22.32%和35.29%,果实果皮红色度显著提高,而2016年夏季较冷凉,棚架套袋果实的总酚、黄烷醇、类黄酮含量分别比篱架套袋降低10.72%、10.97%和45.04%。2015年花后77 d,棚架套袋果实的花色苷单体含量较高,但到花后91 d时,篱架套袋果实花色苷单体含量补偿性增加。成熟果实果皮中均检测到21种花色苷单体,棚架套袋葡萄花色苷双糖苷化、甲基化及总修饰度均高于篱架套袋果实,但有19种花色苷单体含量低于篱架套袋果实。花后63~105 d,除Vv OMT外,棚架与篱架套袋葡萄花色苷代谢途径中Vv UFGT、Vv LDOX、Vv5GT和Vv PPO基因表达变化趋势一致,但相关酶活性差异较大。由此可见,水平叶幕可降低果实微域环境的温湿度,增加套袋果实品质及花色苷单体修饰程度,但降低花色苷单体含量,并影响花色苷代谢途径中相关酶基因的表达量及活性。
DOI:10.3321/j.issn:0513-353X.2005.01.019URLMagsci [本文引用: 2]
以7年生棚架和篱架栽培模式下的玫瑰香葡萄为试材, 采用气相色谱—质谱联机分析了挥发性物质成分相对含量在成熟期的变化。结果表明, 随着采收期推迟, 两种栽培模式下葡萄的酯类物质相对含量均明显增加, 酮醛类化合物相对含量减少, 萜类物质总的相对含量也有下降趋势; 不同形态萜类物质相对含量比例因采收时间不同而有差异, 前期结合态比例明显高于游离态, 后期两者比例相当。篱架葡萄成熟后期酯类物质相对含量高于棚架, 而酮醛类物质和萜类物质相对含量低于棚架。桉叶油素主要以结合态存在, β - 月桂烯则主要以游离态存在, 两种物质在棚架葡萄上的相对含量明显高于篱架。
DOI:10.3321/j.issn:0513-353X.2005.01.019URLMagsci [本文引用: 2]
以7年生棚架和篱架栽培模式下的玫瑰香葡萄为试材, 采用气相色谱—质谱联机分析了挥发性物质成分相对含量在成熟期的变化。结果表明, 随着采收期推迟, 两种栽培模式下葡萄的酯类物质相对含量均明显增加, 酮醛类化合物相对含量减少, 萜类物质总的相对含量也有下降趋势; 不同形态萜类物质相对含量比例因采收时间不同而有差异, 前期结合态比例明显高于游离态, 后期两者比例相当。篱架葡萄成熟后期酯类物质相对含量高于棚架, 而酮醛类物质和萜类物质相对含量低于棚架。桉叶油素主要以结合态存在, β - 月桂烯则主要以游离态存在, 两种物质在棚架葡萄上的相对含量明显高于篱架。
URLMagsci [本文引用: 2]
以6 年生玫瑰香葡萄为试材,研究了Y 型架和篱架不同叶幕层的光照强度和产量分布,测定了果实品质和芳香化合物。结果表明:两种架型的光照强度与叶幕层高度、宽度分布有显著的相关性,Y 型架在高0.2 ~ 0.6 m 和1.8 m 处的光照强度优于篱架;但在1.0 ~ 1.4 m 则低于篱架。Y 型架61.61%的产量集中在定植行两侧0.3 m,高度1.0 ~ 1.4 m 的范围内;篱架55.90%的产量集中在定植行两侧0.15 m,高度0.6 ~ 1.0 m 的范围内。Y 型架果实总糖和抗坏血酸含量分别为16.13%和8.72 mg · kg<sup>-1</sup>,显著高于篱架(14.54%和7.21 mg · kg<sup>-1</sup>);Y 型架果皮原花色素含量为4.72 mg · g<sup>-1</sup>,显著低于篱架(6.30 mg · g<sup>-1</sup>)。Y 型架和篱架果实中检测到芳香化合物种类为32 种和30 种;萜醇化合物的相对含量分别为32.91%和6.33%;其中,Y 型架果实中里那醇的相对含量是篱架的5.7 倍,可见,Y 型架葡萄果实风味优于篱架。
URLMagsci [本文引用: 2]
以6 年生玫瑰香葡萄为试材,研究了Y 型架和篱架不同叶幕层的光照强度和产量分布,测定了果实品质和芳香化合物。结果表明:两种架型的光照强度与叶幕层高度、宽度分布有显著的相关性,Y 型架在高0.2 ~ 0.6 m 和1.8 m 处的光照强度优于篱架;但在1.0 ~ 1.4 m 则低于篱架。Y 型架61.61%的产量集中在定植行两侧0.3 m,高度1.0 ~ 1.4 m 的范围内;篱架55.90%的产量集中在定植行两侧0.15 m,高度0.6 ~ 1.0 m 的范围内。Y 型架果实总糖和抗坏血酸含量分别为16.13%和8.72 mg · kg<sup>-1</sup>,显著高于篱架(14.54%和7.21 mg · kg<sup>-1</sup>);Y 型架果皮原花色素含量为4.72 mg · g<sup>-1</sup>,显著低于篱架(6.30 mg · g<sup>-1</sup>)。Y 型架和篱架果实中检测到芳香化合物种类为32 种和30 种;萜醇化合物的相对含量分别为32.91%和6.33%;其中,Y 型架果实中里那醇的相对含量是篱架的5.7 倍,可见,Y 型架葡萄果实风味优于篱架。
DOI:10.1111/j.1750-3841.2008.00736.xURL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 3]
DOI:10.3864/j.issn.0578-1752.2014.07.015URLMagsci [本文引用: 2]
【目的】检测‘亚历山大’葡萄果实中萜类物质的种类和含量,分析果实发育过程中单萜类物质的积累与相关基因表达的关系,为揭示玫瑰香型葡萄香气物质的积累规律和香味育种奠定基础。【方法】从果实转色后至成熟期,每周取样一次,每次随机取样80—100粒,去籽去梗后榨汁,离心取上清液用于香气物质测定,使用顶空固相微萃取方法萃取样品中的挥发性成分,对采集到的质谱图用NIST 05谱库检索,并根据已有标样的色谱保留时间和保留指数,确定香气成分的化学组成,利用已有的化合物制备标准曲线进行定量。根据DXS的ORF序列设计引物,以充分成熟的样品RNA反转录获得的cDNA为模板,通过多次独立PCR扩增获得‘亚历山大’DXS的ORF片段,根据测序结果分析单核苷酸多态性位点。根据萜类生物合成途径中的8个关键酶基因序列设计引物,使用Ubiquitin,EF1-α和GAPDH 3个看家基因做为内参,进行实时定量PCR,检测这些基因的转录丰度。【结果】从转色后至成熟期,在检测到的所有单萜中里那醇和香叶醇的含量远高于其它单萜,多种萜类浓度逐渐上升,其中里那醇、月桂烯、柠檬烯上升6—8倍,香叶醇、萜品醇、香叶醛和萜品油烯上升2—3倍,玫瑰醚和橙花醛含量变化较小,果实中里那醇、香叶醇、氧化玫瑰和月桂烯的含量高于嗅闻阈值。在单萜生物合成早期途径中,DXS1的表达从转色开始缓慢上升,在花后15周迅速上调,上升约5倍。DXS3则从12周开始迅速上升。DXR表现出波动变化规律。HDR的表达量在前期合成酶基因中是最高的,能达到DXS1、DXS3、DXR的10—20倍;在合成途径中期的2个基因FPPS和GPPS,从转色后至成熟期表达量持续上调,FPPS的表达量上升了4倍,GPPS的表达量上升了2倍;在合成途径的后期,Liner-syn和Terp-syn在花后的11—15周都出现表达量升高,所有基因在花后17周出现明显的表达下降。单萜总量从花后12—16周迅速积累,这与同期DXS3、DXR、HDR、GPPS、FPPS的表达上调的趋势相一致。通过相关性分析结果显示萜类总量与DXS3的相关系数为0.831。‘亚历山大’DXS1的cDNA的开放阅读框全长2 151 bp,编码716个氨基酸,通过多次独立克隆后对测序结果进行比对,在该序列中发现了16个单核苷酸多态性位点,导致4个位置编码的氨基酸发生变化。【结论】单萜合成途径中多个关键酶基因后期表达上调,导致成熟过程中单萜化合物含量上升2—8倍,DXS3与单萜总量的积累具有显著的相关性。
DOI:10.3864/j.issn.0578-1752.2014.07.015URLMagsci [本文引用: 2]
【目的】检测‘亚历山大’葡萄果实中萜类物质的种类和含量,分析果实发育过程中单萜类物质的积累与相关基因表达的关系,为揭示玫瑰香型葡萄香气物质的积累规律和香味育种奠定基础。【方法】从果实转色后至成熟期,每周取样一次,每次随机取样80—100粒,去籽去梗后榨汁,离心取上清液用于香气物质测定,使用顶空固相微萃取方法萃取样品中的挥发性成分,对采集到的质谱图用NIST 05谱库检索,并根据已有标样的色谱保留时间和保留指数,确定香气成分的化学组成,利用已有的化合物制备标准曲线进行定量。根据DXS的ORF序列设计引物,以充分成熟的样品RNA反转录获得的cDNA为模板,通过多次独立PCR扩增获得‘亚历山大’DXS的ORF片段,根据测序结果分析单核苷酸多态性位点。根据萜类生物合成途径中的8个关键酶基因序列设计引物,使用Ubiquitin,EF1-α和GAPDH 3个看家基因做为内参,进行实时定量PCR,检测这些基因的转录丰度。【结果】从转色后至成熟期,在检测到的所有单萜中里那醇和香叶醇的含量远高于其它单萜,多种萜类浓度逐渐上升,其中里那醇、月桂烯、柠檬烯上升6—8倍,香叶醇、萜品醇、香叶醛和萜品油烯上升2—3倍,玫瑰醚和橙花醛含量变化较小,果实中里那醇、香叶醇、氧化玫瑰和月桂烯的含量高于嗅闻阈值。在单萜生物合成早期途径中,DXS1的表达从转色开始缓慢上升,在花后15周迅速上调,上升约5倍。DXS3则从12周开始迅速上升。DXR表现出波动变化规律。HDR的表达量在前期合成酶基因中是最高的,能达到DXS1、DXS3、DXR的10—20倍;在合成途径中期的2个基因FPPS和GPPS,从转色后至成熟期表达量持续上调,FPPS的表达量上升了4倍,GPPS的表达量上升了2倍;在合成途径的后期,Liner-syn和Terp-syn在花后的11—15周都出现表达量升高,所有基因在花后17周出现明显的表达下降。单萜总量从花后12—16周迅速积累,这与同期DXS3、DXR、HDR、GPPS、FPPS的表达上调的趋势相一致。通过相关性分析结果显示萜类总量与DXS3的相关系数为0.831。‘亚历山大’DXS1的cDNA的开放阅读框全长2 151 bp,编码716个氨基酸,通过多次独立克隆后对测序结果进行比对,在该序列中发现了16个单核苷酸多态性位点,导致4个位置编码的氨基酸发生变化。【结论】单萜合成途径中多个关键酶基因后期表达上调,导致成熟过程中单萜化合物含量上升2—8倍,DXS3与单萜总量的积累具有显著的相关性。
DOI:10.3969/j.issn.2095-1191.2017.05.018URL [本文引用: 2]
[目的]探讨V形篱架及T形架栽培模式对酿酒葡萄凌丰生长及产量的影响,为南方湿热地区推广种植酿酒葡萄提供参考依据.[方法]通过田间观察,记录V形篱架和T形架栽培凌丰葡萄的物候期、树体生长势和结果习性,测定、分析其果实产量和品质.[结果]V形篱架栽培模式下,凌丰葡萄的萌芽期、开花期、果实转色期和果实成熟期均早于T形架3~5 d.一年两收栽培模式下,第一茬夏果V形篱架栽培的成枝率和结果枝率分别为75.8%和67.2%,分别显著低于T形架栽培1.8%(绝对值,下同)和5.1%(P0.05,下同),结果枝率为82.1%,显著高于V形篱架栽培8.1%,产量比V形篱架栽培高3.98%,但差异不显著.在两茬果测量光照强度的低处(70和100 cm)和高处(100和150 cm),V形篱架栽培的自然光照强度均显著强于T形架栽培,两种架形栽培凌丰葡萄的可溶性固形物含量差异不显著.[结论]以V形篱架栽培的酿酒葡萄凌丰枝叶分布较均匀,园内相对光照强度较高,有利于提高葡萄产量和品质,在南方湿热地区酿酒葡萄产区具有一定的推广应用价值.
DOI:10.3969/j.issn.2095-1191.2017.05.018URL [本文引用: 2]
[目的]探讨V形篱架及T形架栽培模式对酿酒葡萄凌丰生长及产量的影响,为南方湿热地区推广种植酿酒葡萄提供参考依据.[方法]通过田间观察,记录V形篱架和T形架栽培凌丰葡萄的物候期、树体生长势和结果习性,测定、分析其果实产量和品质.[结果]V形篱架栽培模式下,凌丰葡萄的萌芽期、开花期、果实转色期和果实成熟期均早于T形架3~5 d.一年两收栽培模式下,第一茬夏果V形篱架栽培的成枝率和结果枝率分别为75.8%和67.2%,分别显著低于T形架栽培1.8%(绝对值,下同)和5.1%(P0.05,下同),结果枝率为82.1%,显著高于V形篱架栽培8.1%,产量比V形篱架栽培高3.98%,但差异不显著.在两茬果测量光照强度的低处(70和100 cm)和高处(100和150 cm),V形篱架栽培的自然光照强度均显著强于T形架栽培,两种架形栽培凌丰葡萄的可溶性固形物含量差异不显著.[结论]以V形篱架栽培的酿酒葡萄凌丰枝叶分布较均匀,园内相对光照强度较高,有利于提高葡萄产量和品质,在南方湿热地区酿酒葡萄产区具有一定的推广应用价值.
DOI:10.3864/j.issn.0578-1752.2015.19.018URL [本文引用: 1]
【目的】早熟、具有玫瑰香味的鲜食葡萄具有较广阔的市场前景,了解各类香气组分及相关代谢途径在果实发育过程中的变化规律,把握香气组分形成的关键时期,为早熟葡萄品种品质调控技术的提出提供理论依据。【方法】以‘贝达’嫁接的4年生早熟葡萄品种‘瑞都香玉’为试材,使用顶空固相微萃取结合气相色谱与质谱联用技术,测定在幼果发育期(花后第3周)至过熟期(花后第12周)果实中挥发性组分的变化。根据各组分的变化探究相关代谢途径的更替,并利用主成分分析确定发育阶段与各类香气组分间的关系。【结果】根据可溶性固形物及可滴定酸含量的变化可以确定‘瑞都香玉’葡萄在花后第5周即可进入果实转色/软化期,花后第9周进入果实成熟期。里那醇、橙花醇、香叶醇、香茅醇、萜品醇等主要呈香型萜烯类组分峰值出现的时期不同,橙花醇、香叶醇含量在成熟初期达到最大值,里那醇、萜品醇含量在成熟中期达到峰值,香茅醇含量在成熟期具有波动性。E-2-己烯醛、己醛为成熟期主要的C6化合物,其含量在转色期以后即可达到峰值,成熟期时含量降低;Z-3-己烯醛主要出现在幼果发育期,转色期时含量处于较低水平。β-紫罗兰酮、大马士酮等降异戊二烯衍生物及苯甲醛等芳香族化合物在成熟期时含量处于较低水平或消失。综合各挥发性组分的含量变化可得,萜烯类组分在各发育期均有积累,过熟期含量降低;酯类物质集中出现在幼果发育期,成熟初期含量有所升高;醛酮类物质集中出现在转色初期;C6化合物在转色期有不同程度的升高,成熟期后含量有减少趋势;醇类物质在转为成熟期后含量开始减少;芳香族化合物集中在转色期以前;酸类物质在转为成熟期后有所增加。转色期以前为酯类物质、芳香族化合物变化的关键时期,转色期为醛酮类物质变化的关键时期,转色期至成熟期为成熟期特征萜烯类物质、C6化合物、酸类物质变化的关键时期。【结论】果实发育早期亚麻酸裂解途径Z-3-己烯醛支路活跃,进入转色期后,脂肪酸代谢其他支路与萜烯类物质合成路径活跃性增强;成熟期发育过程中,主要香气成分的积累都有减弱,转色期转为成熟期时为果实香型形成的关键时期。‘瑞都香玉’主要呈香型萜烯成分的积累具有差异性,里那醇、萜品醇含量在成熟期仍有增加,综合考虑C6组分与萜烯类成分的变化,果实香气成熟期稍晚于生理成熟期,适当延迟采收有助于其特征香型的形成。
DOI:10.3864/j.issn.0578-1752.2015.19.018URL [本文引用: 1]
【目的】早熟、具有玫瑰香味的鲜食葡萄具有较广阔的市场前景,了解各类香气组分及相关代谢途径在果实发育过程中的变化规律,把握香气组分形成的关键时期,为早熟葡萄品种品质调控技术的提出提供理论依据。【方法】以‘贝达’嫁接的4年生早熟葡萄品种‘瑞都香玉’为试材,使用顶空固相微萃取结合气相色谱与质谱联用技术,测定在幼果发育期(花后第3周)至过熟期(花后第12周)果实中挥发性组分的变化。根据各组分的变化探究相关代谢途径的更替,并利用主成分分析确定发育阶段与各类香气组分间的关系。【结果】根据可溶性固形物及可滴定酸含量的变化可以确定‘瑞都香玉’葡萄在花后第5周即可进入果实转色/软化期,花后第9周进入果实成熟期。里那醇、橙花醇、香叶醇、香茅醇、萜品醇等主要呈香型萜烯类组分峰值出现的时期不同,橙花醇、香叶醇含量在成熟初期达到最大值,里那醇、萜品醇含量在成熟中期达到峰值,香茅醇含量在成熟期具有波动性。E-2-己烯醛、己醛为成熟期主要的C6化合物,其含量在转色期以后即可达到峰值,成熟期时含量降低;Z-3-己烯醛主要出现在幼果发育期,转色期时含量处于较低水平。β-紫罗兰酮、大马士酮等降异戊二烯衍生物及苯甲醛等芳香族化合物在成熟期时含量处于较低水平或消失。综合各挥发性组分的含量变化可得,萜烯类组分在各发育期均有积累,过熟期含量降低;酯类物质集中出现在幼果发育期,成熟初期含量有所升高;醛酮类物质集中出现在转色初期;C6化合物在转色期有不同程度的升高,成熟期后含量有减少趋势;醇类物质在转为成熟期后含量开始减少;芳香族化合物集中在转色期以前;酸类物质在转为成熟期后有所增加。转色期以前为酯类物质、芳香族化合物变化的关键时期,转色期为醛酮类物质变化的关键时期,转色期至成熟期为成熟期特征萜烯类物质、C6化合物、酸类物质变化的关键时期。【结论】果实发育早期亚麻酸裂解途径Z-3-己烯醛支路活跃,进入转色期后,脂肪酸代谢其他支路与萜烯类物质合成路径活跃性增强;成熟期发育过程中,主要香气成分的积累都有减弱,转色期转为成熟期时为果实香型形成的关键时期。‘瑞都香玉’主要呈香型萜烯成分的积累具有差异性,里那醇、萜品醇含量在成熟期仍有增加,综合考虑C6组分与萜烯类成分的变化,果实香气成熟期稍晚于生理成熟期,适当延迟采收有助于其特征香型的形成。
URLMagsci [本文引用: 1]
香气是葡萄酒的重要感官质量指标之一。葡萄酒中香气物质主要受葡萄品种、生态条件、果实成熟质量及酿造工艺技术等因素影响,香气物质的种类和含量对葡萄酒的风格和典型性起决定作用。本文详细阐述了葡萄与葡萄酒中各类香气物质及影响其形成和组成的各种因素,以期为葡萄酒工业化生产提供理论指导。
URLMagsci [本文引用: 1]
香气是葡萄酒的重要感官质量指标之一。葡萄酒中香气物质主要受葡萄品种、生态条件、果实成熟质量及酿造工艺技术等因素影响,香气物质的种类和含量对葡萄酒的风格和典型性起决定作用。本文详细阐述了葡萄与葡萄酒中各类香气物质及影响其形成和组成的各种因素,以期为葡萄酒工业化生产提供理论指导。
DOI:10.1016/j.foodchem.2014.01.005URLPMID:24518327 [本文引用: 1]
Aroma is an important quality characteristic in Muscat grapes and constitutes a major concern for viticulturist and grapevine breeders. For this reason, Muscat aroma variability was characterised in a segregating progeny and in a collection of table grapes, to assess the usefulness of the presence or absence of rose oxide for predicting Muscat genotypes. Simple tasting and an analysis of free and bound aroma compounds, including rose oxide, linalool oxide, linalool, -terpineol, citronellol, nerol, geraniol, benzyl alcohol and 2-phenylethanol, were carried out. The association between Muscat score and the compounds considered as active odorants according to their odour activity values was also evaluated. The results obtained pointed to a highly significant correlation between the presence/absence of rose oxide in grapes and the presence/absence of Muscat aroma. Thus, this analysis could be a useful tool for identifying Muscat cultivars in a more objective way than sensory analysis.
DOI:10.13925/j.cnki.gsxb.20160195URL [本文引用: 1]
【目的】探讨不同类型果袋对‘阳光玫瑰’葡萄香气组分及合成相关基因表达的影响,为葡萄生产栽培选择适宜的果袋提供理论指导。【方法】以‘阳光玫瑰’葡萄为材料,套普通单层白纸袋、透明微孔塑料袋、双层外黄内黑纸袋和单层绿纸袋4种不同果袋,以普通单层白纸袋为对照(以下简称白袋、透明袋、黑袋和绿袋)。采用实时荧光定量PCR法分析脱氧木酮糖磷酸合酶基因(DXS3)、香叶基二磷酸合酶基因(GPPS)、里那醇合成酶基因(LIS)的表达变化,利用GC-MS技术测定葡萄香气组分。【结果】与对照相比,套透明袋果实较大,可滴定酸含量偏低,但可溶性固形物含量高,糖酸比高,差异显著;套黑纸袋和绿袋果实可溶性固形物含量较低。套透明袋果实中单萜类物质特征香气里那醇含量丰富,香叶醇、橙花醇、萜品醇等辅助类香气含量也很高,玫瑰香气浓郁;而套黑袋果实则特征香气含量最少;套绿袋果实香气物质种类少。套透明袋果实DXS3、GPPS和LIS基因表达量上调,促进单萜物质生成,单萜物质含量丰富,而套黑袋抑制了基因表达,推迟了基因表达高峰,影响单萜物质生成。套绿袋果实和对照相比基因表达量没有显著性差异。【结论】套不同的果袋对‘阳光玫瑰’葡萄果实香气组分及合成相关基因表达量的变化有显著的差异。
DOI:10.13925/j.cnki.gsxb.20160195URL [本文引用: 1]
【目的】探讨不同类型果袋对‘阳光玫瑰’葡萄香气组分及合成相关基因表达的影响,为葡萄生产栽培选择适宜的果袋提供理论指导。【方法】以‘阳光玫瑰’葡萄为材料,套普通单层白纸袋、透明微孔塑料袋、双层外黄内黑纸袋和单层绿纸袋4种不同果袋,以普通单层白纸袋为对照(以下简称白袋、透明袋、黑袋和绿袋)。采用实时荧光定量PCR法分析脱氧木酮糖磷酸合酶基因(DXS3)、香叶基二磷酸合酶基因(GPPS)、里那醇合成酶基因(LIS)的表达变化,利用GC-MS技术测定葡萄香气组分。【结果】与对照相比,套透明袋果实较大,可滴定酸含量偏低,但可溶性固形物含量高,糖酸比高,差异显著;套黑纸袋和绿袋果实可溶性固形物含量较低。套透明袋果实中单萜类物质特征香气里那醇含量丰富,香叶醇、橙花醇、萜品醇等辅助类香气含量也很高,玫瑰香气浓郁;而套黑袋果实则特征香气含量最少;套绿袋果实香气物质种类少。套透明袋果实DXS3、GPPS和LIS基因表达量上调,促进单萜物质生成,单萜物质含量丰富,而套黑袋抑制了基因表达,推迟了基因表达高峰,影响单萜物质生成。套绿袋果实和对照相比基因表达量没有显著性差异。【结论】套不同的果袋对‘阳光玫瑰’葡萄果实香气组分及合成相关基因表达量的变化有显著的差异。
DOI:10.7685/jnau.201603020URL [本文引用: 1]
[目的]研究不同质量浓度细胞分类素(CPPU)对葡萄果实香气组分及合成相关基因表达的影响,为阐明香气物质的合成提供理论依据。[方法]以‘阳光玫瑰’葡萄为材料,在花期时用25 mg·L^-1GA3处理花序,花后2周时用25 mg·L^-1GA3+0 mg·L^-1CPPU处理果穗作为对照(CK),设置25 mg·L^-1GA3+5 mg·L^-1CPPU(Ⅰ)、25 mg·L^-1GA3+10 mg·L^-1CPPU(Ⅱ)、25 mg·L^-1GA3+15 mg·L^-1CPPU(Ⅲ)3个处理,采用实时荧光定量PCR法研究香气合成相关基因脱氧木酮糖磷酸合酶基因(DXS3)、香叶基二磷酸合酶基因(GPPS)和里那醇合成酶基因(LIS)的表达变化,并利用GC-MS技术测定葡萄香气组分。[结果]与对照相比,处理Ⅰ和Ⅱ的果实显著增大、增重,可滴定酸含量和可溶性固形物有所下降,外观品质提高,处理Ⅲ因CPPU浓度过高而引起葡萄品质下降。与对照相比,香气种类减少,萜类物质特征香气里那醇、顺式氧化里那醇、香叶醇等相对含量降低,风味变淡,且CPPU浓度过高,不利于香气的产生。与对照相比,经不同浓度CPPU处理的果实中香气合成相关基因DXS3、GPPS、LIS表达量高峰推迟且峰值下调,浓度越高,推迟时间越久,峰值更低,基因表达受抑制,这与萜类物质相对含量降低相一致。[结论]25 mg·L^-1GA3+5 mg·L^-1CPPU处理能够改善‘阳光玫瑰’葡萄外观品质并保证内在品质。
DOI:10.7685/jnau.201603020URL [本文引用: 1]
[目的]研究不同质量浓度细胞分类素(CPPU)对葡萄果实香气组分及合成相关基因表达的影响,为阐明香气物质的合成提供理论依据。[方法]以‘阳光玫瑰’葡萄为材料,在花期时用25 mg·L^-1GA3处理花序,花后2周时用25 mg·L^-1GA3+0 mg·L^-1CPPU处理果穗作为对照(CK),设置25 mg·L^-1GA3+5 mg·L^-1CPPU(Ⅰ)、25 mg·L^-1GA3+10 mg·L^-1CPPU(Ⅱ)、25 mg·L^-1GA3+15 mg·L^-1CPPU(Ⅲ)3个处理,采用实时荧光定量PCR法研究香气合成相关基因脱氧木酮糖磷酸合酶基因(DXS3)、香叶基二磷酸合酶基因(GPPS)和里那醇合成酶基因(LIS)的表达变化,并利用GC-MS技术测定葡萄香气组分。[结果]与对照相比,处理Ⅰ和Ⅱ的果实显著增大、增重,可滴定酸含量和可溶性固形物有所下降,外观品质提高,处理Ⅲ因CPPU浓度过高而引起葡萄品质下降。与对照相比,香气种类减少,萜类物质特征香气里那醇、顺式氧化里那醇、香叶醇等相对含量降低,风味变淡,且CPPU浓度过高,不利于香气的产生。与对照相比,经不同浓度CPPU处理的果实中香气合成相关基因DXS3、GPPS、LIS表达量高峰推迟且峰值下调,浓度越高,推迟时间越久,峰值更低,基因表达受抑制,这与萜类物质相对含量降低相一致。[结论]25 mg·L^-1GA3+5 mg·L^-1CPPU处理能够改善‘阳光玫瑰’葡萄外观品质并保证内在品质。
DOI:10.1186/1471-2229-10-241URLPMID:3095323 [本文引用: 1]
pAbstract/p pBackground/p pThe sweet, floral flavor typical of Muscat varieties (Muscats), due to high levels of monoterpenoids (geraniol, linalool and nerol), is highly distinct and has been greatly appreciated both in table grapes and in wine since ancient times. Muscat flavor determination in grape (itVitis vinifera /itL.) has up to now been studied by evaluating monoterpenoid levels through QTL analysis. These studies have revealed co-localization of 1-deoxy-D-xylulose 5-phosphate synthase (itVvDXS/it) with the major QTL positioned on chromosome 5./p pResults/p pWe resequenced itVvDXS /itin an itad hoc /itassociation population of 148 grape varieties, which included muscat-flavored, aromatic and neutral accessions as well as muscat-like aromatic mutants and non-aromatic offsprings of Muscats. Gene nucleotide diversity and intragenic linkage disequilibrium (LD) were evaluated. Structured association analysis revealed three SNPs in moderate LD to be significantly associated with muscat-flavored varieties. We identified a putative causal SNP responsible for a predicted non-neutral substitution and we discuss its possible implications for flavor metabolism. Network analysis revealed a major star-shaped cluster of reconstructed haplotypes unique to muscat-flavored varieties. Moreover, muscat-like aromatic mutants displayed unique non-synonymous mutations near the mutated site of Muscat genotypes./p pConclusions/p pThis study is a crucial step forward in understanding the genetic regulation of muscat flavor in grapevine and it also sheds light on the domestication history of Muscats. itVvDXS /itappears to be a possible human-selected locus in grapevine domestication and post-domestication. The putative causal SNP identified in Muscat varieties as well as the unique mutations identifying the muscat-like aromatic mutants under study may be immediately applied in marker-assisted breeding programs aimed at enhancing fragrance and aroma complexity respectively in table grape and wine cultivars./p
DOI:10.1093/jxb/err231URLPMID:21868399 [本文引用: 1]
Grape berries of Muscat cultivars (Vitis viniferaL.) contain high levels of monoterpenols and exhibit a distinct aroma related to this composition of volatiles. A structural gene of the plastidial methyl-erythritol-phosphate (MEP) pathway, 1-deoxy-D-xylulose 5-phosphate synthase (VvDXS), was recently suggested as a candidate gene for this trait, having been co-localized with a major quantitative trait locus for linalool, nerol, and geraniol concentrations in berries. In addition, a structured association study discovered a putative causal single nucleotide polymorphism (SNP) responsible for the substitution of a lysine with an asparagine at position 284 of the VvDXS protein, and this SNP was significantly associated with Muscat-flavoured varieties. The significance of this nucleotide difference was investigated by comparing the monoterpene profiles with the expression ofVvDXSalleles throughout berry development in Moscato Bianco, a cultivar heterozygous for the SNP mutation. Although correlation was detected between theVvDXStranscript profile and the accumulation of free monoterpenol odorants, the modulation ofVvDXSexpression during berry development appears to be independent of nucleotide variation in the coding sequence. In order to assess how the non-synonymous mutation may enhance Muscat flavour, anin vitrocharacterization of enzyme isoforms was performed followed byin vivooverexpression of eachVvDXSallele in tobacco. The results showed that the amino acid non-neutral substitution influences the enzyme kinetics by increasing the catalytic efficiency and also dramatically affects monoterpene levels in transgenic lines. These findings confirm a functional effect of theVvDXSgene polymorphism and may pave the way for metabolic engineering of terpenoid contents in grapevine.
DOI:10.1007/s13258-017-0574-zURL [本文引用: 1]
Linalool, -terpineol, citronellol, nerol, geraniol, and geraniol acid are the main monoterpenes in the grapevine ( Vitis vinifera L.) with the concentrations of these substances differing in different varieties. This research performed association analyses using a core collection of 92 grape accessions to identify causal SNPs in the gene 1-deoxy- d -xylulose-5-phosphate synthase ( VvDXS ), a regulator of monoterpene metabolism. We evaluated the free linalool, -terpineol, citronellol, nerol, geraniol, and geraniol acid content of 92 grape varieties in two consecutive years (2014 and 2015). Twenty-two SNPs were found in the VvDXS coding region and were used in an association analysis. We found that the linalool, -terpineol, nerol, and geraniol acid levels were higher in varieties with a G/T genotype at P852 than in varieties with other genotypes at this site. Additionally, we found a novel polymorphism site P1678 (A/G) with a functional effect on terpenoid content that was related to the regulation of gene transcription. This study revealed the relationship between VvDXS and aroma, with the identified polymorphism site potentially assisting in the screening of aroma compounds at early stages in fruit development in grapevines.
DOI:10.1186/1471-2229-10-226URLPMID:3017849 [本文引用: 1]
pAbstract/p pBackground/p pTerpenoids are among the most important constituents of grape flavour and wine bouquet, and serve as useful metabolite markers in viticulture and enology. Based on the initial 8-fold sequencing of a nearly homozygous Pinot noir inbred line, 89 putative terpenoid synthase genes (itVvTPS/it) were predicted by itin silico /itanalysis of the grapevine (itVitis vinifera/it) genome assembly abbrgrpabbr bid=B11/abbr/abbrgrp. The finding of this very large itVvTPS /itfamily, combined with the importance of terpenoid metabolism for the organoleptic properties of grapevine berries and finished wines, prompted a detailed examination of this gene family at the genomic level as well as an investigation into itVvTPS /itbiochemical functions./p pResults/p pWe present findings from the analysis of the up-dated 12-fold sequencing and assembly of the grapevine genome that place the number of predicted itVvTPS /itgenes at 69 putatively functional itVvTPS/it, 20 partial itVvTPS/it, and 63 itVvTPS /itprobable pseudogenes. Gene discovery and annotation included information about gene architecture and chromosomal location. A dense cluster of 45 itVvTPS /itis localized on chromosome 18. Extensive FLcDNA cloning, gene synthesis, and protein expression enabled functional characterization of 39 itVvTPS/it; this is the largest number of functionally characterized itTPS /itfor any species reported to date. Of these enzymes, 23 have unique functions and/or phylogenetic locations within the plant itTPS /itgene family. Phylogenetic analyses of the itTPS /itgene family showed that while most itVvTPS /itform species-specific gene clusters, there are several examples of gene orthology with itTPS /itof other plant species, representing perhaps more ancient itVvTPS/it, which have maintained functions independent of speciation./p pConclusions/p pThe highly expanded itVvTPS /itgene family underpins the prominence of terpenoid metabolism in grapevine. We provide a detailed experimental functional annotation of 39 members of this important gene family in grapevine and comprehensive information about gene structure and phylogeny for the entire currently known itVvTPS /itgene family./p
DOI:10.1071/FP12326URL [本文引用: 1]
Grape (Vitis vinifera L.) flavour management in the vineyard requires knowledge of the derivation of individual flavour and aroma characteristics. Some of the most prevalent wine grape aroma constituents are terpenoids and this study represents a wide report about grape terpene synthase (TPS) gene transcript profiling in different tissues of two aromatic grapevine varieties, particularly flowers and developing berries, correlated with the accumulation patterns of free aroma compounds. All investigated genes belonging to the TPS-a and TPS-b subfamilies reached the highest expression in accordance with the peak of accumulation of the respective compounds. In the TPS-g subfamily, only one of the genes characterised for linalool synthases showed major transcript abundance in ripening berries, whereas the only geraniol synthase had a peak of expression in green berries and at the beginning of ripening, when geraniol concentration started to increase and overcome the linalool concentration. The genes identified in this study as being mainly responsible for linalool and geraniol synthesis during berry development, and the phenological phases in which they are mostly expressed, should be of interest to viticulturists and wine makers to improve decision making along the chain of production.
DOI:10.1016/j.phytochem.2014.06.007URLPMID:25014656 [本文引用: 1]
Plants produce a plethora of volatile organic compounds (VOCs) which are important in determining the quality and nutraceutical properties of horticultural food products, including the taste and aroma of wine. Given that some of the most prevalent grape aroma constituents are terpenoids, we investigated the possible variations in the relative expression of terpene synthase (TPS) genes that depend on the organ. We thus analysed mature leaves, young leaves, stems, young stems, roots, rachis, tendrils, peduncles, bud flowers, flowers and berries of cv Moscato bianco in terms of their VOC content and the expression of 23 TPS genes. In terms of the volatile characterization of the organs by SPME/GC–MS analysis, flower buds and open flowers appeared to be clearly distinct from all the other organs analysed in terms of their high VOC concentration. Qualitatively detected VOCs clearly separated all the vegetative organs from flowers and berries, then the roots and rachis from other vegetative organs and flowers from berries, which confirms the specialization in volatile production among different organs. Our real-time RT-PCR results revealed that the majority of TPS genes analysed exhibited detectable transcripts in all the organs investigated, while only some were found to be expressed specifically in one or just a few organs. In most cases, we found that the known products of the in vitro assay of VvTPS enzymes corresponded well to the terpenes found in the organs in which the encoding gene was expressed, as in the case of (E)-β-caryophyllene synthases, α-terpineol synthase and α-farnesene synthase. In addition, we found groups of homologous TPS genes, such as (E)-β-caryophyllene and β-ocimene synthases, expressed distinctively in the various tissues. This thus confirmed the subfunctionalization events and a specialization on the basis of the organs in which they are mostly expressed.
DOI:10.1016/j.phytochem.2004.03.018URLPMID:15184006 [本文引用: 1]
The flavour and aroma of certain Vitis vinifera grape varieties is dominated by volatile terpenes and small volatile aldehydes. Monoterpenes contribute to the final grape and wine aroma and flavour in form of free volatiles and as glycoside conjugates of monoterpene alcohols. Typical monoterpenol components of the cultivar Gewürztraminer and other aroma-rich grape varieties are linalool, geraniol, nerol, citronellol, and α-terpineol. In a functional genomics effort to identify genes for the formation of monoterpene alcohols in V. vinifera, a database of full-length cDNA sequences was screened in silico and yielded two clones for putative monoterpene synthases. The gene products were functionally characterized by expression in Escherichia coli, in vitro enzyme assay and gas chromatography-mass spectrometry (GC-MS) product identification as multi-product (61)-α-terpineol synthases.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}