 ,中国农业科学院北京畜牧兽医研究所,北京100193
,中国农业科学院北京畜牧兽医研究所,北京100193Cloning Expression Analysis and Transformation of MsGAI Gene from Medicago sativa L
ZHANG Han, WANG XueMin, LIU XiQiang, MA Lin, WEN HongYu, WANG Zan,Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing, 100193通讯作者:
责任编辑: 李莉
收稿日期:2018-07-18接受日期:2018-09-7网络出版日期:2019-01-16
| 基金资助: |
Received:2018-07-18Accepted:2018-09-7Online:2019-01-16
作者简介 About authors
张涵,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (8066KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张涵, 王学敏, 刘希强, 马琳, 温红雨, 王赞. 紫花苜蓿MsGAI的克隆、表达及遗传转化[J]. 中国农业科学, 2019, 52(2): 201-214 doi:10.3864/j.issn.0578-1752.2019.02.002
ZHANG Han, WANG XueMin, LIU XiQiang, MA Lin, WEN HongYu, WANG Zan.
0 引言
【研究意义】紫花苜蓿(Medicago sativa L.)是一种多年生优质豆科牧草,具有抗逆性强、适应性广、产量高、适口性良好、营养物质丰富等优点,享有“牧草之王”的美誉[1]。此外,紫花苜蓿根系发达,不仅具有固氮作用[2],还可以改善土壤结构、促进微生物活性和营养循环[3]。2011—2015年中国苜蓿干草年平均产量为7.1×108 kg,消费量年平均1.42×109 kg(数据来源:http://m.askci.com/),供需不平衡的严峻形势下,提高中国紫花苜蓿的产量已亟不可待。【前人研究进展】赤霉素(gibberllin,GAs)属于四环二萜生长因子,是种子萌发、胚轴伸长、茎的伸长、以及花的形成等植物发育过程中的基本调节剂[4]。在提高植物生物量方面发挥重要作用,DELLA蛋白是赤霉素信号转导途径中关键的负调节因子,参与GA信号传导并在其中发挥负调控的作用;赤霉素通过泛素蛋白酶降解的途径,来解除DELLA蛋白对其转导途径中的抑制作用。植物体内GA含量升高时,与赤霉素受体(Gibberllin Insensitive Dwarf1)GID结合形成一个二聚体,再与DELLA蛋白形成一个三元复合结构,从而引起DELLA构象发生变化,使SLY1元件能与GRAS域结合,被F-BOX蛋白识别,导致DELLA蛋白通过泛素-蛋白酶途径快速降解,从而使GA发挥正常效应[5]。DELLA蛋白的这种抑制作用在农业生产中起到有利作用,20世纪60—70年代育种学家选育出新的谷物品种,该品种共同特点为植株低矮且高产,比其他品种更具抗倒伏的作用,还具有耐风沙的重要特点[6,7]。这些新品种都是赤霉素合成或应答相关基因发生突变导致[8,9,10]。DELLA蛋白属于转录调控因子,具有保守的C端GRAS结构域,同时含有聚合Ser、Thr区域[11,12,13]和VHIID、SH2、SAW阻遏结构域[14]。在DELLA蛋白结构的中部,出现DELLA和TVHYNP 2个保守的酸性结构域和核定位信号NLS[15];当在转基因植物中表达时,DELLA-GFP融合蛋白定位在细胞核中[16]。拟南芥(Arabidopsis thaliana)中的5个DELLA蛋白存在功能冗余现象,其中,RGA与GAI对植物的营养生长和开花时间有明显影响[17];而RGL1与RGL2在种子萌发过程中发挥重要作用[18]。小麦(Triticum aestivum)Rht-B1/Rht-D1和玉米(Zea mays)dwarf-8(d8)是拟南芥赤霉素不敏感GAI的直系同源基因[19]。这些基因编码类似核转录因子和蛋白质,含有类似于SH2结构域,表明磷酸酪氨酸可能参与赤霉素信号传导。含有GAI等位基因的水稻(Oryza sativa)植株对赤霉素的反应降低并且出现矮化的现象,表明突变的GAI同源物可广泛增加作物产量[20]。GAI突变会影响GA接收以及随后的信号转导,但不会引起GA缺乏。先前对玉米d8和小麦Rht突变体的生理和遗传分析表明,这些突变都属于功能获得性突变[21],降低对GA的反应同时增加内源GA水平[22]。FU等[23]和DILL等[24]指出GAI过表达会导致植株低矮以及GA含量降低,并且这种效应的强度与转基因表达的水平相关。FOSTER等[25]克隆了苹果(Malus domestica)DELLA蛋白的6个基因,将MdRGL2a转入拟南芥,转基因植株出现开花延迟、节间变短等特征。【本研究切入点】目前,关于DELLA蛋白家族的研究多集中在作物及一些模式植物中,牧草中少有报道,且对DELLA蛋白在植物组织中表达模式和胁迫条件下的表达变化探索较少。紫花苜蓿作为牧草中的典型代表,DELLA蛋白在紫花苜蓿中的分子机理尚不明确。【拟解决的关键问题】本研究利用同源克隆的方法得到紫花苜蓿MsGAI,并对该基因编码蛋白的理化性质、组织表达特异性、不同处理下的表达模式进行分析,同时构建了MsGAI植物超表达载体,并获得了转基因紫花苜蓿材料,以期为解析GAI调控紫花苜蓿生物量的机理,进而开展分子遗传改良奠定基础。1 材料与方法
1.1 试验材料
1.1.1 植物材料 试验于2016年7月至2017年12月进行,试验材料为紫花苜蓿‘中苜1号’(Medicago sativa L. Zhongmu No.1)品种,由中国农业科学院北京畜牧兽医研究所保存。1.1.2 试验试剂 PROMEGA总RNA提取试剂盒,
购自普洛麦格公司,TransScript Green One-Step qRT-PCR SuperMix、pEASY-Blunt Simple Cloning Kit、Trans5α Chemically Competent Cell、EasyPure Quick Gel Extraction Kit、DNA凝胶试剂盒,购自北京全式金生物技术有限公司。试验所用的内切酶、连接酶购自NEB(北京)有限公司。DNA提取试剂盒,购自天根生化科技(北京)有限公司。
1.2 紫花苜蓿MsGAI的克隆
使用Eastep Super提取紫花苜蓿总RNA;利用TransScript Green One-Step qRT-PCR反转录体系合成cDNA。根据蒺藜苜蓿(Medicago truncatula)MtGAI(GenBank登录号XM_013605137.1)的cDNA序列,设计特异引物MsGAI_F/R(表1),以紫花苜蓿cDNA为模板进行PCR扩增,反应体系为25 μL:cDNA 1 μL、KOD-plus 1 μL、10×Buffer for KOD 2.5 μL、25 mmol·L-1 MgSO4 1 μL、2 mmol·L-1 dNTPs 2.5 μL、MsGAI引物各0.75 μL和PCR grade water 15.5 μL。反应程序为94℃ 5 min;94℃ 30 s,60℃ 30 s,72℃ 2 min,34个循环;72℃ 5 min。产物经1%琼脂糖凝胶电泳检测、纯化,并与Peasy-T1-Blunt载体连接,转化,挑取单克隆进行菌液PCR验证,阳性克隆送上海英潍捷基生物技术有限公司测序。
Table 1
表1
表1试验中所用引物序列
Table 1
| 引物名称 Primers | 序列 Sequence of primers (5′-3′) | 用途 Application |
|---|---|---|
| MsGAI | F: AAACTTCAACCCATAAACTC | 基因克隆 Gene cloning |
| R: ACTTAAGGGTACCCTGAG | ||
| 121-MsGAI | F: TGCTCTAGAATGAAGAGAGAACACCA | pBI121载体构建 Vector construction |
| R: CGCGGATCCTCACTTGGACTCATTTTG | ||
| Ms_Actin | F: CAAAAGATGGCAGATGCTGAGGAT | 内参基因 Internal control |
| R: CATGACACCAGTATGACGAGGTCG | ||
| QGAI | F: CCACCACCTTAACAGCAGCA | 荧光定量 Real-time PCR |
| R: GAGCACTACCCATAACCATCTC | ||
| M13 | F: GTAAAACGACGGCCAGT | 亚克隆引物 The primers for subcloning |
| R: CAGGAAACAGCTATGAC | ||
| 35s | F: GGTGGCTCCTACAAATGCCA | pBI121载体构建 Vector construction |
| R: GAAACGCAGCACGATACGC | ||
| Promoter | F: GGTACACGCTAAGACGCTAC | 启动子克隆 Promoter cloning |
| R: TTGCTGCTGTTAAGGTGG |
新窗口打开|下载CSV
1.3 MsGAI的生物信息学分析
利用SnapGene软件分析MsGAI开放阅读框及其氨基酸序列;通过Protparam(http://strbio.sourceforge. net/javadocs/org/strbio/net/ProtParam.html)分析MsGAI的理化性质;利用TMHMM(http://WWW.cbs.dtu.dk/ services/TMHMM)对MsGAI蛋白进行跨膜结构的预测分析;利用SOPMA(http://www.sopma.org/)在线预测该蛋白的二级结构;利用SingnalP(http://WWW. cbs.dtu.dk/services/SingnalP-4.1)分析MsGAI信号肽;利用TargetP(http://WWW.cbs.dtu.dk/services/TargetP)程序预测MsGAI蛋白的亚细胞定位。使用ClustalX软件与其他物种进行多序列比对;利用MEGA7.0软件构建分子进化树,同时利用Neighbor-Joining(NJ)法构建系统发育树,Bootstrap值设定为500,通过Bootstrap对生成的系统进化树进行校正;利用SWISS-MODEL(https://swissmodel. expasy.org/)在线预测MsGAI的三级结构;利用DNAMAN软件进行氨基酸同源序列比对;利用PlantCARE(http://www.plantcarescience.com/)预测分析MsGAI启动子。
1.4 MsGAI启动子的克隆和分析
为了深入分析MsGAI的表达,将克隆得到的MsGAI序列进行比对(https://www.alfalfatoolbox.org/ doblast/),挑选得分最高的基因组序列,选择其起始密码子前2 000 bp左右的基因序列,设计特异性引物Promoter(表1),以‘中苜1号’DNA为模板,进行PCR扩增。利用PlantCARE,对得到的启动子序列进行分析。1.5 MsGAI的组织特异性表达分析
提取‘中苜1号’根、茎、叶、花瓣、花萼、雄蕊和雌蕊的总RNA,反转录成cDNA后,进行实时荧光定量PCR。根据得到的MsGAI序列设计特异引物QGAI(表1),以紫花苜蓿Actin为内参基因,利用ABI7500 Real-Time PCR system(美国ABI公司),依照TaKaRa SYBY Premix Ex Taq说明书进行PCR扩增。反应程序为95℃ 5 min;95℃ 5 s,58℃ 35 s,共40个循环。3次生物学重复,2次技术重复。采用2-??CT方法[26]计算MsGAI的表达量。1.6 不同处理下MsGAI的表达分析
‘中苜1号’种子经氯气消毒,平铺在有滤纸的培养皿中,用无菌水将滤纸浸湿,在光照培养箱(16 h光照,8 h黑暗)中培养至种子露白,发芽后的种子移至MS培养基上,培养4周。分别用0.3 mol·L-1 NaCl、15%的PEG、0.1x10-3 mol·L-1 GA3和ABA处理0、2、4、8、12和24 h;黑暗处理0、12、24、48和72 h。1.7 MsGAI超表达载体的构建
根据MsGAI的ORF序列设计带有BamHⅠ和XbaⅠ双酶切位点的上下游引物121-MsGAI(表1)。以含有MsGAI的T载体质粒为模板,扩增含有酶切位点的目的片段,用XbaⅠ/BamHⅠ双酶切目的片段和pBI121空载体,连接、转化。筛选阳性克隆送上海英潍捷基生物技术有限公司测序。超表达载体构建示意图如图1。图1
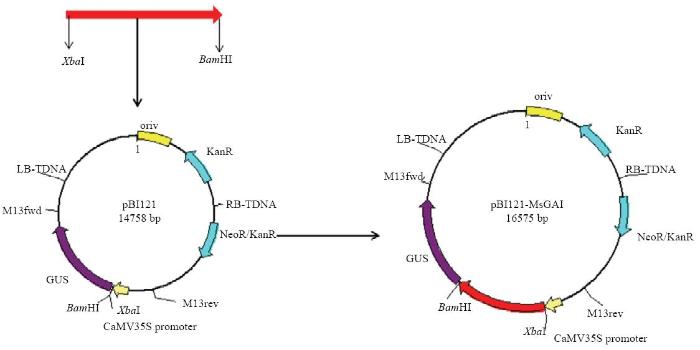 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图135S:MsGAI-gus超表达载体的构建
Fig. 1Structure of 35S:MsGAI-gus over-expression vector
1.8 紫花苜蓿的转化
以‘中苜1号’无菌苗为材料,挑选颜色为深绿色,肉眼观测无虫眼和病斑的叶片。经70%酒精消毒2 min,5%次氯酸钠消毒5 min,无菌水清洗3次后置于灭菌培养皿中。消毒后的外植体,在农杆菌溶液(OD600为0.6—0.8)重悬液中重悬15—30 min。在共培养基上培养48 h,然后转到诱导培养基,2—3周后将愈伤组织转入分化培养基,分化培养1—2个月后,愈伤组织将发育成丛生芽,经继代培养后,生成不定苗,将不定苗转至生根培养基,待不定苗生根后,移栽到营养土与蛭石比例1﹕1的花盆中,置人工气候室中培养。各类培养基的制备及具体操作流程,参照李俊等[27]方法。1.9 转基因植株的鉴定
提取转基因植株DNA,以121-MsGAI和35s/GUS为引物进行PCR扩增。将检测为阳性的转基因植株叶片浸泡在GUS染色液中,37℃过夜,次日脱色处理,观察结果。根据PCR验证及GUS染色结果,挑选长势良好的6个(L5、L7、L8、L9、L11和L12)转基因株系,利用特异引物QGAI进行qRT-PCR分析。1.10 不同处理下转基因植株的表达分析
在选取的6个转基因株系中,分别挑选出高(L5)、中(L8)、低(L11)3个株系,移入基质为营养土的花盆中生长。剪取植株下部成熟茎,通过扦插法进行扩繁。每个株系选取6个单株分别进行NaCl和PEG处理,即用0.3 mol·L-1 NaCl溶液和15%PEG均匀喷洒植物叶片,溶液分别在叶片正反面呈现挂珠状态。分别于0、2、4、8、12和24 h取样待用。2 结果
2.1 MsGAI的获得及生物信息学分析
PCR扩增得到2 005 bp的目的片段(图2),利用SnapGene软件对序列进行分析,该基因片段包含一个1 818 bp的开放阅读框,共编码605个氨基酸。该序列编码的蛋白理论等电点为4.71,相对分子量为71.11 kD,不稳定系数为40.28,属于不稳定蛋白,该蛋白平均亲水系数为-0.43,该蛋白为亲水性蛋白。利用DNAMAN软件将MsGAI与其他物种氨基酸序列进行同源比对,发现该蛋白序列含有GRAS家族典型的DELLA和GRAS结构域(图3),包含大量与鹰嘴豆、蒺藜苜蓿、陆地棉(Gossypium hirsutum)、小麦、玉米等物种DELLA蛋白同源的氨基酸序列。MsGAI氨基酸序列与豆科植物的相似度很高,与蒺藜苜蓿、鹰嘴豆、豌豆(Pisum Sativum)的相似度分别为96%、78%和79%。证明该序列为紫花苜蓿的GAI,故命名为MsGAI。图2
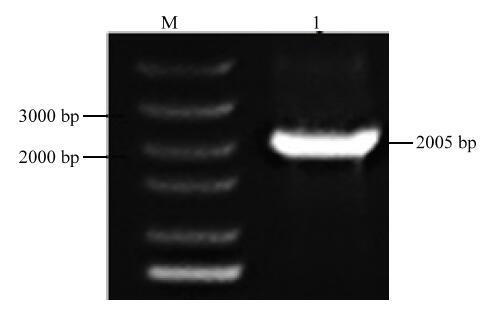 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2紫花苜蓿MsGAI的PCR扩增结果
M:DL5000分子标记;1:PCR扩增产物
Fig. 2PCR amplified product of MsGAI
M: DL5000 marker; 1: PCR amplified product
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3MsGAI与其他物种同源蛋白氨基酸序列比对
MsGAI:紫花苜蓿Medicago sativa;OsSLR1:水稻Oryza sativa,XP_015631543.1;MtGAI:蒺藜苜蓿Medicago truntula,XP_013460591.1;OsGAI:水稻Oryza sativa,Q7G7J6;CaGAI:鹰嘴豆Cicer arietinum,XP_004503135.1;GhGAI:陆地棉Gossypium hirsutum,XP_016744906.1;ZmDWARF8:玉米Zea mays,NC_024459.2;TaRHT-1:小麦Triticum aestivum,Q9ST59.1;VvGAI:葡萄Vitis vinifera,XP_002266267.1。方框分别表示DELLA域、TVHYNP和SAW域The DELLA domain, TVHYNP and SAW domains are represented in the boxes respectively
Fig. 3Amino acid sequence alignment of MsGAI with other homologous proteins
通过对MsGAI蛋白进行分析,结果表明,该蛋白有46.06%的α-螺旋,7.11%的β转角,34.62%的无规则卷曲,12.21%延伸链(图4)。该蛋白没有跨膜结构,可能为非跨膜蛋白,存在信号肽段的可能性很小。MsGAI可能定位在叶绿体,cTP(chloroplast transit peptide)得分为0.634(表2)。
图4
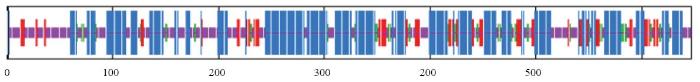 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4MsGAI蛋白二级结构的预测
蓝色:α-螺旋;红色:延伸链;绿色:β转角;橙色:无规则卷曲
Fig. 4Prediction of second structure of MsGAI
Blue: α-helix; Red: Extended chain; Green: β-turn; Orange: Random coil
Table 2
表2
表2亚细胞定位预测结果
Table 2
| 名称 Name | 长度 Length(bp) | 叶绿体 cTP | 线粒体 mTP | 信号肽 SP | Loc | RC |
|---|---|---|---|---|---|---|
| MsGAI | 605 | 0.634 | 0.097 | 0.036 | c | 5 |
新窗口打开|下载CSV
通过MEGA7.0软件,利用NJ法构建系统发育树,并且用Bootstrap进行校正,结果表明,MsGAI与蒺藜苜蓿GAI亲缘关系最近,其次与红三叶(Trifolium pratense)等双子叶豆科植物亲缘关系较近,与单子叶植物亲缘关系较远(图5)。
图5
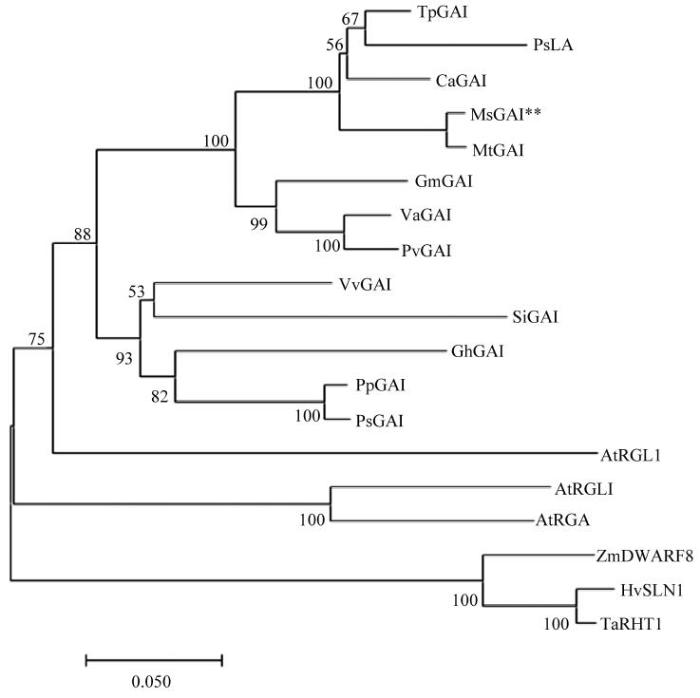 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5MsGAI蛋白与其他物种GAI蛋白的系统发育树
TpGAI:红三叶Trifolium pratense,PNY03470.1;PsLA:豌豆Pisum sativum,ABI30654.1;CaGAI:鹰嘴豆Cicer arietinum,XP_004503135.1;MsGAI:紫花苜蓿Medicago sativa;MtGAI:蒺藜苜蓿Medicago truntula,XP_013460591.1;GmGAI:大豆Glycine max,XP_003552980.1;VaGAI:赤豆Vigna angularis,XP_017418978.1;PvGAI:菜豆Phaseolus vulgaris,BAF62637.1;VvGAI:葡萄Vitis vinifera,ARR27421.1;SiGAI:芝麻Sesamum indicum,XP_011097451.1;GhGAI:陆地棉Gossypium hirsutum,XP_016744906.1;PpGAI:沙梨Pyrus pyrifolia,ANJ78486.1;PsGAI:李子Prunus salicina, AQQ12221.1; AtRGL1:拟南芥Arabidopsis thaliana,NC_003070.9;AtGAI:拟南芥Arabidopsis thaliana,NC_003070.9;AtRGA:拟南芥Arabidopsis thaliana,NC_003071.7;ZmDWARF8:玉米Zea mays,NC_024459.2;HvSLN1:青稞Hordeum vulgare,Q8W127.1;TaRHT1:小麦Triticum aestivum,Q9ST59.1。节点上的数值表示自举值,标尺上的数字表示遗传距离Numbers at node indicates bootstrap value derived from 1000 replicates. The scale bar indicates genetic distance
Fig. 5Phylogenetic tree of MsGAI and other GAI proteins
2.2 MsGAI启动子的生物信息学分析
利用PlantCARE启动子预测元件分析GAI启动子(表3)。结果显示,该基因启动子包含许多顺式作用元件,其中有光响应、昼夜节律响应元件,也存在与植物生长发育相关的元件,例如与分生组织表达相关的作用元件。同时还包含与干旱胁迫、热应激效应及低温响应相关的顺式元件。
Table 3
表3
表3PlantCARE启动子预测结果
Table 3
| 顺式作用元件 Cis-elements | 序列 Sequence (5'-3') | 功能 Function | 数量 No. |
|---|---|---|---|
| 3-AF1 binding site | AAGAGATATTT | 光响应元件 Light responsive element | 1 |
| A-box | CCGTCC | 顺式作用调控元件 Cis-acting regulatory element | 2 |
| ACE | AAAACGTTTA | 光响应顺式元件 Cis-acting element involved in light responsiveness | 1 |
| ARE | TGGTTT | 厌氧诱导必需元件 Cis-acting regulatory element essential for the anaerobic induction | 2 |
| ATCT-motif | AATCTAATCT | 参与光响应的保守DNA模块的部分元件 Part of a conserved DNA module involved in light responsiveness | 2 |
| BOX-4 | ATTAAT | 参与光反应元件 Part of a conserved DNA module involved in light responsiveness | 3 |
| Box-W1 | TTGACC | 真菌诱导子反应元件 Fungal elicitor responsive element | 1 |
| CAAT-box | CAAAT | 启动子和增强子区域中常见的顺式作用元件 Common cis-acting element in promoter and enhancer regions | 47 |
| CCAAT-box | CAACGG | 结合位点MYBHv1 binding site MYBHv1 | 1 |
| CCGTCC-box | CCGTCC | 与分生组织特异性激活相关的顺式调控元件 Cis-acting regulatory element related to meristem specific activation | 2 |
| GAG-motif | AGAGAGT | 部分光响应元件 Part of a light responsive element | 1 |
| GT1-motif | GGTTAA | 光响应元件 Light responsive element | 1 |
| HSE | AAAAAATTTC | 热应激响应顺式元件 Cis-acting element involved in heat stress responsiveness | 1 |
| LAMP-element | CCAAAACCA | 部分光响应元件 Part of a light responsive element | 1 |
| LTR | CCGAAA | 参与低温响应的元件 Cis-acting element involved in low-temperature responsiveness | 1 |
| MBS | TAACTG | 参与干旱诱导MYB结合位点 MYB binding site involved in drought-induction | 1 |
| MRE | AACCTAA | 参与光反应的MYB结合位点 MYB binding site involved in light responsiveness | 1 |
| Skn-1_motif | GTCAT | 胚乳表达所需的顺式作用调控元件 Cis-acting regulatory element required for endosperm expression | 2 |
| TATA-box | ATTATA | 在转录起始的-30附近的核心启动子元件 Core promoter element around -30 of transcription start | 74 |
| chs-CMA2a | GCAATTCC | 部分光响应元件 Part of a light responsive element | 1 |
| circadian | CAANNNNATC | 参与昼夜节律顺行调控元件 Cis-acting regulatory element involved in circadian control | 3 |
新窗口打开|下载CSV
2.3 MsGAI的组织特异性表达及不同处理条件下的表达分析
采用qRT-PCR技术分析MsGAI在紫花苜蓿各个组织中相对表达量,结果表明,MsGAI在紫花苜蓿的根、茎、叶、花瓣、花萼、雄蕊、雌蕊中均有不同程度表达。其中根中表达量最高,花瓣次之,茎中表达量最少(图6)。图6
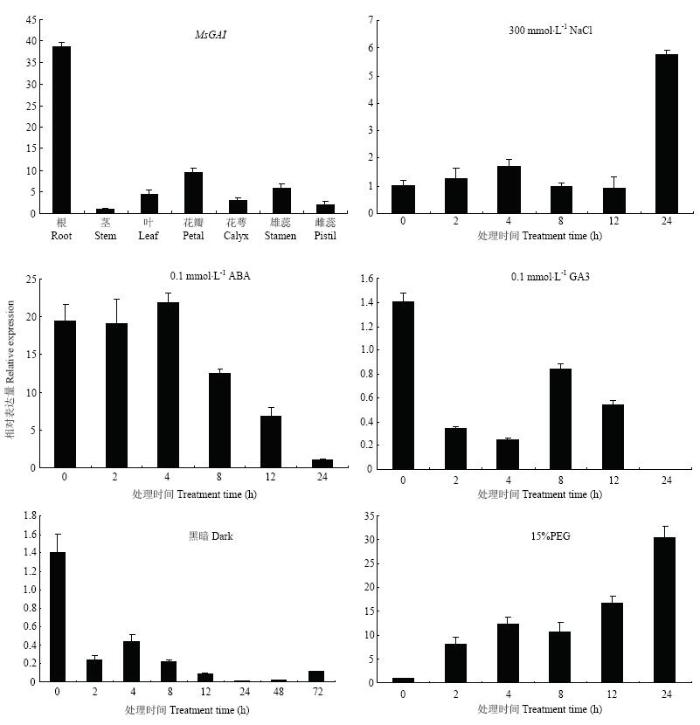 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6MsGAI的组织特异性表达及不同处理下的表达模式
Fig. 6Expression pattern of MsGAI in different tissues and different treatments
通过对不同处理条件下紫花苜蓿MsGAI的表达进行分析(图6)。GA3处理后,表达量总体呈现下降的趋势,在8 h有一个小幅的上升,24 h几乎检测不到MsGAI表达。ABA处理后,MsGAI的表达量在4 h达到峰值,之后逐渐降低。NaCl处理下,前期表达水平变化幅度不大,但在24 h表达量显著升高。PEG模拟干旱胁迫,MsGAI的表达量逐渐升高,到24 h时表达量升高到对照的30倍。黑暗处理后,MsGAI表达量迅速下降,在处理2 h后表达量降低了近10倍,并一直保持较低的表达水平。
2.4 35S:MsGAI-gus超表达载体的构建及紫花苜蓿的遗传转化
利用XbaⅠ和BamHⅠ对构建完善的pBI121植物超表达载体质粒进行双酶切检测,获得条带大小与预期一致(图7)。证明MsGAI已正向插入pBI121表达载体中,超表达载体35S:MsGAI-gus构建成功。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图735S:MsGAI-gus重组质粒的酶切鉴定
M:DL 5000 DNA marker;1:pBI121载体;2:35S:MsGAI-gus双酶切片段
Fig. 7Enzyme digestion identification of 35S:MsGAI-gus
M: DL 5000 DNA marker; 1: pBI121 vector; 2: Enzyme digestion fragments of 35S:MsGAI-gus
采用农杆菌介导的愈伤组织培养法对紫花苜蓿进行遗传转化(图8)。经过4个月的组织培养,目的片段成功转入紫花苜蓿,再生苗经GUS组织染色,呈现蓝色(图8-E),后将转基因幼苗移入人工气候室培养。
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8农杆菌介导的紫花苜蓿组织培养
A:外植体;B:愈伤组织;C:成熟小胚;D:阳性苗;E:GUS染色;F:成活的转基因植株
Fig. 8Transgenic Medicago sativa L. plants obtained after Agrobacterium tumefaciens-mediated
A: Exophyte; B: Callus; C: Mature embryo; D: Positive plants; E: Gus staining; F: Transgenic plant
待植株生长1月后,提取转基因植株DNA,进行PCR检测(图9)。对载体中的GUS进行PCR检测,结果显示,3对引物均扩增出大小相符的PCR片段。(图9)。
图9
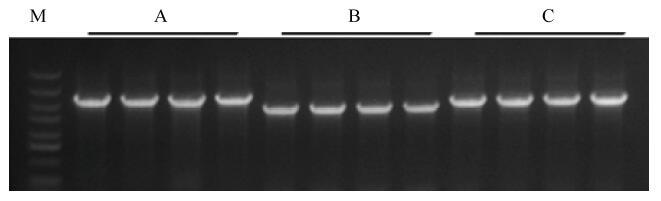 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9部分转基因紫花苜蓿的PCR检测
A:pBI121载体引物检测结果(1—4);B:MsGAI引物检测结果(5—8);C:上游载体引物下游基因引物检测结果(9—12)
Fig. 9PCR detection results of partial transgenic medicago sativa L.
A: Detection results used pBI121 vector primers; B: Detection results used MsGAI gene primers; C: Detection results used combination of pBI121 vector primers and MsGAI gene primers
2.5 转基因植株表达分析
对转基因植株进行qRT-PCR检测,结果表明,与对照相比,转基因紫花苜蓿中MsGAI的表达量较高,其中L5株系表达量最高,L11表达量相对较低(图10)。在15%PEG处理下,L5、L8、L11表达量上调,其中L5株系的表达量明显高于L11株系;与对照组相比,转基因植株各时间段MsGAI表达量均提高。300 mol·L-1 NaCl处理后,3个株系的MsGAI表达量均出现不同程度的提高,L5最为显著。图10
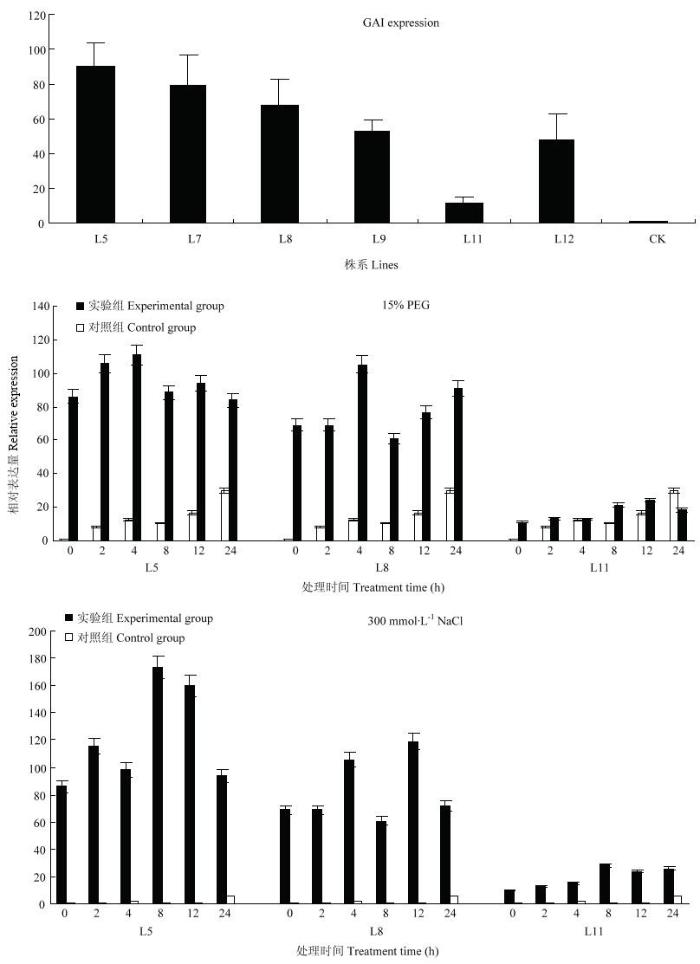 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10转基因紫花苜蓿中MsGAI表达量及不同处理下的表达模式
Fig. 10Expression pattern of MsGAI in transgenic medicago sativa L. and different treatments
3 讨论
3.1 MsGAI在豆科物种中高度保守
DELLA蛋白定位在细胞核中,蛋白序列结构特征分析以及蛋白核定位表明,DELLA蛋白属于一类潜在的转录因子[28]。在没有GA的情况下,DELLA蛋白阻遏植物生长发育;当DELLA蛋白上的GA信号感知域接收到信号以后,阻遏作用被抑制[29]。DELLA蛋白基因在许多植物中被成功克隆,功能研究表明,DELLA蛋白对植物种子萌发、开花时间、植株生物量方面均有重要影响[30]。本研究利用同源克隆的方法从紫花苜蓿中成功分离GAI基因,该基因编码的蛋白具有GRAS家族的典型结构域和保守区,其中包含DELLA蛋白N端保守结构域DELLA和TVHYNP,C端保守结构SAW。通过构建不同物种的DELLA蛋白系统发育树结果表明,紫花苜蓿MsGAI基因与蒺藜苜蓿、鹰嘴豆、红三叶、大豆(Glycine max)、豌豆等豆科植物的相似度较高,其中与蒺藜苜蓿的相似度高达96%。说明这类基因在进化过程中高度保守,具有相似的功能。基于此理论,参考模式物种同源基因的作用机制,探索出紫花苜蓿所特有的MsGAI功能,不失为一种有效的基因功能研究方法。
3.2 MsGAI响应紫花苜蓿非生物胁迫
MsGAI在紫花苜蓿的多个组织中均有表达,在根中的表达量最高,花中次之,茎中最低(图6)。因此推测,MsGAI在紫花苜蓿各个组织的生长发育过程中均可发挥作用。本研究结果显示,GA3处理后,MsGAI的表达量迅速降低,24 h后表达量降到最低,这有可能是因为外施赤霉素后,赤霉素与受体结合,然后与DELLA蛋白结合形成三元结构,最终DELLA蛋白被降解,导致编码DELLA蛋白的基因表达量下降。这一结果也与DELLA蛋白作为赤霉素信号通路中的阻遏因子的结论一致[31]。资料表明,ABA可通过影响细胞质子泵的生物活性来调节植物体的生长发育,植物受到外界胁迫时,ABA通过增强质子泵活性,从而刺激多种逆境蛋白的基因表达[32]。ZENTELLA等[33]提出DELLA蛋白能够诱导E3连接酶基因XERICO表达上调来促进ABA的积累。对谷物糊粉层的研究表明,ABA通过作用于DELLA蛋白下游来抑制GA反应[34]。因此,ABA可能参与了DELLA蛋白调控GA的信号途径,DELLA蛋白与ABA在GA调控植物生长发育过程中可能存在协同作用。本研究中,ABA处理初期,基因表达量变化不显著,4 h后,MsGAI表达量迅速下降。同时,将克隆得到的MsGAI启动子进行序列分析,结果显示,在MsGAI启动子区域含有大量与ABA抗逆相关的作用元件。模拟干旱胁迫处理后,紫花苜蓿对照组与转基因组,MsGAI的表达量逐渐上升并一直处于较高水平,同时盐胁迫也诱导该基因的表达,由此推断,MsGAI可能通过与ABA相互协同参与紫花苜蓿对盐及干旱的胁迫响应。
4 结论
从紫花苜蓿中分离1个DELLA蛋白基因MsGAI,包含1 818 bp的开放阅读框,该基因序列具有响应赤霉素的DELLA和VHYNP结构域,MsGAI与蒺藜苜蓿MtGAI的亲缘关系最近。MsGAI在根中表达量最高。转基因MsGAI紫花苜蓿对干旱、盐胁迫处理具有明显响应。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.7606/j.issn.1000-4025.2016.11.2159URL [本文引用: 1]

赤霉素受体(GID)是赤霉素信号转导途径的重要成员,直接影响着赤霉素对植物体效应的发挥。该研究利用同源克隆的方法,首次从紫花苜蓿中克隆得到1个赤霉素受体基因,命名为 MsGID 1b。序列分析发现,MsGID 1b基因开放阅读框长度为1053 bp,编码350个氨基酸,推测其蛋白质分子量为39.839 kD,是一个无信号肽和跨膜结构的亲水性蛋白。序列比对结果表明,MsGID 1b 基因与蒺藜苜蓿 MtGID 1b 基因的核苷酸序列相似性为98%,氨基酸序列相似性为99%,且具有 HSL 家族典型的 HGG 和 GXSXG 保守结构域及 GA、DELLA 蛋白结合位点。荧光定量 PCR 分析表明,MsGID 1b 基因在紫花苜蓿各组织中的表达丰度依次为:根>盛花>初花>茎>叶>荚果;经 GA3、ABA、NaCl、PEG 和黑暗诱导后该基因表达上调,尤其是在 GA3诱导下,MsGID 1b 基因的表达量一直维持在较高水平,表明 MsGID 1b 基因可能参与紫花苜蓿的抗逆调控。
DOI:10.7606/j.issn.1000-4025.2016.11.2159URL [本文引用: 1]
赤霉素受体(GID)是赤霉素信号转导途径的重要成员,直接影响着赤霉素对植物体效应的发挥。该研究利用同源克隆的方法,首次从紫花苜蓿中克隆得到1个赤霉素受体基因,命名为 MsGID 1b。序列分析发现,MsGID 1b基因开放阅读框长度为1053 bp,编码350个氨基酸,推测其蛋白质分子量为39.839 kD,是一个无信号肽和跨膜结构的亲水性蛋白。序列比对结果表明,MsGID 1b 基因与蒺藜苜蓿 MtGID 1b 基因的核苷酸序列相似性为98%,氨基酸序列相似性为99%,且具有 HSL 家族典型的 HGG 和 GXSXG 保守结构域及 GA、DELLA 蛋白结合位点。荧光定量 PCR 分析表明,MsGID 1b 基因在紫花苜蓿各组织中的表达丰度依次为:根>盛花>初花>茎>叶>荚果;经 GA3、ABA、NaCl、PEG 和黑暗诱导后该基因表达上调,尤其是在 GA3诱导下,MsGID 1b 基因的表达量一直维持在较高水平,表明 MsGID 1b 基因可能参与紫花苜蓿的抗逆调控。
DOI:10.2134/agronj1996.00021962003600060010xURL [本文引用: 1]
Abstract Alfalfa (Medicago sativa L.) is a deeply rooted perennial that may be able to remove NO3 from the subsoil beneath the rooting zone of annual crops. Our main objectives were to characterize the potential of two alfalfa cultivars (Agate and Ineffective Agate, an ineffectively nodulated near isoline of Agate) to remove NO3 from the subsoil and to evaluate the influence of available subsoil NO3 on symbiotic N2 fixation. Low ( 0.3 mM) or high (20 mM) NO3-N concentrations were supplied through a subsoil irrigation system installed in a Hubbard loamy sand soil (sandy, mixed Udorthentic Haploboroll) at Becker, MN. Nitrate uptake and N2 fixation were evaluated during two regrowth periods using 15N. During fall regrowth in the establishment year, supply of 20 mM 15N-enriched NO3 through subsoil irrigation did not decrease symbiotic N2 fixation; during summer regrowth of the first production year, symbiotic N2 fixation decreased only 40% (from 7.77 to 4.60 g N m-2). Even though it had higher yields, Agate alfalfa removed less subsoil NO3-N (1.52 and 5.51 g N m-2) than did Ineffective Agate (3.12 and 7.13 g N m-2) in each of two harvests where 15N was applied. Root length and mass after 2 yr were not altered by NO3 treatment or cultivar. This is the first time that direct measurements of subsoil nitrate removal have been made in the field for an N2-fixing crop. Because it is unlikely that differences in NO3 uptake efficiency could account for such cultivar differences, we propose that assimilation of atmospheric N2 reduced the capacity of the fixing crop to absorb NO3. These results suggest that non-N2-fixing alfalfa cultivars would be more useful for bioremediation of nitrate-contaminated sites than standard cultivars.
DOI:10.2134/jeq2001.30130xURLPMID:11215664 [本文引用: 1]
Abstract By 19%, standard remediation techniques had significantly reduced the concentration of nitrate nitrogen (NO3- -N) in local ground water at the site of a 1989 anhydrous ammonia spill, but NO3- -N concentrations in portions of the site still exceeded the public drinking water standard. Our objective was to determine whether local soil and ground water quality could be improved with alfalfa (Medicago sativa L.). A 3-yr study was conducted in replicated plots (24 by 30 m) located hydrologically upgradient of the ground water under the spill site. Three alfalfa entries ['Agate', Ineffective Agate (a non-N2-fixing elite germplasm similar to Agate), and MWNC-4 (an experimental germplasm)] were seeded in the spring of 1996. Corn (Zea mays L.) or wheat (Triticum aestivum L.) was seeded adjacent to the alfalfa each year. Crops were irrigated with N-containing ground water to meet water demand. During the 3-yr period, about 540 kg of inorganic N was removed from the aquifer through irrigation of 4.9 million L water. Cumulative N removal from the site over 3 yr was 972 kg N ha(-1) in Ineffective Agate alfalfa hay, compared with 287 kg N ha(-1) for the annual cereal grain. Soil solution NO3- concentrations were reduced to low and stable levels by alfalfa, but were more variable under the annual crops. Ground water quality improved, as evidenced by irrigation water N concentration. We do not know how much N was removed by the N2-fixing alfalfas, but it appears that either fixing or non-N2-fixing alfalfa will effectively remove inorganic N from N-affected sites.
DOI:10.1016/S1360-1385(00)01790-8URLPMID:11120474 [本文引用: 1]
The identification of most of the genes involved in the metabolic pathways for gibberellin hormones has helped us to understand these pathways and their regulation. Many of these enzymes are multifunctional and therefore fewer enzymes than might be expected are required to synthesize the various gibberellin structures. However, several of the enzymes are encoded by multiple genes that are regulated differently, adding unexpected genetic complexity. Several endogenous and environmental factors modify the expression of gibberellin biosynthesis genes, including developmental stage, hormonal status and light. A future challenge will be to dissect the complex, interacting pathways that mediate the regulation of gibberellin metabolism.
.
DOI:10.1199/tab.0103URLPMID:22303234 [本文引用: 1]
Abstract Bioactive gibberellins (GAs) are diterpene phytohormones that modulate growth and development throughout the whole life cycle of the plant. Arabidopsis genes encoding most GA biosynthesis and catabolism enzymes, as well as GA receptors (GIBBERELLIN INSENSITIVE DWARF1, GID1) and early GA signaling components have been identified. Expression studies on the GA biosynthesis genes are beginning to reveal the potential sites of GA biosynthesis during plant development. Biochemical and genetic analyses demonstrate that GA de-represses its signaling pathway by binding to GID1s, which induce degradation of GA signaling repressors (DELLAs) via an ubiquitin-proteasome pathway. To modulate plant growth and development, the GA pathway is also regulated by endogenous signals (other hormones) and environmental cues (such as light, temperature and salt stress). In many cases, these internal and external cues directly affect GA metabolism and bioactive GA levels, and indirectly alter DELLA accumulation and GA responses. Importantly, direct negative interaction between DELLA and PIF3 and PIF4 (2 phytochrome interacting transcription factors) appears to integrate the effects of light and GA on hypocotyl elongation.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/dnares/9.1.11URLPMID:11939564 [本文引用: 1]
A rice semidwarfing gene, sd-1, known as the green revolution gene, was isolated by positional cloning and revealed to encode gibberellin 20-oxidase, the key enzyme in the gibberellin biosynthesis pathway. Analysis of 3477 segregants using several PCR-based marker technologies, including cleaved amplified polymorphic sequence, derived-CAPS, and single nucleotide polymorphisms revealed 1 ORF in a 6-kb candidate interval. Normal-type rice cultivars have an identical sequence in this region, consisting of 3 exons (558, 318, and 291 bp) and 2 introns (105 and 1471 bp). Dee-Geo-Woo-Gen-type sd-1 mutants have a 383-bp deletion from the genome (278-bp deletion from the expressed sequence), from the middle of exon 1 to upstream of exon 2, including a 105-bp intron, resulting in a frame-shift that produces a termination codon after the deletion site. The radiation-induced sd-1 mutant Calrose 76 has a 1-bp substitution in exon 2, causing an amino acid substitution (Leu [CTC] to Phe [TTC]). Expression analysis suggests the existence of at least one more locus of gibberellin 20-oxidase which may prevent severe dwarfism from developing in sd-1 mutants.
DOI:10.1038/416701aURL [本文引用: 1]
[本文引用: 1]
.
DOI:10.1046/j.1365-313X.1999.00431.xURLPMID:10341448 [本文引用: 1]
Mutations at the SCARECROW (SCR) locus in Arabidopsis thaliana result in defective radial patterning in the root and shoot. The SCR gene product contains sequences which suggest that it is a transcription factor. A number of Arabidopsis Expressed Sequence Tags (ESTs) have been identified that encode gene products bearing remarkable similarity to SCR throughout their carboxyl-termini, indicating that SCR is the prototype of a novel gene family. These ESTs have been designated SCARECROW-LIKE (SCL). The gene products of the GIBBERELLIN-INSENSITIVE (GAI) and the REPRESSOR of ga1 3 (RGA) loci show high structural and sequence similarity to SCR and the SCLs. Sequence analysis of the products of the GRAS (GAI, RGA, SCR) gene family indicates that they share a variable amino-terminus and a highly conserved carboxyl-terminus that contains five recognizable motifs. The SCLs have distinct patterns of expression, but all of those analyzed show expression in the root. One of them, SCL3, has a tissue-specific pattern of expression in the root similar to SCR. The importance of the GRAS gene family in plant biology has been established by the functional analyses of SCR, GAI and RGA.
DOI:10.1038/22307URL [本文引用: 1]
.
[本文引用: 1]
DOI:10.1104/pp.010918URL [本文引用: 1]
DOI:10.1105/tpc.010319URLPMID:11826299 [本文引用: 1]
The slender rice1 mutant (slr1) shows a constitutive gibberellin (GA) response phenotype. To investigate the mode of action of SLR1, we generated transgenic rice expressing a fusion protein consisting of SLR1 and green fluorescent protein (SLR1-GFP) and analyzed the phenotype of the transformants and the subcellular localization of GFP in vivo. SLR1-GFP worked in nuclei to repress the GA signaling pathway; its overproduction caused a dwarf phenotype. Application of GA3 to SLR1-GFP overproducers induced GA actions such as shoot elongation, downregulation of GA 20-oxidase expression, and upregulation of SLR1 expression linked with the disappearance of the nuclear SLR1-GFP protein. We also performed domain analyses of SLR1 using transgenic plants overproducing different kinds of truncated SLR1 proteins. The analyses revealed that the SLR1 protein can be divided into four parts: a GA signal perception domain located at the N terminus, a regulatory domain for its repression activity, a dimer formation domain essential for signal perception and repression activity, and a repression domain at the C terminus. We conclude that GA signal transduction is regulated by the appearance or disappearance of the nuclear SLR1 protein, which is controlled by the upstream GA signal.
.
DOI:10.1101/gad.11.23.3194URLPMID:9389651 [本文引用: 1]
The Arabidopsis gai mutant allele confers a reduction in gibberellin (GA) responsiveness. Here we report the molecular cloning of GAI and a closely related gene GRS. The predicted GAI (wild-type) and gai (mutant) proteins differ only by the deletion of a 17-amino-acid segment from within the amino-terminal region. GAI and GRS contain nuclear localization signals, a region of homology to a putative transcription factor, and motifs characteristic of transcriptional coactivators. Genetic analysis indicates that GAI is a repressor of GA responses, that GA can release this repression, and that gai is a mutant repressor that is relatively resistant to the effects of GA. Mutations at SPY and GAR2 suppress the gai phenotype, indicating the involvement of GAI, SPY, and GAR2 in a signaling pathway that regulates GA responses negatively. The existence of this pathway suggests that GA modulates plant growth through derepression rather than through simple stimulation.
.
[本文引用: 1]
.
DOI:10.1101/gad.969002URLPMID:11877383 [本文引用: 1]
The germination of Arabidopsis seeds is promoted by gibberellin (GA). Arabidopsis GAI, and RGA are genes encoding key GA signal-transduction components (GAI and RGA) that mediate GA regulation of stem elongation. The Arabidopsis genome contains two further genes, RGL1 and RGL2, that encode proteins (RGL1 and RGL2) that are closely related to GAI and RGA. Here, we show that RGL2 regulates seed germination in response to GA, and that RGL1, GAI, and RGA do not. In addition, we show that RGL2 transcript levels rise rapidly following seed imbibition, and then decline rapidly as germination proceeds. In situ GUS staining revealed that RGL2 expression in imbibed seeds is restricted to elongating regions of pre-emergent and recently emerged radicles. These observations indicate that RGL2 is a negative regulator of GA responses that acts specifically to control seed germination rather than stem elongation. Furthermore, as RGL2 expression is imbibition inducible, RGL2 may function as an integrator of environmental and endogenous cues to control seed germination.
.
[本文引用: 1]
.
DOI:10.1007/BF02341024URL [本文引用: 1]
The endogenous gibberellins (GAs) from shoots of the GA-insensitive mutant, gai, of Arabidopsis thaliana were analyzed and compared with the GAs from the Landsberg erecta (Ler) line. Twenty GAs were identified in Ler plants by full-scan gas chromatography-mass spectrometry (GC-MS) and Kovats retention indices (KRI's). These GAs are members of the early-13-hydroxylation pathway (GA53, GA44, GA19, GA17, GA20, GA1, GA29, and GA8), the non-3,13-hydroxylation pathway (GA12, GA15, GA24, GA25, GA9, and GA51), and the early-3-hydroxylation pathway (GA37, GA27, GA36, GA13, GA4, and GA34). The same GAs, except GA53, GA44, GA37, and GA29 were detected in the gai mutant by the same methods. In addition, extracts from gai plants contained GA41 and GA71. Both lines also contained several unknown GAs. In Ler plants these were mainly hydroxy-GA12 derivatives, whereas in the gai mutant hydroxy-GA24, hydroxy-GA25, and hydroxy-GA9 compounds were detected. Quantification of seven GAs by GC-selected ion monitoring (SIM), using internal standards, and comparisons of the ion intensities in the SIM chromatograms of the other thirteen GAs, demonstrated that the gai mutant had reduced levels of all C20-dicarboxylic acids (GA53, GA44, GA19, GA12, GA15, GA24, GA37, GA27, and GA36). In contrast, gai plants had increased levels of C20-tricarboxylic acid GAs (GA17, GA25, and GA41) and of all C19-GAs (GA20, GA1, GA8, GA9, GA51, GA4, GA34, and GA71) except GA29. The 3 -hydroxylated GAs, GA1, and GA4, and their respective 2 -hydroxylated derivatives, GA8 and GA34, were the most abundant GAs found in shoots of the gai mutant. Thus, the gai mutation in Arabidopsis results in a phenotype that resembles GA-deficient mutants, is insensitive to both applied and endogenous GAs, and contains low levels of C20-dicarboxylic acid GAs and high levels of C19-GAs. This indicates that the GAI gene controls a step beyond the synthesis of an active GA. The gai mutant is presumably a GA-receptor mutant or a mutant with a block in the transduction pathway between the receptor and stem elongation.
.
DOI:10.1073/pnas.87.20.7983URLPMID:2236013 [本文引用: 1]
Twenty gibberellins (GAs) have been identified in extracts from shoots of the Landsberg erecta line of Arabidopsis thaliana by full-scan gas chromatography-mass spectrometry and Kovats retention indices. Eight of them are members of the early-13-hydroxylation pathway (GA53, GA44, GA19, GA17, GA20, GA1, GA29, and GA8), six are members of the early-3-hydroxylation pathway (GA37, GA27, GA36, GA13, GA4, and GA34), and the remaining six are members of the non-3,13-hydroxylation pathway (GA12, GA15, GA24, GA25, GA9, and GA51). Seven of these GAs were quantified in the Landsberg erecta line of Arabidopsis and in the semidwarf ga4 and ga5 mutants by gas chromatography-selected ion monitoring (SIM) using internal standards. The relative levels of the remaining 13 GAs were compared by the use of ion intensities only. In comparison with the Landsberg erecta line, the ga4 mutant had reduced levels of the 3-hydroxy- and 3,13-dihydroxy-GAs, and it accumulated the 13-hydroxy-GAs, except GA53, and the non-3,13-hydroxy-GAs, except GA12. The GA4 gene encodes, therefore, a protein with 3 -hydroxylation activity. The ga5 mutant had reduced levels of the C19-GAs, which indicates that the product of the GA5 gene catalyzes the elimination of C-20 at the aldehyde level. The ga5 mutant also had increased levels of certain C20-GAs, which indicates existence of an additional control, possibly hydroxylation of C-20. The growth-response data, as well as the accumulation of GA9in the ga4 mutant, indicate that GA9is not active in Arabidopsis, but it must be 3 -hydroxylated to GA4to become bioactive. It is concluded that the reduced levels of the 3 -hydroxy-GAs, GA1and GA4, are the cause of the semidwarf growth habit of both mutants.
.
DOI:10.1104/pp.108.2.475URLPMID:12228487 [本文引用: 1]
The semidominant gibberellin-insensitive (gai) mutant of Arabidopsis thaliana shows impairment in multiple responses to the plant hormone gibberellin A3, which include effects on seed germination, stem elongation, apical dominance, and rapid flowering in short days. Results presented here show that the gai mutation also interferes with development of fertile flowers in continuous light. Mutagenesis of the gai mutant resulted in recovery of 17 independent mutants in which the gibberellin-insensitive phenotype is partially or completely suppressed. Sixteen of the suppressor mutations act semidominantly to restore gibberellin responsiveness. One representative of this class, the gar1 mutation, could not be genetically separated from the gai locus and is proposed to cause inactivation of the gai gene. The exceptional gar2 mutation partially suppresses the gai phenotype, is completely dominant, and is not linked to the gai locus. The gar2 mutation may define a new gene involved in gibberellin signaling. A recessive allele of the spindly (SPY) locus, spy-5, was also found to partially suppress the gai mutant phenotype.
.
DOI:10.1038/nature01387URLPMID:12610625 [本文引用: 1]
The growth of plant organs is influenced by a stream of the phytohormone auxin that flows from the shoot apex to the tip of the root. However, until now it has not been known how auxin regulates the cell proliferation and enlargement that characterizes organ growth. Here we show that auxin controls the growth of roots by modulating cellular responses to the phytohormone gibberellin (GA). GA promotes the growth of plants by opposing the effects of nuclear DELLA protein growth repressors, one of which is Arabidopsis RGA (for repressor of gal-3). GA opposes the action of several DELLA proteins by destabilizing them, reducing both the concentration of detectable DELLA proteins and their growth-restraining effects. We also show that auxin is necessary for GA-mediated control of root growth, and that attenuation of auxin transport or signalling delays the GA-induced disappearance of RGA from root cell nuclei. Our observations indicate that the shoot apex exerts long-distance control on the growth of plant organs through the effect of auxin on GA-mediated DELLA protein destabilization.
.
[本文引用: 1]
[本文引用: 1]
DOI:10.1006/meth.2001.1262URL [本文引用: 1]
URL [本文引用: 1]
东方山羊豆(Galega orientalis L.)属于豆科山羊豆属,为优质多年生豆科牧草,具有重要的生态和经济价值。提高牧草生物量、抗逆性是牧草育种的主要内容。植物基因资源挖掘与利用,是牧草遗传改良的重要技术途径。本论文选择东方山羊豆赤霉素受体、水通道蛋白作为研究对象,开展了基因功能验证及转基因研究工作。 本试验主要取得如下进展: 1.通过酵母双杂交实验表明,东方山羊豆赤霉素受体(GoGID)可以与拟南芥DELLA蛋白在GA存在条件下互作。通过突变体回复实验表明,GoGID能够部分回复拟南芥突变体gid1ac;完全回复突变体gid1ab为正常表型。表明GoGID编码蛋白能够部分替代AtGIDs功能,具有生物学功能。 2.构建表达载体pBI121-GoGID-GUS,通过农杆菌介导的花序浸染法转化拟南芥。经过后代的筛选、扩繁得到T3代转基因株系。研究表明,转基因拟南芥植株高度、鲜重、干重、种子产量都增加,其中植株干重比对照提高90%~160%。 3.采用农杆菌介导的愈伤侵染法,转化紫花苜蓿。转基因苜蓿及对照经扦插繁殖,用于生物学观察及实验。结果表明,转基因紫花苜蓿GroupⅠ、GroupⅡ组,植株干重比对照提高8%-46%;GroupⅢ干重与对照相似,但叶/茎比提高。 4.在东方山羊豆一个受盐诱导EST基础上,克隆得到与植物AQP同源的基因。该基因编码289个氨基酸,与植物的PIP基因序列同源性最高,属于PIP1亚类,命名为GoPIP1,在Genbank中登录HM991477。 5. GoPIP1基因在根、茎、叶中都有表达,其中在根中表达量最高。受盐、干旱强烈诱导。亚细胞分析表明,定位于细胞膜。在非胁迫条件下,转基因植株地上部/地下部比例提高,叶片失水速率增加;在盐份条件下,转基因株系与对照基本没有差异,而且GoPIP1基因没有促进离子转运。在模拟干旱条件下,转基因拟南芥鲜重降低、含水量降低,体内脯氨酸含量降低、丙二醛含量提高;转基因拟南芥,离体叶片失水速率增加,叶片渗透势增加;同时转基因植株中,ABA合成途径及受ABA诱导基因表达量增加。GoPIP1增加了转基因拟南芥对干旱敏感性。
URL [本文引用: 1]
东方山羊豆(Galega orientalis L.)属于豆科山羊豆属,为优质多年生豆科牧草,具有重要的生态和经济价值。提高牧草生物量、抗逆性是牧草育种的主要内容。植物基因资源挖掘与利用,是牧草遗传改良的重要技术途径。本论文选择东方山羊豆赤霉素受体、水通道蛋白作为研究对象,开展了基因功能验证及转基因研究工作。 本试验主要取得如下进展: 1.通过酵母双杂交实验表明,东方山羊豆赤霉素受体(GoGID)可以与拟南芥DELLA蛋白在GA存在条件下互作。通过突变体回复实验表明,GoGID能够部分回复拟南芥突变体gid1ac;完全回复突变体gid1ab为正常表型。表明GoGID编码蛋白能够部分替代AtGIDs功能,具有生物学功能。 2.构建表达载体pBI121-GoGID-GUS,通过农杆菌介导的花序浸染法转化拟南芥。经过后代的筛选、扩繁得到T3代转基因株系。研究表明,转基因拟南芥植株高度、鲜重、干重、种子产量都增加,其中植株干重比对照提高90%~160%。 3.采用农杆菌介导的愈伤侵染法,转化紫花苜蓿。转基因苜蓿及对照经扦插繁殖,用于生物学观察及实验。结果表明,转基因紫花苜蓿GroupⅠ、GroupⅡ组,植株干重比对照提高8%-46%;GroupⅢ干重与对照相似,但叶/茎比提高。 4.在东方山羊豆一个受盐诱导EST基础上,克隆得到与植物AQP同源的基因。该基因编码289个氨基酸,与植物的PIP基因序列同源性最高,属于PIP1亚类,命名为GoPIP1,在Genbank中登录HM991477。 5. GoPIP1基因在根、茎、叶中都有表达,其中在根中表达量最高。受盐、干旱强烈诱导。亚细胞分析表明,定位于细胞膜。在非胁迫条件下,转基因植株地上部/地下部比例提高,叶片失水速率增加;在盐份条件下,转基因株系与对照基本没有差异,而且GoPIP1基因没有促进离子转运。在模拟干旱条件下,转基因拟南芥鲜重降低、含水量降低,体内脯氨酸含量降低、丙二醛含量提高;转基因拟南芥,离体叶片失水速率增加,叶片渗透势增加;同时转基因植株中,ABA合成途径及受ABA诱导基因表达量增加。GoPIP1增加了转基因拟南芥对干旱敏感性。
DOI:10.3969/j.issn.1674-3466.2006.05.006URL [本文引用: 1]
The phytohormone gibberellins (GA) play an important role in controlling and modulating diverse developmental processes, such as seed germination, hypocotyls elongation, leaf expansion and flowering time.In recent years, there are significant progresses in understanding of GA biosynthesis pathway and GA signaling in Arabidopsis and rice. This review highlights GA biosynthesis pathway and their regulation, including a new pathway identified in rice, and molecular model of“ De-repress” in GA signaling. GA promotes plant growth via 26S proteasome-dependent proteolysis of DELLA proteins repressors, one of the key component in GAsignaling, and also depends on GA-mediated interaction between GA receptor and DELLA proteins. Finally, this paper discussed the cross-talking between GA and other hormones, and modulation of adaptation to environments.
DOI:10.3969/j.issn.1674-3466.2006.05.006URL [本文引用: 1]
The phytohormone gibberellins (GA) play an important role in controlling and modulating diverse developmental processes, such as seed germination, hypocotyls elongation, leaf expansion and flowering time.In recent years, there are significant progresses in understanding of GA biosynthesis pathway and GA signaling in Arabidopsis and rice. This review highlights GA biosynthesis pathway and their regulation, including a new pathway identified in rice, and molecular model of“ De-repress” in GA signaling. GA promotes plant growth via 26S proteasome-dependent proteolysis of DELLA proteins repressors, one of the key component in GAsignaling, and also depends on GA-mediated interaction between GA receptor and DELLA proteins. Finally, this paper discussed the cross-talking between GA and other hormones, and modulation of adaptation to environments.
DOI:10.1016/j.plaphy.2006.06.012URL [本文引用: 1]
.
DOI:10.1105/tpc.107.054999URLPMID:17933900 [本文引用: 1]
Bioactive gibberellins (GAs) are phytohormones that regulate growth and development throughout the life cycle of plants. DELLA proteins are conserved growth repressors that modulate all aspects of GA responses. These GA-signaling repressors are nuclear localized and likely function as transcriptional regulators. Recent studies demonstrated that GA, upon binding to its receptor, derepresses its signaling pathway by binding directly to DELLA proteins and targeting them for rapid degradation via the ubiquitin-proteasome pathway. Therefore, elucidating the signaling events immediately downstream of DELLA is key to our understanding of how GA controls plant development. Two sets of microarray studies followed by quantitative RT-PCR analysis allowed us to identify 14 early GA-responsive genes that are also early DELLA-responsive in Arabidopsis thaliana seedlings. Chromatin immunoprecipitation provided evidence for in vivo association of DELLA with promoters of eight of these putative DELLA target genes. Expression of all 14 genes was downregulated by GA and upregulated by DELLA. Our study reveals that DELLA proteins play two important roles in GA signaling: (1) they help establish GA homeostasis by direct feedback regulation on the expression of GA biosynthetic and GA receptor genes, and (2) they promote the expression of downstream negative components that are putative transcription factors/regulators or ubiquitin E2/E3 enzymes. In addition, one of the putative DELLA targets, XERICO, promotes accumulation of abscisic acid (ABA) that antagonizes GA effects. Therefore, DELLA may restrict GA-promoted processes by modulating both GA and ABA pathways.
.
DOI:10.1111/jipb.12313URLPMID:25418692 [本文引用: 1]
Plant responses to abiotic stresses are coordinated by arrays of growth and developmental programs.Gibberellic acid(CA) and abscisic acid(ABA) play critical roles in the developmental programs and environmental responses,respectively,through complex signaling and metabolism networks.However,crosstalk between the two phytohormones in stress responses remains largely unknown.In this study,we report that CIBBERELLIN-INSENSITIVE DWARF 1(GID1),a soluble receptor for GA,regulates stomatal development and patterning in rice(Oryza sativa L.).The gid1 mutant showed impaired biosynthesis of endogenous ABA under drought stress conditions,but it exhibited enhanced sensitivity to exogenous ABA.Scanning electron microscope and infrared thermal image analysis indicated an increase in the stomatal conductance in the gid1 mutant under drought conditions.Interestingly,the gid1 mutant had increased levels of chlorophyll and carbohydrates under submergence conditions,and showed enhanced reactive oxygen species(ROS)-scavenging ability and submergence tolerance compared with the wild-type.Further analyses suggested that the function of GID1 in submergence responses is partially dependent on ABA,and GA signaling by GID1 is involved in submergence tolerance by modulating carbohydrate consumption.Taken together,these findings suggest GID1 plays distinct roles in stomatal response and submergence tolerance through both the ABA and GA signaling pathways in rice.
DOI:10.1111/j.1365-313X.2007.03122.xURLPMID:17488236 [本文引用: 1]
Plants can sense neighbour competitors through light-quality signals and respond with shade-avoidance responses. These include increased shoot elongation, which enhances light capture and thus competitive power. Such plant lant interactions therefore profoundly affect plant development in crowded populations. Shade-avoidance responses are tightly coordinated by interactions between light signals and hormones, with essential roles for the phytochrome B photoreceptor [sensing the red:far red (R:FR) ratio] and the hormone gibberellin (GA). The family of growth-suppressing DELLA proteins are targets for GA signalling and are proposed to integrate signals from other hormones. However, the importance of these regulators has not been studied in the ecologically relevant, complex realm of plant canopies. Here we show that DELLA abundance is regulated during growth responses to neighbours in dense Arabidopsis stands. This occurs in a R:FR-dependent manner in petioles, depends on GA, and matches the induction kinetics of petiole elongation. Similar interactions were observed in the growth response of seedling hypocotyls and are general for a second canopy signal, reduced blue light. Enhanced DELLA stability in the gai mutant inhibits shade-avoidance responses, indicating that DELLA proteins constrain shade-avoidance. However, using multiple DELLA knockout mutants, we show that the observed DELLA breakdown is not sufficient to induce shade-avoidance in petioles, but plays a more central role in hypocotyls. These data provide novel information on the regulation of shade-avoidance under ecologically important conditions, defining the importance of DELLA proteins and GA and unravelling the existence of GA- and DELLA-independent mechanisms.
.
DOI:10.1105/tpc.107.054999URLPMID:17933900 [本文引用: 1]
Bioactive gibberellins (GAs) are phytohormones that regulate growth and development throughout the life cycle of plants. DELLA proteins are conserved growth repressors that modulate all aspects of GA responses. These GA-signaling repressors are nuclear localized and likely function as transcriptional regulators. Recent studies demonstrated that GA, upon binding to its receptor, derepresses its signaling pathway by binding directly to DELLA proteins and targeting them for rapid degradation via the ubiquitin-proteasome pathway. Therefore, elucidating the signaling events immediately downstream of DELLA is key to our understanding of how GA controls plant development. Two sets of microarray studies followed by quantitative RT-PCR analysis allowed us to identify 14 early GA-responsive genes that are also early DELLA-responsive in Arabidopsis thaliana seedlings. Chromatin immunoprecipitation provided evidence for in vivo association of DELLA with promoters of eight of these putative DELLA target genes. Expression of all 14 genes was downregulated by GA and upregulated by DELLA. Our study reveals that DELLA proteins play two important roles in GA signaling: (1) they help establish GA homeostasis by direct feedback regulation on the expression of GA biosynthetic and GA receptor genes, and (2) they promote the expression of downstream negative components that are putative transcription factors/regulators or ubiquitin E2/E3 enzymes. In addition, one of the putative DELLA targets, XERICO, promotes accumulation of abscisic acid (ABA) that antagonizes GA effects. Therefore, DELLA may restrict GA-promoted processes by modulating both GA and ABA pathways.
DOI:10.2307/3871414URL [本文引用: 1]
The antagonism between gibberellins (GA) and abscisic acid (ABA) is an important factor regulating the developmental transition from embryogenesis to seed germination. In barley aleurone layers, the expression of genes encoding α-amylases and proteases is induced by GA but suppressed by ABA. It has been shown that an ABA-induced protein kinase, PKABA1, mediates the ABA suppression of α-amylase expression. Using a barley aleurone transient expression system, we have now localized the site of action of PKABA1 relative to other signal transduction components governing the expression of α-amylase. The expression of α-amylase can be transactivated by the transcription factor GAMyb, which is itself induced by GA. A truncated GAMyb containing the DNA binding domain but lacking the transactivation domain prevents the GA induction of α-amylase, further supporting the notion that GAMyb mediates the GA induction of α-amylase expression. Although ABA and PKABA1 strongly inhibit the GA induction of α-amylase, they have no effect on GAMyb-transactivated α-amylase expression. Using a GAMyb promoter-β-glucuronidase construct, we also show that both ABA and PKABA1 repress the GA induction of GAMyb. In the slender mutant, GAMyb and α-amylase are highly expressed, even in the absence of GA. However, this constitutive expression can still be inhibited by ABA, PKABA1, or an inhibitor of cGMP synthesis. On the basis of these observations, we suggest that PKABA1 acts upstream from the formation of functional GAMyb but downstream from the site of action of the Slender gene product. Because PKABA1 inhibits the GA induction of the GAMyb promoter-β-glucuronidase construct, it appears that at least part of the action of PKABA1 is to downregulate GAMyb at the transcriptional level.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}