Expression Profiling and Functional Characterization of Rice Transcription Factor OsWRKY68
CHEN Yue, WANG TianXingZi, YANG Shuo, ZHANG Tong, MA JinJiao, YAN GaoWei, LIU YuQing, ZHOU Yan, SHI JiaNan, LAN JinPing, WEI Jian, DOU ShiJuan, LIU LiJuan, YANG Ming, LI LiYun, LIU GuoZhen,College of Life Sciences, Hebei Agricultural University, Baoding 071001, Hebei
Abstract 【Objective】 There are nearly 100 WRKY transcription factor members in rice genome, many of them are involved in plant growth and development, biotic and abiotic stress responses. Molecular biology & bioinformatics lab identified that the expression of OsWRKY68 protein was induced after inoculation with Xanthomonas oryzae pv. oryzae (Xoo) in rice. The aim of this study is attempt to further explore the function of OsWRKY68. 【Method】Rice TP309 samples of different tissues at different developmental stages, including germination, seedling, tillering, booting and flowering stages of root, stem, leaf, sheath, cushion, panicle, anther, husk, seed, abiotic stress (4℃, 44℃, 48℃, submerge, NaCl, PEG, constant light, constant dark) and hormone treatments (abscisic acid, methyl jasmonate, salicylic acid, ethephon) were collected. Total protein were extracted and analyzed by Western blot (WB) systematically using OsWRKY68-specific antibody. The expression patterns of OsWRKY68 protein isolated from different tissues at different developmental stages, and tissues obtained from abiotic stresses and hormone treatments were investigated. RNA interfering vector was constructed and transformed to wildtype TP309 rice variety via Agrobacterium tumefaciens strategy. Identification of transgenic plants were carried out by PCR and WB. The phenotype of OsWRKY68 RNAi transgenic plants were monitored and plant height, tiller number, spike length, spikelet number and seed-setting rate were measured.【Result】By comparing the abundance of OsWRKY68 protein in different tissues, it was found that OsWRKY68 protein was expressed in a constitutive way during the normal growth and development of rice, the abundance of OsWRKY68 protein expressed among different tissues were not varied too much. However, different levels of OsWRKY68 were observed. The expression level of OsWRKY68 in anthers at flowering stage was higher than that in mature panicles, panicle axis and husk. It was not expressed in sheaths at tillering and booting stages, but it was expressed in sheaths at flowering stage. In panicles, the abundance of OsWRKY68 was decreased gradually along with the growth of the young panicle. By investigating the expression patterns of OsWRKY68 protein under abiotic stress and hormone treatments, it was found that the abundance of OsWRKY68 protein decreased steadily under salt stress. The expression of OsWRKY68 protein increased steadily at constant light treatment, a specific band (designated as OsWRKY68 +) with higher molecular weight appeared at three days and enhanced in the following timepoints. After methyl jasmonate (MeJA) and ethephon (ET) treatments, OsWRKY68 + band appeared also and its intensity increased as the treatments continues. Four homozygous OsWRKY68 RNAi transgenic lines (Y316, Y317, Y326 and Y337) were checked by PCR and WB analyses and verified at T3 generation. The abundance of OsWRKY68 protein in RNAi transgenic plants was lower than that in wildtype TP309. Phenotypic investigation revealed significant reduction in plant height, tiller number and seed setting rate in transgenic plants.【Conclusion】Rice OsWRKY68 protein plays an important role in the process of normal growth and development of rice. Knocking down the abundance of OsWRKY68 protein via RNAi affected the normal growth of rice. In addition, the data of expression patterns suggested that the function of OsWRKY68 protein may be involved with salt stress, light, MeJA and ethene-mediated signal transduction pathways. Keywords:rice;WRKY transcription factor;expression patterns;western blot;RNA interference
PDF (1035KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 陈悦, 王田幸子, 杨烁, 张彤, 马金姣, 燕高伟, 刘玉晴, 周艳, 史佳楠, 兰金苹, 魏健, 窦世娟, 刘丽娟, 杨明, 李莉云, 刘国振. 水稻转录因子OsWRKY68蛋白质的表达特征及其功能特性[J]. 中国农业科学, 2019, 52(12): 2021-2032 doi:10.3864/j.issn.0578-1752.2019.12.001 CHEN Yue, WANG TianXingZi, YANG Shuo, ZHANG Tong, MA JinJiao, YAN GaoWei, LIU YuQing, ZHOU Yan, SHI JiaNan, LAN JinPing, WEI Jian, DOU ShiJuan, LIU LiJuan, YANG Ming, LI LiYun, LIU GuoZhen. Expression Profiling and Functional Characterization of Rice Transcription Factor OsWRKY68[J]. Scientia Acricultura Sinica, 2019, 52(12): 2021-2032 doi:10.3864/j.issn.0578-1752.2019.12.001
pTCK303和pCAMBIA2300质粒由中国科学院遗传与发育生物学研究所江光怀博士提供。pEASY-T1质粒购自生工生物工程(上海)股份有限公司。含有水稻OsWRKY68全长的cDNA质粒AK072938(Os04g51560)购自日本农业生物资源研究所水稻基因组资源中心(Rice Genome Resource Center,National Institute of Agrobiological Sciences)。
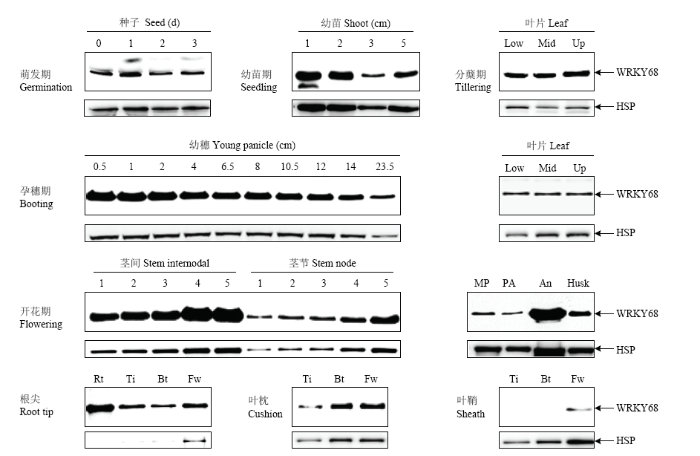
WB条件:上样量10 μL,电泳条件:80 V,20 min;160 V,90 min,胶浓度10% tricine,电泳槽Mini PROTEAN Tetra cell,转膜条件:100 V,60 min,膜PVDF,一抗:抗OsWRKY68抗体,抗OsHSP82抗体,二抗:羊抗兔二抗,羊抗鼠二抗。HSP:水稻HSP82;WRKY68:水稻OsWRKY68。下同。An:花药;Bt:孕穗期;Fw:开花期;Husk:颖壳;Low:叶片下部;Mid:叶片中部;MP:成熟穗;PA:穗轴;Rt:幼根;Ti:分蘖期;Up:叶片上部 Fig. 1Expression profiling of OsWRKY68 protein in different tissues at different developmental stages
WB conditions: Sample loading volume 10 μL. Electrophoresis: 80 V, 20 min, 160 V, 90 min, Gel concentration 10% tricine. Electrophoresis apparatus Mini PROTEAN Tetra cell. Membrane (PVDF) transfer: 100 V, 60 min, Primary Antibody: anti-OsWRKY68 polyclonal antibody (BPI, AbP80049-A-S), anti-HSP82 monoclonal antibody (BPI, AbM51099-31-PU), Secondary antibody: goat anti-rabbit secondary antibody (BPI, AbP-71001-D-HRP), goat anti-mouse secondary antibody (BPI, AbP-71003-D-HRP). HSP: Rice OsHSP82; WRKY68: Rice OsWRKY68. The same as below. An: Anther; Bt: Booting; Fw: Flowering; Low: Lower part of leave; Mid: Middle part of leave; MP: Mature panicle; PA: Panicle axis; Rt: Root; Ti: Tillering; Up: Upper part of leave
A:OsWRKY68蛋白质在水稻苗期NaCl(200 mmol·L-1)处理下的表达特征;B:OsWRKY68蛋白质在水稻离体叶片恒光处理下的表达特征。CK:对照;CL:恒光处理 Fig. 2Expression profiling of OsWRKY68 protein under abiotic stresses
A: Expression profiling of rice OsWRKY68 protein at seedling stage under 200 mmol·L-1 NaCl treatment; B: Expression profiling of rice OsWRKY68 protein under constant light in vitro. CK: Control; CL: Constant light
VIANA VE, BUSANELLOC , DA MAIAL C, PEGORAROC, COSTA DEOLIVEIRA A. Activation of rice WRKY transcription factors: an army of stress fighting soldiers? Current Opinion Plant Biology, 2018,45:268-275. DOI:10.1016/j.pbi.2018.07.007URL [本文引用: 1]
ROSS CA, LIUY, SHEN QJ . The WRKY gene family in rice ( Oryza sativa). Journal of Integrative Plant Biology, 2007,49(6):827-842. [本文引用: 1]
Rice WRKY WorkingGroup . Nomenclature report on rice WRKY's - Conflict regarding gene names and its solution Rice (New York), 2012,5(1):3. [本文引用: 1]
CAI YH, CHEN XJ, XIEK, XING QK, WU YW, LIJ, DU CH, SUN ZX, GUO ZJ . Dlf1, a WRKY transcription factor, is involved in the control of flowering time and plant height in rice Plos One, 2014,9(7):e102529. DOI:10.1371/journal.pone.0102529URL [本文引用: 1]
LEEH, CHAJ, CHOIC, CHOIN, JI HS, PARK SR, LEES, HWANG DJ . Rice WRKY11 plays a role in pathogen defense and drought tolerance Rice (New York), 2018,11(1):5. [本文引用: 1]
QIU DY, XIAOJ, XIE WB, LIU HB, LI XH, XIONG LZ, WANG SP . Rice gene network inferred from expression profiling of plants overexpressing OsWRKY13, a positive regulator of disease resistance Molecular Plant, 2008,1(3):538-551. [本文引用: 4]
XIAOJ, CHENG HT, LI XH, XIAO JH, XU CG, WANG SP . Rice WRKY13 regulates cross talk between abiotic and biotic stress signaling pathways by selective binding to different cis-elements Plant Physiology, 2013,163(4):1868-1882. DOI:10.1104/pp.113.226019URL [本文引用: 1]
QIU DY, XIAOJ, DING XH, XIONGM, CAIM, CAO YL, LI XH, XU CG, WANG SP . OsWRKY13 mediates rice disease resistance by regulating defense-related genes in salicylate- and jasmonate- dependent signaling Molecular Plant-Microbe Interactions, 2007,20(5):492. DOI:10.1094/MPMI-20-5-0492URL [本文引用: 2]
ABBRUSCATOP, NEPUSZT, MIZZIL, CORVO MD, MORANDINIP, FUMASONII, MICHELC, PACCANAROA, GUIDERDONIE, SCHAFFRATHU . OsWRKY22, a monocot WRKY gene, plays a role in the resistance response to blast Molecular Plant Pathology, 2012,13(8):828-841. DOI:10.1111/mpp.2012.13.issue-8URL [本文引用: 1]
PENG XX, HU YJ, TANG XK, ZHOU PL, DENG XB, WANG HH . Isolation and expression profiles jasmonic acid application and of rice WRKY30 induced by fungal pathogen infection Scientia Agricultura Sinica, 2011,21(44):2454-2461. (in Chinese) [本文引用: 1]
SHEN HS, LIU CT, ZHANGY, MENG XP, ZHOUX, CHU CC, WANG XP . OsWRKY30 is activated by MAP kinases to confer drought tolerance in rice Plant Molecular Biology, 2012,80(3):241-253. DOI:10.1007/s11103-012-9941-yURL [本文引用: 1]
ZHANGJ, PENG YL, GUO ZJ . Constitutive expression of pathogen-inducible OsWRKY31 enhances disease resistance and affects root growth and auxin response in transgenic rice plants Cell Research, 2008,18(4):508-521. DOI:10.1038/cr.2007.104 [本文引用: 1]
HAN M KC, LEEJ, LEE SK, JEON JS . OsWRKY42 represses OsMT1d and induces reactive oxygen species and leaf senescence in rice Molecules & Cells, 2014,37(7):532. [本文引用: 1]
PILLAI SE, KUMARC, PATEL HK, SONTI RV . Overexpression of a cell wall damage induced transcription factor, OsWRKY42, leads to enhanced callose deposition and tolerance to salt stress but does not enhance tolerance to bacterial infection BMC Plant Biology, 2018,18(1):177. DOI:10.1186/s12870-018-1391-5 [本文引用: 2]
SHIMONOM, KOGAH, AKAGIA, HAYASHIN, GOTOS, SAWADAM, KURIHARAT, MATSUSHITAA, SUGANOS, JIANG CJ . Rice WRKY45 plays important roles in fungal and bacterial disease resistance Molecular Plant Pathology, 2011,13(1):83-94. [本文引用: 1]
SHIMONOM, SUGANOS, NAKAYAMAA, JIANG CJ, ONOK, TOKIS, TAKATSUJIH . Rice WRKY45 plays a crucial role in benzothiadiazole-inducible blast resistance The Plant Cell, 2007,19(6):2064-2076. DOI:10.1105/tpc.106.046250URL [本文引用: 2]
CHENG HT, LIU HB, DENG YK, XIAO JX, LI XH, WANG SP . The WRKY45-2-WRKY13-WRKY42 transcriptional regulatory cascade is required for rice resistance to fungal pathogen Plant Physiology, 2015,167(3):1087-1099. DOI:10.1104/pp.114.256016URL [本文引用: 3]
QIU YP, YU DQ . Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis. Environmental and Experimental Botany, 2009,65:35-47. [本文引用: 1]
WANGJ, ZHOUL, SHIH, CHERNM, YUH, YIH, HEM, YIN JJ, ZHU XB, LIY, LI WT, LIU JL, WANG JC, CHEN XQ, QINGH, WANG YP, LIU GF, WANG WM, LIP, WU XJ, ZHU LH, ZHOU JM, RONALDP, LI SG, LI JY, CHEN XW . A single transcription factor promotes both yield and immunity in rice Science, 2018,361:1026-1028. DOI:10.1126/science.aat7675URL [本文引用: 1]
WEIT, OUBIN, LI JB, ZHAOY, GUO DS, ZHU YY, CHEN ZL, GU HY, LI CY, QIN GJ . Transcriptional profiling of rice early response to Magnaporthe oryzae identified OsWRKYs as important regulators in rice blast resistance. PLoS ONE, 2013,8(3):e59720. [本文引用: 1]
RAINERIJ, WANG SH, PELEGZ, BLUMWALDE, CHAN RL . The rice transcription factor OsWRKY47 is a positive regulator of the response to water deficit stress Plant Molecular Biology, 2015,88(4/5):401-413. DOI:10.1007/s11103-015-0329-7URL [本文引用: 1]
TIAN XJ, LI XF, ZHOU WJ, REN YK, WANG ZY, LIU ZQ, TANG JQ, TONG HN, FANGJ, BU QY . Transcription factor OsWRKY53 positively regulates brassinosteroid signaling and plant architecture Plant Physiology, 2017,175(3):1337-1349. DOI:10.1104/pp.17.00946URL [本文引用: 1]
CHUJOT, TAKAIR, AKIMOTO-TOMIYAMAC, ANDOS, MINAMIE, NAGAMURAY, KAKUH, SHIBUYAN, YASUDAM, NAKASHITAH, UMEMURAK, OKADAA, OKADAK, NOJIRIH, YAMANEH . Involvement of the elicitor-induced gene OsWRKY53 in the expression of defense-related genes in rice Biochimica et Biophysica Acta-Biomembranes, 2007,1769(7/8):497-505. DOI:10.1016/j.bbaexp.2007.04.006URL [本文引用: 1]
PENGY, BARTLEY LE, CHEN XW, DARDICKC, CHERNM, RUANR, CANLAS PE, RONALD PC . OsWRKY62 is a negative regulator of basal and Xa21-mediated defense against Xanthomonas oryzae pv. oryzae in rice. Molecular Plant, 2008,1(3):446-458. [本文引用: 1]
VO K TX, KIM CY, HOANG TV, LEE SK, SHIRSEKARG, SEO YS, LEE SW, WANG GL, JEON JS . OsWRKY67 plays a positive role in basal and XA21-mediated resistance in rice. Frontiers in Plant Science, 2018,8:2220. [本文引用: 1]
LIU XQ, BAI XQ, WANG XJ, CHU CC . OsWRKY71, a rice transcription factor, is involved in rice defense response Journal of Plant Physiology, 2007,164(8):969-979. DOI:10.1016/j.jplph.2006.07.006URL [本文引用: 1]
SONGY, CHEN LG, ZHANG LP, YU DQ . Overexpression of OsWRKY72 gene interferes in the abscisic acid signal and auxin transport pathway of Arabidopsis. Journal of Biosciences, 2010,35(3):459-471. [本文引用: 1]
DAI XY, WANG YY, ZHANG WH . OsWRKY74, a WRKY transcription factor, modulates tolerance to phosphate starvation in rice Journal of Experimental Botany, 2016,67(3):947-960. DOI:10.1093/jxb/erv515URL [本文引用: 1]
SEO YS, CHERNM, BARTLEY LE, HANM, JUNG KH, LEEI, WALIAH, RICHTERT, XUX, CAO PJ . Towards establishment of a rice stress response interactome Plos Genetics, 2011,7(4):e1002020. DOI:10.1371/journal.pgen.1002020URL [本文引用: 1]
LIU QQ, ZHANG CQ, XUY, LUY, YU HX, GU MH . The WRKY transcription factor OsWRKY78 regulates stem elongation and seed development in rice Planta, 2011,234(3):541-554. DOI:10.1007/s00425-011-1423-yURL [本文引用: 1]
WANG HH, HAO JJ, CHEN XJ, HAO ZN, WANGX, LOU YG, PENG YL, GUO ZJ . Overexpression of rice WRKY89 enhances ultraviolet B tolerance and disease resistance in rice plants Plant Molecular Biology, 2007,65(6):799-815. DOI:10.1007/s11103-007-9244-xURL [本文引用: 1]
YANGS, ZHOUL, MIAO LY, SHI JN, SUN CQ, FANW, LAN JP, CHENH, LIU LJ, DOU SJ, LIU GZ, LI LY . The expression and binding properties of the rice WRKY68 protein in the Xa21-mediated resistance response to Xanthomonas oryzae pv.Oryzae. Journal of Integrative Agriculture, 2016,15(11):2451-2460. [本文引用: 6]
NURUZZAMAN M SA, SATOHK, KUMARA, LEUNGH, KIKUCHIS . Comparative transcriptome profiles of the WRKY gene family under control, hormone-treated, and drought conditions in near-isogenic rice lines reveal differential, tissue specific gene activation Journal of Plant Physiology, 2014,171(1):2-13. DOI:10.1016/j.jplph.2013.09.010URL [本文引用: 2]
SATOHK, SAJIS, ITOS, SHIMIZUH, SAJIH, KIKUCHIS . Gene response in rice plants treated with continuous fog influenced by pH, was similar to that treated with biotic stress Rice, 2014,7(1):10. DOI:10.1186/s12284-014-0010-9URL [本文引用: 2]
RAMAMOORTHYR, JIANG SY, KUMARN, VENKATESH PN, RAMACHANDRANS . A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments Plant Cell Physiology, 2008,49(6):865-879. DOI:10.1093/pcp/pcn061URL [本文引用: 2]
LIU GZ, LIU SQ, WUL, XU NZ . Antibody-based rice proteomics- the beginning and perspectives Scientia Sinica Viate, 2011,41(3):173-177. (in Chinese) [本文引用: 1]
ZHANG JS, MA JJ, ZHANGT, CHENY, WEIJ, ZHANGL, SHI JN, XUS, YAN GW, DU TM, DOU SJ, LI LY, LIU LJ, LIU GZ . The establishment and application of rice protein sample library RiceS-A300 Scientia Agricultura Sinica, 2018,51(19):3625-3638. (in Chinese) [本文引用: 1]
LI XM, BAIH, WANG XY, LI LY, CAO YH, WEIJ, LIU YM, LIU LJ, GONG XD, WUL, LIU SQ, LIU GZ . Identification and validation of rice reference proteins for western blotting Journal of Experimental Botany, 2011,62:4763-4772. DOI:10.1093/jxb/err084URL [本文引用: 1]
BAIH, LAN JP, GANQ, WANG XY, HOU MM, CAO YH, LI LY, LIU LJ, HAO YJ, YIN CC, WUL, ZHU LH, LIU GZ . Identification and expression analysis of components involved in rice Xa21-mediated disease resistance signaling. Plant Biology, 2012,14:914-922. [本文引用: 1]
LANDIS, HAUSMAN JF, GUERRIEROG, ESPOSITOS . Poaceae vs. abiotic stress: Focus on drought and salt stress, recent insights and perspectives Frontiers in Plant Science, 2017,8:1214. DOI:10.3389/fpls.2017.01214URL [本文引用: 1]
TAOZ, KOU YJ, LIU HB, LI XH, XIAO JH, WANG SP . OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice Journal of Experimental Botany, 2011,62(14):4863-4874. DOI:10.1093/jxb/err144URL [本文引用: 2]
CHENH, LAI ZB, SHI JW, XIAOY, CHEN ZX, XU XP . Roles of Arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biology, 2010,10:281. [本文引用: 1]
LI LY, SHI JN, YANGS, SUN CQ, LIU GZ . Functional annotation of rice WRKY transcription factors based on their transcriptional features Hereditas, 2016,38(2):126-136. (in Chinese) [本文引用: 1]
 ,河北农业大学生命科学学院,河北保定 071001
,河北农业大学生命科学学院,河北保定 071001
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT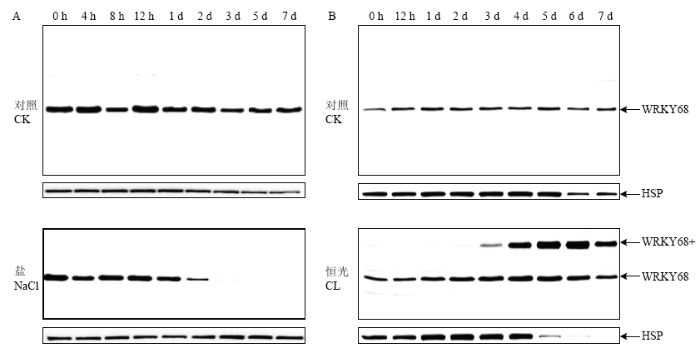 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT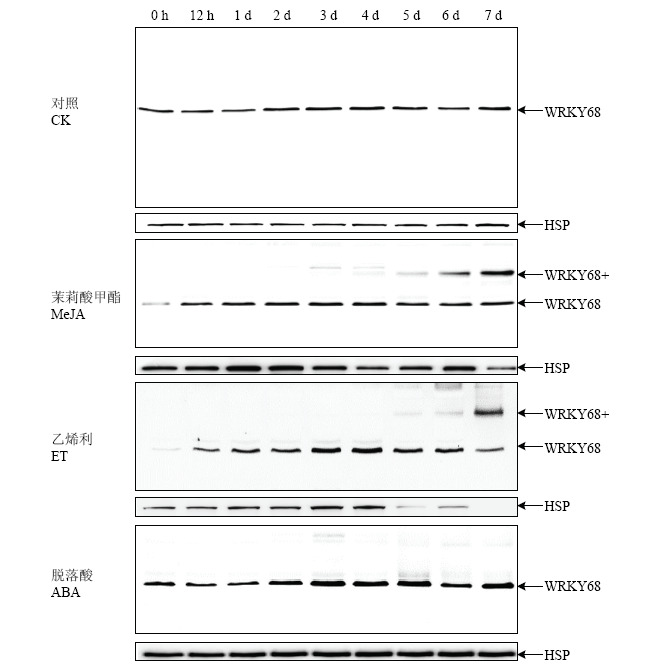 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT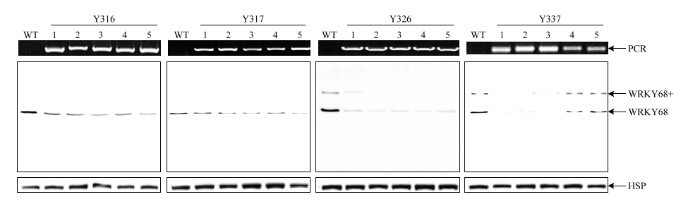 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT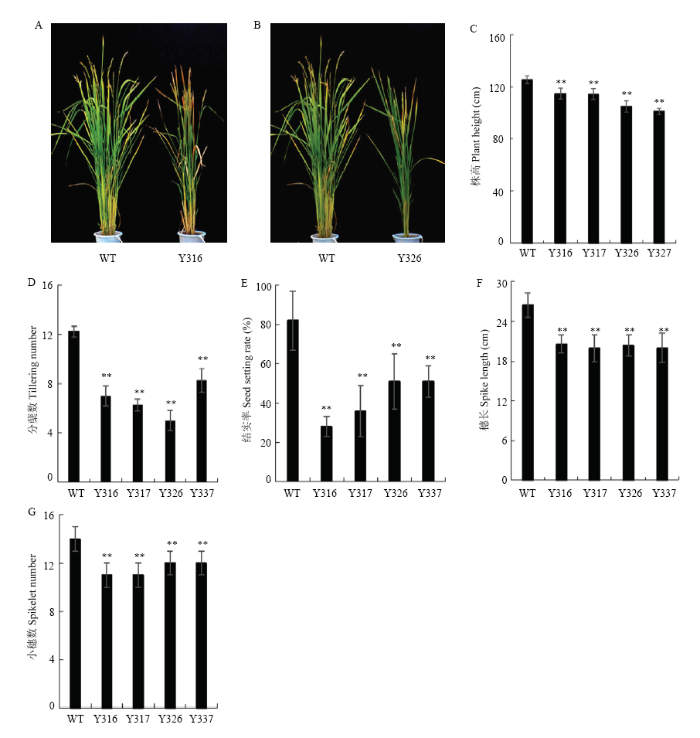 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}