 ,1, 程宪国,1
,1, 程宪国,1Enrichment Profile of GA4 is an Important Regulatory Factor Triggering Rice Dwarf
HUANG ShengCai1, WANG Bing1, XIE GuoQiang2, LIU ZhongLai2, ZHANG MeiJuan1, ZHANG ShuQing,1, CHENG XianGuo,1通讯作者:
收稿日期:2018-11-6接受日期:2018-12-9网络出版日期:2019-03-01
| 基金资助: |
Received:2018-11-6Accepted:2018-12-9Online:2019-03-01
作者简介 About authors
黄升财,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (2445KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
黄升财, 王冰, 谢国强, 刘中来, 张美娟, 张树清, 程宪国. 赤霉素GA4是水稻矮化特征的重要调节因子[J]. 中国农业科学, 2019, 52(5): 786-800 doi:10.3864/j.issn.0578-1752.2019.05.002
HUANG ShengCai, WANG Bing, XIE GuoQiang, LIU ZhongLai, ZHANG MeiJuan, ZHANG ShuQing, CHENG XianGuo.
0 引言
【研究意义】水稻是人类生存至关重要的粮食作物之一,水稻产量通常由分蘖数、穗粒数、单粒重及株高等农艺性状决定。随着人口压力逐渐增加,耕地面积逐渐减小,气候和环境恶化等问题的出现,人类对粮食产量和质量的要求愈来愈高。探明水稻矮化和分蘖增多的分子机制,为培育抗倒伏及高有效分蘖的水稻品种提供科学的理论支持,对保证人类粮食安全具有重要的意义。【前人研究进展】植物生长高度在很大程度上受内源激素的影响,包括赤霉素[1,2]、油菜素类固醇[3,4]以及独脚金内酯[5,6]等。其中赤霉素(GAs)是一类二萜类化合物,可促进植物生命周期各个阶段的生长:种子萌发、茎伸长、叶片膨大、开花和花发育[7]。赤霉素的合成受古巴焦磷酸合成酶(copalyl pyrophosphate synthase,CPS)、内根-贝壳杉烯合成酶(ent-kaurene synthase,KS)、内根-贝壳杉烯19-氧化酶(ent-kaurene oxidase,KO)、GA-20-氧化酶(gibberellin- 20-oxidase,GA20ox)及GA-3-氧化酶(gibberellin-3- oxidase,GA3ox)等多个酶统一调控[8]。在拟南芥中,ga1 (CPS突变体)[9]、ga2 (KS突变体)[10]、ga3 (KO突变体)[11]均出现矮化现象。水稻赤霉素合成基因OsKS[12]、OsKO (d35 )[13]及OsGA20ox1 (sd1 )[1]突变也会使水稻出现不同程度的矮化。一般情况下,赤霉素合成突变体除了株高会发生变化之外,另外一个特征是对外源GA敏感。而水稻gid1[14]和slr1[15]突变体均对外源GA不敏感,表明GID1(赤霉素特异性受体)和SLR1(DELLA蛋白)参与植物体内赤霉素信号的转导。相关研究表明,当细胞外GA浓度较低时,GID1不与GA结合,其N端结构域(N-Ex)呈疏松状态,使DELLA蛋白与GA早期应答基因结合,并通过抑制应答基因的翻译过程进而使其失活。当细胞外GA浓度较高时,GA诱导GID1构象发生变化,并与其结合形成GA-GID1复合物。该复合物具有疏水性的表面以便与DELLA蛋白形成三聚体,促进DELLA与泛素E3连接酶复合体结合,诱导DELLA蛋白被26S蛋白酶降解,进而GA信号得以释放[16]。矮化突变体除了株高变矮之外,通常还伴随着花发育异常和分蘖增多等现象。尹昌喜[17]研究表明OsGA3ox2 下调表达及OsGA2ox1 上调表达导致穗茎节间GA1含量减少是水稻出现半矮化和包穗的原因。而张玲[18]和WANG等[19]分别利用图位克隆和BSA鉴定了新的基因DDF2 和DDF3 ,这两个基因的突变均能导致水稻半矮化且花发育异常。独脚金内酯(strigolactone,SL)是一组类胡萝卜素衍生的内酯,拟南芥多丛枝突变体[20]及水稻多分蘖突变体[21]均证实了SL在抑制腋芽生长方面具有保守作用。【本研究切入点】笔者在转基因水稻中获得稳定遗传的半矮化突变体,与已被鉴定的其他水稻矮化植株的表型有较大差异。【拟解决的关键问题】本研究利用转录组学及生物信息学分析确定造成突变体表型异常的主要基因,并利用高效液相色谱-质谱联仪对突变体内的激素进行检测,与转录组测序结果相互验证。矮化成因的解析对克服过度矮化障碍因子及培育抗倒伏高产水稻品种具有理论与应用价值。1 材料与方法
1.1 突变体材料
在水稻日本品种Kitaake(Oryza sativa Japonica )幼胚组培过程中,获得一株半矮化突变体,通过网室自然条件下土壤培养3代之后,未出现性状分离,表型稳定,突变体用sd13 (semi-dwarf 13)表示。1.2 突变体表型分析
挑选发育饱满的野生型和突变体种子于玻璃培养皿中,自来水浸泡,置于30℃恒温培养箱中暗发芽1 d。第2天用湿润的滤纸代替自来水继续培养,第4天转移到光照培养箱中,光照条件为白天16 h 28℃/黑夜8 h 22℃。第5天转移到土壤中,水稻土和蛭石比例为2﹕1,培养白盆的尺寸54 cm×35 cm×12 cm(长×宽×高),土壤重量为4 kg,每天上午和下午各浇一次水。培养地点是中国农业科学院资源区划所东区网室。在水稻生长至抽穗期时,用直尺量取地上部高度(5次生物学重复)。选取5株水稻进行有效分蘖数统计。利用立体式显微镜(LECIA M165 FC,德国)观察水稻小穗及花内部构造的差异。小穗采自穗的同一位置,然后用镊子将外稃和内稃分开露出雄蕊和雌蕊。选取穗茎节间的茎,用刀片刮至茎透明为止,刮的时候需滴两滴蒸馏水以确保茎保持湿润,然后利用光学显微镜(LEICA DM6 B,德国)观察细胞大小,并统计30个细胞的长度。在水稻成熟之后,选取10株统计其结实率。然后用游标卡尺测量种子长和宽,重复6次。
1.3 转录组测序
对苗期生长3周的水稻进行外源GA处理,喷施20 mL浓度为1.5×10-4 mol·L-1的GA3,3 d喷施1次,共3次,于喷施后第8天选取水稻第2片叶进行转录组测序,所有处理均按3次生物学重复进行。首先用Easy Pure Plant RNA(北京全式金)试剂盒提取RNA,然后用带有Oligo(dT)的磁珠富集mRNA,加入fragmentation buffer将mRNA打断成短片段,随后,用六碱基随机引物(random hexamers)以mRNA为模板进行反转录合成一链cDNA,再加入缓冲液、dNTPs和DNA聚合酶Ⅰ合成二链cDNA。其次利用AMPure XP beads纯化双链cDNA,对纯化后的双链cDNA进行末端修复、加A、加接头。通过AMPure XP beads对双链cDNA进行片段大小选择,最后进行PCR扩增以构建cDNA文库。使用Agilent 2100对文库的插入片段大小进行检测,文库质检合格后,利用Illumina高通量测序平台进行测序。使用DEseq和DEseq2检测转基因与野生型株系之间的差异表达基因,差异基因筛选的条件为:差异倍数≥2和Q值(或FDR)≤0.01。然后使用Gene Ontology(GO富集)[22]和KEGG富集[23]对差异基因进行分析。
1.4 突变体外源GA敏感性检测
土培水稻外源喷施GA处理与转录组测序的水稻一致,在转录组测序取样之前,观察并记录表型差异。水培水稻外源GA处理是待水稻在恒温培养箱中发芽3 d后,进行水培,营养液(Hoagland,pH5.8)3 d更换1次,并在营养液中分别添加浓度为1.5×10-4、3×10-5和6×10-6 mol·L-1的GA3,10 d后,观察突变体的表型。1.5 qRT-PCR分析
待水稻生长至孕穗期,进行外源GA处理,与转录组测序取样之前的处理方式相同,之后选取花、穗茎节间的茎及第2片叶提取RNA。首选用液氮进行研磨,之后用Easy Pure Plant RNA试剂盒(北京全式金)进行RNA提取。将提取后的RNA用浓度测定仪(Nanodrop 2000,美国)测定其浓度。利用反转录试剂盒Transscript One-step gDNA Removal and cDNA Synthesis SuperMix(北京全式金)将1.5 μg RNA反转录为cDNA。使用ChamQTM Universal SYBR qPCR Master Mix(南京诺唯赞)进行荧光定量PCR,总反应溶液体积为20 μL,包括10 μL 2×ChamQTM Universal SYBR qPCR Master Mix、0.4 μL正反引物(表1)、2 μL cDNA模板和7.2 μL无RNA酶水,重复3次。使用ABI 7500(美国)荧光定量PCR仪进行定量PCR扩增,PCR扩增条件为94℃ 30 s;95℃ 10 s,60℃ 30 s,40个循环。以β-Actin 作为内参,扩增结束后使用2-ΔΔCt公式计算相对表达量。Table 1
表1
表1荧光定量PCR引物
Table 1
| 基因 Gene | 引物序列 Primer sequence (5′-3′) |
|---|---|
| β-Actin | F: CTGACAGGATGAGCAAGGAG R: GGCAATCCACATCTGCTGGA |
| OsGA13ox | F: AGAAGTGGAGAAAAGACTACGG R: CAATGATCTTTCTCTGGTGTGC |
| D14 | F: AGAAAGAGAGAGAAGAAGCGAG R: CGCGCTCCCCTTTTATATACTA |
新窗口打开|下载CSV
1.6 赤霉素含量检测
1.6.1 赤霉素的提取 称取新鲜水稻叶片约1 g于液氮中研磨粉碎,加入10 mL异丙醇/盐酸提取缓冲液和8 μL 1 μg·mL-1内标溶液,4℃震荡30 min;随后加入20 mL二氯甲烷,4℃震荡30 min;之后13 000 r/min(4℃)离心5 min,取下层有机相,避光用氮吹仪(杭州美欧)吹干有机相,用400 μL甲醇(0.1%甲酸)溶解,0.22 μm滤膜过滤,最后用高效液相色谱(Aglient1290,美国Aglient)-质谱(SCIEX-6500Qtrap,美国AB)联仪检测。1.6.2 标准曲线绘制 以甲醇(0.1%甲酸)为溶剂配制梯度为0.1、0.2、0.5、2.0、5.0、20.0、50.0和200.0 ng·mL-1的GA1和GA4标准溶液,并加入终浓度为20.0 ng·mL-1的内标溶液。每个浓度2个重复。
1.6.3 液相和质谱条件 液相条件:色谱柱采用poroshell 120 SB-C18反相色谱柱(2.1 mm×150 mm,2.7 μm);柱温:30℃;流动相:A﹕B=(甲醇/0.1%甲酸)﹕(水/0.1%甲酸);洗脱梯度:0—1 min,A=20%;1—9 min,A递增至80%;9—10 min,A=80%;10—10.1 min,A递减至20%;10.1—15min,A=20%;进样体积:2 μL。
质谱条件:气帘气为15 psi;喷雾电压:4 500 V;雾化气压力:65 psi;辅助气压力:70 psi;雾化温度:400℃。
1.7 统计分析
利用Excel 2010和SPSS 12.0软件对实验数据进行统计分析,用Duncan法进行差异显著性检验。2 结果
2.1 半矮化突变体sd13 的表型差异
水稻半矮化突变体的表型发育异常。与野生型相比,突变体株高变矮且分蘖数增多(图1-A)并出现包穗现象,穗不能完全伸出剑叶鞘(图1-B和图1-C);突变体种子变小(图1-D)且小穗发育异常,外稃与内稃上的表皮毛消失(图2-A和图2-C);花发育延迟,雄蕊变小(图2-E和图2-G);茎节间细胞长度减小(图3)。统计分析显示突变体株高减少56.29%,且各个节间长度均减小(图4-A),差异极显著;突变体有效分蘖数比WT高出47.44%(图4-B),方差分析差异极显著;突变体结实率低,仅为WT的12.62%(图4-C);种子长度和宽度均减小,经方差分析差异显著(P <0.05)(图4-D和图4-E)。利用显微镜对突变体的茎纵切片统计分析,发现突变体的细胞长度减少约23%,差异极显著(P <0.01)(图4-F)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1野生型和突变体的表型差异
A:孕穗期野生型和突变体形态;B:野生型小穗(比例尺,2 cm);C:突变体小穗(比例尺,2 cm);D:野生型和突变体成熟种子大小
Fig. 1Phenotypic differences between the WT and the sd13
A: Morphological characteristics at booting stage; B: Spikelet in the WT (Scale bar, 2 cm); C: Spikelet in the sd13 (Scale bar, 2 cm); D: Mature seed size
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2水稻野生型和突变体的微观表型
A—D:GA喷施前后野生型和突变体水稻小穗形态(比例尺,0.4 mm);E—H:GA喷施前后野生型和突变体水稻花的特征(比例尺,0.4 mm)
Fig. 2Microscopic phenotypes of the WT and the sd13
A-D: The spikelet morphology of the WT and the sd13 before and after the application of GA3 (Scale bar, 0.4 mm); E-H: The flower characteristic of the WT and the sd13 before and after the application of GA3(Scale bar, 0.4 mm)
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3喷施GA3前后野生型和突变体水稻穗茎节间茎细胞大小(比例尺,25 μm)
Fig. 3The apical internode cell size of the WT and the sd13 before and after spraying GA (Scale bar, 25 μm)
图4
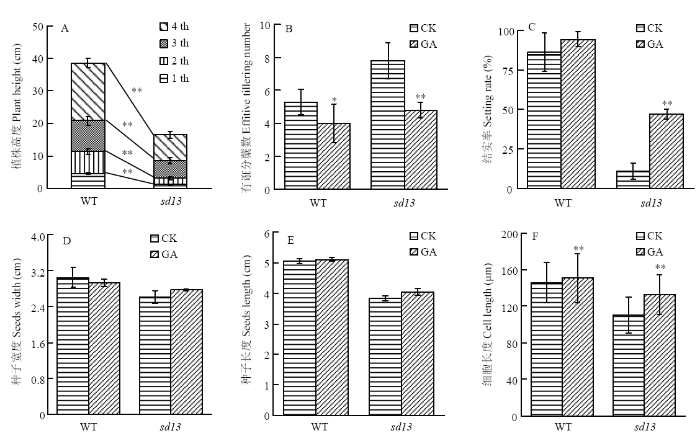 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4野生型和突变体水稻表型及茎细胞长度统计分析
**代表差异极显著,P <0.01;*代表差异显著,P <0.05。下同
Fig. 4Statistical analyses of phenotype and stem cell lengths of the WT and the sd13
** indicates a extremely significant difference, P <0.01; * indicates a significant difference, P <0.05. The same as below
2.2 突变体对外源GA敏感性检测
为了深入探究突变体遗传突变的分子应答与赤霉素合成基因之间是否存在依存关系,对水稻进行外源喷施GA3处理,结果表明,经赤霉素处理的突变体的株高、分蘖、包穗、结实率、种子大小、表皮毛和茎细胞长度均得到一定程度的恢复(图2-C和图2-D,图4-B—图4-F,图5-A)。与喷施GA3前相比,野生型和突变体在喷施GA3之后,有效分蘖数均减小,差异均达显著水平。喷施GA3后,与野生型相比,虽然突变体结实率差异仍然极显著,但是与喷施GA3之前相比显著增高(P <0.01)。种子的长与宽在喷施GA3后均得到一定程度的恢复,但是与喷施GA前差异不明显。在喷施GA3后,突变体表皮毛出现了一定的恢复,但是没有测试具体数量指标。突变体在喷施GA3后,其茎细胞长度显著变长(P <0.01)。水培的突变体经过GA3处理也出现相似的现象(图5-B),结果显示,在营养液中添加不同浓度GA3之后,突变体株高均增加,其中以低浓度处理(6×10-6 mol·L-1)的突变体恢复最佳,说明突变体极有可能是GA敏感型突变体。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5GA处理后突变体的表型变化
A:土培突变体喷施GA后的表型变化;B:营养液添加不同浓度GA后突变体表型变化,GA①、GA②和GA③分别代表GA3浓度为1.5×10-4、3.0×10-5和6.0×10-6 mol·L-1
Fig. 5Phenotypic changes of the sd13 after GA treatment
A: Phenotypic changes after application of GA by soil culture mutants; B: Mutant phenotypic changes after adding different concentrations of GA in nutrient solution, the GA①, GA② and GA③ represents the concentration of GA3 is 1.5×10-4, 3.0×10-5 and 6.0×10-6 mol·L-1, respectively
2.3 基于转录组学分析的差异基因表达特征
为了深入探究sd13 遗传突变的分子机制,将培养3周的水稻进行转录组测序。测序结果显示,与WT相比,突变体共有631个差异基因,其中326个上调,305个下调(图6-B)。GO(Gene Ontology)富集显示,差异基因主要富集在电子转移过程,涉及电子转移的基因有50个(图6-C)。KEGG(Kyoto Encyclopedia of Genes and Genomes)富集显示差异基因主要参与糖酵解和磷酸戊糖等代谢途径(图6-D)。说明半矮化突变体的某一个或者多个基因突变导致了下游参与电子转移基因的异常表达,异常表达的基因极有可能是一些与糖代谢有关的活性酶。图6
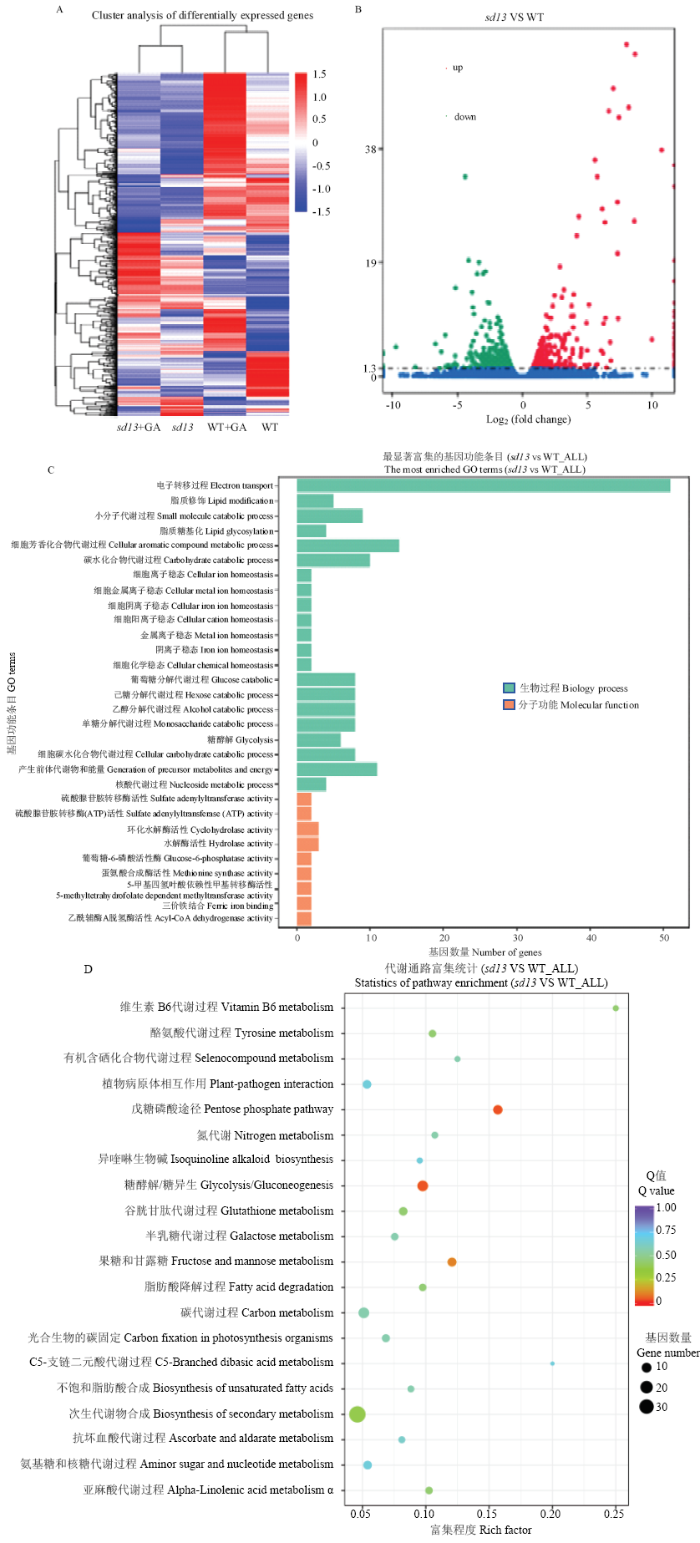 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6转录组测序结果
A:差异基因聚类图。每一行表示一个基因,每一列表示一个样,颜色从红到蓝,表示表达水平从大到小,颜色相近聚类区内的基因表达模式相近,说明这些基因可能具有相似的功能或参与调控同一条代谢通路;B:差异基因火山图。差异表达显著的基因用红点(上调)和绿点(下调)表示,差异表达不显著的基因用蓝点表示;C:差异基因GO富集图。绿色代表生物过程,橙色代表分子功能;D:差异基因KEGG富集散点图。点的大小表示此pathway中差异表达基因个数多少,而点的颜色对应于不同的Qvalue值,值越小说明富集越显著
Fig. 6Transcriptome sequencing results
A: Differential gene clustering. Each row represents a gene and each column represents a sample. The color is from red to blue, indicating that the expression level is from large to small, and the expression patterns of genes in similar clusters are similar, indicating that these genes may have similar functions or participate in regulation of the same metabolic pathway; B: Differential gene volcano map. Genes with significant differential expression are indicated by red dot (up-regulation) and green dot (down-regulation), and genes with insignificant differential expression are represented by blue dots; C: Differential gene GO enrichment map. Green represents biological processes and orange represents molecular function; D: Differential gene KEGG enrichment scatter plot. The size of the dot indicates the number of differentially expressed genes in the pathway, and the color of the dot corresponds to a different Qvalue. The smaller the value, the more significant the enrichment
为进一步分析挖掘差异表达基因的累积分布特征,对突变体中喷施GA前后得到的8个表达不一致的差异基因进行了分析(表2),结果表明,这8个差异基因对外源GA较为敏感。利用生物信息学分析,发现有一个与赤霉素合成相关的OsGA13ox ,在突变体中表达明显上调,而喷施GA后表达出现下调。OsGA13ox 在植物体内负责将GA12转化为GA53,而GA12和GA53又分别转化为GA4和GA1,GA4的活性强于GA1,暗示半矮化突变体极有可能是OsGA13ox 表达异常引起的。对突变体的花、茎和叶提取RNA进行qRT-PCR验证,结果(图7)显示,OsGA13ox 在突变体各器官中的表达均高于野生型,尤其在茎中的表达量较高且与WT相比差异极显著;OsGA13ox 在突变体叶中的表达量也较高,与WT相比差异极显著;但在花中的差异不明显。对突变体进行GA处理之后,OsGA13ox 的表达量出现不同程度的下降,其中在茎和叶中与喷施GA之前相比差异极显著。说明突变体是由OsGA13ox 表达异常引起的,且OsGA13ox 对外源GA敏感。
Table 2
表2
表2突变体喷施GA前后表达相反的基因
Table 2
| 基因 Gene | 功能 Function |
|---|---|
| Os01g0115600 | 蛋白激酶功能,结合ATP Kinase function, binding with ATP |
| Os01g0781700 | ATP结合功能ATP binding |
| Os10g0463800 | 功能未知Unknown function |
| Os03g0332100 | GA13ox,催化GA12转化为GA53,决定GA4和GA1的比例[24] GA13ox, Converts GA12 into GA53, determines the ratio of GA4 and GA1[24] |
| Os04g0223500 | 黄素腺嘌呤二核苷酸,单加氧酶活性 FAD, monooxygenase activity |
| Os04g0606000 | 酰基转移酶活性 Transferase activity, transferring acyl groups |
| Os06g0364500 | 多糖结合功能Polysaccharide binding |
| Os12g0231700 | 推测是转位子蛋白Transposon protein, putative |
新窗口打开|下载CSV
图7
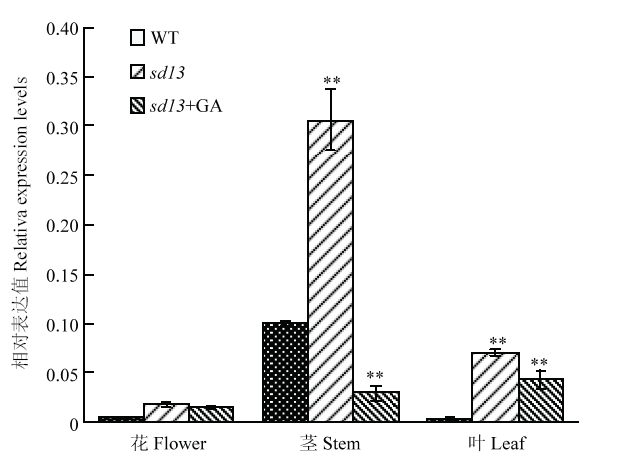 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7OsGA13ox 在突变体各器官中的相对表达量
Fig. 7Relative expression of the OsGA13ox gene in flowers, stem and leaf of the sd13
2.4 水稻突变体GA的富集特征
为明确突变体中OsGA13ox 表达上调是否引起水稻中GA4含量变化,用高效液相和质谱联仪对突变体中的GA1和GA4进行检测分析。结果表明,与WT相比,突变体中的GA1含量无显著变化,但活性更强的GA4减少了94.9%(图8),差异极显著(P <0.01),与预测结果一致,说明半矮化突变体是由GA4减少引起的。图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8水稻野生型和突变体中GA的检测结果
A:野生型水稻GA4检测峰图;B:突变体水稻GA4检测峰图;C:野生型和突变体水稻中GA1含量;D:野生型和突变体水稻中GA4含量
Fig. 8GA test results in the WT and the sd13
A: The GA4 peak map of WT; B: The GA4 peak map of the sd13 ; C: GA1 content in the WT and the sd13 ; D: GA4 content in the WT and the sd13
2.5 GA对水稻分蘖的影响
赤霉素(GA)和独脚金内酯(SL)通常参与调控植物的枝条分枝或分蘖[25],并且GA和SL之间存在着竞争DELLA蛋白结合位点的关系[26]。转录组测序分析韦恩图显示,野生型和突变体有12个共同对GA应答的基因(图9-A),利用GO注释获得这12个基因的功能(表3)。有趣的是,最后3个基因Os03g0203200 、Os03g0251350 和Os03g0856500 在突变体中也下调表达。Os03g0203200 是D14蛋白,作为独脚金内酯(SL)的特异性受体,可与赤霉素受体GID1竞争DELLA蛋白结合位点,调控植物的生长发育[27]。qRT-PCR结果显示(图9-B),与野生型相比,D14 在突变体中下调表达,差异极显著。野生型和突变体喷施GA后,D14 均上调表达,其中野生型D14 上调表达较高,差异极显著(P <0.01),而突变体中虽然也出现了上调表达,但与喷施GA之前相比,差异不显著。Table 3
表3
表3野生型和突变体共表达的12个GA应答基因
Table 3
| 基因 Gene | 功能 Function |
|---|---|
| Novel00727 | Lnc RNA,可能参与调控植物的表型Lnc RNA, probably participate in regulating the phenotype of plants |
| Os01g0191200 | 磷酸激酶活性 Acid phosphatase activity |
| Os01g0198900 | 功能未知 Unknow function |
| Os01g0368900 | 谷胱甘肽-二硫化物氧化还原酶活性 Has a glutathione-disulfide oxidoreductase activity |
| Os01g0501800 | 参与光合系统Ⅱ的组装和稳定 Involved in photosystem II assembly and stabilization |
| Os01g0600900 | 光受体,捕捉光信号 The light-harvesting complex functions as a light receptor |
| Os01g0948200 | 转录因子 DNA-binding transcription factor activity |
| Os03g0159900 | 功能未知 Unknow function |
| Os03g0196250 | 功能未知 Unknow function |
| Os03g0203200 | D14蛋白参与独脚金内酯(SL)信号通路;与赤霉素竞争DELLA结合位点[27] Involved in strigolactone signaling pathway; Competition with GA for DELLA binding sites[27] |
| Os03g0251350 | 转录因子,调控长日照开花[28] Probable transcription factor involved in the regulation of flowering time under long day[28] |
| Os03g0856500 | 与核糖体小亚基结合,负调控蛋白翻译 Ribosomal small subunit binding, negative regulation of translational elongation |
新窗口打开|下载CSV
图9
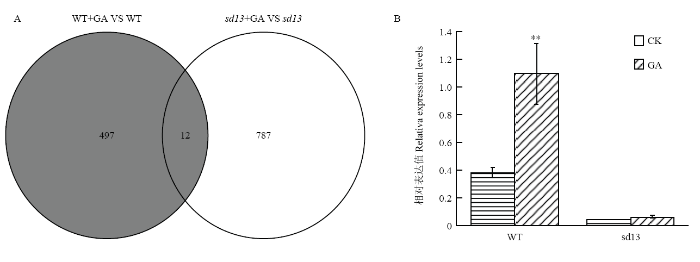 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9差异基因韦恩图及D14 在野生型和突变体茎中的相对表达量
Fig. 9Venn diagram of differential genes and relative expression levels of D14 in the stems of the WT and the sd13
3 讨论
3.1 OsGA13ox 异常表达使GA4含量减少是突变体半矮化的原因
许多研究表明水稻的矮化及半矮化与赤霉素合成或信号转导受阻有关,ITOH等[29]研究表明水稻OsGA3ox2 突变是d18 突变体矮化的原因,而LO等[30]发现超表达OsGA2ox 水稻也会出现矮化等现象。这些现象产生的原因是GA3ox氧化酶是控制合成GA1和GA4的关键因子,而GA2ox氧化酶可以使GAs钝化失活[31]。本研究发现一株半矮化突变体,其株高变矮,分蘖增多,且花发育异常,结实率低。该突变体对外源GA敏感,并且喷施GA3能使表型得到一定程度的恢复,说明该突变体为功能缺失型突变体。赤霉素的合成主要受内根-贝壳杉烯合成酶(KS)、内根-贝壳杉烯19-氧化酶(KO)、GA-13-羟化酶(GA13ox)及GA-20-氧化酶(GA20ox)等酶的统一调控(图10)。已有研究表明,在拟南芥[32,33]和水稻[34]中,GA1的生物活性低于GA4,而GA13ox作为调控GA4和GA1的枢纽,对植物体内赤霉素的相对稳定具有十分重要的作用。MAGOME等[24]发现OsGA13ox 在水稻中超表达可导致水稻半矮化,而GA13突变体却表现出植株增高,穗茎节间变长。造成这种现象的原因是GA13ox 超表达致使活性更高的GA4富集减少,ga13 突变体则相反,由此可说明GA13ox 在控制植物体内GA4和GA1的相对稳定发挥着重要的作用。本研究转录组测序和qRT-PCR结果显示,与WT相比,半矮化突变体OsGA13ox 表达上调,使活性更高的GA4含量减少,破坏GA4和GA1之间的相对平衡,这与前人研究结论相似,再次证明了GA13ox对植物体内赤霉素的相对平衡是至关重要的。图10
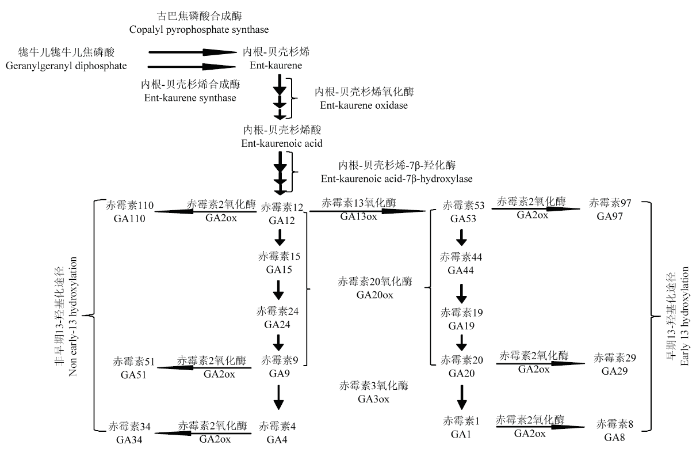 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10植物体内赤霉素合成途径[31]
Fig. 10Gibberellin synthesis pathway in plants[31]
3.2 sd13 的突变基因可能是能够抑制OsGA13ox 基因表达的转录因子
通常基因的转录水平受转录因子的调节,ZHANG等[35]研究发现水稻转录因子OsWRKY71是赤霉素信号转导的抑制子,而OsGAMYB是GA诱导型转录激活因子,其启动子中含有W盒功能性保守基序TGAC,OsWRKY71可以与该基序特异性结合而抑制OsGAMYB的激活,导致GA信号的转导受到阻碍。ZHANG等[36]又发现OsWRKY24 也可以抑制赤霉素信号的转导。GUO等[37]研究表明拟南芥GD1 是编码具有抑制活性的含B3结构域的转录因子,gd1 突变体致使拟南芥出现矮化等现象,同时发现赤霉素(GA)失活基因OsGA2ox3 的表达显著增加;并且验证了gd1 突变体中OsGA20ox1 、OsGA20ox2 和OsGA3ox2 3种GA生物合成基因表达减少,导致内源GA4富集明显降低。外源GA处理不仅可以诱导拟南芥GD1 表达,而且能够部分缓解gd1 突变体的矮化程度,说明GD1 能够抑制植物体内赤霉素失活基因的表达。因此,推测本研究中OsGA13ox 的异常表达可能与具有抑制活性的转录因子突变有关。利用植物转录因子数据库(http://planttfdb.cbi.pku.edu.cn)对OsGA13ox 转录起始位点前500位和后100位启动子序列进行基序预测,结果如表4所示。除此之外,据报道,茉莉酸(JA)可以通过强烈抑制GA20ox 和GA13ox 的转录拮抗GA的合成[38],说明高水平JA可能拮抗茎中GA的生物合成。所以与OsGA13ox 相关的转录因子是否突变有待于进一步深入研究,并且探索茉莉酸合成基因或其他基因是否与OsGA13ox 的异常表达有关。Table 4
表4
表4OsGA13ox 启动子序列中的基序预测结果
Table 4
| 基因 Gene | 基序 Motif | 家族 Family | 功能 Function |
|---|---|---|---|
| Os03g0224200 | CAGTATCTTT | ARR-B | 参与细胞分裂素信号的转导[39] Participation in the transduction of cytokinin signaling[39] |
| Os06g0130600 | CACTCACATTCTCTCTGCACA | BBR-BPC | 调控大麦BKn3 的表达[40] Regulate the expression of BKn3 gene in barley[40] |
| Os10g0419200 | TTTTTTTTTTTG | C2H2 | 促进植物营养生长到生殖生长的转变[41] Promote the transformation of plant vegetative growth to reproductive growth[41] |
| Os07g0236700 | GTATCTTTTTTTTTTTGCGCC | Dof[42] | 可能参与碳水化合物代谢基因的光调节,植物防御机制 Possible involvement in light regulation of carbohydrate metabolism genes, plant defense mechanisms |
| Os03g0821200 | AACAAAGCTAAAACATAAGGG | Dof[42] | 参与种子萌发,发芽后糊粉中的赤霉素反应等基因的调控 Involving in seed germination, regulation of gibberellin reactions in aleurone after germination |
| Os03g0324300 | GTCTAGATTCATTAC | EIL | 乙烯信号响应的正向调控因子[43] Positive regulator of ethylene signal response[43] |
| Os03g0324200 | GTCTAGATTCATTAC | EIL | 在植物受伤时,参与乙烯信号的转导[44] Participation in the transduction of ethylene signals when plants are injured[44] |
| Os09g0490200 | TAATGAATCTA | EIL | 参与乙烯信号转导 Participation in ethylene signal transduction |
| Os11g0681200 | GAAACACGCGCGCGC | FAR1 | 参与抵抗疾病[45] Enhance the resistance of disease[45] |
| Os02g0680700 | TAGATATTA | MYB_related[46] | 功能未知 Unknown function |
| Os08g0157600 | TAGATATTA | MYB_related[46] | 功能未知 Unknown function |
新窗口打开|下载CSV
3.3 GA可能通过调控D14 的表达间接调控水稻分蘖
拟南芥ga1-3 突变体拥有更多的分枝,超表达OsGA2ox 的水稻也表现出分蘖增多[30],而水稻d61 突变体中赤霉素合成基因上调表达使得分蘖减少[47],说明GA能够抑制水稻分蘖。ITO等[25]已证明外源GA能够使水稻分蘖异常突变体得到恢复。除赤霉素之外,独脚金内酯(SL)也能够抑制枝条分枝或者分蘖[48],且SL的合成受GA信号调控[49]。D14蛋白为SL的特异性受体,二者能够和DELLA蛋白结合形成复合物,从而调控下游基因的表达,间接调节枝条分枝或者分蘖。本研究发现,与WT相比,突变体的D14 下调表达,而D14 作为SL的特异性受体,下调表达会使SL信号减弱,从而使分蘖增多,这与突变体的分蘖增多相符合。外源喷施GA使D14 上调表达,会增强SL信号从而使分蘖减少,而WT喷施GA后分蘖减少也验证了这一猜测。本研究推测GA可能通过D14 的表达间接调控植物体的枝条分枝或分蘖。4 结论
半矮化突变体是由OsGA13ox 异常表达导致GA4富集减少引起的;赤霉素可能通过D14 间接调控水稻的分蘖。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1073/pnas.132266399URL [本文引用: 2]
DOI:10.1038/22307URL [本文引用: 1]
DOI:10.2307/3871176URLPMID:11006334 [本文引用: 1]

Brassinosteroids (BRs) are plant growth-promoting natural products required for plant growth and development. Physiological studies have demonstrated that exogenous BR, alone or in combination with auxin, enhance bending of the lamina joint of rice. However, little is known about the function of endogenous BR in rice or other grass species. We report here the phenotypical and molecular characterization of a rice dwarf mutant, d61, that is less sensitive to BR compared to the wild type. We cloned a rice gene, OsBRI1, with extensive sequence similarity to that of the Arabidopsis BRI gene, which encodes a putative BR receptor kinase. Linkage analysis showed that the OsBRI1 gene is closely linked to the d61 locus. Single nucleotide substitutions found at different sites of the d61 alleles would give rise to amino acid changes in the corresponding polypeptides. Furthermore, introduction of the entire OsBRI1 coding region, including the 5 and 3 flanking sequences, into d61 plants complemented the mutation to display the wild-type phenotype. Transgenic plants carrying the antisense strand of the OsBRI1 transcript showed similar or even more severe phenotypes than those of the d61 mutants. Our results show that OsBRI1 functions in various growth and developmental processes in rice, including (1) internode elongation, by inducing the formation of the intercalary meristem and the longitudinal elongation of internode cells; (2) bending of the lamina joint; and (3) skotomorphogenesis.
[本文引用: 1]
DOI:10.1111/j.1365-313X.2007.03210.xURLPMID:17655651 [本文引用: 1]
Plant architecture is mostly determined by shoot branching patterns. Apical dominance is a well-known control mechanism in the development of branching patterns, but little is known regarding its role in monocots such as rice. Here, we show that the concept of apical dominance can be applied to tiller bud outgrowth of rice. In dwarf10 ( d10 ), an enhanced branching mutant of rice, apical dominance can be observed, but the inhibitory effects of the apical meristem was reduced. D10 is a rice ortholog of MAX4/RMS1/DAD1 that encodes a carotenoid cleavage dioxygenase 8 and is supposed to be involved in the synthesis of an unidentified inhibitor of shoot branching. D10 expression predominantly occurs in vascular cells in most organs. Real-time polymerase chain reaction analysis revealed that accumulation of D10 mRNA is induced by exogenous auxin. Moreover, D10 expression is upregulated in six branching mutants, d3 , d10 , d14 , d17 , d27 and high tillering dwarf ( htd1 ). No such effects were found for D3 or HTD1 , the MAX2 and MAX3 orthologs, respectively, of rice. These findings imply that D10 transcription might be a critical step in the regulation of the branching inhibitor pathway. In addition, we present observations that suggest that FINE CULM1 ( FC1 ), a rice ortholog of teosinte branched 1 ( tb1 ), possibly works independently of the branching inhibitor pathway.
DOI:10.1093/pcp/pci022URLPMID:15659436 [本文引用: 1]
Abstract In this study, we analyzed five tillering dwarf mutants that exhibit reduction of plant stature and an increase in tiller numbers. We show that, in the mutants, axillary meristems are normally established but the suppression of tiller bud activity is weakened. The phenotypes of tillering dwarf mutants suggest that they play roles in the control of tiller bud dormancy to suppress bud activity. However, tillering dwarf mutants show the dependence of both node position and planting density on their growth, which implies that the functions of tillering dwarf genes are independent of the developmental and environmental control of bud activity. Map-based cloning of the D3 gene revealed that it encodes an F-box leucine-trich repeat (LRR) protein orthologous to Arabidopsis MAX2/ORE9. This indicates the conservation of mechanisms controlling axillary bud activity between monocots and eudicots. We suggest that tillering dwarf mutants are suitable for the study of bud activity control in rice and believe that future molecular and genetic studies using them may enable significant progress in understanding the control of tillering and shoot branching.
DOI:10.1016/j.pbi.2004.11.015URLPMID:15653404 [本文引用: 1]
The importance of gibberellin (GA) in vegetative and reproductive development has been known for some time. Recent studies have uncovered new roles of GA in leaf differentiation, photomorphogenesis and pollen-tube growth. Significant contributions to our understanding of GA-regulated morphogenesis include the identification of upstream regulators of GA biosynthesis, the elucidation of the function of GA signaling components, and the isolation of downstream targets. In addition, the mechanisms of interactions between GA and other hormone pathways are beginning to be revealed at the molecular level.
DOI:10.3321/j.issn:1006-687X.2008.04.028URL [本文引用: 1]
赤霉素是一种高效能的广谱植物生长调节剂,能够促进植物的生长发育,具有重要的生物学功能.利用突变体研究赤霉素生物合成途径和信号转导是目前的研究热点.本文对赤霉素的研究历史、合成途径、关键酶基因、作用机理以及最新的研究进展等作一综述.
DOI:10.3321/j.issn:1006-687X.2008.04.028URL [本文引用: 1]
赤霉素是一种高效能的广谱植物生长调节剂,能够促进植物的生长发育,具有重要的生物学功能.利用突变体研究赤霉素生物合成途径和信号转导是目前的研究热点.本文对赤霉素的研究历史、合成途径、关键酶基因、作用机理以及最新的研究进展等作一综述.
DOI:10.1104/pp.104.040279URL [本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.119.2.507URLPMID:9952446 [本文引用: 1]
The Arabidopsis GA3 cDNA was expressed in yeast (Saccharomyces cerevisiae) and the ability of the transformed yeast cells to metabolize ent-kaurene was tested. We show by full-scan gas chromatography-mass spectrometry that the transformed cells produce ent-kaurenoic acid, and demonstrate that the single enzyme GA3 (ent-kaurene oxidase) catalyzes the three steps of gibberellin biosynthesis from ent-kaurene to ent-kaurenoic acid.
DOI:10.1007/s00299-004-0896-6URLPMID:15668792 [本文引用: 1]
We have isolated a severe dwarf transposon ( Ds ) insertion mutant in rice ( Oryza sativa L.), which could be differentiated early in the seedling stage by reduced shoot growth and dark green leaves, and later by severe dwarfism and failure to initiate flowering. These mutants, however, showed normal seed germination and root growth. One of the sequences flanking Ds , rescued from the mutant, was of a chromosome 4-located putative ent -kaurene synthase ( KS ) gene, encoding the enzyme catalyzing the second step of the gibberellin (GA) biosynthesis pathway. Dwarf mutants were always homozygous for this Ds insertion and no normal plants homozygous for this mutation were recovered in the segregating progeny, indicating that the Ds insertion mutation is recessive. As mutations in three recently reported rice GA-responsive dwarf mutant alleles and the dwarf mutation identified in this study mapped to the same locus, we designate the corresponding gene OsKS1 . The osks1 mutant seedlings were responsive to exogenous gibberellin (GA 3 ). OsKS1 transcripts of about 2.3 kb were detected in leaves and stem of wild-type plants, but not in germinating seeds or roots, suggesting that OsKS1 is not involved in germination or root growth. There are at least five OsKS1 -like genes in the rice genome, four of which are also represented in rice expressed sequence tag (EST) databases. All OsKS1 -like genes are transcribed with different expression patterns. ESTs corresponding to all six OsKS genes are represented in other cereal databases including barley, wheat and maize, suggesting that they are biologically active.
DOI:10.1023/B:PLAN.0000038261.21060.47URL [本文引用: 1]
A rice ( Oryza sativa L.) semi-dwarf cultivar, Tan-Ginbozu ( d35 Tan-Ginbozu), contributed to the increase in crop productivity in Japan in the 1950s. Previous studies suggested that the semi-dwarf...
DOI:10.1038/nature04028URLPMID:16193045 [本文引用: 1]
Gibberellins (GAs) are phytohormones that are essential for many developmental processes in plants. It has been postulated that plants have both membrane-bound and soluble GA receptors; however, no GA receptors have yet been identified. Here we report the isolation and characterization of a new GA-insensitive dwarf mutant of rice, gid1. The GID1 gene encodes an unknown protein with similarity to the hormone-sensitive lipases, and we observed preferential localization of a GID109“green fluorescent protein (GFP) signal in nuclei. Recombinant glutathione S-transferase (GST)09“GID1 had a high affinity only for biologically active GAs, whereas mutated GST09“GID1 corresponding to three gid1 alleles had no GA-binding affinity. The dissociation constant for GA4 was estimated to be around 10-7 M, enough to account for the GA dependency of shoot elongation. Moreover, GID1 bound to SLR1, a rice DELLA protein, in a GA-dependent manner in yeast cells. GID1 overexpression resulted in a GA-hypersensitive phenotype. Together, our results indicate that GID1 is a soluble receptor mediating GA signalling in rice.
DOI:10.1111/j.1365-313X.2003.01990.xURLPMID:14756772 [本文引用: 1]
The phytohormone gibberellin (GA) controls growth and development in plants. Previously, we identified a rice F-box protein, gibberellin-insensitive dwarf2 (GID2), which is essential for GA-mediated DELLA protein degradation. In this study, we analyzed the biological and molecular biological properties of GID2. Expression of GID2 preferentially occurred in rice organs actively synthesizing GA. Domain analysis of GID2 revealed that the C-terminal regions were essential for the GID2 function, but not the N-terminal region. Yeast two-hybrid assay and immunoprecipitation experiments demonstrated that GID2 is a component of the SCF complex through an interaction with a rice ASK1 homolog, OsSkp15. Furthermore, an in vitro pull-down assay revealed that GID2 specifically interacted with the phosphorylated Slender Rice 1 (SLR1). Taken these results together, we conclude that the phosphorylated SLR1 is caught by the SCF GID2 complex through an interacting affinity between GID2 and phosphorylated SLR1, triggering the ubiquitin-mediated degradation of SLR1.
DOI:10.1104/pp.110.161554URLPMID:20921186 [本文引用: 1]
This article highlights recent advances in our understanding of the molecular mechanisms of GA metabolism, transport, perception, and signalling, and the regulatory circuit between the GA pathway and other pathways to control plant growth and development in response to internal and external cues.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1101/gad.256603URLPMID:12815068 [本文引用: 1]
Shoot branching is inhibited by auxin transported down the stem from the shoot apex. Auxin does not accumulate in inhibited buds and so must act indirectly. We show that mutations in the MAX4 gene of Arabidopsis result in increased and auxin-resistant bud growth. Increased branching in max4 shoots is restored to wild type by grafting to wild-type rootstocks, suggesting that MAX4 is required to produce a mobile branch-inhibiting signal, acting downstream of auxin. A similar role has been proposed for the pea gene, RMS1. Accordingly, MAX4 and RMS1 were found to encode orthologous, auxin-inducible members of the polyene dioxygenase family.
DOI:10.1038/nature12870URLPMID:24336200 [本文引用: 1]
Strigolactones (SLs) are a group of newly identified plant hormones that control plant shoot branching. SL signalling requires the hormone-dependent interaction of DWARF 14 (D14), a probable candidate SL receptor, with DWARF 3 (D3), an F-box component of the Skp-Cullin-F-box (SCF) E3 ubiquitin ligase complex. Here we report the characterization of a dominant SL-insensitive rice (Oryza sativa) mutant dwarf 53 (d53) and the cloning of D53, which encodes a substrate of the SCFD3 ubiquitination complex and functions as a repressor of SL signalling. Treatments with GR24, a synthetic SL analogue, cause D53 degradation via the proteasome in a manner that requires D14 and the SCFD3 ubiquitin ligase, whereas the dominant form of D53 is resistant to SL-mediated degradation. Moreover, D53 can interact with transcriptional co-repressors known as TOPLESS-RELATED PROTEINS. Our results suggest a model of SL signalling that involves SL-dependent degradation of the D53 repressor mediated by the D14-D3 complex.
DOI:10.1186/gb-2010-11-2-r14URLPMID:20132535 [本文引用: 1]
GOseq is a method for GO analysis of RNA-seq data that takes into account the length bias inherent in RNA-seq We present GOseq, an application for performing Gene Ontology (GO) analysis on RNA-seq data. GO analysis is widely used to reduce complexity and highlight biological processes in genome-wide expression studies, but standard methods give biased results on RNA-seq data due to over-detection of differential expression for long and highly expressed transcripts. Application of GOseq to a prostate cancer data set shows that GOseq dramatically changes the results, highlighting categories more consistent with the known biology.
DOI:10.1093/nar/gkm882URLPMID:18077471 [本文引用: 1]
KEGG (http://www.genome.jp/kegg/) is a database of biological systems that integrates genomic, chemical and systemic functional information. KEGG provides a reference knowledge base for linking genomes to life through the process of PATHWAY mapping, which is to map, for example, a genomic or transcriptomic content of genes to KEGG reference pathways to infer systemic behaviors of the cell or the organism. In addition, KEGG provides a reference knowledge base for linking genomes to the environment, such as for the analysis of drug-target relationships, through the process of BRITE mapping. KEGG BRITE is an ontology database representing functional hierarchies of various biological objects, including molecules, cells, organisms, diseases and drugs, as well as relationships among them. KEGG PATHWAY is now supplemented with a new global map of metabolic pathways, which is essentially a combined map of about 120 existing pathway maps. In addition, smaller pathway modules are defined and stored in KEGG MODULE that also contains other functional units and complexes. The KEGG resource is being expanded to suit the needs for practical applications. KEGG DRUG contains all approved drugs in the US and Japan, and KEGG DISEASE is a new database linking disease genes, pathways, drugs and diagnostic markers.
DOI:10.1073/pnas.1215788110URLPMID:23319637 [本文引用: 3]
Bioactive gibberellins (GAs) control many aspects of growth and development in plants. GA(1) has been the most frequently found bioactive GA in various tissues of flowering plants, but the enzymes responsible for GA(1) biosynthesis have not been fully elucidated due to the enzymes catalyzing the 13-hydroxylation step not being identified. Because of the lack of mutants defective in this enzyme, biological significance of GA 13-hydroxylation has been unknown. Here, we report that two cytochrome P450 genes, CYP714B1 and CYP714B2, encode GA 13-oxidase in rice. Transgenic Arabidopsis plants that overexpress CYP714B1 or CYP714B2 show semidwarfism. There was a trend that the levels of 13-OH GAs including GA(1) were increased in these transgenic plants. Functional analysis using yeast or insect cells shows that recombinant CYP714B1 and CYP714B2 proteins can convert GA(12) into GA(53) (13-OH GA(12)) in vitro. Moreover, the levels of 13-OH GAs including GA(1) were decreased, whereas those of 13-H GAs including GA(4) (which is more active than GA(1)) were increased, in the rice cyp714b1 cyp714b2 double mutant. These results indicate that CYP714B1 and CYP714B2 play a predominant role in GA 13-hydroxylation in rice. The double mutant plants appear pheno-typically normal until heading, but show elongated uppermost internode at the heading stage. Moreover, CYP714B1 and CYP714B2 expression was up-regulated by exogenous application of bioactive GAs. Our results suggest that GA 13-oxidases play a role in fine-tuning plant growth by decreasing GA bioactivity in rice and that they also participate in GA homeostasis.
DOI:10.1584/jpestics.D18-013URL [本文引用: 2]
[本文引用: 1]
DOI:10.1038/ncomms3613URLPMID:24131983 [本文引用: 3]
Abstract Strigolactones (SLs) are phytohormones that inhibit shoot branching and function in the rhizospheric communication with symbiotic fungi and parasitic weeds. An 02±/0205-hydrolase protein, DWARF14 (D14), has been recognized to be an essential component of plant SL signalling, although its precise function remains unknown. Here we present the SL-dependent interaction of D14 with a gibberellin signalling repressor SLR1 and a possible mechanism of phytohormone perception in D14-mediated SL signalling. D14 functions as a cleavage enzyme of SLs, and the cleavage reaction induces the interaction with SLR1. The crystal structure of D14 shows that 5-hydroxy-3-methylbutenolide (D-OH), which is a reaction product of SLs, is trapped in the catalytic cavity of D14 to form an altered surface. The D14 residues recognizing D-OH are critical for the SL-dependent D14-SLR1 interaction. These results provide new insight into crosstalk between gibberellin and SL signalling pathways.
DOI:10.1007/s00425-015-2426-xURLPMID:26542958 [本文引用: 2]
Main conclusion OsNF-YC2 and OsNF-YC4 proteins regulate the photoperiodic flowering response through the modulation of three flowering-time genes ( Ehd1, Hd3a , and RFT1 ) in rice. Plant NUCLEAR...
DOI:10.1073/pnas.141239398URL [本文引用: 1]
We have cloned two gibberellin (GA) 3 -hydroxylase genes, OsGA3ox1 and OsGA3ox2, from rice by screening a genomic library with a DNA fragment obtained by PCR using degenerate primers. We have used full-scan GC-MS and Kovats retention indices to show function for the two encoded recombinant fusion proteins. Both proteins show 3 -hydroxylase activity for the steps GA20to$GA_1, \> GA_5$to$GA_3, \>GA_{44}$to GA38and GA9to GA4. In addition, indirect evidence suggests that the OsGA3ox1 protein also has 2,3-desaturase activity, which catalyzes the steps GA9to 2,3-dehydro-GA9and GA20to GA5(2,3-dehydro GA20), and 2 -hydroxylase activity, which catalyzes the steps GA1to GA8and GA4to GA34. Molecular and linkage analysis maps the OsGA3ox1 gene to the distal end of the short arm of chromosome 5; the OsGA3ox2 gene maps to the distal end of the short arm of chromosome 1 that corresponds to the D18 locus. The association of the OsGA3ox2 gene with the d18 locus is confirmed by sequence and complementation analysis of three d18 alleles. Complementation of the d18-AD allele with the OxGA3ox2 gene results in transgenic plants with a normal phenotype. Although both genes show transient expression, the highest level for OsGA3ox1 is from unopened flower. The highest level for OsGA3ox2 is from elongating leaves.
DOI:10.1105/tpc.108.060913URL [本文引用: 2]
DOI:10.1007/s00344-015-9546-1URLPMID:4622167 [本文引用: 3]
Gibberellin research has its origins in Japan in the 19th century, when a disease of rice was shown to be due to a fungal infection. The symptoms of the disease including overgrowth of the seedling and sterility were later shown to be due to secretions of the fungusGibberella fujikuroi(now reclassified asFusarium fujikuroi), from which the name gibberellin was derived for the active component. The profound effect of gibberellins on plant growth and development, particularly growth recovery in dwarf mutants and induction of bolting and flowering in some rosette species, prompted speculation that these fungal metabolites were endogenous plant growth regulators and this was confirmed by chemical characterisation in the late 1950s. Gibberellins are now known to be present in vascular plants, and some fungal and bacterial species. The biosynthesis of gibberellins in plants and the fungus has been largely resolved in terms of the pathways, enzymes, genes and their regulation. The proposal that gibberellins act in plants by removing growth limitation was confirmed by the demonstration that they induce the degradation of the growth-inhibiting DELLA proteins. The mechanism by which this is achieved was clarified by the identification of the gibberellin receptor from rice in 2005. Current research on gibberellin action is focussed particularly on the function of DELLA proteins as regulators of gene expression. This review traces the history of gibberellin research with emphasis on the early discoveries that enabled the more recent advances in this field.
DOI:10.1104/pp.117.4.1195URLPMID:9701576 [本文引用: 1]
The gibberellins (GAs) are a complex family of diterpenoid compounds, some of which are potent endogenous regulators of plant growth. As part of a feedback control of endogenous GA levels, active GAs negatively regulate the abundance of mRNA transcripts encoding GA biosynthesis enzymes. For example, Arabidopsis GA4 gene transcripts encode GA 3尾-hydroxylase, an enzyme that catalyzes the conversion of inactive to active GAs. Here we show that active GAs regulate GA4 transcript abundance in a dose-dependent manner, and that down-regulation of GA4 transcript abundance is effected by GA4(the product of 3 -hydroxylation) but not by its immediate precursor GA9(the substrate). Comparison of several different GA structures showed that GAs active in promoting hypocotyl elongation were also active in regulating GA4 transcript abundance, suggesting that similar GA:receptor and subsequent signal transduction processes control these two responses. It is interesting that these activities were not restricted to 3 -hydroxylated GAs, being also exhibited by structures that were not 3 -hydroxylated but that had another electronegative group at C-3. We also show that GA-mediated control of GA4 transcript abundance is disrupted in the GA-response mutants gai and spy-5. These observations define a sensitive homeostatic mechanism whereby plants may regulate their endogenous GA levels.
DOI:10.1073/pnas.87.20.7983URLPMID:2236013 [本文引用: 1]
Twenty gibberellins (GAs) have been identified in extracts from shoots of the Landsberg erecta line of Arabidopsis thaliana by full-scan gas chromatography-mass spectrometry and Kovats retention indices. Eight of them are members of the early-13-hydroxylation pathway (GA53, GA44, GA19, GA17, GA20, GA1, GA29, and GA8), six are members of the early-3-hydroxylation pathway (GA37, GA27, GA36, GA13, GA4, and GA34), and the remaining six are members of the non-3,13-hydroxylation pathway (GA12, GA15, GA24, GA25, GA9, and GA51). Seven of these GAs were quantified in the Landsberg erecta line of Arabidopsis and in the semidwarf ga4 and ga5 mutants by gas chromatography-selected ion monitoring (SIM) using internal standards. The relative levels of the remaining 13 GAs were compared by the use of ion intensities only. In comparison with the Landsberg erecta line, the ga4 mutant had reduced levels of the 3-hydroxy- and 3,13-dihydroxy-GAs, and it accumulated the 13-hydroxy-GAs, except GA53, and the non-3,13-hydroxy-GAs, except GA12. The GA4 gene encodes, therefore, a protein with 3 -hydroxylation activity. The ga5 mutant had reduced levels of the C19-GAs, which indicates that the product of the GA5 gene catalyzes the elimination of C-20 at the aldehyde level. The ga5 mutant also had increased levels of certain C20-GAs, which indicates existence of an additional control, possibly hydroxylation of C-20. The growth-response data, as well as the accumulation of GA9in the ga4 mutant, indicate that GA9is not active in Arabidopsis, but it must be 3 -hydroxylated to GA4to become bioactive. It is concluded that the reduced levels of the 3 -hydroxy-GAs, GA1and GA4, are the cause of the semidwarf growth habit of both mutants.
DOI:10.1105/tpc.106.043729URLPMID:17644730 [本文引用: 1]
GIBBERELLIN INSENSITIVE DWARF1 (GID1) encodes a soluble gibberellin (GA) receptor that shares sequence similarity with a hormone-sensitive lipase (HSL). Previously, a yeast two-hybrid (Y2H) assay revealed that the GID1-GA complex directly interacts with SLENDER RICE1 (SLR1), a DELLA repressor protein in GA signaling. Here, we demonstrated, by pulldown and biomolecular fluorescence complementation (BiFC) experiments, that the GA-dependent GID1-SLR1 interaction also occurs in planta. GA was found to have the highest affinity to GID1 in Y2H assays and is the most effective form of GA in planta. Domain analyses of SLR1 using Y2H, gel filtration, and BiFC methods revealed that the DELLA and TVHYNP domains of SLR1 are required for the GID1-SLR1 interaction. To identify the important regions of GID1 for GA and SLR1 interactions, we used many different mutant versions of GID1, such as the spontaneous mutant GID1s, N- and C-terminal truncated GID1s, and mutagenized GID1 proteins with conserved amino acids replaced with Ala. The amino acid residues important for SLR1 interaction completely overlapped the residues required for GA binding that were scattered throughout the GID1 molecule. When we plotted these residues on the GID1 structure predicted by analogy with HSL tertiary structure, many residues were located at regions corresponding to the substrate binding pocket and lid. Furthermore, the GA-GID1 interaction was stabilized by SLR1. Based on these observations, we proposed a molecular model for interaction between GA, GID1, and SLR1.
DOI:10.1104/pp.103.034967URLPMID:15047897 [本文引用: 1]
The molecular mechanism by which GA regulates plant growth and development has been a subject of active research. Analyses of the rice (Oryza sativa) genomic sequences identified 77 WRKY genes, among which OsWRKY71 is highly expressed in aleurone cells. Transient expression of OsWRKY71 by particle bombardment specifically represses GA-induced Amy32b alpha-amylase promoter but not abscisic acid-induced HVA22 or HVA1 promoter activity in aleurone cells. Moreover, OsWRKY71 blocks the activation of the Amy32b promoter by the GA-inducible transcriptional activator OsGAMYB. Consistent with its role as a transcriptional repressor, OsWRKY71 is localized to nuclei of aleurone cells and binds specifically to functionally defined TGAC-containing W boxes of the Amy32b promoter in vitro. Mutation of the two W boxes prevents the binding of OsWRKY71 to the mutated promoter, and releases the suppression of the OsGAMYB-activated Amy32b expression by OsWRKY71, suggesting that OsWRKY71 blocks GA signaling by functionally interfering with OsGAMYB. Exogenous GA treatment decreases the steady-state mRNA level of OsWRKY71 and destabilizes the GFP:OsWRKY71 fusion protein. These findings suggest that OsWRKY71 encodes a transcriptional repressor of GA signaling in aleurone cells.
DOI:10.1007/s11103-009-9463-4URLPMID:19199048 [本文引用: 1]
Abscisic acid (ABA) and gibberellins (GAs) control several developmental processes including seed maturation, dormancy, and germination. The antagonism of these two hormones is well-documented. However, recent data from transcription profiling studies indicate that they can function as agonists in regulating the expression of many genes although the underlying mechanism is unclear. Here we report a rice WRKY gene, OsWRKY24 , which encodes a protein that functions as a negative regulator of both GA and ABA signaling. Overexpression of OsWRKY24 via particle bombardment-mediated transient expression in aleurone cells represses the expression of two reporter constructs: the β-glucuronidase gene driven by the GA-inducible Amy32b α-amylase promoter ( Amy32b - GUS ) and the ABA-inducible HVA22 promoter ( HVA22 - GUS ). OsWRKY24 is unlikely a general repressor because it has little effect on the expression of the luciferase reporter gene driven by a constitutive ubiquitin promoter ( UBI - Luciferase ). As to the GA signaling, OsWRKY24 differs from OsWRKY51 and 6171, two negative regulators specifically function in the GA signaling pathway, in several ways. First, OsWRKY24 contains two WRKY domains while OsWRKY51 and 6171 have only one; both WRKY domains are essential for the full repressing activity of OsWRKY24. Second, binding of OsWRKY24 to the Amy32b promoter appears to involve sequences in addition to the TGAC cores of the W-boxes. Third, unlike OsWRKY71, OsWRKY24 is stable upon GA treatment. Together, these data demonstrate that OsWRKY24 is a novel type of transcriptional repressor that inhibits both GA and ABA signaling.
DOI:10.1111/tpj.2013.75.issue-3URL [本文引用: 1]
DOI:10.1111/tpj.12058URLPMID:23190261 [本文引用: 1]
Hormones play pivotal roles in regulating plant development, growth, and stress responses, and cross-talk among different hormones fine-tunes various aspects of plant physiology. Jasmonic acid (JA) is important for plant defense against herbivores and necrotic fungi and also regulates flower development; in addition, Arabidopsis mutants over-producing JA usually have stunted stems and wound-induced jasmonates suppress Arabidopsis growth, suggesting that JA is also involved in stem elongation. Gibberellins (GAs) promote stem and leaf growth and modulate seed germination, flowering time, and the development of flowers, fruits, and seeds. However, little is known about the interaction between the JA and GA pathways. Two calcium-dependent protein kinases, CDPK4 and CDPK5, are important suppressors of JA accumulation in a wild tobacco species, Nicotiana attenuata. The stems of N.attenuata silenced in CDPK4 and CDPK5 (irCDPK4/5 plants) had dramatically increased levels of JA and exhibited stunted elongation and had very high contents of secondary metabolites. Genetic analysis indicated that the high JA levels in irCDPK4/5 stems accounted for the suppressed stem elongation and the accumulation of secondary metabolites. Supplementation of GA3 to irCDPK4/5 plants largely restored normal stem growth to wild-type levels. Measures of GA levels indicated that over-accumulation of JA in irCDPK4/5 stems inhibited the biosynthesis of GAs. Finally, we show that JA antagonizes GA biosynthesis by strongly inhibiting the transcript accumulation of GA20ox and possibly GA13ox, the key genes in GA production, demonstrating that high JA levels antagonize GA biosynthesis in stems.
DOI:10.1016/j.gene.2006.06.020URLPMID:16919402 [本文引用: 2]
We identified four histidine kinase (HK) genes of a cytokinin receptor family, two histidine-containing phosphotransmitter (HPt) genes, thirteen A-type response regulator (RR) genes and six B-type RR genes in the rice genome. The HK genes ( OHK2, OHK3, OHK4 and OHK5 for Oryza sativa HK), the HPt genes ( OHP1 and OHP2 for O. sativa HPt) and the B-type RR genes ( ORR1, ORR2, ORR3, ORR4 and ORR6 for O. sativa RR) except one ( ORR5) showed expression in various organs. ORR5 was expressed in callus and flower. Three A-type RR genes ( OsRR4, OsRR9 and OsRR10 for O. sativa RR) showed cytokinin-induced expression, and three ( OsRR8, OsRR12 and OsRR13) showed expression in flower. We also identified two other genes named OHK1 and CHARK ( CHASE domain Receptor-like serine/threonine Kinase). OHK1 encodes an HK similar to Arabidopsis CKI1, which is involved in female gametophyte development. CHARK encodes a protein with an extracellular cytokinin-perceiving CHASE domain and a cytoplasmic serine/threonine kinase domain which are connected with a single transmembrane domain. The presence of all four gene families and CHARK in the rice genome suggests that a cytokinin signal is transduced by the phosphotransfer mechanism as is the case in Arabidopsis, and that rice may have an additional novel signalling pathway involving serine/threonine phosphorylation.
DOI:10.1046/j.1365-313X.2003.01767.xURLPMID:12795701 [本文引用: 2]
In the dominant mutant Hooded (K) , the barley gene BKn3 is overexpressed as a result of a duplication of 305 bp in intron IV. When fused to a cauliflower mosaic virus 35S minimal promoter, the 305 bp element activates gene expression in tobacco, as does a 655 bp BKn3 promoter sequence. Both DNA fragments contain a (GA) 8 repeat (GA/TC) 8 . A one-hybrid screen using the 305 bp element as the DNA target led to the cloning of the barley b recombinant (BBR) protein, which binds specifically to the (GA/TC) 8 repeat. BBR is nuclear targeted and is a characterized nuclear localization signal (NLS) sequence, a DNA-binding domain extended up to 90 aa at the C-terminus and a putative N-terminal activation domain. The corresponding gene has no introns and is ubiquitously expressed in barley tissues. In co-transfection experiments, BBR activates (GA/TC) 8 -containing promoters, and its overexpression in tobacco leads to a pronounced leaf shape modification. BBR has properties of a GAGA-binding factor, but the corresponding gene has no sequence homology to Trl and Psq of Drosophila , which encode functionally analogous proteins. In Arabidopsis , (GA/TC) 8 repeats occur particularly within 1500 bp upstream of gene start codons included in some homeodomain genes of different classes. The data presented suggest that expression of the barley BKn3 is regulated, at least in part, by the binding of the transcription factor BBR to GA/TC repeats.
DOI:10.1073/pnas.0806019105URL [本文引用: 2]
Transition from the vegetative phase to reproductive phase is a crucial process in the life cycle of higher plants. Although the molecular mechanisms of flowering regulation have been extensively characterized in a number of plant species, little is known regarding how the transition process initiates. Here, we show that the Rice Indeterminate 1 (RID1) gene acts as the master switch for the transition from the vegetative to reproductive phase. RID1 encodes a Cys-2/His-2-type zinc finger transcription factor that does not have an ortholog in Arabidopsis spp. A RID1 knockout (rid1), mutated by T-DNA insertion, never headed after growing for >500 days under a range of growth conditions and is thus referred to as a never-flowering phenotype. This mutation-suppressed expression of the genes is known to be involved in flowering regulation, especially in the Ehd1/Hd3a pathway and a series of RFT homologs. RID1 seems to be independent of the circadian clock. A model was proposed to place RID1 in the molecular pathways of flowering regulation in rice, for which there are two indispensable elements. In the first, RID1 is controlling the phase transition and initiation of floral induction. In the other, the Hd3a/RFL1/FTL complex acts as the immediate inducer of flowering. Loss of function in either element would cause never-flowering. Once the phase transition is induced with the activation of RID1, flowering signal is transduced and regulated through the various pathways and eventually integrated with FT-like proteins to induce flowering.
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s00438-009-0483-1URLPMID:19798512 [本文引用: 2]
Ethylene and jasmonic acid (JA) have been proposed as key compounds for wound signaling in plants. In Arabidopsis , ETHYLENE INSENSITIVE3 (EIN3), which is an essential transcription factor for ethylene signaling, is regulated at the post-transcriptional level, while transcriptional regulation of EIN3 or EIN3-LIKE ( EIL ) genes has not been well documented. The expression of 6 rice EIL genes ( OsEIL1-6 ) was analyzed and only OsEIL1 and 2 were found to be wound-inducible EIL . OsEIL2 was also induced by JA. Electrophoretic mobility shift assays showed that recombinant OsEIL1 and 2 proteins bound to specific DNA sequences that are recognized by a wound-inducible tobacco EIL. Accumulation of OsEIL1 and 2 transcripts reached a maximum at 1 and 0.5 h after wounding, respectively, and the corresponding DNA-binding activity in nuclear extracts of rice leaves was increased at 1 h after wounding. Candidates for OsEIL-target genes were selected by microarray analysis of wounded rice and by promoter sequence analyses of wound-inducible genes identified by microarray analysis. In OsEIL1 - and/or 2 -suppressed rice plants, the expression of at least four of 18 candidate genes analyzed was down-regulated. These results indicate the importance of inducible OsEILs in wound signaling in rice.
[本文引用: 2]
DOI:10.1016/S0378-1119(96)00521-5URLPMID:8996094 [本文引用: 2]
Abstract A novel myb-like gene (AtmybL2) was isolated from an Arabidopsis thaliana cDNA library. The single copy gene was localised on chromosome I. A gene specific transcript is preferentially found in leaves. The predicted gene product consists of a conservative N-terminal myb-domain known to be involved in DNA-binding and a unique proline-rich C-terminal part. Remarkably, the myb-domain includes only one of the typical two or three tryptophan repeats found in other myb-like proteins.
DOI:10.1007/s00425-010-1263-1URLPMID:20830595 [本文引用: 1]
A dwarf mutant, dwarf 62 (d62), was isolated from rice cultivar 93-11 by mutagenesis with -rays. Under normal growth conditions, the mutant had multiple abnormal phenotypes, such as dwarfism, wide and dark-green leaf blades, reduced tiller numbers, late and asynchronous heading, short roots, partial male sterility, etc. Genetic analysis indicated that the abnormal phenotypes were controlled by the recessive mutation of a single nuclear gene. Using molecular markers, the D62 gene was fine mapped in 131-kb region at the short arm of chromosome 6. Positional cloning of D62 gene revealed that it was the same locus as DLT/OsGRAS-32, which encodes a member of the GRAS family. In previous studies, the DLT/OsGRAS-32 is confirmed to play positive roles in brassinosteroid (BR) signaling. Sequence analysis showed that the d62 carried a 2-bp deletion in ORF region of D62 gene which led to a loss-of-function mutation. The function of D62 gene was confirmed by complementation experiment. RT-PCR analysis and promoter activity analysis showed that the D62 gene expressed in all tested tissues including roots, stems, leaves and panicles of rice plant. The d62 mutant exhibited decreased activity of -amylase in endosperm and reduced content of endogenous GA1. The expression levels of gibberellin (GA) biosynthetic genes including OsCPS1, OsKS1, OsKO1, OsKAO, OsGA20ox2/SD1 and OsGA2ox3 were significantly increased in d62 mutant. Briefly, these results demonstrated that the D62 (DLT/OsGRAS-32) not only participated in the regulation of BR signaling, but also influenced GA metabolism in rice.
DOI:10.1038/nature07272URLPMID:18690207 [本文引用: 1]
Abstract Shoot branching is a major determinant of plant architecture and is highly regulated by endogenous and environmental cues. Two classes of hormones, auxin and cytokinin, have long been known to have an important involvement in controlling shoot branching. Previous studies using a series of mutants with enhanced shoot branching suggested the existence of a third class of hormone(s) that is derived from carotenoids, but its chemical identity has been unknown. Here we show that levels of strigolactones, a group of terpenoid lactones, are significantly reduced in some of the branching mutants. Furthermore, application of strigolactones inhibits shoot branching in these mutants. Strigolactones were previously found in root exudates acting as communication chemicals with parasitic weeds and symbiotic arbuscular mycorrhizal fungi. Thus, we propose that strigolactones act as a new hormone class-or their biosynthetic precursors-in regulating above-ground plant architecture, and also have a function in underground communication with other neighbouring organisms.
DOI:10.1104/pp.17.00301URLPMID:28404726 [本文引用: 1]
Strigolactones (SLs) are a class of plant hormones that regulate diverse physiological processes, including shoot branching and root development. They also act as rhizosphere signaling molecules to stimulate the germination of root parasitic weeds and the branching of arbuscular mycorrhizal fungi. Although various types of cross talk between SLs and other hormones have been reported in physiological analyses, the cross talk between gibberellin (GA) and SLs is poorly understood. We screened for chemicals that regulate the level of SLs in rice (Oryza sativa) and identified GA as, to our knowledge, a novel SL-regulating molecule. The regulation of SL biosynthesis by GA is dependent on the GA receptor GID1 and F-box protein GID2. GA treatment also reduced the infection of rice plants by the parasitic plant witchers weed (Striga hermonthica). These data not only demonstrate, to our knowledge, the novel plant hormone cross talk between SL and GA, but also suggest that GA can be used to control parasitic weed infections.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}