 ,中国农业科学院蜜蜂研究所农业部授粉昆虫生物学重点实验室,北京100093
,中国农业科学院蜜蜂研究所农业部授粉昆虫生物学重点实验室,北京100093Infection Status and Genetic Relationship of Crithidia bombi in Different Regions of China
TANG YuJie, WANG LiuHao, LI Kai, LI JiLian,Key Laboratory of Pollinating Insect Biology of the Ministry of Agriculture, Institute of Apicultural Research, Chinese Academy of Agricultural Sciences, Beijing 100093通讯作者:
责任编辑: 岳梅
收稿日期:2018-10-12接受日期:2018-11-7网络出版日期:2019-03-16
| 基金资助: |
Received:2018-10-12Accepted:2018-11-7Online:2019-03-16
作者简介 About authors
唐裕杰,E-mail: whtangyujie1993@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (484KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
唐裕杰, 王刘豪, 李凯, 李继莲. 我国不同地区熊蜂短膜虫的感染情况及亲缘关系[J]. 中国农业科学, 2019, 52(6): 1110-1118 doi:10.3864/j.issn.0578-1752.2019.06.014
TANG YuJie, WANG LiuHao, LI Kai, LI JiLian.
0 引言
【研究意义】熊蜂(Bombus spp.)是自然生态系统中重要的授粉昆虫,在维持自然界和农业生态系统平衡方面发挥着至关重要的作用。我国是全世界熊蜂种类最丰富的国家之一[1,2]。目前全球已知的熊蜂种类约250种[3]。我国已知的熊蜂种类多达125种,约占全球熊蜂已知物种的50%[4]。熊蜂具有个体大、寿命长、吻较长、全身密被绒毛等特点,采集能力强于蜜蜂,对弱光、低温等较恶劣的环境条件有良好的适应能力[5]。目前,我国饲养熊蜂的主要用途是为设施农业、大田作物、果蔬等授粉,同时也对外出口至其他国家[6]。商品化熊蜂群的应用规模越来越大,其在不同国家间的进出口不可避免会传播部分病原虫害,其中寄生虫是影响熊蜂生态分布和商业化发展的主要因素。熊蜂短膜虫(Crithidia bombi)是危害熊蜂健康的常见寄生虫之一,其主要寄生于熊蜂肠道内,且传播广泛、数量多,对熊蜂的繁育与产业化产生巨大的影响[7,8]。在野外环境中,熊蜂可能会从巢内的同伴或在被污染的花上觅食的同伴处感染这种寄生虫[9]。研究熊蜂短膜虫的感染情况,关注熊蜂健康,保护熊蜂的物种多样性,对更好地保护生态系统具有重要意义。【前人研究进展】熊蜂短膜虫属于动鞭毛虫纲、动质目、锥虫亚目、短膜虫属,其形态特点为只有锥虫亚目6个发育期中的无鞭毛期和前鞭毛期,无鞭毛期是随昆虫粪便排出的感染期,而前鞭毛期为肠道寄生期[10]。早期的研究报道证实,在熊蜂肠道中发现了短膜虫属的Crithidia bombi和细滴虫属的Leptomonas sp.。武文杰认为自然界中的熊蜂短膜虫存在着多重感染的现象;熊蜂短膜虫对熊蜂的成活率有较为明显的影响,且不同株的短膜虫对熊蜂的影响不同[11,12,13]。在真核生物的核糖体中,内转录间隔区(internal transcribed space,ITS)位于18S和5.8S rDNA之间以及5.8S和28S rDNA之间,故ITS区包括ITS-1区和ITS-2区。因为ITS区域两端分别为大亚基和小亚基,所以不像rRNA基因一样受到功能上的限制,且18S、5.8S和28S基因组序列在绝大多数的生物中趋于保守,在生物种间变化相对较小,稳定性强,而ITS区域进化速度较编码区快,在核苷酸序列和长度上也存在很强的变异性[14,15,16]。基于这些特点,ITS区域成为许多寄生虫分子分类学研究的有用标记。ZHU等[17]通过对15种蛔虫rDNA的ITS-1区和ITS-2区进行测序,发现了不同种的ITS序列存在不同程度的差异,进而分析了15种蛔虫的系统进化;张立海等[18]利用2个引物扩增松材线虫(Bursaphelenchus xylophilus)和拟松材线虫(B. mucronatus)的ITS序列,得到ITS-1区的扩增产物大小约308 bp,该区可作为松材线虫PCR鉴定的靶向序列,并测出这两个种间的ITS序列差异为32—39 bp,而松材线虫种内差异不超过1 bp;周荣琼等[19]通过对国内3株安氏隐孢子(Cryptosporidium andersoni)的ITS-1序列进行扩增并分析,发现3株安氏隐孢子的ITS-1序列基本一致,仅其中1株与其他2株有3个碱基的差异,与GenBank注册的C. muris和C. parvum存在种间差异,且差异显著,这充分说明了ITS-1序列可以作为C. andersoni种的遗传标记,从而为隐孢子虫属的种间鉴定打下了基础;邹志文等[20]研究发现,ITS序列片段分析结果支持尼氏真绥螨(Euseius nicholsi)和卵圆真绥螨(E. ovalis)现在的分类地位,而小新绥螨属与钝绥螨属似乎未达到属间差异,其分类地位有待进一步确定;艾琳等[21]扩增了黑龙江绒山羊和绵羊的东毕吸虫(Orientobilharzia spp.)的ITS序列,发现这两种东毕吸虫的ITS序列总长度均为875 bp,且序列相似性为99.9%,证实了黑龙江绒山羊和绵羊的东毕吸虫为同一个种;LIU等[22]通过测得不同牛梨形虫ITS序列来构建牛梨形虫的系统发育树,发现所有的牛巴贝斯虫和泰勒虫各独立占有一支,故通过ITS序列差异来反映这些种的亲缘关系更为理想。上述研究均证实了随着分子生物学技术的发展,rDNA-ITS序列分析已经广泛应用于原虫和蠕虫等寄生虫的分类、鉴定、诊断和系统发育的研究,且能够取得很好的效果。【本研究切入点】近年来我国关于熊蜂短膜虫的研究报道相对缺乏。本研究利用特异的ITS引物扩增待检测的熊蜂肠道总DNA,之后进行凝胶电泳,通过是否扩增出675 bp的熊蜂短膜虫ITS基因片段来判断待检测的蜂群是否感染了熊蜂短膜虫,通过进一步测序,分析不同蜂种、不同地区寄生的熊蜂短膜虫之间的亲缘关系。【拟解决的关键问题】调查研究我国内蒙古、甘肃、青海、四川4个省(自治区)熊蜂短膜虫的感染率,并由其序列同源性构建系统发育树,旨在进一步探究我国部分地区短膜虫的流行规律及不同地区、不同蜂种间寄生的短膜虫的亲缘关系,为商品化熊蜂的短膜虫病的进出口检疫以及熊蜂短膜虫的分类鉴定提供理论依据。1 材料与方法
1.1 熊蜂样本采集
供试熊蜂样本均于2017年7—8月采自内蒙古、甘肃、青海、四川等地。在采集样本时,将捕虫网捕获得到的熊蜂置于无水乙醇中,妥善保存熊蜂样本,同时记录采集地区的海拔高度和经纬度(表1)。之后对不同蜂种、不同地区熊蜂短膜虫的感染率进行卡方检验分析(SPSS 22.0)。Table 1
表1
表1各地区所调查的熊蜂的种类和数量
Table 1
| 省(自治区) Province (autonomous region) | 纬度 Latitude | 经度 Longitude | 海拔 Elevation (m) | 蜂种 Species | 采集数 Collection number | 感染数 Infected number |
|---|---|---|---|---|---|---|
| 内蒙古 Inner Mongolia | 48°41.448′ | 122°45.290′ | 419 | 密林熊蜂B. patagiatus | 41 | 5 |
| 48°41.448′ | 122°45.290′ | 419 | 明亮熊蜂B. lucorum | 4 | 3 | |
| 49°31.497′ | 117°16.951′ | 712 | 西伯熊蜂B. sibiricus | 50 | 0 | |
| 49°28.953′ | 119°40.439′ | 602 | 盗熊蜂B. filchnerae | 2 | 2 | |
| 49°28.953′ | 119°40.439′ | 602 | 藓状熊蜂B. muscorum | 1 | 0 | |
| 甘肃 Gansu | 36°34.728′ | 102°58.077′ | 2096 | 黑尾熊蜂B. melanurus | 7 | 1 |
| 36°34.728′ | 102°58.077′ | 2096 | 西伯熊蜂B. sibiricus | 1 | 1 | |
| 36°49.855′ | 102°39.003′ | 2210 | 火红熊蜂B. pyrosoma | 19 | 13 | |
| 36°49.855′ | 102°39.003′ | 2210 | 明亮熊蜂B. lucorum | 2 | 1 | |
| 36°49.855′ | 102°39.003′ | 2210 | 盗熊蜂B. filchnerae | 3 | 1 | |
| 34°25.522′ | 102°17.784′ | 3454 | 稳纹熊蜂B. waltoni | 7 | 0 | |
| 34°25.522′ | 102°17.784′ | 3454 | 拉熊蜂B. laesus | 1 | 0 | |
| 34°42.456′ | 102°30.892′ | 3288 | 密林熊蜂B. patagiatus | 52 | 12 | |
| 34°42.456′ | 102°30.892′ | 3288 | 兴熊蜂B. impetuosus | 2 | 0 | |
| 38°09.402′ | 100°11.705′ | 2912 | 红束熊蜂B. rufofasciatus | 103 | 15 | |
| 38°09.402′ | 100°11.705′ | 2912 | 苏氏熊蜂B. sushkini | 3 | 2 | |
| 38°09.402′ | 100°11.705′ | 2912 | 伪猛熊蜂B. personatus | 2 | 0 | |
| 38°09.402′ | 100°11.705′ | 2912 | 克什米尔熊蜂B. kashmirensis | 2 | 1 | |
| 38°09.402′ | 100°11.705′ | 2912 | 越熊蜂B. supremus | 3 | 1 | |
| 37°56.221′ | 100°57.951′ | 3418 | 猛熊蜂B. difficillimus | 4 | 1 | |
| 37°59.971′ | 100°45.020′ | 3214 | 拉达克熊蜂B. ladakhensis | 6 | 0 | |
| 37°59.971′ | 100°45.020′ | 3214 | 昆仑熊蜂B. keriensis | 7 | 1 | |
| 38°13.043′ | 100°56.944′ | 2835 | 小雅熊蜂B. lepidus | 3 | 0 | |
| 青海 Qinghai | 37°10.689′ | 102°03.091′ | 2596 | 明亮熊蜂B. lucorum | 10 | 9 |
| 37°10.689′ | 102°03.091′ | 2596 | 火红熊蜂B. pyrosoma | 11 | 7 | |
| 37°10.689′ | 102°03.091′ | 2596 | 西氏熊蜂B. sichelii | 5 | 4 | |
| 37°10.689′ | 102°03.091′ | 2596 | 小雅熊蜂B. lepidus | 9 | 7 | |
| 37°10.689′ | 102°03.091′ | 2596 | 密林熊蜂B. patagiatus | 1 | 0 | |
| 37°10.689′ | 102°03.091′ | 2596 | 兴熊蜂B. impetuosus | 3 | 0 | |
| 37°41.408′ | 100°34.319′ | 3461 | 稳纹熊蜂B. waltoni | 10 | 0 | |
| 37°41.408′ | 100°34.319′ | 3461 | 拉达克熊蜂B. ladakhensis | 3 | 0 | |
| 37°41.408′ | 100°34.319′ | 3461 | 伪猛熊蜂B. personatus | 2 | 0 | |
| 37°41.408′ | 100°34.319′ | 3461 | 红束熊蜂B. rufofasciatus | 1 | 0 | |
| 37°41.408′ | 100°34.319′ | 3461 | 克什米尔熊蜂B. kashmirensis | 1 | 0 | |
| 四川 Sichuan | 28°19.672′ | 103°07.999′ | 2062 | 兴熊蜂B. impetuosus | 80 | 12 |
| 28°19.672′ | 103°07.999′ | 2062 | 小雅熊蜂B. lepidus | 12 | 2 | |
| 28°19.672′ | 103°07.999′ | 2062 | 三条熊蜂B. trifasciatus | 8 | 5 | |
| 28°19.672′ | 103°07.999′ | 2062 | 白背熊蜂B. festivus | 40 | 28 | |
| 28°19.672′ | 103°07.999′ | 2062 | 常熊蜂B. avanus | 10 | 10 | |
| 30°02.905′ | 101°58.049′ | 2833 | 疏熊蜂B. remotus | 159 | 32 | |
| 28°32.822′ | 102°10.638′ | 1785 | 弗里熊蜂B. friseanus | 317 | 86 | |
| 合计Total | 1007 | 262 |
新窗口打开|下载CSV
1.2 熊蜂肠道总DNA提取
将采样保存的熊蜂解剖,取其中肠后放入1.5 mL的离心管中,10 000×g离心2 min,收集离心管下方的沉淀物。向沉淀中依次加入500 mmol·L-1的NaCl 60 μL、100 mmol·L-1的Tris-HCl(pH 8.0)30 μL、50 mmol·L-1的EDTA(pH 8.0)150 μL、10%的SDS溶液30 μL、50 μg·μL-1蛋白酶K溶液30 μL,并混匀,然后在液氮中反复冻融3次,55℃水浴48 h。消化好的悬液按DNA提取试剂盒(Wizard ? SV 96 Genomic DNA Purification System,Promega)的操作步骤提取熊蜂肠道的基因组DNA,于-20℃冰箱保存备用。1.3 引物设计和PCR扩增
利用ITS引物Cri-its-b12及Cri-its-f4扩增熊蜂短膜虫ITS序列[23],上游引物Cri-its-b12序列为5′-AGGAAGCCAAGTCATCCATCGC-3′,下游引物Cri-its-f4序列为5′-GGTCTATGTGATTCCGTGGTTT CC-3′,由华大基因公司合成。PCR反应体系为25 μL,包括5 μL的5×GoTaq Buffer、1.5 μL的MgC12(25 mmol·L-1,Promega公司),2.0 μL的dNTPs(200 μmol·L-1,TaKaRa公司),引物Cri-its-b12和引物Cri-its-f4各1 μL(10 μmol·L-1),0.625 U/管的GoTaq DNA聚合酶,5 μL的模板DNA(约5 ng·μL-1),8.875 μL的ddH2O。反应在Eppendorf Master cycler PCR仪上进行。反应参数:预变性95℃ 4 min,变性95℃ 1 min,退火64℃ 1 min,延伸72℃ 1 min,循环32次,最后延伸72℃ 10 min。之后进行电泳检测条带有无,2%的琼脂糖凝胶,GlodView核酸染料染色,在10 g·L-1的TAE电泳液中电泳,全自动凝胶成像系统成像并拍照留存。1.4 ITS基因片段克隆与测序
1.4.1 PCR产物与载体的连接 利用琼脂糖凝胶回收试剂盒(DP209,天根生化科技有限公司)切胶回收PCR扩增产物,然后再在PCR管中依次加入PCR纯化产物1—3 μL(根据产物浓度而定)和pMD19-T载体1 μL,然后加ddH2O至5 μL,即总量为5 μL,最后加入Solution I溶液5 μL,混匀,16℃反应30 min。1.4.2 连接产物的转化 从-80℃冰箱取出感受态细胞,轻轻放置在冰浴环境中使其融化;缓慢地加入目的DNA,轻轻上下颠倒混匀,冰浴30 min,42℃水浴热激30 s,然后快速转移,冰浴2—3 min,该过程避免离心管剧烈摇晃;然后向离心管中加入500 μL不含氨苄的LB液体培养基,混匀,置于37℃恒温培养箱中,200 r/min培养1 h,待细胞复苏;吸取已转化的感受态细胞加入到含氨苄的LB固体培养基上,涂板,使细胞均匀地分布于平板上,再将平板倒置于37℃培养箱中培养过夜。
1.4.3 菌液PCR 取阳性重组菌落的菌液接种于含氨苄的LB液体培养基中,于恒温培养箱中培养过夜,然后取一定量的菌液进行PCR扩增。反应的总体系为25 μL,反应参数:预变性95℃ 4 min,变性95℃ 1 min,退火64℃ 1 min,延伸72℃ 1 min,循环32次,最后延伸72℃ 10 min。然后进行电泳检测条带有无,2%的琼脂糖凝胶,GlodView染料染色,在10 g·L-1的TAE电泳液中电泳,全自动凝胶成像系统成像并摄像。
1.4.4 重组质粒的抽提及酶切鉴定 经过菌液PCR筛选出来阳性克隆菌株后,采用质粒小提试剂盒(DP103,天根生化科技有限公司)抽提重组质粒,用限制性内切酶HindⅢ和BamHⅠ进行双酶切鉴定。
1.4.5 熊蜂短膜虫ITS序列的测序 将经过菌液PCR方法筛选,以及重组质粒酶切鉴定后的阳性菌落送至北京诺赛基因组研究中心有限公司测序。ITS基因片段的测序结果采用BioEdit编辑,并进行Blast比对分析,用DNAman 9.0对分离的rDNA-ITS序列进行分析比较,MEGA 6.0构建系统发育树以确定短膜虫的系统进化[24,25,26]。
2 结果
2.1 不同种熊蜂的短膜虫感染情况
检测了25种熊蜂的短膜虫感染情况,共计1 007只。检测到感染短膜虫的熊蜂有20种,共计262只,感染率为26.0%,样本量在30只以上的熊蜂有表2所示8种。其中白背熊蜂与火红熊蜂的短膜虫感染率极显著高于其他6种熊蜂(P<0.01),而西伯熊蜂的短膜虫感染率极显著低于其他7种熊蜂(P<0.01)。Table 2
表2
表2不同种熊蜂的短膜虫感染情况
Table 2
| 蜂种 Species | 样本数(只) Sample number | 感染率 Infection rate (%) | 卡方值 χ2 value | P值 P value |
|---|---|---|---|---|
| 白背熊蜂B. festivus | 40 | 70.0A | 36.368 | 1.63E-09 |
| 火红熊蜂B. pyrosoma | 30 | 66.7A | 23.695 | 0.000001 |
| 弗里熊蜂B. friseanus | 317 | 27.1B | 0.057 | 0.812 |
| 疏熊蜂B. remotus | 159 | 20.1BC | 2.931 | 0.087 |
| 密林熊蜂B. patagiatus | 94 | 18.1BC | 3.178 | 0.075 |
| 红束熊蜂B. rufofasciatus | 104 | 14.4C | 7.274 | 0.007 |
| 兴熊蜂B. impetuosus | 85 | 14.1C | 6.323 | 0.012 |
| 西伯熊蜂B. sibiricus | 51 | 2.0D | 15.457 | 0.000084 |
新窗口打开|下载CSV
2.2 同一地区不同种熊蜂的短膜虫感染情况
在调查的熊蜂中,对样品数≥30只的熊蜂短膜虫感染率进行分析比较(表3),四川的弗里熊蜂、疏熊蜂、兴熊蜂的短膜虫感染率依次为27.1%、20.1%和15.0%,且这3种熊蜂感染率无显著性差异(P>0.05);而白背熊蜂的短膜虫感染率为70.0%,极显著高于前3种熊蜂(P<0.01)。甘肃的密林熊蜂和红束熊蜂的短膜虫感染率依次为23.1%和14.6%,且这两种熊蜂感染率差异不显著(P>0.05)。内蒙古的密林熊蜂的短膜虫感染率为12.2%,而西伯熊蜂未发现感染,故密林熊蜂的感染率极显著高于西伯熊蜂(P<0.01)。Table 3
表3
表3同一地区不同种熊蜂的短膜虫感染情况
Table 3
| 蜂种 Species | 四川Sichuan | 甘肃Gansu | 内蒙古Inner Mongolia | |||
|---|---|---|---|---|---|---|
| 样本数(只) Sample number | 感染率 Infection rate (%) | 样本数(只) Sample number | 感染率 Infection rate (%) | 样本数(只) Sample number | 感染率 Infection rate (%) | |
| 白背熊蜂B. festivus | 40 | 70.0A | — | — | — | — |
| 弗里熊蜂B. friseanus | 317 | 27.1B | — | — | — | — |
| 疏熊蜂B. remotus | 159 | 20.1B | — | — | — | — |
| 兴熊蜂B. impetuosus | 80 | 15.0B | — | — | — | — |
| 密林熊蜂B. patagiatus | — | — | 52 | 23.1A | 41 | 12.2A |
| 红束熊蜂B. rufofasciatus | — | — | 103 | 14.6A | — | — |
| 西伯熊蜂B. sibiricus | — | — | — | — | 50 | 0B |
新窗口打开|下载CSV
2.3 同种熊蜂在不同地区的短膜虫感染情况
由表4可以看出,同种熊蜂在不同地区感染的短膜虫感染率之间差异均不显著。密林熊蜂在内蒙古的短膜虫感染率为12.2%,在甘肃为23.1%;火红熊蜂在甘肃的短膜虫感染率为68.4%,在青海为63.6%,两个省份的差异虽不显著,但火红熊蜂的短膜虫感染率在这两个省份均较高;稳纹熊蜂在甘肃和青海两个省份均未发现感染。另外从表1可以看出,跨3个地区分布的明亮熊蜂虽然总的样本量不多,但感染率均较高。Table 4
表4
表4同种熊蜂在不同地区的短膜虫感染情况
Table 4
| 省(自治区)Province (autonomous region) | 密林熊蜂B. patagiatus | 火红熊蜂B. pyrosoma | 稳纹熊蜂B. waltoni | |||
|---|---|---|---|---|---|---|
| 样本数(只) Sample number | 感染率 Infection rate (%) | 样本数(只) Sample number | 感染率 Infection rate (%) | 样本数(只) Sample number | 感染率 Infection rate (%) | |
| 内蒙古Inner Mongolia | 41 | 12.2A | — | — | — | — |
| 甘肃 Gansu | 52 | 23.1A | 19 | 68.4A | 7 | 0A |
| 青海 Qinghai | — | — | 11 | 63.6A | 10 | 0A |
新窗口打开|下载CSV
2.4 不同地区熊蜂的短膜虫感染情况
由表5可以看出,青海、四川、甘肃、内蒙古4个地区的熊蜂短膜虫感染率分别为48.2%、28.0%、22.0%、10.2%。其中青海的短膜虫感染率最高,且极显著高于其他3个地区(P<0.01);内蒙古的短膜虫感染率最低,且极显著低于其他3个省份(P<0.01)。Table 5
表5
表5不同地区熊蜂的短膜虫感染情况
Table 5
| 省(自治区)Province (autonomous region) | 样本数(只) Sample number | 感染率 Infection rate (%) | 卡方值 χ2 value | P值 P value |
|---|---|---|---|---|
| 青海 Qinghai | 56 | 48.2A | 12.668 | 0.000372 |
| 四川 Sichuan | 626 | 28.0B | 0.457 | 0.499 |
| 甘肃 Gansu | 227 | 22.0B | 1.945 | 0.163 |
| 内蒙古Inner Mongolia | 98 | 10.2C | 12.655 | 0.000375 |
新窗口打开|下载CSV
2.5 不同级型熊蜂的短膜虫感染情况
由表6可以看出,雄蜂和工蜂的短膜虫感染率分别为44.7%、24.5%,蜂王未发现感染。其中,雄蜂的感染率极显著高于其他两种级型(P<0.01)。Table 6
表6
表6不同级型熊蜂的短膜虫感染情况
Table 6
| 级型 Caste | 样本数(只)Sample number | 感染率 Infection rate (%) | 卡方值 χ2 value | P值 P value |
|---|---|---|---|---|
| 雄蜂 Male | 76 | 44.7A | 12.467 | 0.000414 |
| 工蜂 Worker | 929 | 24.5B | 0.556 | 0.456 |
| 蜂王 Queen | 2 | 0C | 0.703 | 0.402 |
新窗口打开|下载CSV
2.6 感染熊蜂的短膜虫亲缘关系
基于NJ算法构建不同地区熊蜂感染的短膜虫的亲缘关系(图1)。甘肃地区检测的9种熊蜂中,有7种熊蜂感染的短膜虫亲缘关系较近,而另外2种亲缘关系较远,分别为黑尾熊蜂和猛熊蜂。内蒙古地区检测的3种熊蜂感染的短膜虫亲缘关系均较近。青海地区检测的4种熊蜂短膜虫亲缘关系均较近。四川地区较为复杂,疏熊蜂、兴熊蜂和弗里熊蜂感染的短膜虫种类不一,感染的短膜虫亲缘关系既与整体保持一致性,又有各自特殊的进化分支。总体而言,大部分熊蜂感染的短膜虫亲缘关系均较近,除了甘肃的黑尾熊蜂和猛熊蜂,四川的疏熊蜂、兴熊蜂和弗里熊蜂感染的短膜虫与其他短膜虫亲缘关系较远。此外,还发现甘肃的黑尾熊蜂和四川的弗里熊蜂感染的短膜虫亲缘关系很近,部分采自四川的疏熊蜂感染的短膜虫接近于Crithidia expoeki。图1
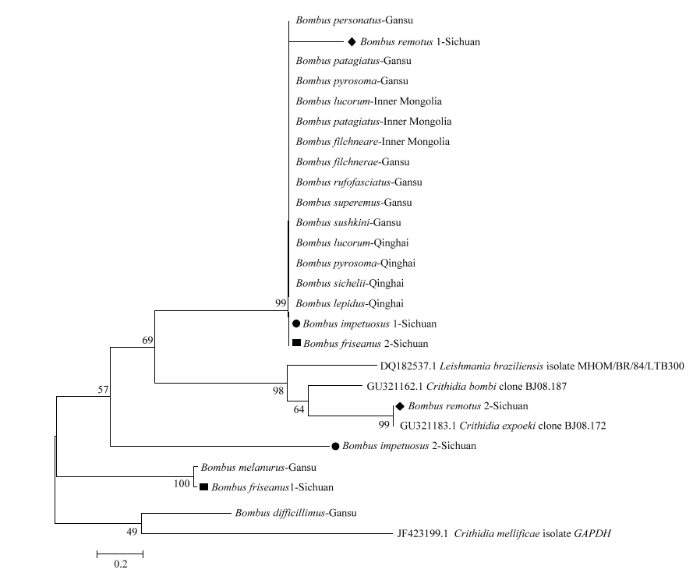 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1基于NJ算法构建的不同地区感染熊蜂的短膜虫近缘种间的系统进化树
Fig. 1Phylogenetic tree of C. bombi of closely related species from different regions based on NJ algorithm
3 讨论
本研究发现,熊蜂短膜虫对我国熊蜂的感染广泛,在检测的内蒙古、甘肃、青海、四川4个省(自治区)的25种1 007只熊蜂中,有20种262只被短膜虫感染,其感染率为26.0%。样本量在30只以上的8种熊蜂分别为白背熊蜂、火红熊蜂、弗里熊蜂、疏熊蜂、密林熊蜂、红束熊蜂、兴熊蜂和西伯熊蜂,其中白背熊蜂和火红熊蜂的短膜虫感染率极显著高于其他6种熊蜂,而西伯熊蜂的短膜虫感染率极显著低于其他7种熊蜂,说明短膜虫存在寄主专一性,这是由于不同种类寄主的生理生化特性和免疫特征不同[27],以及不同基因型的短膜虫与不同基因型的熊蜂种群间相互作用的结果。四川地区的白背熊蜂的雄蜂感染率极高,这导致雄蜂整体发病水平偏高,同时,四川地区的白背熊蜂成为最容易感染短膜虫的蜂种,推测该地区的白背熊蜂为较易感特定种属的短膜虫。另外,青海地区的短膜虫感染率最高,内蒙古地区的短膜虫感染率最低,这可能与不同地区熊蜂的种类有关,例如,青海地区的火红熊蜂、明亮熊蜂、西氏熊蜂的感染率都较高,均为易感染短膜虫的蜂种;而内蒙古地区的西伯熊蜂的短膜虫感染率是样本量在30只以上的所有蜂种中相对最低的,推测正是由于西伯熊蜂占比较大而使得内蒙古地区的整体感染率水平相对其他省份较低。对我国不同地区熊蜂短膜虫感染情况的统计分析侧重于样本量≥30的种类,这些种类是各个地区的优势种。但数量少于30只的样本也可观察到差异,从表1可以看出,跨3个地区分布的明亮熊蜂虽然总的样本量不多,但感染率均很高,推测明亮熊蜂亦为易感病的蜂种,因样本量不足,有待进一步采样检测和验证。根据统计结果也可以看出,同种熊蜂在不同地区的短膜虫感染率差异均不显著,但火红熊蜂在甘肃和青海地区的感染率均远高于其他种熊蜂水平,推测火红熊蜂较易感染短膜虫,这与前述结果一致;而在甘肃和青海两个省份采集的稳纹熊蜂样品中均未发现感染短膜虫,推测稳纹熊蜂为较抗短膜虫的蜂种。
YOURTH等研究发现,在继代移殖短膜虫的蜂群中,短膜虫对熊蜂宿主的感染能力在同一蜂群内没有增加;在未处理的蜂群中,感染短膜虫的工蜂数量下降,推测这种感染情况的减少很可能是由于在同一蜂群中继代移殖短膜虫,会逐渐丧失短膜虫的某些株型,混合型在特定株型的连续传代过程中被“过滤”,而没有相应增加存活株型的整体强度[9]。在意大利和瑞士,短膜虫感染率水平与宿主级型相关,蜂王的感染率为54%—81%,工蜂为75%—80%,雄蜂为47%—71%[28]。我国熊蜂的雄蜂和工蜂的感染率分别为44.7%和24.5%,雄蜂感染率略低于国外水平,而工蜂的感染率远低于国外水平,说明我国熊蜂更抗短膜虫;另外,相比于工蜂,雄蜂的短膜虫感染率更高,说明我国短膜虫更倾向于寄生雄蜂,这为今后防治熊蜂短膜虫提供了理论依据。
通过对四省/区熊蜂所感染的短膜虫的ITS序列进行测序分析发现,内蒙古、甘肃、青海地区熊蜂感染的短膜虫总体进化关系较为一致,同种短膜虫在这几个地区的感染率也相对一致,而四川与其他3个地区虽然有重叠的进化关系,但分化相对较远。熊蜂所感染的短膜虫间的亲缘关系总体来说较强,但也受到地理位置以及其寄主熊蜂蜂种的影响。本研究取样范围仍不够宽泛,我国其他省/区的短膜虫感染情况仍不明确,故有待于进一步扩大调查研究范围。同时,采集样本时,对蜂王的采集较为缺乏,不具充分理由说明蜂王是否抗熊蜂短膜虫。
4 结论
我国熊蜂广泛感染熊蜂短膜虫,不同蜂种、不同地理环境和不同级型间熊蜂短膜虫的感染率均有差异,熊蜂群体经过长期进化演变,不同地区形成优势种类,故不同地区所捕捉到的熊蜂种类不同。熊蜂短膜虫存在寄主专一性,且短膜虫对熊蜂宿主的感染具有适应性,这与不同基因型的短膜虫种群和熊蜂种群间相互作用有关。此外,我国部分地区熊蜂感染的短膜虫的亲缘关系总体来说较强,但也受地理位置及其寄主的影响。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 1]

Bumble bees are among the minority of groups of organisms for which there is some evidence that most species have already been described. Nonetheless, a synoptic revision of the group has been delayed, in part by the difficulties imposed by an unusually high ratio of names to species (averaging more than 11). To explore some of the factors contributing to this phenomenon, historical and geographical trends in the naming of bumble bees are summarised. This shows that most taxa were named by European authors, beginning with the most widespread European species, moving later to not only the more narrowly distributed species and to species from other parts of the world, but also to taxa at progressively lower nomenclatural ranks, particularly within the more widespread European species. Nearly half of all of these names have been published since the last world-wide checklist in 1922. In attempting to bring this up to date, the present checklist adopts broad interpretations of species and recognises a total of 239 recent species (including the social parasites but excluding fossil taxa), with 24 new synonyms and 29 provisional synonyms. The list also includes notes on alternative interpretations of taxonomic status and on nomenclatural problems, drawing attention to those cases where further research is most urgently needed. In particular, suggestions are presented for an application to the International Commission on Zoological Nomenclature to use its Plenary Power in order to conserve current usage of the commonly used names atratus, balteatus, distinguendus, flavifrons, humilis, hyperboreus, mesomelas, mixtus, norvegicus, polaris, pyrenaeus, soroeensis and variabilis.
DOI:10.1017/S1477200008002843URL [本文引用: 1]
Bumblebees are important pollinators in mountainous regions. The highland region of Sichuan and Chongqing (together, Sichuan in the former broad sense) includes part of the greatest hotspot of bumblebee diversity worldwide, with half of the species of China and more than a fifth of the world''s species. In this paper we present the first review of this diverse but particularly poorly known fauna, drawing on 6705 bees from selective samples made by the authors during field work and from museum collections, together with 1123 literature records (an overlapping set). Among this material we recognise 56 species that have 847 names (including infrasubspecific names). One new species, Bombus (Pyrobombus) wangae, is described from Sichuan and Gansu. B. laesus is found to have a colour pattern in this part of its range that was previously undescribed. Six species are recorded from the Sichuan090006Chongqing region for the first time (B. avanus, B. branickii, B. difficillimus B. humilis, B. norvegicus, B. tibetanus), of which B. avanus is only the second published record worldwide. One species, B. braccatus, is endemic to Sichuan and is confirmed as extant in 2005. We provide diagnoses and keys to species for both sexes. Colour variation is described, distributions within the Sichuan090006Chongqing region are mapped, altitudinal and seasonal activity are plotted, and the authors090005 records of food plants are listed.
DOI:10.1111/j.1095-8312.2007.00784.xURL [本文引用: 1]
Bumble bees ( Bombus Latreille) occupy a wide diversity of habitats, from alpine meadows to lowland tropical forest, yet they appear to be similar in morphology throughout their range, suggesting that behavioural adaptations play a more important role in colonizing diverse habitats. Notwithstanding their structural homogeneity, bumble bees exhibit striking inter- and intraspecific variation in colour pattern, purportedly the outcome of mimetic evolution. A robust phylogeny of Bombus would provide the framework for elucidating the history of their wide biogeographical distribution and the evolution of behavioural and morphological adaptations, including colour pattern. However, morphological studies of bumble bees have discovered too few phylogenetically informative characters to reconstruct a robust phylogeny. Using DNA sequence data, we report the first nearly complete species phylogeny of bumble bees, including most of the 250 known species from the 38 currently recognized subgenera. Bayesian analysis of nuclear (opsin, EF-102±, arginine kinase, PEPCK) and mitochondrial (16S) sequences results in a highly resolved and strongly supported phylogeny from base to tips, with clear-cut support for monophyly of most of the conventional morphology-based subgenera. Most subgenera fall into two distinct clades ( short-faced and long-faced ) associated broadly with differences in head morphology. Within the short-faced clade is a diverse New World clade, which includes nearly one-quarter of the currently recognized subgenera, many of which are restricted to higher elevations of Central and South America. The comprehensive phylogeny provides a firm foundation for reclassification and for evaluating character evolution in the bumble bees. 0008 2007 The Linnean Society of London, Biological Journal of the Linnean Society , 2007, 91 , 161090009188.
DOI:10.17520/biods.2018068URL [本文引用: 1]
熊蜂是众多野生植物和农作物的重要传粉者,在维持自然和农业生态系统平衡中发挥着十分重要的作用。中国地形多样、植被丰富,是全球熊蜂多样性的热点地区,但过去缺乏系统调查研究,中国熊蜂多样性现状及其利用特性不详。本文介绍了近20年在中国熊蜂资源系统调查及其人工利用方面取得的主要研究结果:(1)2002–2017年采集熊蜂标本5万余号,先后揭示了一系列疑难类群的分类地位,确认中国已知熊蜂125种,占全球熊蜂已知物种总数的50%,表明中国是全球熊蜂物种资源最丰富的国家;(2)首次报道了中国熊蜂物种名录,其中22种为中国特有种。我国青藏高原东部向黄土高原、秦岭山地和四川盆地过渡的地带,是全球熊蜂多样性分布中心;(3)1998–2017年,明亮熊蜂(Bombus lucorum)、密林熊蜂(B.patagiatus)、红光熊蜂(B.ignitus)、火红熊蜂(B.pyrosoma)、重黄熊蜂(B.picipes)和兰州熊蜂(B.lantschouensis)6种本土熊蜂先后被驯养成功,其中密林熊蜂和兰州熊蜂的繁殖性状优良,已应用于我国设施作物传粉服务。同时,我们从栖息地、食物资源、外来物种、农药使用等方面提出了保护中国熊蜂资源的策略,以期为野生传粉昆虫特别是熊蜂资源的保护与可持续利用提供参考资料。
DOI:10.17520/biods.2018068URL [本文引用: 1]
熊蜂是众多野生植物和农作物的重要传粉者,在维持自然和农业生态系统平衡中发挥着十分重要的作用。中国地形多样、植被丰富,是全球熊蜂多样性的热点地区,但过去缺乏系统调查研究,中国熊蜂多样性现状及其利用特性不详。本文介绍了近20年在中国熊蜂资源系统调查及其人工利用方面取得的主要研究结果:(1)2002–2017年采集熊蜂标本5万余号,先后揭示了一系列疑难类群的分类地位,确认中国已知熊蜂125种,占全球熊蜂已知物种总数的50%,表明中国是全球熊蜂物种资源最丰富的国家;(2)首次报道了中国熊蜂物种名录,其中22种为中国特有种。我国青藏高原东部向黄土高原、秦岭山地和四川盆地过渡的地带,是全球熊蜂多样性分布中心;(3)1998–2017年,明亮熊蜂(Bombus lucorum)、密林熊蜂(B.patagiatus)、红光熊蜂(B.ignitus)、火红熊蜂(B.pyrosoma)、重黄熊蜂(B.picipes)和兰州熊蜂(B.lantschouensis)6种本土熊蜂先后被驯养成功,其中密林熊蜂和兰州熊蜂的繁殖性状优良,已应用于我国设施作物传粉服务。同时,我们从栖息地、食物资源、外来物种、农药使用等方面提出了保护中国熊蜂资源的策略,以期为野生传粉昆虫特别是熊蜂资源的保护与可持续利用提供参考资料。
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
蜜蜂科(Apidae)是膜翅目(Hymenoptera)细腰亚目(Apocrita)针尾部(ACULEATA)蜜蜂总科中的一个科,一个很大的科,全世界共有25~30万种,分属于8亚科4000多属。象许多其它的膜翅目成虫一样,蜜蜂科的雌性成虫都访花以吸收花蜜并采集花粉。但...
.
URL [本文引用: 1]
蜜蜂科(Apidae)是膜翅目(Hymenoptera)细腰亚目(Apocrita)针尾部(ACULEATA)蜜蜂总科中的一个科,一个很大的科,全世界共有25~30万种,分属于8亚科4000多属。象许多其它的膜翅目成虫一样,蜜蜂科的雌性成虫都访花以吸收花蜜并采集花粉。但...
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 2]
DOI:10.1038/2121385a0URL [本文引用: 1]
KNOWLEDGE of the structure and life cycles of the medically important Haemoflagellates of man and lower animals has increased during this century and it became necessary to define the developmental stages of the leishmanias and trypanosomes in their mammalian and insect hosts. In the course of their cyclical development, flagellates of the genera Leishmania and Trypanosoma pass through stages comparable with those of the monogenetic Trypanosomatidae and so it has been customary to refer to them by names derived from those genera in which the corresponding stages are the most characteristic forms.
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
利用等位基因酶电泳,对直接从宿主体内分离和克隆的40个熊蜂短股虫克隆进行了遗传变异检测。这40个熊蜂短股虫克隆分别从被两个蜂王感染的8只工蜂体内得到。共检测了GPI、MDH、ME、6PGD和PGM等五种酶。根据PGM,发现这40个熊蜂短股虫克隆中具有两种变异型。结果说明自然界中,熊蜂短膜虫存在着多重感染的现象。利用脉冲场梯度凝胶电泳,对熊蜂短膜央两种变异型的染色体规模DNA进行了比较,没有发现染色体规模DNA的区别。结合等位基因酶和染色体规模DNA电泳的结果,说明两种变异型的亲缘关系密切。对熊蜂短膜虫两种变异型生长曲线的比较结果,发现它们的增殖速度具有差别。
URLMagsci [本文引用: 1]
利用等位基因酶电泳,对直接从宿主体内分离和克隆的40个熊蜂短股虫克隆进行了遗传变异检测。这40个熊蜂短股虫克隆分别从被两个蜂王感染的8只工蜂体内得到。共检测了GPI、MDH、ME、6PGD和PGM等五种酶。根据PGM,发现这40个熊蜂短股虫克隆中具有两种变异型。结果说明自然界中,熊蜂短膜虫存在着多重感染的现象。利用脉冲场梯度凝胶电泳,对熊蜂短膜央两种变异型的染色体规模DNA进行了比较,没有发现染色体规模DNA的区别。结合等位基因酶和染色体规模DNA电泳的结果,说明两种变异型的亲缘关系密切。对熊蜂短膜虫两种变异型生长曲线的比较结果,发现它们的增殖速度具有差别。
DOI:10.3969/j.issn.1674-5507.1998.02.019URL [本文引用: 1]
Genetic relationship of Crithidia bombi and their host bumble bees, Bombus terrestris, were studied by the survival of bees from different colonies infected with different strains of C. bombi. The results will add to our limited knowledge of natural animal-parasite systems and provide comparative data for the exploration of the genetic relationships between parasites and their hosts.
DOI:10.3969/j.issn.1674-5507.1998.02.019URL [本文引用: 1]
Genetic relationship of Crithidia bombi and their host bumble bees, Bombus terrestris, were studied by the survival of bees from different colonies infected with different strains of C. bombi. The results will add to our limited knowledge of natural animal-parasite systems and provide comparative data for the exploration of the genetic relationships between parasites and their hosts.
DOI:10.1086/417338URL [本文引用: 1]
DOI:10.3969/j.issn.1673-758X.2002.05.021 [本文引用: 1]
篇首:吸虫分类是一个复杂而又有争议性的话题,长期以来,****们对吸虫分类进行了各种各样的研究。传统的 吸虫分类主要依据虫体形态、中间宿主地理分布、交叉繁殖及生理生化等参数,后来应用了蛋白质和酶学方法。随着分子生物学的技术发展,应用DNA技术研究种 株的基因差异,分析种株的遗传变异及分类被认为是一种直接而可靠的方法。本文就近10年来吸虫种株在核糖体RNA、线粒体DNA、随机扩增的多态性DNA 分析等方面的研究进展作一综述。
DOI:10.3969/j.issn.1673-758X.2002.05.021 [本文引用: 1]
篇首:吸虫分类是一个复杂而又有争议性的话题,长期以来,****们对吸虫分类进行了各种各样的研究。传统的 吸虫分类主要依据虫体形态、中间宿主地理分布、交叉繁殖及生理生化等参数,后来应用了蛋白质和酶学方法。随着分子生物学的技术发展,应用DNA技术研究种 株的基因差异,分析种株的遗传变异及分类被认为是一种直接而可靠的方法。本文就近10年来吸虫种株在核糖体RNA、线粒体DNA、随机扩增的多态性DNA 分析等方面的研究进展作一综述。
DOI:10.3321/j.issn:0366-6964.2005.01.015URLMagsci [本文引用: 1]
通过对国内来源于不同宿主的ZS人株、SH人株、CN猪株、QH绵羊株4个弓形虫虫株,以及国际标准强毒株RH株的核糖体DNA内转录间隔区(ITS)及5.8S DNA序列进行PCR扩增、克隆、测序和序列分析,旨在对国内不同宿主间弓形虫虫株的遗传变异情况进行分析和验证,为分子遗传学和分子诊断学研究提供资料。结果显示:QH绵羊株、ZS人株、SH人株、CN猪株的ITS及5.8S序列完全一致,且与GenBank上注册RH株的ITS及5.8S序列也一致;仅实验室传代保存的RH株的ITS2序列与其它4株的ITS2有2个碱基的差异。结果表明ITS可作为分子标记用于弓形虫与其它原虫的种间鉴定,但不适合用于弓形虫种内遗传变异的研究。
DOI:10.3321/j.issn:0366-6964.2005.01.015URLMagsci [本文引用: 1]
通过对国内来源于不同宿主的ZS人株、SH人株、CN猪株、QH绵羊株4个弓形虫虫株,以及国际标准强毒株RH株的核糖体DNA内转录间隔区(ITS)及5.8S DNA序列进行PCR扩增、克隆、测序和序列分析,旨在对国内不同宿主间弓形虫虫株的遗传变异情况进行分析和验证,为分子遗传学和分子诊断学研究提供资料。结果显示:QH绵羊株、ZS人株、SH人株、CN猪株的ITS及5.8S序列完全一致,且与GenBank上注册RH株的ITS及5.8S序列也一致;仅实验室传代保存的RH株的ITS2序列与其它4株的ITS2有2个碱基的差异。结果表明ITS可作为分子标记用于弓形虫与其它原虫的种间鉴定,但不适合用于弓形虫种内遗传变异的研究。
DOI:10.1007/PL00008561URLPMID:11002982 [本文引用: 1]
Abstract The nuclear ribosomal DNA (rDNA) region spanning the first (ITS-1) and second (ITS-2) internal transcribed spacers was sequenced for 15 taxa of ascaridoid nematodes. The length of the ITS-1 and ITS-2 sequences in the 15 taxa ranged from 392-500 bp and 240 348 bp, respectively. While nucleotide variation of 0-2.9% in the ITS-1 and/or ITS-2 sequences was detected within taxa where multiple samples were sequenced, significantly higher level of nucleotide difference (9.4-66.6%) was detected between the taxa, except for Ascaris suum and A. lumbricoides whose taxonomic status remains uncertain. These interspecific differences were linked with the considerable size differences (0-108 bp) in the rDNA spacers. Phenograms based on the genetic differences among the 15 taxa showed some concordance with previous classification schemes derived from morphological data.
DOI:10.3321/j.issn:0412-0914.2001.01.013URL [本文引用: 1]
本文为克服形态鉴定的不足,用分子生物学的方法鉴别松材线虫。线虫核糖体DNA(rDNA) 的内部转录间隔区(ITS1)区(约308 bp)的测序结果显示:松材线虫种内区别很小,不超过1 bp;拟松材线虫种内区别较大,最大达7 bp;这2种线虫的种间区别为32~39 bp。根据以上测序结果,本文结合单条线虫DNA的提取技术,对14个松材线虫和拟松材线虫样本进行了单链构象多态性(PCR—SSCP)分析,结果表明 PCR-SSCP分析技术可明确区分这2种线虫,该技术可为单条松材线虫的鉴定提供一套灵敏而可靠的方法。
DOI:10.3321/j.issn:0412-0914.2001.01.013URL [本文引用: 1]
本文为克服形态鉴定的不足,用分子生物学的方法鉴别松材线虫。线虫核糖体DNA(rDNA) 的内部转录间隔区(ITS1)区(约308 bp)的测序结果显示:松材线虫种内区别很小,不超过1 bp;拟松材线虫种内区别较大,最大达7 bp;这2种线虫的种间区别为32~39 bp。根据以上测序结果,本文结合单条线虫DNA的提取技术,对14个松材线虫和拟松材线虫样本进行了单链构象多态性(PCR—SSCP)分析,结果表明 PCR-SSCP分析技术可明确区分这2种线虫,该技术可为单条松材线虫的鉴定提供一套灵敏而可靠的方法。
DOI:10.3969/j.issn.1008-0589.2005.05.014URL [本文引用: 1]
通过对国内三株安氏隐孢子虫(Cryptosporidium andersoni)即GD株、HN株和AH株的rDNA的内转录间隔区Ⅰ(ITS_1)序列进行PCR扩增、克隆、测序和序列分析,旨在确定ITS_1是否可作为C.andersoni分子分类的遗传标记。结果表明:GD株、HN株和AH株的ITS_1序列基本一致,仅AH株有三个碱基的差异;但与GenBank注册的C.muris和C.parvum存在种间差异,而且差异显著。说明ITS_1可作为C.andersoni种的遗传标记,从而为隐孢子虫属的种间鉴定以及进一步的分子流行病学调查和分子诊断学研究奠定了基础。
DOI:10.3969/j.issn.1008-0589.2005.05.014URL [本文引用: 1]
通过对国内三株安氏隐孢子虫(Cryptosporidium andersoni)即GD株、HN株和AH株的rDNA的内转录间隔区Ⅰ(ITS_1)序列进行PCR扩增、克隆、测序和序列分析,旨在确定ITS_1是否可作为C.andersoni分子分类的遗传标记。结果表明:GD株、HN株和AH株的ITS_1序列基本一致,仅AH株有三个碱基的差异;但与GenBank注册的C.muris和C.parvum存在种间差异,而且差异显著。说明ITS_1可作为C.andersoni种的遗传标记,从而为隐孢子虫属的种间鉴定以及进一步的分子流行病学调查和分子诊断学研究奠定了基础。
DOI:10.3864/j.issn.0578-1752.2011.23.023URLMagsci [本文引用: 1]
【目的】通过分析比较钝绥螨ITS基因片段,探讨其作为分子手段应用于种类鉴定,同时应用此分子标记来分析钝绥螨的亲缘关系,为研究其系统发育和完善分类系统提供参考。【方法】对采自江西桔园的尼氏真绥螨、江原钝绥螨、津川钝绥螨和东方钝绥螨进行总DNA提取,PCR扩增,测序,并从GenBank中下载5种钝绥螨的ITS序列,应用软件分析ITS片段长度、A+T%含量、变异位点和进化关系。【结果】得到总长度为644—655 bp的ITS序列片段,序列中A+T碱基比例较高,为58.2%。整个ITS基因片段存在204个碱基变异位点(ITS1:172;ITS2:22;5.8S:10)和32个碱基插入/缺失。真绥螨属的尼氏真绥螨和卵圆真绥螨的分类地位较近,遗传距离为0.013;而小新绥螨属的4个种与钝绥螨属的3个种在构建系统发育树时未能明显地分为2支,其中胡瓜新小绥螨和东方钝绥螨的遗传距离最近,为0.083。【结论】ITS序列片段分析结果支持尼氏真绥螨和卵圆真绥螨现在的分类地位,而小新绥螨属与钝绥螨属似乎未达到属间差异,其分类地位有待进一步确定。
DOI:10.3864/j.issn.0578-1752.2011.23.023URLMagsci [本文引用: 1]
【目的】通过分析比较钝绥螨ITS基因片段,探讨其作为分子手段应用于种类鉴定,同时应用此分子标记来分析钝绥螨的亲缘关系,为研究其系统发育和完善分类系统提供参考。【方法】对采自江西桔园的尼氏真绥螨、江原钝绥螨、津川钝绥螨和东方钝绥螨进行总DNA提取,PCR扩增,测序,并从GenBank中下载5种钝绥螨的ITS序列,应用软件分析ITS片段长度、A+T%含量、变异位点和进化关系。【结果】得到总长度为644—655 bp的ITS序列片段,序列中A+T碱基比例较高,为58.2%。整个ITS基因片段存在204个碱基变异位点(ITS1:172;ITS2:22;5.8S:10)和32个碱基插入/缺失。真绥螨属的尼氏真绥螨和卵圆真绥螨的分类地位较近,遗传距离为0.013;而小新绥螨属的4个种与钝绥螨属的3个种在构建系统发育树时未能明显地分为2支,其中胡瓜新小绥螨和东方钝绥螨的遗传距离最近,为0.083。【结论】ITS序列片段分析结果支持尼氏真绥螨和卵圆真绥螨现在的分类地位,而小新绥螨属与钝绥螨属似乎未达到属间差异,其分类地位有待进一步确定。
DOI:10.3969/j.issn.1672-3619.2007.03.005URL [本文引用: 1]
目的扩增绒山羊和绵羊源东毕吸虫的ITS rDNA序列并进行分析比较。方法运用PCR方法,以保守引物BD1和BD2扩增从黑龙江绒山羊和绵羊体内分离的东毕吸虫(Oriemobilharzia spp.)rDNA的内转录间隔区(ITS-1及ITS-2)。PCR产物经测序后进行序列分析。结果黑龙江绒山羊和绵羊东毕吸虫的ITS序列总长均为875bp,其中ITS-1序列长为384Up,5.8S序列长为159Up,ITS-2序列长为332Up。黑龙江绒山羊和绵羊东毕吸虫的ITS序列相似性为99.9%,只有一个碱基差异,存在于ITS-2序列中。结论本研究在国际上首次报道了绒山羊和绵羊东毕吸虫的ITS序列,证实绒山羊和绵羊源东毕吸虫代表同一个种。这些结果为东毕吸虫分子生物学的进一步研究奠定了基础,并为寄生虫分子系统学研究提供了新资料。
DOI:10.3969/j.issn.1672-3619.2007.03.005URL [本文引用: 1]
目的扩增绒山羊和绵羊源东毕吸虫的ITS rDNA序列并进行分析比较。方法运用PCR方法,以保守引物BD1和BD2扩增从黑龙江绒山羊和绵羊体内分离的东毕吸虫(Oriemobilharzia spp.)rDNA的内转录间隔区(ITS-1及ITS-2)。PCR产物经测序后进行序列分析。结果黑龙江绒山羊和绵羊东毕吸虫的ITS序列总长均为875bp,其中ITS-1序列长为384Up,5.8S序列长为159Up,ITS-2序列长为332Up。黑龙江绒山羊和绵羊东毕吸虫的ITS序列相似性为99.9%,只有一个碱基差异,存在于ITS-2序列中。结论本研究在国际上首次报道了绒山羊和绵羊东毕吸虫的ITS序列,证实绒山羊和绵羊源东毕吸虫代表同一个种。这些结果为东毕吸虫分子生物学的进一步研究奠定了基础,并为寄生虫分子系统学研究提供了新资料。
[本文引用: 1]
[P]. (
[本文引用: 1]
[P]. (
[本文引用: 1]
DOI:10.1016/j.ympev.2011.08.031URLPMID:21930222 [本文引用: 1]
Pinnularia is an ecologically important and species-rich genus of freshwater diatoms (Bacillariophyceae) showing considerable variation in frustule morphology. Interspecific evolutionary relationships were inferred for 36 Pinnularia taxa using a five-locus dataset. A range of fossil taxa, including newly discovered Middle Eocene forms of Pinnularia, was used to calibrate a relaxed molecular clock analysis and investigate temporal aspects of the genus’ diversification. The multi-gene approach resulted in a well-resolved phylogeny of three major clades and several subclades that were frequently, but not universally, delimited by valve morphology. The genus Caloneis was not recovered as monophyletic, confirming that, as currently delimited, this genus is not evolutionarily meaningful and should be merged with Pinnularia. The Pinnularia–Caloneis complex is estimated to have diverged between the Upper Cretaceous and the early Eocene, implying a ghost range of at least 10million year (Ma) in the fossil record.
DOI:10.1016/S0022-2836(05)80360-2URL [本文引用: 1]
ABSTRACT A new approach to rapid sequence comparison, basic local alignment search tool (BLAST), directly approximates alignments that optimize a measure of local similarity, the maximal segment pair (MSP) score. Recent mathematical results on the stochastic properties of MSP scores allow an analysis of the performance of this method as well as the statistical significance of alignments it generates. The basic algorithm is simple and robust; it can be implemented in a number of ways and applied in a variety of contexts including straightforward DNA and protein sequence database searches, motif searches, gene identification searches, and in the analysis of multiple regions of similarity in long DNA sequences. In addition to its flexibility and tractability to mathematical analysis, BLAST is an order of magnitude faster than existing sequence comparison tools of comparable sensitivity.
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}