 ,1,2, �ųɸ�3, �Ž�ΰ3
,1,2, �ųɸ�3, �Ž�ΰ3Cloning and Identification of Long-Chain Non-Coding RNA Linc24063 and Its Correlation with the Expression Level of miRNAs in Yak
WANG Hui1,2, CHAI ZhiXin1,2, ZHU JiangJiang1,2, ZHONG JinCheng,1,2, ZHANG ChengFu3, Xin JinWei3ͨѶ����:
���α༭: �ּ���
�ո�����:2018-11-5��������:2019-05-9�����������:2019-07-16
| ��������: |
Received:2018-11-5Accepted:2019-05-9Online:2019-07-16
����� About authors
����,E-mail: wanghui892321@sina.cn��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (984KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
����, ��־��, �콭��, �ӽ��, �ųɸ�, �Ž�ΰ. ��ţLinc24063�Ŀ�¡����������miRNAs ����ˮƽ������Է���[J]. �й�ũҵ��ѧ, 2019, 52(14): 2538-2547 doi:10.3864/j.issn.0578-1752.2019.14.012
WANG Hui, CHAI ZhiXin, ZHU JiangJiang, ZHONG JinCheng, ZHANG ChengFu, Xin JinWei.
0 ����
���о����塿�ڻ�����,lncRNA��ͨ��miRNA���ӵ������á�һ����,lncRNA����ΪmiRNA������,��miRNA�л������Խ��miRNA;��һ����,lncRNA����Ϊ��������ԴRNA��ceRNA��,�������ͬmiRNA���Ԫ����mRNA����miRNA,���Ʒ��롢���ڿɱ���л����mRNA����[1]����ţ��Ҫ�ֲ��ں���3 000 m���ϵ���ظ�ԭ����,Ϊ���������ṩ�⡢�̲�Ʒ����ţ�̾��иߵ��ס�����֬�ʵ��ص�,�о�lncRNA����ţ������֯�еľ��幦�ܼ����ػ���,�Ը��õ�������ţ�Ŵ���Դ������Ҫ���塣��ǰ���о���չ������ţ�Ȳ��鶯����,�������������Ŵ��ľ��Ӻ����ӵı����Ŵ����β�ͬ,���½ӽ�1%�ĵ����ʱ�������������ױ�������,�����ױ������ı����Ŵ�������Ϊ������ӡ��[2,3]������Ϊֹ,�����ࡢС��ţ������Ȳ��鶯���зֱ������350����236����45����18��ӡ�ǻ���http://igc.otago.ac.nz��,��Щӡ�ǻ���ͨ���Դص���ʽ���ڡ�Dlk1-Dio3ӡ����λ����Ⱦɫ��14q32��С��Ⱦɫ��12q��ţȾɫ�� 21q������Ⱦɫ��18q��,��������̥����[4]����֢�ķ���[5]��ϸ����ֳ�ֻ�[6]���������л����ͨ·[7]�з�����Ҫ���ö����ܹ�ע,��Ŀǰ�о��϶��ӡ������֮һ��Dlk1- Dio3ӡ�������3����Դ����ı��뵰����Dlk1��Dio3�� Rtl1/Peg11����ĸϵ����ij����DZ���RNA����lncRNA:Meg3��Meg8��Meg9����miRNA��snoRNA[8]���о�����,Dlk1-Dio3ӡ���������ּ�߶ȱ���,�㷺���뵽��������������������С�Dlk1-Dio3ӡ������ļ�����С���ϸ��������ˮƽ���[9];Dlk1������ض��ܸ�ϸ��[10]����ϸ������ֳ��ֻ�[7];Dlk1������������������ӷ�Сϸ���ΰ�ϸ������Ϯ����[11]��LncRNA�dz��ȴ���200 bp��һ��DZ���RNA,����̥����[12]��ϸ����ֳ�ֻ�[13]�����ӷ���[14]�ȶ������������з�����Ҫ�ĵ������á�Dlk1-Dio3ӡ�������ж���߱���Ҫ���ܵ�lncRNA,���֬��ʳ������С��Meg3������,Meg3ͨ������FoxO1�ı�����ǿ�����ȵ��صֿ�,����Met3����תС���ɸ�֬��ʳ�����ѪҺ���������������ϵ�[14]��������,�����ڲ�ͬ����Dlk1-Dio3ӡ����䷢�ֲ�����������lncRNA,������Meg8��Meg9��Meg9��Dio3֮��ֱ��ֲ�������2��lncRNA,��lncRNA B830012L 14Rik��AK044800,lncRNA B830012L 14Rik��С����ԡ����ࡢ���������֯�й㷺����[15],��lncRNA AK044800��Ҫ����̥ʱ�ڵ�ǰ����֯�б���[16];��ţDlk1-Dio3ӡ����lncRNA��������,���������Linc24061��Linc24063�� Linc24064����lncRNAs,�����������ࡢ���༰������֯�о��㷺����[17,18]�������о�����㡿ǰ���о�����,Dlk1-Dio3ӡ�����ڵ�lncRNA,�ܲ�������������ѧ���̵��ء�������ţ��,δ����ӡ������lncRNA���о��������������Ĺؼ����⡿���о�ּ����ţDlk1- Dio3ӡ����Meg8��Meg9�������ţLinc24063,ͨ��������Ϣѧ�����miRNA����Ԥ��,�����֯�����������������������֯�����miRNAs�������,�Ӷ�Ϊ��ţLinc24063ͨ��miRNA���ӹ��ܵĵ��ػ����о��ṩ�������ݡ�1 �����뷽��
1.1 �������
2018��4��,��������������������������,ѡȡ���ڵ�һ�����ڡ����ؽӽ���������12ͷĸ��ţ,����ɼ��ѳ������ࡢ���ࡢ���ࡢ���ٺʹ�����֯,DEPC������ϴ������ֽ��װ,Ѹ������Һ������,����ʵ���ұ��á�1.2 RNA��ȡ����ţLinc24063����
����֯RNA��ȡ����Trizol��Invitrogen���Լ�˵���顣RNA�����Ⱥ��������ϸ���á�����ţDlk1-Dio3ӡ����Meg8��Meg9������н�������������ţ������ȶ�,�����ţLinc24063��NCBI��¼��:KU956000.1����������ţ���������и߶ȱ���,���ݴ���������Primer premier 5�����ţLinc24063����5�� RACE��cDNAĩ�˿�����������3�� RACE�������ţ����֯RNA������Ϻ�,����SMARTer RACE cDNA Amplification Kit��Clontech, Palo Alto, CA, USA�����ղ���˵�������RACE����,��ȡLinc24063��5��˺�3��ˡ�5�� RACE����Ϊ:5��-TTAAACATTCCTAACATCTGCC TAC-3��,3�� RACE����Ϊ5��-GGGACCCTGAGGCCCAGTCCATTC-3�䡣
1.3 ��ţLinc24063���з���
����NCBI BLAST����ţLinc24063���������ּ�ı����Լ�Ⱦɫ���ϵĶ�λ���з���;������������Coding Potential Calculator��http://cpc2.cbi.pku.edu.cn/����Linc24063����֪lncRNA Meg9����¼��:NR_132275.2���ͱ������β-�ҵ��ף�CSN2,��¼��:MH378280.1���ı�����������Ԥ��;����miRanda��mireap����������Linc24063����õ�miRNAs;����DAVID����������https://david.ncifcrf.gov/������Linc24063����õ�miRNAs�İл������GO������KEGG�ź�ͨ·������1.4 ��ţLinc24063ԭ�˱���
����ţLinc24063ȫ���ǿ�¡��Pet-28a�����EcoR���Hind��ø��λ����,���Linc24063-28aԭ�˱�������,ת����BL21����̬ϸ��,��ú���������Linc24063-28a�������,��ȡ������EcoR���Hind��ø�м�����CSN2-28aԭ�˱�������Ϊ��ʵ���ұ��档Linc24063-28a��CSN2-28a�յ����ﲽ���������[19]��ȡ1 mL�յ���ľ�Һ����,��100 ��L��1��SDS-PAGE Loading Buffer��������,��ˮ�����10 min,��֤������ȫ�ѽ⡣10 000��g����5 min��,ȡ����10 ��L����SDS-PAGE�����������ÿ���˹����R-250ȾɫҺȾɫ,��ɫҺ��ɫ,��������
1.5 ��ţLinc24063��֯�������
����PrimeScript TM RT reagent Kit with gDNA Eraser��TaKaRa, �������Լ��н�����֯��RNA��ת¼ΪcDNA,Linc24063�������ļ�����SYBR Premix Ex TaqTM�Լ��У�Takara��˵����,�ڲλ���ѡ������嵰��S9��RPS9,��¼��:XM_005899362.2�����㷺���ﵰ�ף�UXT, ��¼��:XM_014483477.1��[20];����miRNA�������ļ������Stem-loop���� [21],�ڲλ���ѡ��5S rRNA������Bio-Rad CFX96������,�����������ǽ�����ţLinc24063��miRNAs��֯����������������ϵ��Ϊ:2��SYBR? Premix Ex TaqTM ��10 ��L,cDNAģ��1 ��L,�����������1 ��L,ddH2O 7 ��L��qPCR����Ϊ:95 �� 30 s;95 �� 5 s,60 �� 30 s,39��ѭ������������������1��Table 1
��1
��1ʵʱӫ�ⶨ��PCR����
Table 1
| �������� Gene name | �������У�5��-3�䣩 Primer sequence (5��-3��) | �����С Product length (bp) |
|---|---|---|
| Linc24063 | F:ACTGTGAACATAGATGGTGAGG | 141 |
| R:CACGGGGTCACAAAGAGTCA | ||
| RPS9 | F:GTGGTGAACATCCCGTCCTT | 130 |
| R:GCCACTGCACCTTGTAACAC | ||
| UXT | F:CGAGGCTTTCATCTCTGACG | 138 |
| R:TCCGAGTGATTAGCTTCCTGG | ||
| 18S rRNA | F:GTGGTGTTGAGGAAAGCAGACA | 79 |
| R:CGATCCCTGTCCTCACCTCATC | ||
| MiR-200a | RT:GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACaacatcgt | |
| F:CTGGtaacactgtctggta | 70 | |
| MiR-24 | RT:GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACctgttcct | |
| F:GGAtggctcagttcagc | 69 | |
| MiR-27a | RT:GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACcggaactt | |
| F:GGCacttcacagtggct | 69 | |
| MiR-141 | RT:GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACccatcttt | |
| F:GGCataacactgtctggt | 69 | |
| miRNA universal primer | GTGCAGGGTCCGAGGT |
�´��ڴ�|����CSV
1.6 ���ݷ���
������������������ʹ��2-����ct��,����Linc24063�Ķ���ѡ��UXT��RPS9�ļ���ƽ��ֵ���з���������SPSS 19.0Ƥ��ѷϵ����˫β����ӦLinc24063��miRNA������ԡ��������|R |��0.8Ϊ��ǿ���,0.6��|R |��0.8Ϊǿ���,0.4��|R |��0.6Ϊ�ж����,|R |��0.4Ϊ����ء������ʾΪƽ��ֵ+����*,P<0.05Ϊ��������,**,P<0.01Ϊ���켫������2 ���
2.1 �����DZ���RNA Linc24063��������
����SMARTer RACE cDNA Amplification Kit ������ţ�����DZ���RNA Linc24063,�������,5�� RACEƬ�δ�СΪ476 bp,3�� RACEƬ�δ�СΪ356 bp��ͼ1-A��,�����������Linc24063��С758 bp��ͼ1-B��,λ����ţ21��Ⱦɫ���Dlk1-Dio3ӡ����NCBI blast�������ּ䱣����,���������ţLinc24063��ţ������ɽ������������Էֱ�Ϊ98%��90%��90%,�����˺�С������������Խϵ͡�ͼ1
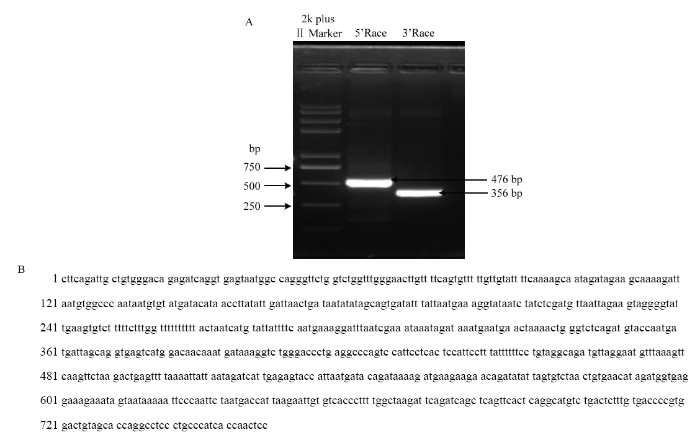 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1Linc24063 5�� Race��3�� Race��Ӿͼ��A�������У�B��
Fig. 1The 5�� Race and 3�� Race electrophoresis analysis (A) and the sequence (B) of Linc24063
2.2 �����DZ���RNA Linc24063�ļ���
������������Coding Potential Calculator�Գ����DZ���RNA Linc24063����֪lncRNA Meg9�ͱ������CSN2�ı�������Ԥ�����,�������:Linc24063�볤���DZ���RNA Meg9����,����DZ�ܾ��ϵ�,���������ڱ������CSN2,���ڷDZ���RNA����2����Table 2
��2
��2Linc24063��Meg9��CSN2�������DZ�ܷ���
Table 2
| ���� Gene | ����/�DZ��� Coding/Noncoding | ����DZ�� Coding probability | Fickett �� Fickett score | �ȵ�� Isoelectric point |
|---|---|---|---|---|
| Linc24063 | Noncoding | 0.01 | 0.35 | 9.52 |
| Meg9 | Noncoding | 0.17 | 0.36 | 7.90 |
| CSN2 | coding | 1.00 | 0.45 | 5.89 |
�´��ڴ�|����CSV
Ϊ��֤Linc24063�Ƿ�߱���������,���о���Linc24063����pET-28aԭ�˱�������,��ȡ��������EcoR���Hind��˫ø�м���,��ͼ2-A��֪,Linc24063-28a��CSN2-28a�ֱ��һ��758 bp��780 bp��С��Ƭ��,��һ��ͨ��������֤Ŀ��Ƭ�γɹ�����ԭ�˱������塣����ԭ�˱���ϵͳ�������Linc24063�ͱ������CSN2���з���,�������:Linc24063��30 kD����û�м����ױ���,�����䲻����Ч�ķ��뵰��,������ͬ����������,�������CSN2��30 kD�����е��ױ���,����CSN2�ܺܺõı��ﵰ�ף�ͼ2-B���������о��������,��ţLinc24063��һ�������ij����DZ���lncRNA��
ͼ2
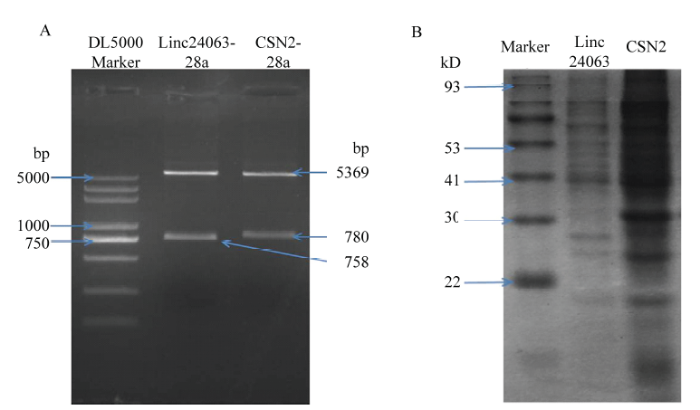 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2Linc24063��ԭ�˱���
��A��Linc24063��CSN2ԭ�˱�������ø�м���;��B��Linc24063��CSN2ԭ�˱���
Fig. 2The prokaryotic expression of Linc24063
(A) Identification of prokaryotic expression vector of Linc24063 and CSN2 by restriction endonuclease. (B) The prokaryotic expression of Linc24063 and CSN2
2.3 �����DZ���RNA Linc24063��֯������
����RT-qPCR����,�����о���Linc24063����ţ��ͬ��֯�ı����ס������ʾ,Linc24063�����ٱ��������,��������������֯��P<0.05��,��δ��Ժ�����,�ڸ�����ѳ��б������ϵͣ�ͼ3����ͼ3
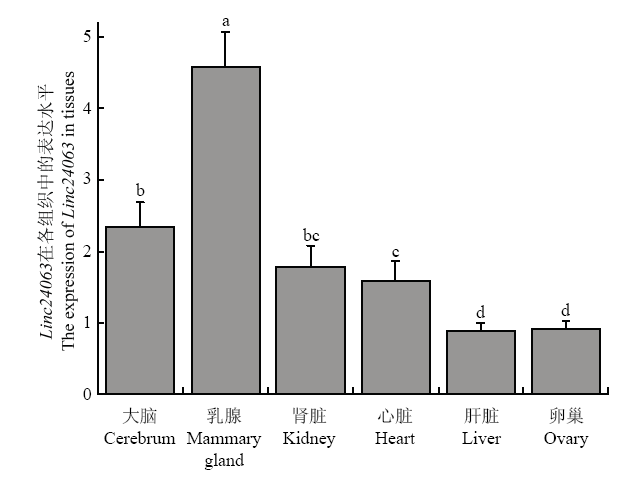 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3Linc24063�ڸ���֯�еı�����
Сд��ĸa,b��ʾ��������,P<0.05
Fig. 3The expression of Linc24063 in tissues
The lowercase a, b means significant difference, P<0.05
2.4 �����DZ���RNA Linc24063������Ϣѧ����
����miRbase 22���ݿ�����ͨţ����ţ�Ľ�Դ���֣����������е�miRNAs,����miRanda��mireap����������Linc24063��������õ�miRNA,������21�����ּ䱣��miRNAs����3����Table 3
��3
��3��Linc24063����õı���miRNAs
Table 3
| Name | ���������У�5��-3�䣩Mature sequence(5��-3��) |
|---|---|
| bta-miR-137 | UUAUUGCUUAAGAAUACGCGUAG |
| bta-miR-139 | UCUACAGUGCACGUGUCUCCAGU |
| bta-miR-141 | UAACACUGUCUGGUAAAGAUGG |
| bta-miR-154a | UAGGUUAUCCGUGUAGCCUUCG |
| bta-miR-200a | UAACACUGUCUGGUAACGAUGUU |
| bta-miR-221 | AGCUACAUUGUCUGCUGGGUUU |
| bta-miR-223 | UGUCAGUUUGUCAAAUACCCCA |
| bta-miR-24 | UGGCUCAGUUCAGCAGGAACAG |
| bta-miR-27a | AGGGCUUAGCUGCUUGUGAGCA |
| bta-miR-30b-5p | UGUAAACAUCCUACACUCAGCU |
| bta-miR-369-3p | AAUAAUACAUGGUUGAUCUUU |
| bta-miR-374a | UUAUAAUACAACCUGAUAAGUG |
| bta-miR-376b | AUCAUAGAGGAAAAUCCAUGUU |
| bta-miR-378b | ACUGGACUUGGAGUCAGAAGGC |
| bta-miR-378c | ACUGGACUUGGAGUCAGAAGU |
| bta-miR-455-3p | GCAGUCCAUGGGCAUAUACACU |
| bta-miR-487a | AAUCAUACAGGGACAUCCAGU |
| bta-miR-500 | UAAUCCUUGCUACCUGGGUGAGA |
| bta-miR-543 | AAACAUUCGCGGUGCACUUCUU |
| bta-miR-551a | GCGACCCAAUCUUGGUUUCCA |
| bta-miR-655 | AUAAUACAUGGUUAACCUCUCU |
�´��ڴ�|����CSV
��2.3�о�����,Linc24063�����ٵı��������,�����Ʋ��������ٷ�������ϳɹ����о�����Ҫ����,�ɴ˽��ǰ����������miRNAs�������о�[22,23],����13��miRNAs������������֯����Linc24063��������á�����13��miRNAs�İл������GO������KEGG�ź�ͨ·����,�������,��ЩmiRNAs����������RNA�ۺ�øII��ػ����ת¼������ѧ�����У���4����KEGG�ź�ͨ·��������,��ЩmiRNAs�������뵽TGF-beta�ź�ͨ·��PI3K-Akt�ź�ͨ·��ErbB�ź�ͨ·��MAPK�ź�ͨ·�ȣ���5����
Table 4
��4
��4��Linc24063����õ�miRNAs�л����ǰ10��GO������Ŀ
Table 4
| GO��Ŀ Go Term | ������Ŀ Gene count | �������� Fold enrichment | Pֵ P value |
|---|---|---|---|
| Nucleusϸ���� | 201 | 1.63 | 7.98E-14 |
| Cytoplasmϸ���� | 171 | 1.28 | 2.06E-04 |
| Regulation of transcription from RNA polymerase II promoter RNA�ۺ�øII������ת¼�ĵ��� | 116 | 1.98 | 4.76E-06 |
| Nucleoplasm ���� | 83 | 1.48 | 2.68E-04 |
| Metal ion binding�������ӽ�� | 71 | 1.42 | 0.00231 |
| Cytosol��Һ | 60 | 1.37 | 0.011998 |
| RNA polymerase II core promoter proximal region sequence-specific DNA binding DNA�����Խ��RNA�ۺ�øII���������ӽ������� | 52 | 2.92 | 3.51E-08 |
| Transcription, DNA-templated DNAΪģ���ת¼ | 42 | 1.57 | 0.004197 |
| Transcriptional activator activity, RNA polymerase II core promoter proximal region RNA�ۺ�øII���������ӽ���ת¼������� | 33 | 3.05 | 2.15E-06 |
| Sequence-specific binding���������Խ�� | |||
| Chromatin bindingȾɫ�ʽ�� | 30 | 2.22 | 8.71E-05 |
�´��ڴ�|����CSV
Table 5
��5
��5��Linc24063����õ�miRNAs�л�����ź�ͨ·
Table 5
| �ź�ͨ·��Ŀ Pathway term | ������ Input number | Pֵ P-value | У����Pֵ Corrected P-value |
|---|---|---|---|
| TGF-beta signaling pathway TGF-β�ź�ͨ· | 16 | 1.46E-06 | 8.22E-05 |
| PI3K-Akt signaling pathway PI3K-Akt�ź�ͨ· | 28 | 0.000267 | 0.0063 |
| Axon guidance ��ͻ���� | 18 | 0.000392 | 0.00865 |
| ErbB signaling pathway ErbB �ź�ͨ· | 10 | 0.002087 | 0.03393 |
| Chronic myeloid leukemia������ϸ����Ѫ�� | 9 | 0.002965 | 0.04313 |
| FoxO signaling pathway FoXO�ź�ͨ· | 13 | 0.003366 | 0.04738 |
| Pathways in cancer ��֢ͨ· | 27 | 0.003372 | 0.04783 |
| Insulin signaling pathway�ȵ����ź� | 13 | 0.003495 | 0.04786 |
| Maturity onset diabetes of the young���� | 5 | 0.003572 | 0.04789 |
| MAPK signaling pathway MAPK�ź�ͨ· | 19 | 0.003611 | 0.04952 |
�´��ڴ�|����CSV
2.5 �����DZ���RNA Linc24063��miRNAs��������Է���
Ϊ̽��Linc24063��miRNAs�Ĺ��������,���о�����RT-qPCR��������֯�м��Linc24063��miR-200a ��miR-24��miR-141��miR-27a�ı�����,��������Ƥ��ѷ����Է�����ͼ4��,�������:Linc24063��12ͷ��ţ������֯�еı�������miR-200a��R=-0.834,P=0.001����miR-141��R=-0.657,P=0.02���ı����������������,��miR-27a�ı�������������أ�R=0.647,P=0.023��,����miR-24����������ԣ�P>0.05���������о��������,Linc24063����ţ������֯�п���ͨ������miR-200a��miR-141��miR-27a����ı���,�Ӷ����뵽���ٷ�������ɷֺϳɼ���л�����������С�ͼ4
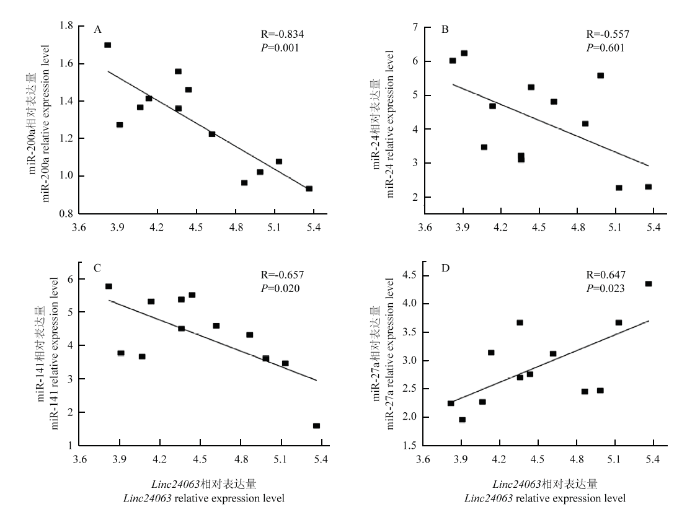 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ412ͷ��ţ������֯Linc24063��miRNAs����������Է���
Fig. 4The correlation between expression levels of Linc24063 and miRNAs in 12 mammary gland of yaks
3 ����
Ŀǰ,����lncRNA���о�����������ģʽ���P����,������ţ������Ҳ��չ�˴����о�,���ǹ�����ţlncRNA���о����١����о��״�ͨ��RACE������ȡ����������ţLinc24063,λ����ţ21��Ⱦɫ���Dlk1-Dio3ӡ����,�ڷ�ۻ����ţ�����о��нϸߵı����ԣ�>90%��,��ǰ�˶�Dlk1-Dio3ӡ������о�������ӡ���������ּ���н�ǿ�ı����ԵĽ��һ��[24]����֯���������������,Linc24063��������6����֯�о��б���,���������б��������,����Ǵ��Ժ�����,����Linc24063����Щ��֯�п��ܾ��߱�����ѧ���ܡ�������Ϣѧ�����������:��ɸѡ��21����Linc24063����õ����ּ䱣��miRNAs �����ǰ���о���miRNAs��������֯�еı������о�[22,23],����ɸѡ��13��miRNAs������������֯����Linc24063��������á�����13��miRNAs�İл������GO������KEGG�ź�ͨ·��������:��Linc24063����õ�miRNAs�İл�����TGF-beta�ź�ͨ·��PI3K-Akt�ź�ͨ·���ȵ����ź�ͨ·������������������ͻ������Axon guidance���ź�ͨ·�С�TGF-beta�ź�ͨ·[25]��PI3K-Akt�ź�ͨ·[26,27]���ȵ����ź�ͨ·�������ٷ�������֬�鵰�ϳ�������ص��ź�ͨ·,���Linc24063��������֯�б�������ߵ��ص�,�Ʋ�Linc24063���������ٷ�������֬�鵰�ϳɵ�����ѧ�����з�����Ҫ���á�
Ϊ̽��Linc24063���ܵ����û���,���о������ѡ��4�����������������õ�miRNAs,��������֯�ж������������������Է������������,Linc24063��miR-200a��miR-141��miR-27a��������ء�ǰ���о�����miR-200a��������Ƥϸ����ͨ������β-�ҵ���E-��ճ������ά����Ƥϸ���ļ���[28],ͬʱ����֬���鵰�ϳ��ź�ͨ·�з�����Ҫ����[29];miR-141�� miR-27a�ɷֱ�ͨ������STAT5���ס�PPAR�õ��ı���,���뵽�鵰�ס���֬�ĺϳ���[30]���ɴ˿����Ʋ�,Linc24063����ͨ������miR-200a��miR-141��miR-27a���뵽��ţ�鵰����֬������ϳ���,��������幦�ܺ����û��ƻ�������о���
4 ����
��ţDlk1-Dio3ӡ�������һ���ڷ�ۻ�����б��صij����DZ���RNA Linc24063,ȫ��758 bp�������ټ�������֯�о��нϸ߱���ˮƽ,������Ϣѧ��������Linc24063���ܲ��뵽PI3K-Akt���ȵ��ص�����֬�鵰��������ص��ź�ͨ·�С���������֯��Linc24063����֬�鵰�ϳ���ص�miRNAs,��miR-200a��miR-27a��miR-141��������������ԡ������о����ΪLinc24063�Ĺ����о��ṩ�˻������ݡ��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
DOI:10.1016/j.cell.2011.07.014URL [��������: 1]
[��������: 1]
DOI:10.1016/S0065-2660(02)46006-3URL [��������: 1]
DOI:10.1093/hmg/ddy235URL [��������: 1]
DOI:10.1080/17476348.2017.1355241URL [��������: 1]
DOI:10.1007/s00018-014-1698-9URL [��������: 1]
DOI:10.1016/j.cmet.2018.02.022URL [��������: 2]
DOI:10.1101/gr.172701URL [��������: 1]
DOI:10.1074/jbc.M110.131995URL [��������: 1]
DOI:10.1038/ng.1110URL [��������: 1]
DOI:10.3892/ol.2017.6019URL [��������: 1]
DOI:10.11843/j.issn.0366-6964.2017.01.001URLMagsci [��������: 1]

<p>��̥������һ�����ӵĹ��̣��䷢���ܵ��˺ܶ�ת¼�������ӵĵ��ء����Ÿ�ͨ����Ȳ������ķ�չ���о����ַDZ���RNA��ncRNA���Զ�����̥������XȾɫ��ʧ��Ա���ء��Բ�����������ʮ����Ҫ�ĵ������á����У�miRNA��Ҫͨ���������mRNA��3′UTR��ϲ��������̥��������ػ��� lncRNAͨ��ת¼���Ż�������Ⱦɫ����������ػ���Ӷ��鵼���鶯��XȾɫ��ʧ�������WȾɫ��ļ���������piRNAͨ����Ĭת������ά����ֳϸ��DNA�����ԣ���������ֳϸ���γɼ��Ҳ��Ա���أ�circRNA������ΪmiRNA�ĺ�����ض����Բ��ķ������������miRNA��lncRNA��piRNA��circRNA��ncRNA�ǶȲ���������̥���������е��о���չ��Ϊ��һ���о�ncRNA������ض�����̥�������̵����û����ṩ�ο���</p>
DOI:10.11843/j.issn.0366-6964.2017.01.001URLMagsci [��������: 1]
<p>��̥������һ�����ӵĹ��̣��䷢���ܵ��˺ܶ�ת¼�������ӵĵ��ء����Ÿ�ͨ����Ȳ������ķ�չ���о����ַDZ���RNA��ncRNA���Զ�����̥������XȾɫ��ʧ��Ա���ء��Բ�����������ʮ����Ҫ�ĵ������á����У�miRNA��Ҫͨ���������mRNA��3′UTR��ϲ��������̥��������ػ��� lncRNAͨ��ת¼���Ż�������Ⱦɫ����������ػ���Ӷ��鵼���鶯��XȾɫ��ʧ�������WȾɫ��ļ���������piRNAͨ����Ĭת������ά����ֳϸ��DNA�����ԣ���������ֳϸ���γɼ��Ҳ��Ա���أ�circRNA������ΪmiRNA�ĺ�����ض����Բ��ķ������������miRNA��lncRNA��piRNA��circRNA��ncRNA�ǶȲ���������̥���������е��о���չ��Ϊ��һ���о�ncRNA������ض�����̥�������̵����û����ṩ�ο���</p>
URL [��������: 1]
URL [��������: 1]
URL [��������: 2]
URL [��������: 2]
DOI:10.1016/j.bbrc.2015.11.048URL [��������: 1]
DOI:10.1007/s10735-011-9337-3URL [��������: 1]
DOI:10.1007/s10059-013-2275-zURL [��������: 1]
DOI:10.11843/j.issn.0366-6964.2016.09.013URLMagsci [��������: 1]
<p>ּ�ڼ���ţ<em>Dlk</em>1-<em>Dio</em>3ӡ�������ڵ���ӡ��lncRNAs�����о�ѡȡλ��<em>Meg</em>8��<em>Meg</em>9������2��������б�ǩ��EST����ͨ��RT-PCR��¡���в����з�����������������lncRNA������������Ϊ<em>Linc</em>24062��<em>Linc</em>24063������<em>Linc</em>24062��<em>Linc</em>24063��ţ8����֯���ġ��Ρ�Ƣ���Ρ�������������Ƥ��֬���ʹ��ԣ��еı������2��lncRNAs�ڱ�������֯�о�������û��ڵ��������̬�ԣ�SNP��λ���ֱ�Ӳ���ͨ���Ƚ��Ӻ���ţ��SNPλ�㴦������PCR����������RT-PCR��������IJ����ͼ����<em>Linc</em>24062��<em>Linc</em>24063��ӡ��״̬������<em>Linc</em>24062��<em>Linc</em>24063��ţ�������֯�о�Ϊ����λ������˵��<em>Linc</em>24062��<em>Linc</em>24063��ţ����ӡ�ǵġ�</p>
DOI:10.11843/j.issn.0366-6964.2016.09.013URLMagsci [��������: 1]
<p>ּ�ڼ���ţ<em>Dlk</em>1-<em>Dio</em>3ӡ�������ڵ���ӡ��lncRNAs�����о�ѡȡλ��<em>Meg</em>8��<em>Meg</em>9������2��������б�ǩ��EST����ͨ��RT-PCR��¡���в����з�����������������lncRNA������������Ϊ<em>Linc</em>24062��<em>Linc</em>24063������<em>Linc</em>24062��<em>Linc</em>24063��ţ8����֯���ġ��Ρ�Ƣ���Ρ�������������Ƥ��֬���ʹ��ԣ��еı������2��lncRNAs�ڱ�������֯�о�������û��ڵ��������̬�ԣ�SNP��λ���ֱ�Ӳ���ͨ���Ƚ��Ӻ���ţ��SNPλ�㴦������PCR����������RT-PCR��������IJ����ͼ����<em>Linc</em>24062��<em>Linc</em>24063��ӡ��״̬������<em>Linc</em>24062��<em>Linc</em>24063��ţ�������֯�о�Ϊ����λ������˵��<em>Linc</em>24062��<em>Linc</em>24063��ţ����ӡ�ǵġ�</p>
DOI:10.1007/s10709-016-9939-5URL [��������: 1]
DOI:10.1016/j.bbagrm.2016.05.003URL [��������: 1]
DOI:10.1002/jcp.v232.5URL [��������: 1]
DOI:10.1093/nar/gni178URL [��������: 2]
DOI:10.1186/1471-2164-13-731URL [��������: 2]
DOI:10.3168/jds.2016-11461URL [��������: 1]
DOI:10.1016/j.tig.2008.03.011URL [��������: 1]
DOI:10.1023/A:1009568532177URL [��������: 1]
DOI:10.1016/j.mce.2017.04.025URL [��������: 1]
DOI:10.1021/acs.jafc.7b05305URL [��������: 1]
DOI:10.1371/journal.pone.0065127URL [��������: 1]
URLMagsci [��������: 1]
<P><FONT face=Verdana>ּ�ڹ���pAd��pri��miR��200a�����ٲ�����ʹ������ɽ��������Ƥϸ�����ȶ����� miR��200a���о� miR��200a ����֬�ϳ���ػ��� mRNA �����Ӱ�졣����ũ������ DNA Ϊģ������ pri��miR��200a������ Ad��Easy ϵͳ���������ٲ������� pAd��pri��miR��200a,���� HEK 293 ϸ�����н��в����İ�װ��������TCI50 ���ⶨ�����ζȡ�qRT��PCR ��� miR��200a �� 16 ����֬�ϳ���ػ��� mRNA ����ˮƽ�����з��������ʾ�� pri��miR��200a ���� pre��miR��200a 86 bp �Ͳ������й� 297 bp��ø�н��֤ʵ pAd��pri��miR��200a �����ɹ��� Ad��pri��miR��200a �ζȴ� 8��109 PFU��mL-1��ʵʱ�������������Ad��pri��miR��200a ��MOI Ϊ 200����Ⱦ��ɽ��������Ƥϸ�� 72 h��miR��200a �ı������϶��ո߳� 2.4 ����ͬʱ miR��200a �Ĺ��������� 10 ������ mRNA �������µ���6 ������ mRNA �������ϵ�������֬�����ͷ�ϳ���ػ��� FASN��֬��������ػ��� TIP47 ��֬����������ػ��� FABP4 �����ϴֱ��½��� 0.47��0.89�� 0.65 ������ TAG ������ػ��� DGAT1 ��֬����ػ��� HSL �϶��շֱ������� 0.52�� 1.49 �������о���õ������ٲ��� Ad��pri��miR��200a ����ɽ��������Ƥϸ�����ȶ����� miR��200a��ͬʱ miR��200a ������Ӱ����֬�ϳ���ػ��� mRNA ����ˮƽ��</FONT></P><P><FONT face=Verdana></FONT> </P>
URLMagsci [��������: 1]
<P><FONT face=Verdana>ּ�ڹ���pAd��pri��miR��200a�����ٲ�����ʹ������ɽ��������Ƥϸ�����ȶ����� miR��200a���о� miR��200a ����֬�ϳ���ػ��� mRNA �����Ӱ�졣����ũ������ DNA Ϊģ������ pri��miR��200a������ Ad��Easy ϵͳ���������ٲ������� pAd��pri��miR��200a,���� HEK 293 ϸ�����н��в����İ�װ��������TCI50 ���ⶨ�����ζȡ�qRT��PCR ��� miR��200a �� 16 ����֬�ϳ���ػ��� mRNA ����ˮƽ�����з��������ʾ�� pri��miR��200a ���� pre��miR��200a 86 bp �Ͳ������й� 297 bp��ø�н��֤ʵ pAd��pri��miR��200a �����ɹ��� Ad��pri��miR��200a �ζȴ� 8��109 PFU��mL-1��ʵʱ�������������Ad��pri��miR��200a ��MOI Ϊ 200����Ⱦ��ɽ��������Ƥϸ�� 72 h��miR��200a �ı������϶��ո߳� 2.4 ����ͬʱ miR��200a �Ĺ��������� 10 ������ mRNA �������µ���6 ������ mRNA �������ϵ�������֬�����ͷ�ϳ���ػ��� FASN��֬��������ػ��� TIP47 ��֬����������ػ��� FABP4 �����ϴֱ��½��� 0.47��0.89�� 0.65 ������ TAG ������ػ��� DGAT1 ��֬����ػ��� HSL �϶��շֱ������� 0.52�� 1.49 �������о���õ������ٲ��� Ad��pri��miR��200a ����ɽ��������Ƥϸ�����ȶ����� miR��200a��ͬʱ miR��200a ������Ӱ����֬�ϳ���ػ��� mRNA ����ˮƽ��</FONT></P><P><FONT face=Verdana></FONT> </P>
DOI:10.1016/j.gene.2013.03.050URL

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}