 ,ɽ��ũҵ��ѧ����Ƽ�ѧԺ,ɽ��̩�� 271018
,ɽ��ũҵ��ѧ����Ƽ�ѧԺ,ɽ��̩�� 271018Effect of Dietary ��-Linolenic Acid Levels on Physiological Function of Apis mellifera ligustica Worker Bee Larvae
YU Jing, ZHANG WeiXing, MA LanTing, XU BaoHua,College of Animal Science and Technology, Shandong Agricultural University, Taian 271018, ShandongͨѶ����:
���α༭: ��÷
�ո�����:2019-03-7��������:2019-03-27�����������:2019-07-01
| ��������: |
Received:2019-03-7Accepted:2019-03-27Online:2019-07-01
����� About authors
�ھ�,E-mail��1391031996@qq.com

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (471KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
�ھ�, ������, ������, �㱣��. ������-������ˮƽ��������۷乤���׳��������ܵ�Ӱ��[J]. �й�ũҵ��ѧ, 2019, 52(13): 2368-2378 doi:10.3864/j.issn.0578-1752.2019.13.015
YU Jing, ZHANG WeiXing, MA LanTing, XU BaoHua.
0 ����
���о����塿֬������ϸ��Ĥ����Ҫ�ɷ�,��ϸ��Ĥ�Ĺ���������Ҫ�����á�֬����Ҳ���۷䷱ֳ�ͷ����������,���۷䶬��������֬���巢������Դ[1]����-�������Ƕ�������һ�ֱ���IJ�����֬����,��������n-3�����֬�����ǰ������,�����ڿ�ת��Ϊ������n-3�����֬����DHA��EPA,�Ӷ�������DHA��EPA���Ƶ���������,�Ի�������������������л����Ҫ����[2]����-������������֬����ϳ�ø��COA-�Ȼ�ø��������������ת��ø��3-�ǻ�-3-���������øA��ԭø��3-hydroxy-3-methyl glutarylcoenzyme A reductase,HMGR���Ȼ���,���ɼ�ǿ�������е���-����,���ٸ��������͵��̴��ĺϳ�,�Ӷ���Ѫ֬���á���-������ͨ�����Ʀ�-6ϵPUFA�Ĵ�л,����ǰ������PGE2��ǰ���ٻ���PGI2��Ѫ˨��TXA2�ĺϳ�,�Ӷ��������ס�����������Ѫ˨�����á��۷��׳����Ҫ֬������Դ�Ƿ�����,����������-�����Ậ������,����-�����������ڲ��ܺϳ�,�����ʳ���л��,���̽���׳���������-��������������ˮƽʮ�ֱ�Ҫ����ǰ���о���չ�������о�����,�벸�鶯������,��-������Ȧ�-3֬������۷����֪��������������Ҫ������[1],Ҳ���о���Ϊ�۷䳦���и�Ũ�ȵ����������������еֿ������ϸ����Ⱦ������[3]��ARIEN��[4]�о�����-������Ȧ�-3֬����ȱ�����۷��Ӫ��Ӱ��,������ι����-�������������۷�����ʹ�������ѧϰ���������½�;��������[5]�о�����,������-������Ȳ�����֬������˹����ϸ��ʺ�ώ����Arma chinensis������������������,��߳ɳ����ʺ���Ⱥ������;Ҧ˼���[6]�Բ�ͬ��������-�������С��������θ30��35 d,����С��Ŀ�������ϸ������Ѫ����Ѫ��ˮƽ�������,˵����-�����������ǿС��������������;�챣�ҵ�[7]�о�����,��-��������п��������ӳ�����������,������-�������뿹������������ǿ����������;������[8]�о�����,���û�������-������ˮƽ�ܹ�Ӱ���۷�Ѫ�ܰ���֬�ʺ���������֬��ø���Լ�������ػ������ˮƽ��,���ڴ��û��������ӹ�����͵���-�����ᶼ�ή��������۷䣨Apis mellifera ligustica���������������о�����㡿Ŀǰ,������-�������������۷�Ӫ�����õ��о�����,�����׳����-�������Ӫ����Ҫ��δ���������������Ĺؼ����⡿���˹�������������۷乤���׳�Ϊ�о�����,ͨ��̽���׳������в�ͬ��-������ˮƽ�����������������������Ժ�����ָ���Ӱ��,��ȷ�����׳����-�������������Ҫ��,Ϊ�ƶ��۷��������ṩ���ݡ�1 �����뷽��
������2018��2019����ɽ��ũҵ��ѧ����Ƽ�ѧԺ��ɡ�1.1 ����
������۷乤���׳�ȡ��ɽ��ũҵ��ѧ�����Ⱥ����������������VANDENBERG��[9]���䷽����,�ڻ��������зֱ�����0.02%��0.04%��0.06%��0.08%����-������,�Ƴ�4����������������1����Table 1
��1
��1�����������
Table 1
| ԭ�� Ingredient | ��-����������ˮƽ Level of ��-linolenic acid (%) | ||||
|---|---|---|---|---|---|
| 0 | 0.02 | 0.04 | 0.06 | 0.08 | |
| ������ Royal jelly ������ Glucose ���� Fructose ��ĸ��ȡ�� Yeast extract ��ˮ Sterile water ��-��������-linolenic acid �ϼ�Total | 50.00 6.00 6.00 1.00 37.00 0 100.00 | 50.00 6.00 6.00 1.00 37.00 0.02 100.00 | 50.00 6.00 6.00 1.00 37.00 0.04 100.00 | 50.00 6.00 6.00 1.00 37.00 0.06 100.00 | 50.00 6.00 6.00 1.00 37.00 0.08 100.00 |
�´��ڴ�|����CSV
��Ҫ�Լ���Total RNA Kit II�Լ��й���OMEGA��˾,ӫ�ⶨ���Լ��й���TransGen��˾,Transcript First Strand cDNA Synthesis Kit�Լ��й���Roche��˾,����ø�������������飨ELISA���Լ��й��Կ�ŵ������Ƽ�����˾,��-�����ᣨŨ��80%���ϣ����Խ����쿭��������˾��
1.2 ���鷽��
�����÷����������������̶���һ�ų�Ƣ�ϲ���,12 h�ڲ����㹻�����ʱ,��Dz�����,�ͷŷ���,����3 d,�����ѷ���ΪС�׳�,��1�����׳档����ѡȡ1 200ֻ1����������۷乤���׳�,600ֻ�����׳滯Ӽ�ʺ�����ָ��ⶨ,����600ֻ������������ָ��ⶨ�������׳���������������Ʒ�Ϊ5��,ÿ��5���ظ�,ÿ���ظ�48ֻ����1�����׳����Ƴ��������¶����˵ļ���200 ��L������24��ϸ����������,���������ں����������У��¶�33��,���ʪ��55%��,�����ڼ�ÿ�������������������6��ĩ���7���,�׳濪ʼ��ֱ�����ű�����ʱ,���׳�ת������ǰ�̺����ֽ��24��ϸ��������������Ӽ,������������������ӱ[10]��1.3 �ⶨָ���뷽��
1.3.1 ��Ӽ�ʺ����ʲⶨ ��������1�쿪ʼ,ÿ���鲢��¼�׳����������,�����������弰ʱ�Ƴ�,ֱ��δ�������׳�ȫ����Ӽ�����·�,��¼�ɹ���Ӽ�������������·��������,ͳ���׳滯Ӽ�ʺ����ʡ�1.3.2 ��֬�������ⶨ �׳��֬�������ⶨ��������÷��[11]�ȷ�-�״����ᷨ���ֱ�ȡ5��7���乤���׳���Ʒ10ֻ,������ǰ��ɳ��صij���ƿ��,����65����¸�������12 h,�������Ʒ����10 mL���Ĺ���,����2 mL�ȷ�-�״���2�s1�����Һ,��ȫ�Զ���Ʒ������ĥ���Ƚ�3 min,����24 h�������Ĺ��м���2 mL�ȷ�-�״���2�s1�����Һ,3 000 r/min����10 min,������������һ���Ĺ���,������м���2 mL�ȷ�-�״���2�s1�����Һ,ͬ����������,������ת����һ���Ĺ���;���������Ĺ��м���1.2 mL 1.6% CaCl2��Һ,ҡ��,����1 h,ͬ����������,��ȥ�ϲ�Һ;��1 mL 2% CaCl2-�ȷ�-�״���3�s8�s4�����Һ�ϲ�Һ�رڻ����������Ĺ���,ͬ����������,��ȥ�ϲ�Һ;�²�Һת���Ѻ�ɳ��صij���ƿ��,70����º�ɳ���,����ƿ���β�ֵ��Ϊ��֬�ء�
1.3.3 �׳�Ѫ�ܰ�֬�ʺ����ⶨ �������Ϊ40 ��L��ëϸѪ����ȡ7���乤���׳�Ѫ�ܰ�,��������Ѫ�ܰʹ����Ѿ������������������1.5 mL���Ĺ���,������������Ĺܷ��ڱ�����,�Է�Ѫ�ܰͱ�����,��Ʒ����-80�汣�档�ⶨʱ,4��������13 000 r/min����5 min,ȡ100 ��L���屸��,��������7020��ȫ�Զ����������Dzⶨ�׳�Ѫ�ܰ��и������������̴������ܶ�֬���͵��ܶ�֬��������
1.3.4 �׳濹����ָ���֬�ʴ�л���������ø���Բⶨ ÿ���ظ����ѡȡ1ֻ6���乤���׳�,��1�s9��������ȼ���һ����������ˮ��2 mL���Ĺ��н�����֯�Ƚ�,�����Ƴ�10%�Ƚ�Һ,��4��������,13 000 r/min����10 min,ȡ100 ��L���屸�á���������ø�������������飨ELISA���ⶨ����ȩ��MDA���������ܳ��������绯ø��T-SOD����������øA�Ȼ�ø��ACC����֬����ϳ�ø��FAS����������ø��PO�����ܾ�ø��lysozyme�����ԡ�
1.3.5 ������ػ���������IJⶨ ʹ��Total RNA Kit II�Լ�����ȡ5��7���乤���׳���RNA,���ó����ֹ��ȼƼ����Ũ�Ⱥʹ��ȡ�����Transcript First Strand cDNA Synthesis Kit�Լ���,����RNA��ת¼ΪcDNA,-20�汣�汸�á��ڲλ���ѡ������ȶ�����-actin,ί���������﹤�̣��Ϻ����ɷ�����˾��ɶ�������������ϳ�,������Ϣ����2������ʵʱӫ�ⶨ���Լ��в���ָ��,������ABI7500ϵͳ���Ŀ�Ļ������Ա�����,��Ӧ��ϵΪ20 ��L����Ӧ������94�� 30 s;94�� 5 s,60�� 34 s,40��ѭ����
Table 2
��2
��2Ŀ�Ļ�����������
Table 2
| Ŀ�Ļ��� Target gene | �������� Primer sequence (5��-3��) | ���ﳤ�� Product length (bp) | GenBank��¼�� GenBank accession number |
|---|---|---|---|
| ������ø���� PO | F: ATGAGGACTTCCTGTCTGTTGCAG R: CTTCGGTAAGCGGTGGTACTGG | 132 | XM_026444741.1 |
| �ܾ�ø���� lysozyme | F: TCGCACTACTCGCCACGATA R: TTTGATGCAGAGCGAGAGCG | 114 | NM_001011614.1 |
�´��ڴ�|����CSV
1.4 ���ݷ���
���ݲ���SAS 9.2�������е����ط��������one-way ANOVA����Turkey������бȽϷ���,P<0.05Ϊ��������,P<0.01Ϊ���켫����,�������ݾ��ԡ�mean��SD������ʽ��ʾ��2 ���
2.1 ������-����������ˮƽ��������۷乤���׳滯Ӽ�ʺ����ʵ�Ӱ��
��������-�����������ˮƽΪ0.02%��0.04%ʱ,��Ӽ�ʺ�����������������������;��-�����������ˮƽΪ0.06%��0.08%ʱ,��Ӽ�ʺ����ʾ��������ڶ����飨P<0.05������3����Table 3
��3
��3��������-����������ˮƽ�Թ����׳滯Ӽ�ʺ����ʵ�Ӱ��
Table 3
| ��Ŀ Item | ��-����������ˮƽLevel of ��-linolenic acid (%) | Pֵ P-value | ||||
|---|---|---|---|---|---|---|
| 0 | 0.02 | 0.04 | 0.06 | 0.08 | ||
| ��Ӽ�� Pupation rate (%) | 82.86��0.80a | 86.31��1.07a | 84.65��0.26a | 73.70��0.71b | 51.14��0.56c | <0.0001 |
| ���� Emergence rate (%) | 60.00��0.39a | 62.45��0.86a | 52.50��0.35b | 23.33��1.33c | 16.67��0.66d | <0.0001 |
�´��ڴ�|����CSV
2.2 ��������-����������ˮƽ��7����������۷乤���׳�Ѫ�ܰ�����ָ���Ӱ��
��������-����������ˮƽΪ0.04%ʱ,Ѫ�ܰ����ܵ��ף�total protein,TP�������������ڶ�����,��������������-������������������׳�Ѫ�ܰ��и���������triglyceride,TG�����ܵ��̴���total cholesterol,TC���ĺ�����P<0.05��������������,��������-����������ˮƽΪ0.02%��0.04%��0.06%ʱ,���ܶ�֬���ף�high density lipoprotein,HDL���ĺ�����������,�����ܶ�֬���ף�low density lipoprotein,LDL���ĺ����������ͣ�P<0.05������4����Table 4
��4
��4��������-����������ˮƽ�Թ����׳�Ѫ�ܰ�����ָ���Ӱ��
Table 4
| ��Ŀ Item | ��-����������ˮƽLevel of ��-linolenic acid (%) | Pֵ P-value | ||||
|---|---|---|---|---|---|---|
| 0 | 0.02 | 0.04 | 0.06 | 0.08 | ||
| �ܵ���TP (g��L-1) | 40.80��0.37ab | 39.38��1.31b | 36.08��1.27c | 42.42��0.74a | 40.70��1.36ab | 0.0013 |
| ��������TG (mmol��L-1) | 3.36��0.39a | 1.59��0.04b | 1.58��0.01b | 2.01��0.21b | 1.56��0.13b | 0.0002 |
| �ܵ��̴�TC (mmol��L-1) | 0.94��0.01a | 0.59��0.03c | 0.63��0.02c | 0.77��0.01b | 0.78��0.03b | <0.0001 |
| ���ܶ�֬����HDL (mmol��L-1) | 0.38��0.01c | 0.60��0.01b | 0.79��0.03a | 0.66��0.07b | 0.40��0.03c | <0.0001 |
| ���ܶ�֬����LDL (mmol��L-1) | 0.17��0.01a | 0.13��0.01b | 0.09��0.01c | 0.14��0.01b | 0.18��0.00a | <0.0001 |
�´��ڴ�|����CSV
2.3 ��������-����������ˮƽ��5��7����������۷乤���׳��֬������Ӱ��
��-����������ˮƽΪ0.02%��0.06%��0.08%ʱ,5���乤���׳�����֬�����������ڶ�����;��-����������ˮƽΪ0.06%��0.08%ʱ,7���乤���׳�����֬�����������ڶ����飨P<0.05������5����Table 5
��5
��5��������-����������ˮƽ�Թ����׳��֬������Ӱ��(�ɻ�,%)
Table 5
| ��Ŀ Item | ��-����������ˮƽ Level of ��-linolenic acid (%) | Pֵ P-value | ||||
|---|---|---|---|---|---|---|
| 0 | 0.02 | 0.04 | 0.06 | 0.08 | ||
| 5�����׳� 5-day-old larva | 7.89��0.54c | 13.80��0.47b | 9.50��0.40c | 16.30��0.23a | 13.62��1.47b | 0.0002 |
| 7�����׳� 7-day-old larva | 12.05��0.82c | 13.49��0.11abc | 12.35��0.07bc | 14.61��0.87a | 15.76��1.01a | 0.0190 |
�´��ڴ�|����CSV
2.4 ��������-����������ˮƽ��6����������۷乤���׳濹����ָ���Ӱ��
����������,����������-����������ˮƽΪ0.02%��0.04%ʱ,6���乤���׳�ı���ȩ��MDA�������������ͣ�P<0.05����ͼ1-A��;��������-����������ˮƽΪ0.04%ʱ,6���乤���׳泬�������绯ø��SOD�������������ڶ����飨P<0.05����ͼ1-B����ͼ1
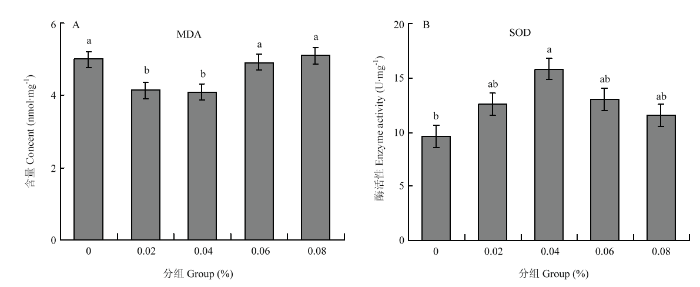 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1��������-����������ˮƽ��6���乤���׳濹����ָ���Ӱ��
���ϲ�ͬСд��ĸ��ʾ����������P<0.05������ͬ
Fig. 1Effect of ��-linolenic acid levels on antioxidant indexes of 6-day-old worker bee larvae
Different lowercases on the bars indicate significant difference (P<0.05). The same as below
2.5 ��������-����������ˮƽ��������۷乤���׳�֬�ʴ�л���ø���Ե�Ӱ��
��������-����������ˮƽΪ0.02%��0.04%ʱ,������۷乤���׳�����������øA�Ȼ�ø��ACC�������������ڶ����飨P<0.05����ͼ2-A������������-����������ˮƽΪ0.02%ʱ,�����׳�����֬����ϳ�ø��FAS����������������������죨P>0.05��,������ˮƽΪ0.04%��0.06%��0.08%ʱ,FAS�����������ڶ����飨P<0.05����ͼ2-B����ͼ2
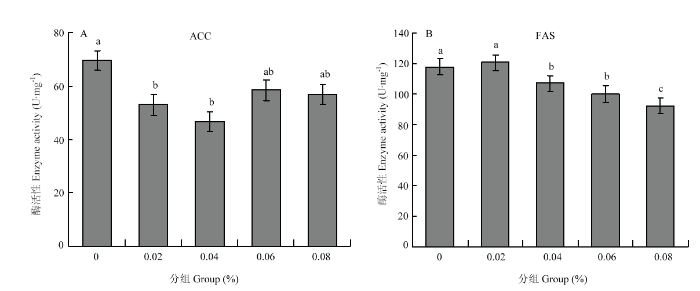 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2��������-����������ˮƽ��6���乤���׳�֬�ʴ�л���ø���Ե�Ӱ��
Fig. 2Effect of ��-linolenic acid levels on lipid metabolization-related enzyme activity of 6-day-old worker bee larvae
2.6 ��������-����������ˮƽ��������۷乤���׳�����������Ӱ��
��������-����������ˮƽΪ0.02%��0.04%��0.06%ʱ,6���乤���׳���ܾ�ø��lysozyme���ͷ�����ø��PO�������������ڶ����飨P<0.05����ͼ3����ͼ3
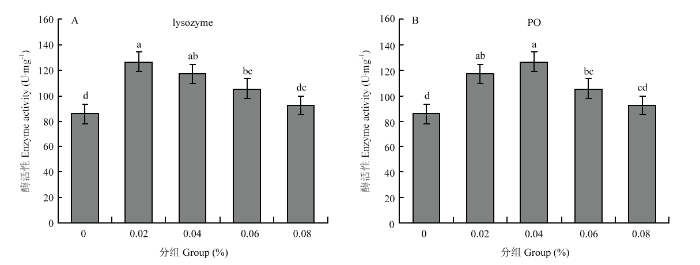 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ36���乤���׳��ܾ�ø�ͷ�����ø����
Fig. 3Lysozyme and PO activities of 6-day-old worker bee larvae
��������-����������ˮƽΪ0.02%��0.04%ʱ,5���乤���׳�lysozyme��PO��Ա������������ڶ����飨P<0.05��,������������-����������ˮƽΪ0.06%��0.08%ʱ,lysozyme��Ա����������Խ��ͣ�ͼ4-A��������������,����������-����������ˮƽΪ0.04%��0.06%ʱ,7���乤���׳�lysozyme��PO��Ա�������������,������������-����������ˮƽΪ0.08%ʱ,lysozyme��PO��Ա�����ȴ�������ͣ�P<0.05����ͼ4-B����
ͼ4
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4������ø���ܾ�ø������Ա�����
Fig. 4Relative expression of PO and lysozyme
3 ����
3.1 ��������-����������ˮƽ��������۷乤���׳滯Ӽ�ʺ����ʵ�Ӱ��
��-��������Ϊn-3ϵ�в�����֬����,�����������۷��������ܵ��о�����,���������ϵ��о��϶ࡣ��褵�[12]�о�����,�����������������ˮƽ������,�����������ܳ��ֳ������߶��͵����ơ�Ҳ�д����о�����,����һ�����������Ͷ��⼦�͵�������������������Ӱ��[13,14,15]���ź��ε�[16]�о�����,��ˮƽ����-��������������������������Ӱ�첻����,����ˮƽ����-�����������������������õ��������ܡ����������,��ˮƽ����-��������������������۷乤���׳�Ļ�Ӽ�ʺ�����������Ӱ��,������������-����������ˮƽ�ﵽ0.06%ʱ�����������׳�Ļ�Ӽ�ʺ����ʡ���������[5]�������ֱַ���������ͺ��������������ιώ��,����ȡʳ�������������ϵ�ώ�������׳����������������,�ɳ����ʺʹ��۳ɳ������վ�����������������ȡʳ��������ϵ�ώ��,���������������ϸ��ʺϵ������档�ɴ˿ɼ�,��-����������������۷��������ܵ�Ӱ���Դ�������,���������-��������������Ͷ������ּ����������������йء�3.2 ��������-����������ˮƽ��������۷乤���׳�֬�ʴ�л��Ӱ��
3.2.1 ��7���乤���׳�Ѫ�ܰ�����ָ���Ӱ�� ����Ѫ�ܰ���Ѫϸ��������Һ��Ѫ���Ļ����,Ϊ�����³´�л�����ʽ����봢��ij���,�����ߡ����ˡ����ϡ������ȷ���������Ҫ����[17]���ܵ��̴����������������ܶ�֬���ס����ܶ�֬���������ٴ���������ӳѪ֬��л���ܵij���ָ��[18]����-��������Խ����൨�̴��ϳɹ����еĹؼ�ø3-�ǻ�-3-���������øA��ԭø�Ļ���,�Ӷ������൨�̴��ĺϳ���,ͬʱҲ������ѪҺ���ܶ�֬������[19,20]����������������ٴ��������,�����֬���ᣨPUFA�����Խ��������������̴��͵��ܶ�֬����ˮƽ,���߸��ܶ�֬����ˮƽ������ȡ[21]�о�����,������-����������ĸ�֬��ʳ�Ĵ���Ѫ���и���������Ѫ���ܵ��̴�ˮƽ�����Խ���;MEDEIROS��[22]֤����-���������������Ѫ���ܵ��̴����������������ܶ�֬�����̴���LDL-C��ˮƽ;Ҧ�ֽܵ�[18]�о�����,����������1.08% ��-������������������ͷ��Ѫ���ܵ��̴�������������,�����ܶ�֬���������������ߡ����о�����,������������-��������������������۷乤���׳�Ѫ�ܰ����������ܵ��̴��ĺ���,0.02%��0.04%���ܵ��̴�����������������������,��-����������ˮƽΪ0.02%��0.04%��0.06%ʱ���������͵��ܶ�֬������,���߸��ܶ�֬���ĺ���,0.04%����ܶ�֬������������������������,�����ܶ�֬���ĺ�����������������,���������о�������Ǻ�,˵�������˵���������,��-��������н�Ѫ֬�����á�
3.2.2 ��6���乤���׳�֬�ʴ�л���ø���Ե�Ӱ�� ��-��������Ϊ�ṹ���ʺʹ�л��������,��������Ҫ����������,����-�������ijЩ����������ͨ���������ø�Ļ�����ʵ�ֵġ���-������Ľ�Ѫ֬����һ������ͨ���Դ�л�ʵĵ�����ʵ��,��һ��������ͨ�������й�֬�����ͺϳ�øϵ��ʵ�ֵġ�MERSMANN��[23]ͨ���ڳ���������������֬������о�����,PUFA��֬����ĺϳ��ٶ��Լ�FAS��ACC�Ļ��Ծ�����������;TOUSSANT��[24]�ֱ��ø���PUFA�ͱ���֬�����������ι����,���������ι����PUFA�����Ĵ������FAS��ACC��������������ι��������֬���������Ĵ���;BLAKE��[25]�о�����,��ιPUFA�Ĵ��������FAS mRNA�������ι����֬��������6%��
�������о��������,PUFA��֬���ϳ�øϵ����������,���뱾��������һ��,��������-����������ˮƽΪ0.04%ʱ,����������FAS��ACC�Ļ���,�Ӷ���Ѫ֬�����á�
3.3 ��������-����������ˮƽ��������۷乤���׳濹����������Ӱ��
���ɻ�ͨ����ǿ�������öԺ�����������ͽ���,ʹDNA�������ѡ�ͻ��,�Ի���Ŀ�����ϵͳ�ʹ���Ӳ�������,���˵Ļ��۵���ϸ��˥�ϻ�����[7]�����������绯ø��SOD��������������Ҫ�Ŀ�����ø,��������������������ɻ�,�Կ�������������ɻ���ϸ����ɵ���,����ʱ������ϸ��,�����ڿ������������������֡�����ȩ��MDA����Ĥ֬����������Ҫ����,�京����ӵط�ӳ����ϸ�������ɻ����������س̶�[26]����˫˫��[27]�о�����,����Ӻ����;��н�ǿ�Ŀ���������,������Ҫ�Ŀ�������������Ϊ��-������,��������Ӻ�������-������Ũ�ȵ�����,��DPPH���ɻ���ABTS+�������Ҳ������,����������Ҳ����ǿ;�챣�ҵ�[7]�о�����,��-��������������߶���Ѫ��SOD�������Ĺ�������øˮƽ,����MDA����,��߶�����ɻ��������ӳ�����;��������[28]�о�����,��ʳ������-���������������������SOD��CAT����,����MDA����,���������Ŀ��������������о�����,����-����������ˮƽΪ0.02%��0.04%ʱ�����������׳�MDA����,��0.04%ˮƽ��-����������������SOD�Ļ��ԡ�������������һ��,˵���������������˵���-�������ܹ���������׳�Ŀ�����������3.4 ��������-����������ˮƽ��������۷乤���׳�����������Ӱ��
�۷�Ѫϸ����������Ϊ�Ǽ�Ӻ���ϸ������������ָ��[29],Ѫϸ����Ŀ����ҷ�Ӧ��������ø��������������Ծ����������,һ����Ϊ,Ѫϸ����Ŀ�����������������͵�����[30,31,32]����Һ���ἤ���ź�ͨ·�����ģ�antimicrobial peptide�����ܾ�ø�ĺϳ�,������������øԭ����ϵͳ,������Эͬ�����赲��ɱ����������[33]��Ҧ˼���[6]�о�����,��-�����������Դٽ�С��ĵ���-����ϸ����ǻ����ϸ������������,���С��NKϸ���Ļ���,��������ǿС���������;˫���[34]�о�����,��ţ���������Ӹ�����-����������Ӽ������Ը��ƻ���ǿ��ţ����Һ���ߺ�ϸ�����߹���;LESSARD��[35]ͨ���ֱ���ι��ţ����n-6��n-3�����֬���������,����ιʳ����n-3 PUFA��������ţ��ιʳ����n-6 PUFA��������ţ���,�ܰ�ϸ����ֳ��Ӧ������ǿ���ܾ�ø�Ǿ���ϸ���ʱ�ͷŵ�һ��ˮ��øC,��ˮ���ܱ��������ѿ�߸˾�����ɫ�˵�����ȸ��������Ծ�ϸ�����ϵ����ռ�,ʹϸ���ܽ�,�ܾ�ø�����������۶�����������״̬����ʿ�µ�[36]�о�����,����������PUFA���Ը��Ʋ�������Һ����ˮƽ,��߿���Ч��,��������ߵ�PUFA���������Ͳ������ܾ�ø����,�Ӷ������˻������߷�Ӧ�����ӹ�����͵IJ�����֬���ᶼ��ʹ���߹��������½�,ֻ�������Ĺ����������������߹��ܡ����о�����,����������-�����������ˮƽΪ0.02%��0.04%��0.06%ʱ,������������׳��ܾ�ø�ͷ�����ø���ԡ�0.02%����ˮƽ����-���������������������5�����׳�lysozyme��PO��Ա�����,��0.04%����ˮƽ����-�����������������7�����׳�lysozyme��PO��Ա�����,������������-����������ˮƽΪ0.08%ʱ,lysozyme��PO��Ա�����ȴ�������͡��ɴ˿���˵��,���˵���-�����������ǿ�۷��������,����������������������۷���������½���
4 ����
������۷乤���׳���������-����������ˮƽ���׳�Ļ�Ӽ�ʡ������Լ�������������������֬�ʴ�л����һ���̶ȵ�Ӱ��;������۷乤���׳���������-�����������ˮƽΪ0.02%��0.04%ʱ,�׳�Ļ�Ӽ�ʺ������������,�������������俹��������������,Ϊ��Ϊ���˵���-����������ˮƽ���ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
DOI:10.1080/0005772X.2001.11099504URL [��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 3]
[��������: 3]
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.1080/00218839.1987.11100743URL [��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.3969/j.issn.1006-267x.2014.02.018Magsci [��������: 1]

��һ��ʵ�������䷽�Ļ����ϣ��������ͷֱ����0�������飩��25%��50%��75%��100%�����������Ƴ�5�ֵȵ����ܣ��ֵ����ʺ���Ϊ35%������Ϊ15 MJ/kg�����������ϣ�������ѭ����ֳϵͳ����ι5������[����(5.81±0.23) g]8�ܣ����о������������ͬˮƽ���Ͷ������������ܡ�����ɡ�������֬�ʴ�л�Ϳ�����ָ���Ӱ�졣��������������������������ˮƽ�����ߣ�������ض������ʣ�SGR�����������ߺ͵����ƣ���25%����ߣ���������75%��100%�飨<em>P</em><0.05����������ϵ����FCR������ֳ��෴�ı仯���ƣ���25%����ͣ���������100%�飨<em>P</em><0.05����������Ч�ʣ�PER���ͳɻ��ʣ�SR���ڸ�������������죨<em>P</em>>0.05����������ȫ��ֵ����ʺ����������������飨<em>P</em><0.05��������֬���������������ڳ�25%������������飨<em>P</em><0.05���������������ͬˮƽ���Ͷ�ȫ������ʺʹֻҷֺ���������Ӱ�죨<em>P</em>>0.05���������������������ˮƽ�����ߣ�������֬����֬ø��LPL�����Ժ��ܿ�����������T-AOC�����ͣ���������ƻ��������ø��MDH�������������ߣ�����������25%������������죨<em>P</em><0.05������������೬�������绯ø��SOD���������������죨<em>P</em>>0.05�����ɴ˿ɼ��������������ͬˮƽ���Ͷ�������������ܡ�������֬�ʴ�л�Ϳ���������������Ӱ�첻ͬ�������������25%����ʱ���������Ч����ã�����ȫ������ͻ��谭�������������Σ��������Ľ�����
.
DOI:10.3969/j.issn.1006-267x.2014.02.018Magsci [��������: 1]
��һ��ʵ�������䷽�Ļ����ϣ��������ͷֱ����0�������飩��25%��50%��75%��100%�����������Ƴ�5�ֵȵ����ܣ��ֵ����ʺ���Ϊ35%������Ϊ15 MJ/kg�����������ϣ�������ѭ����ֳϵͳ����ι5������[����(5.81±0.23) g]8�ܣ����о������������ͬˮƽ���Ͷ������������ܡ�����ɡ�������֬�ʴ�л�Ϳ�����ָ���Ӱ�졣��������������������������ˮƽ�����ߣ�������ض������ʣ�SGR�����������ߺ͵����ƣ���25%����ߣ���������75%��100%�飨<em>P</em><0.05����������ϵ����FCR������ֳ��෴�ı仯���ƣ���25%����ͣ���������100%�飨<em>P</em><0.05����������Ч�ʣ�PER���ͳɻ��ʣ�SR���ڸ�������������죨<em>P</em>>0.05����������ȫ��ֵ����ʺ����������������飨<em>P</em><0.05��������֬���������������ڳ�25%������������飨<em>P</em><0.05���������������ͬˮƽ���Ͷ�ȫ������ʺʹֻҷֺ���������Ӱ�죨<em>P</em>>0.05���������������������ˮƽ�����ߣ�������֬����֬ø��LPL�����Ժ��ܿ�����������T-AOC�����ͣ���������ƻ��������ø��MDH�������������ߣ�����������25%������������죨<em>P</em><0.05������������೬�������绯ø��SOD���������������죨<em>P</em>>0.05�����ɴ˿ɼ��������������ͬˮƽ���Ͷ�������������ܡ�������֬�ʴ�л�Ϳ���������������Ӱ�첻ͬ�������������25%����ʱ���������Ч����ã�����ȫ������ͻ��谭�������������Σ��������Ľ�����
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.1093/ps/80.6.753URL [��������: 1]
Magsci [��������: 1]
<FONT face=Verdana>���о�̽�����������������ᣨALA��ˮƽ�Զ�����2�������������������ܡ�Ѫ֬������ͼ���֬���ṹ�ɼ�����������øA�Ȼ�ø��ACC)�����֬��ת��ø1��CPT1��mRNA��������Ӱ�졣ѡ�ö����������������������������200ֻ������ֳ�5�飬ÿ��40ֻ���ֱ���ι��ͬALA��������0��0.1%��0.2%��0.3%��0.4%����������Ԥ��7 d������23 d���������������ALAˮƽ��ƽ���ղ�ʳ����ADI����������������ADG�������رȣ�F/G����������Ӱ��������P<0.05��������ALAˮƽ����������Ѫ�����������TG����P=0.001 2�������̴���P=0.005 6���͵��ܶ�֬���ף�LDL��ˮƽ��P=0.000 4������Ѫ����ܶ�֬���ף�HDL��Ӱ�첻������P=0.217 7��������ALAˮƽ�Ը���ͱ��������֬���ᣨSPA��Ӱ�첻������P��0.05������������������ͱ������������֬���ᣨMUFA������Ժ�����P<0.05����������߸���ͱ���������֬���ᣨPUFA����P<0.05����n��3ϵ��PUFA����Ժ�����P<0.000 1��������������n��6/n��3������P<0.01������������n��6ϵ��PUFA��Ժ������첻������P>0��05��������ALAˮƽ�Ը���ACC��CPT1����mRNA��������Ӱ�����������P=0.000 2��P=0.000 1�������������ܡ���֯֬���ṹ�ɺ����ʵȶ�濼�ǣ�������2�����������������п�����0.2%������������������n��3PUFA�Ĺ����������Ʒ��</FONT>
Magsci [��������: 1]
<FONT face=Verdana>���о�̽�����������������ᣨALA��ˮƽ�Զ�����2�������������������ܡ�Ѫ֬������ͼ���֬���ṹ�ɼ�����������øA�Ȼ�ø��ACC)�����֬��ת��ø1��CPT1��mRNA��������Ӱ�졣ѡ�ö����������������������������200ֻ������ֳ�5�飬ÿ��40ֻ���ֱ���ι��ͬALA��������0��0.1%��0.2%��0.3%��0.4%����������Ԥ��7 d������23 d���������������ALAˮƽ��ƽ���ղ�ʳ����ADI����������������ADG�������رȣ�F/G����������Ӱ��������P<0.05��������ALAˮƽ����������Ѫ�����������TG����P=0.001 2�������̴���P=0.005 6���͵��ܶ�֬���ף�LDL��ˮƽ��P=0.000 4������Ѫ����ܶ�֬���ף�HDL��Ӱ�첻������P=0.217 7��������ALAˮƽ�Ը���ͱ��������֬���ᣨSPA��Ӱ�첻������P��0.05������������������ͱ������������֬���ᣨMUFA������Ժ�����P<0.05����������߸���ͱ���������֬���ᣨPUFA����P<0.05����n��3ϵ��PUFA����Ժ�����P<0.000 1��������������n��6/n��3������P<0.01������������n��6ϵ��PUFA��Ժ������첻������P>0��05��������ALAˮƽ�Ը���ACC��CPT1����mRNA��������Ӱ�����������P=0.000 2��P=0.000 1�������������ܡ���֯֬���ṹ�ɺ����ʵȶ�濼�ǣ�������2�����������������п�����0.2%������������������n��3PUFA�Ĺ����������Ʒ��</FONT>
[��������: 1]
[��������: 1]
DOI:10.3969/j.issn.1006-267x.2015.03.014Magsci [��������: 2]
�ڰ봿�������䷽�Ļ�����,�ֱ����6��α-�����Ậ��(0.02%��0.55%��1.08%��1.60%��2.13%��2.65%)��6�������Ậ��(0.86%��1.29%��1.73%��2.16%��2.59%��3.03%),���������͡������͡�����͵���������α-�����ᡢ������ĺ���,���Ƶȵ�����(�ֵ����ʺ���Ϊ30.09%,��֬������Ϊ6.87%)��12�ְ봿����������,̽����ͷ������[��ʼ����Ϊ(59.5±0.5) g]������α-�����ᡢ����������˺�������ֳ�����Ϊα-�����������������2����,����6��,ÿ��4���ظ�,ÿ���ظ�20β,��ֳ����Ϊ85 d���������:��α-������������,���ݻع鷽�̼���õ�,������α-�����Ậ���ֱ�Ϊ1.32%��1.33%ʱ,��ͷ��������������ض������ʺ���С������ϵ��;0.02%�������ָ���������ڳ�1.08%�������������(<em>P</em><0.05),�����ȡ�����ָ�����������������(<em>P</em>>0.05);0.02%��Ѫ���ܵ��̴�����������������������0.55%��1.08%��(<em>P</em><0.05),1.60%��Ѫ����ܶ�֬��������������0.55%��2.13%��(<em>P</em><0.05),0.02%��Ѫ����ܶ�֬����������������������(<em>P</em><0.05)����������������,���ݻع鷽�̼���õ�,�����������Ậ���ֱ�Ϊ2.02%��2.03%ʱ,��ͷ��������������ض������ʺ���С������ϵ��;3.03%��ĸ���ָ������������������(<em>P</em><0.05),�����ȡ�����ָ�����������������(<em>P</em>>0.05);1.29%��Ѫ���ܵ��̴�������������1.73%��(<em>P</em><0.05),0.86%��Ѫ���������������������1.73%��(<em>P</em><0.05),1.29%��Ѫ����ܶ�֬��������������0.86%��3.03%��(<em>P</em><0.05),����Ѫ����ܶ�֬����������������(<em>P</em>>0.05)�����ض������ʡ�����ϵ����Ϊ��Ҫ����ָ��,��ϲ���Ѫ������ָ�������ָ��,�õ��ʺ���ͷ���������������ά�������������������������˵������ᡢ�����Ậ���ֱ�Ϊ1.32%~1.33%��2.02%~2.03%��
DOI:10.3969/j.issn.1006-267x.2015.03.014Magsci [��������: 2]
�ڰ봿�������䷽�Ļ�����,�ֱ����6��α-�����Ậ��(0.02%��0.55%��1.08%��1.60%��2.13%��2.65%)��6�������Ậ��(0.86%��1.29%��1.73%��2.16%��2.59%��3.03%),���������͡������͡�����͵���������α-�����ᡢ������ĺ���,���Ƶȵ�����(�ֵ����ʺ���Ϊ30.09%,��֬������Ϊ6.87%)��12�ְ봿����������,̽����ͷ������[��ʼ����Ϊ(59.5±0.5) g]������α-�����ᡢ����������˺�������ֳ�����Ϊα-�����������������2����,����6��,ÿ��4���ظ�,ÿ���ظ�20β,��ֳ����Ϊ85 d���������:��α-������������,���ݻع鷽�̼���õ�,������α-�����Ậ���ֱ�Ϊ1.32%��1.33%ʱ,��ͷ��������������ض������ʺ���С������ϵ��;0.02%�������ָ���������ڳ�1.08%�������������(<em>P</em><0.05),�����ȡ�����ָ�����������������(<em>P</em>>0.05);0.02%��Ѫ���ܵ��̴�����������������������0.55%��1.08%��(<em>P</em><0.05),1.60%��Ѫ����ܶ�֬��������������0.55%��2.13%��(<em>P</em><0.05),0.02%��Ѫ����ܶ�֬����������������������(<em>P</em><0.05)����������������,���ݻع鷽�̼���õ�,�����������Ậ���ֱ�Ϊ2.02%��2.03%ʱ,��ͷ��������������ض������ʺ���С������ϵ��;3.03%��ĸ���ָ������������������(<em>P</em><0.05),�����ȡ�����ָ�����������������(<em>P</em>>0.05);1.29%��Ѫ���ܵ��̴�������������1.73%��(<em>P</em><0.05),0.86%��Ѫ���������������������1.73%��(<em>P</em><0.05),1.29%��Ѫ����ܶ�֬��������������0.86%��3.03%��(<em>P</em><0.05),����Ѫ����ܶ�֬����������������(<em>P</em>>0.05)�����ض������ʡ�����ϵ����Ϊ��Ҫ����ָ��,��ϲ���Ѫ������ָ�������ָ��,�õ��ʺ���ͷ���������������ά�������������������������˵������ᡢ�����Ậ���ֱ�Ϊ1.32%~1.33%��2.02%~2.03%��
DOI:10.1016/j.preteyeres.2004.06.002URL [��������: 1]
DOI:10.3969/j.issn.1006-267x.2014.04.011Magsci [��������: 1]
�����齫������θ���������붯̬����ʵʱ��¼�������ϣ������˸߾���ȫ��������в��ü�˿��ĸ��<em>Candida utilis</em>��CU������ˮƽ��������θ�������Ժ���ά�����Ӱ�졣��������θҺ����5ͷ�����ں�˹̹��ţ�������Ϊ6�飬����CU����ˮƽ�ֱ�Ϊ0�������飩��0.53×10<sup>6</sup>��1.07×10<sup>6</sup>��1.60×10<sup>6</sup>��2.13×10<sup>6</sup>��2.67×10<sup>6</sup> CFU/mL���ⶨ72 h�ۼƲ���������������ѧ��������θ��������ָ�ꡣ�����������CU����ˮƽ�����ߣ������Ӫ�����ʣ������ʡ�����ϴ����ά������ϴ����ά��������ʧ�ʡ���ʱ����������72 h�ۻ�����������������ѧ�������������������ʹﵽ1/2��������������ʱ�䣩�����ﵰ�������ܻӷ���֬����Ũ���Լ����ᡢ������֧��֬���Ậ�����������������ߣ�<em>P</em><0.050��<em>P</em><0.001���������ᡢ���Ậ���Լ��������������������ͣ�<em>P</em><0.001������ˣ�����CU�������θ������ά�����뷢��Ч�ʡ���ȫ������������£�CU��������ˮƽΪ2.67×10<sup>6</sup> CFU/mL��
DOI:10.3969/j.issn.1006-267x.2014.04.011Magsci [��������: 1]
�����齫������θ���������붯̬����ʵʱ��¼�������ϣ������˸߾���ȫ��������в��ü�˿��ĸ��<em>Candida utilis</em>��CU������ˮƽ��������θ�������Ժ���ά�����Ӱ�졣��������θҺ����5ͷ�����ں�˹̹��ţ�������Ϊ6�飬����CU����ˮƽ�ֱ�Ϊ0�������飩��0.53×10<sup>6</sup>��1.07×10<sup>6</sup>��1.60×10<sup>6</sup>��2.13×10<sup>6</sup>��2.67×10<sup>6</sup> CFU/mL���ⶨ72 h�ۼƲ���������������ѧ��������θ��������ָ�ꡣ�����������CU����ˮƽ�����ߣ������Ӫ�����ʣ������ʡ�����ϴ����ά������ϴ����ά��������ʧ�ʡ���ʱ����������72 h�ۻ�����������������ѧ�������������������ʹﵽ1/2��������������ʱ�䣩�����ﵰ�������ܻӷ���֬����Ũ���Լ����ᡢ������֧��֬���Ậ�����������������ߣ�<em>P</em><0.050��<em>P</em><0.001���������ᡢ���Ậ���Լ��������������������ͣ�<em>P</em><0.001������ˣ�����CU�������θ������ά�����뷢��Ч�ʡ���ȫ������������£�CU��������ˮƽΪ2.67×10<sup>6</sup> CFU/mL��
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.1016/j.nutres.2007.04.003URL [��������: 1]
[��������: 1]
DOI:10.1093/jn/111.1.146URL [��������: 1]
DOI:10.1093/jn/120.12.1727URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1046/j.1461-0248.2001.00279.xURL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
.
[��������: 1]
DOI:10.3168/jds.S0022-0302(04)70040-5URL [��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}