 ,1,2,**, 刘钊2,**, 肖爽2, 李芳军2, 吴霞3, 王保民2, 田晓莉,2,*
,1,2,**, 刘钊2,**, 肖爽2, 李芳军2, 吴霞3, 王保民2, 田晓莉,2,*Effects of PSAG12-IPT gene expression on leaf senescence, yield, and fiber quality in cotton
WANG Ye,1,2,**, LIU Zhao2,**, XIAO Shuang2, LI Fang-Jun2, WU Xia3, WANG Bao-Min2, TIAN Xiao-Li,2,*通讯作者: * 田晓莉, E-mail:tianxl@cau.edu.cn
第一联系人:
收稿日期:2020-11-21接受日期:2021-04-26网络出版日期:2021-06-09
| 基金资助: |
Corresponding authors: * E-mail:tianxl@cau.edu.cn
First author contact:
Received:2020-11-21Accepted:2021-04-26Published online:2021-06-09
| Fund supported: |
作者简介 About authors
王晔,E-mail:wangyebua@126.com

摘要
探讨PSAG12-IPT基因导入对棉花叶片衰老性状、产量和纤维品质的影响。本研究以9个转化事件的纯合阳性株系为试验材料, 于2011—2012年在中国农业大学上庄实验站开展研究。结果表明, 转基因株系叶片衰老阶段IPT基因表达量大幅上调, Z+ZR类细胞分裂素含量显著高于野生型冀合321。根据Z+ZR类细胞分裂素含量将9个转PSAG12-IPT基因株系聚类分为抗衰老能力强、弱和中间型3大类, 不同类型株系在叶片衰老阶段的叶绿素和可溶性蛋白含量较野生型增加幅度不同。转基因株系单株成铃数(直径≥2 cm)和铃重与野生型相当或高于野生型, 但各株系第一次收获时的吐絮率显著低于野生型, 且降低幅度与抗衰老能力成正比。这导致抗衰老能力较强和中等的PSAG12-IPT过表达株系(OE-37和OE-38)因有效吐絮铃少而减产, 而抗衰老能力弱的株系(OE-30)籽棉产量与野生型相当。PSAG12-IPT基因过表达株系的纤维长度整齐度指数显著高于野生型, 马克隆值有增加趋势, 其他品质指标与野生型差别不大。
关键词:
Abstract
A chimeric gene of bacterial IPT and the senescence-specific SAG12 promoter from Arabidopsis (PSAG12-IPT) was overexpressed in cotton, and nine independent transgenic lines were obtained. To identify the leaf senescence characteristics, yield, and fiber quality of PSAG12-IPT cotton plants, we performed the experiments at the Shangzhuang Experimental Station of China Agricultural University in 2011 and 2012. The results showed that the expression of IPT genes in transgenic lines was dramatically up-regulated at leaf aging stage, and the content of Z+ZR-type cytokinin was significantly higher than that of wild type Jihe 321. According to the content of Z+ZR-type cytokinin, nine transgenic lines were clustered into three categories with strong, weak, and moderate anti-aging ability. During leaf senescence period, the contents of chlorophyll and soluble proteins in transgenic lines were positively related to the content of Z+ZR-type cytokinin. Compared to wild type, transgenic lines had a higher or equal number of bolls (diameter ≥ 2 cm) per plant and boll weigh, but their boll-opening rate at the first harvest was significantly decreased, and the extent of reduction was positively related to the anti-aging ability. Consequently, the seed cotton yield of PSAG12-IPT lines (OE-37 and OE-38) with strong or moderate anti-aging ability decreased due to less opened bolls, while the line (OE-30) with weak anti-aging ability had a comparable yield to that of wild type. The fiber length uniformity of PSAG12-IPT cotton lines was significantly higher than that of wild type. Compared with wild type, the micronaire results of transgenic lines revealed an increasing trend, and fiber length, breaking strength, and breaking elongation was not significantly different from that of wild type.
Keywords:
PDF (737KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王晔, 刘钊, 肖爽, 李芳军, 吴霞, 王保民, 田晓莉. 转PSAG12-IPT基因对棉花叶片衰老及产量和纤维品质的影响. 作物学报, 2021, 47(11): 2111-2120 DOI:10.3724/SP.J.1006.2021.04251
WANG Ye, LIU Zhao, XIAO Shuang, LI Fang-Jun, WU Xia, WANG Bao-Min, TIAN Xiao-Li.
棉花早衰是指叶片在花铃盛期过早黄化、干枯和脱落[1,2], 早衰发生严重的年份减产幅度可达20%~50% [3], 并导致棉花纤维强度下降和纤维长度变短[4]。
植物生理学的传统观点认为, 根系是细胞分裂素(cytokinin, CTKs)的主要合成部位, 可以通过木质部向地上部输送CTKs调节叶片衰老[5]。有研究表明, 根系供应的CTKs减少可能是大豆[6]和向日葵[7]叶片衰老的主要原因。Barry等[8]首次从根癌农杆菌中分离出异戊烯基转移酶基因IPT (Isopentenyl-trans ferase gene), 其产物异戊烯基转移酶是细胞分裂素生物合成的关键酶, 催化细胞分裂素从头合成的第一步反应, 促使5°-AMP和异戊烯基焦磷酸合成异戊烯基腺核苷酸, 之后转化为各种细胞分裂素。Lohman等[9]从拟南芥中克隆出一组衰老相关基因SAG (senescence-associated gene), 并证明其中的SAG12受衰老高度特异正调控, 仅在自然衰老状态下的组织中表达。Gan和Amasino[10]进一步将SAG12启动子与来自根癌农杆菌的编码IPT基因构建成嵌合基因(PSAG12-IPT)转入烟草。该嵌合基因具有自动调控特性, 即当衰老发生时, PSAG12驱动IPT基因表达, 植株体内CTKs含量增加, 衰老受到抑制; 当CTKs的含量达到一定水平时, PSAG12的活性受到抑制, IPT基因关闭; 当衰老再次开始时, PSAG12-IPT再度表达。因此, 转基因植株体内的CTKs始终维持在一定的水平, 叶片则表现出较高的光合作用和较长的寿命, 衰老得到延缓[10]。此后, 嵌合基因PSAG12-IPT被陆续转入到生菜[11]、水稻[12]、小麦[13]和苎麻[14]等多种作物中, 不仅延缓了叶片的衰老, 而且在一定程度上改善了作物的农艺性状和抗逆能力。
Yu等[15]利用种子特异表达的菜豆蛋白启动子(Ph/P)与IPT基因构建载体转化棉花, 改变了棉花纤维的长度, 且转IPT基因棉花种子中的CTKs含量明显提高, 根系发达, 次生根数目增多。李静等[16]利用花粉管通道技术将含有嵌合基因PSAG12-IPT的pBG121质粒转入早衰型陆地棉品种中棉所10号中, 测定T1代植株的叶绿素和CTKs含量, 发现转基因植株的衰老得到延缓。Liu等[17]通过农杆菌介导法获得转IPT基因棉株, 其叶绿素和CTKs含量及清除活性氧的酶活性显著增加。目前, 对转IPT基因棉花的抗早衰功能研究相对较多, 对产量和品质等经济性状及抗早衰能力与经济性状间的关系研究较少。本研究以陆地棉品种冀合321为受体, 获得了9个转PSAG12-IPT基因纯合株系, 在田间条件下研究了转基因株系的抗衰老能力及产量和纤维品质性状, 分析了抗衰老能力与产量性状的联系, 旨在为培育高产优质抗早衰棉花品种提供基础材料。
1 材料与方法
1.1 试验材料
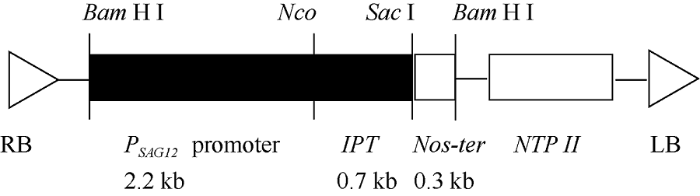
所用质粒PSG529来自于Wisconsin大学Richard M Amasino教授实验室, 经该实验室许可后由中国农业大学付永彩老师转赠。该质粒含有拟南芥衰老特异性启动子PSAG12和根癌农杆菌的异戊烯基转移酶基因IPT, 筛选基因为NTP II (图1)。山西省农业科学院棉花研究所采用农杆菌侵染法, 向野生型陆地棉(Gossypium hirsutum L.)冀合321中导入该质粒, 获得转基因当代株系, 经逐年筛选、鉴定和繁育, 共获得9个转化事件的纯合阳性T3代株系用于本研究, 株系编号分别为OE-9、OE-17、OE-19B、OE- 19L、OE-22、OE-27、OE-30、OE-37、OE-38。图2为各株系目的基因IPT和标记基因NPT II的PCR鉴定结果, 其中IPT基因的上、下游引物分别为5'-GCGTCTAATTTTCGGTCCAA-3'和5'-AA TGGATGACCTTCGAATCC-3', NPT II基因的上、下游引物分别为5'-TCCGGCCGCTTGGGTGGAGA G-3'和5'-CTGGCGCGAGCCCCTGATGCT-3', 结果表明目的基因和标记基因已整合到棉花基因组中。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1质粒PSG529结构图
Fig. 1Construction of plasmid PSG529
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2转PSAG12-IPT基因棉花植株的PCR检测
A: 叶片中IPT基因的PCR结果; B: 叶片中NPT II基因的PCR结果; C: 根系中NPT II基因的PCR结果。
Fig. 2PCR detection of transgenic cotton plants with PSAG12-IPT gene
A: PCR detection of IPT genes in the leaves of transgenic plants; B: PCR detection of NPT II gene in the leaves of transgenic plants; C: PCR detection of NPT II gene in the roots of transgenic plants.
1.2 试验设计
田间试验于2011年和2012年在中国农业大学上庄试验站(40°08′N, 116°10′E)进行。试验地土壤为壤土。试验采用完全随机区组设计, 重复3次。小区行长5.0 m, 行距0.9 m, 2011年为2行区, 小区面积9.0 m2; 2012年为4行区, 小区面积18.0 m2。2011年和2012年分别于5月10日和5月8日播种, 种植密度为37,500株 hm-2。试验地四周设4行保护行, 栽培管理措施同一般生产田。1.3 测定项目与方法
2011年和2012年分别于7月23日和8月4日打顶, 打顶当天标记刚平展的主茎叶片, 叶龄记为 0 d。2011年取叶龄0 d (7月23日)、20 d (8月12日)、36 d (8月28日)、47 d (9月8日)和62 d (9月23日)的叶片, 2012年取叶龄0 d (8月4日)、14 d (8月18日)、28 d (9月1日)、42 d (9月15日)和56 d (9月29日)的叶片, 用于测定相关指标。1.3.1 IPT基因相对表达量检测 用TRIzol试剂盒(Invitrogen, Carlsbad, CA, USA)提取叶片RNA, 用Primer Primier 5.0软件(Primier Biosoft International, Palo Alto, CA, US)设计引物。以UBQ7为内参基因, 引物序列为Sense: 5°-atccgtcctgcatcaagaga-3°, AntiSense: 5°-tcccggctatgtcatctttc-3°。实时荧光定量PCR (ABI 7500 Real-Time PCR System): 用TaKaRa公司SYBR Green I (Molecular probes)做为荧光指示探针。RT-PCR反应体系为15 μL, 在荧光定量PCR仪上按照两步法扩增程序, 即95℃预变性30 s; 95℃变性5 s, 60℃退火30 s, 40个循环; 最后进行溶解曲线的检测收集。根据2-ΔΔCt方法[18]计算基因相对表达量。
1.3.2 生理指标测定 采用间接酶联免疫吸附分析法[19]测定玉米素和玉米素核苷(Zeatin+Zeatin riboside, Z+ZR)类的细胞分裂素含量, 抗体由中国农业大学作物化控研究中心提供。参照Arnon[20]的方法测定叶绿素含量, 参考Read等[21]的方法测定可溶性蛋白含量。
1.3.3 产量及产量构成因素 小区实收测产, 收获2次。2011年的收获时间为10月8日和10月25日, 2012年为10月11日和10月28日。第1次收获时在每小区选取生长一致的棉株8株测定株高、果枝始节、果枝数、铃数、吐絮率等, 称量标记植株的吐絮铃重量, 计算平均铃重, 之后轧花测定衣分。
1.3.4 纤维品质 第1次收获时, 在每小区随机选取棉株中部果枝内围铃(第1~2果节) 30个左右, 轧花后由农业部棉花品质监督检验测试中心用HVI900系统进行测定。
1.4 数据统计
采用SPSS18.0 (SPSS Inc., Chicago, IL, USA)进行聚类分析和方差分析(ANOVA), 用Duncan’s法进行多重比较(P≤0.05), 用SigmaPlot 11.0 (Systat Software Inc, Washington St, USA)绘图。2 结果与分析
2.1 转PSAG12-IPT基因对棉花叶片衰老阶段IPT基因表达量和Z+ZR含量的影响
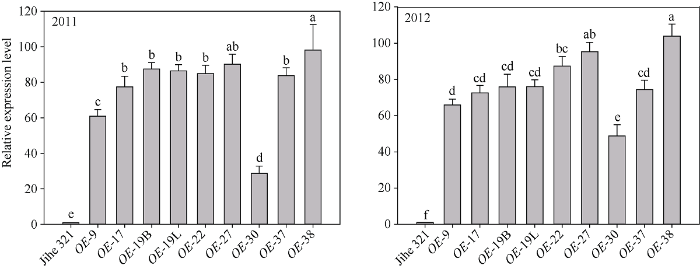
与野生型相比, 转基因株系叶片在开始衰老阶段(叶龄62 d, 2011年; 叶龄56 d, 2012年)异源IPT基因的相对表达量均有提高, 差异达显著水平, 且不同株系提高幅度存在差异, 其中OE-38相对表达量最高、OE-30相对表达量最少, 2年结果趋势一致(图3)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3棉花叶片衰老阶段PSAG12-IPT基因在转基因株系中的表达分析
柱上不同小写字母表示在0.05水平差异显著。
Fig. 3Relative expression of PSAG12-IPT gene in different transgenic lines of cotton at leaf senescence stage
Different lowercase letters above the bars indicate significant difference at the 0.05 probability level.
从图4可以看出, 在叶片开始衰老阶段(叶龄62 d, 2011年; 叶龄56 d, 2012年), PSAG12-IPT基因导入使各转基因株系叶片中Z+ZR含量显著升高(2011年OE-9株系除外), 其中OE-38的增幅最高, 达到71.5%~79.4%。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4转PSAG12-IPT基因对棉花叶片衰老阶段细胞分裂素含量的影响
柱上不同小写字母表示在0.05水平差异显著。
Fig. 4Effects of PSAG12-IPT gene overexpression on cytokinin concentration of cotton leaves at leaf senescence stage
Different lowercase letters above the bars indicate significant difference at the 0.05 probability level.
2.2 转PSAG12-IPT基因棉花株系的聚类分析
根据叶片Z+ZR含量进行聚类分析, 当聚类数为3时, 各类群之间方差达到显著水平。I类为低Z+ZR型, 包括OE-9和OE-30 2个株系, 抗衰老能力较弱; II类为中等Z+ZR型, 包括6个株系; III类为高Z+ZR型, 只有OE-38 1个株系, 抗衰老能力最强。对9个转基因株系叶片衰老阶段IPT表达量进行聚类分析得到相同结果(数据未列出)。从3个类型中各选1个代表性株系, 即抗衰老能力较弱的OE-30、抗衰老能力最强的OE-38和抗衰老能力介于二者之间的OE-37, 用于叶片功能和产量及纤维品质研究。Table 1
表1
表1不同转基因棉花株系叶片细胞分裂素的聚类分析
Table 1
| 类群 Cluster | 株系代号 Code of line | 株系个数 Number of lines | 频率 Frequency (%) | 类平均标准差 Mean of cluster (nmol g-1) | 变幅 Range (nmol g-1) |
|---|---|---|---|---|---|
| I | OE-9, OE-30 | 2 | 22.22 | 75.66 c | 75.45-75.88 |
| II | OE-17, OE-19B, OE-19L, OE-22, OE-27, OE-37 | 6 | 66.67 | 87.43 b | 82.23-93.19 |
| III | OE-38 | 1 | 11.11 | 113.62 a | 113.62 |
新窗口打开|下载CSV
2.3 转PSAG12-IPT基因株系叶片IPT基因表达量和Z+ZR含量变化动态
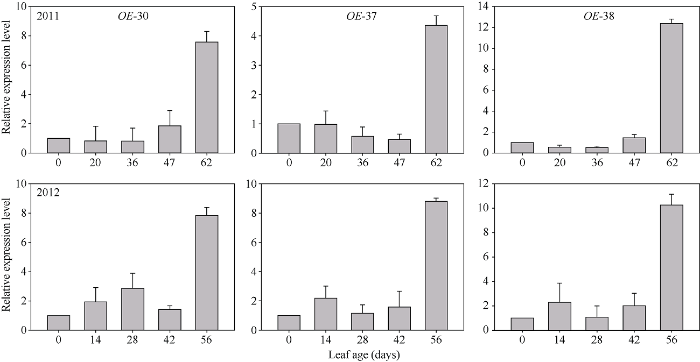
由图5可知, PSAG12-IPT基因在OE-30、OE-37和OE-38号转基因株系标记叶片的不同生长时期均有表达, 随着叶片的衰老其表达量增加。与叶片展开当天(叶龄0 d)相比, OE-30、OE-37和OE-38株系进入衰老阶段的叶片(叶龄56~62 d)其PSAG12-IPT基因表达量分别增加了6.6~6.8倍、3.4~7.8倍、9.3~11.4倍。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5PSAG12-IPT基因在转基因棉株叶片中相对表达量变化动态
Fig. 5Relative expression profile of PSAG12-IPT gene in leaves of transgenic cotton lines
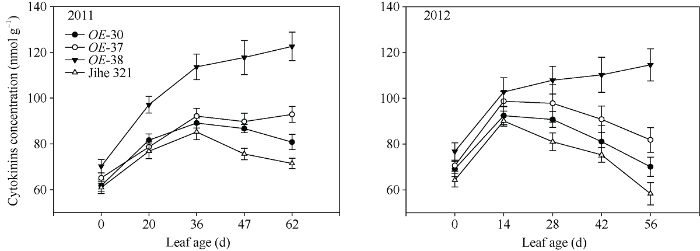
由图6可知, 野生型叶片Z+ZR含量随叶龄增长呈先增加再下降的趋势, IPT过表达株系OE-30 (抗早衰能力弱)和OE-37 (抗早衰能力中等)的变化趋势与野生型相同, 但Z+ZR含量高于野生型, 且提高幅度随叶龄增长而增大。抗早衰能力最强的OE-38, 其叶片Z+ZR含量随叶龄增长先快速增加后缓慢增加, 且在叶龄20 d (2011年)或28 d (2012年)及以后显著高于野生型和其他2个过表达株系, 其中2011年在叶龄62 d、2012年在叶龄56 d时较野生型增加71.5%和96.5%。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6转基因株系和野生型棉株叶片细胞分裂素含量变化动态
Fig. 6Dynamic variations of cytokinin concentrations in leaves of wild type and transgenic cotton lines
2.4 转PSAG12-IPT基因株系叶片叶绿素和可溶性蛋白含量变化动态
由图7可知, 棉花叶片叶绿素含量自叶片展开后即逐渐下降。IPT过表达株系的叶片持绿性较好, 其叶绿素含量在叶片生长的中后期不同程度高于野生型。图7
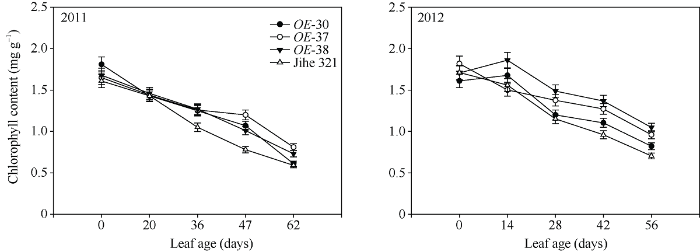 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7转基因株系和野生型棉株叶片叶绿素含量变化动态
Fig. 7Dynamic variations of chlorophyll contents in leaves of wild type and transgenic cotton lines
蛋白质的降解是叶片衰老的基本特征之一。由图8可知, 叶片展开后可溶性蛋白含量快速上升, 但2~3周后即开始下降。与野生型相比, OE-38和OE-37株系的叶片可溶性蛋白含量在叶龄56~62 d之间分别增加了30.3%~43.5%和22.1%~37.6%, OE- 30与野生型的差异较小。
图8
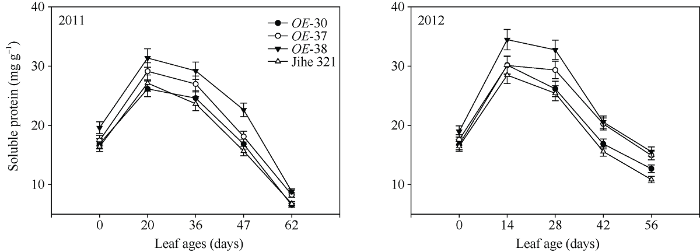 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8转基因株系和野生型棉株叶片可溶性蛋白含量变化动态
Fig. 8Dynamic variations of soluble protein concentrations in leaves of wild type and transgenic cotton lines
2.5 转PSAG12-IPT基因对棉花农艺性状的影响
与野生型相比, IPT过表达株系株高呈降低趋势, 其中2011年达显著水平(表2)。OE-30、OE-37和OE-38的果枝始节均较野生型显著提高, 提高范围为1~2个节位, 说明IPT过表达株系的花芽分化较晚, 有可能导致晚熟。几个转基因株系的果枝数与野生型相比有减少趋势, 但差异不显著。Table 2
表2
表2PSAG12-IPT基因导入对棉花农艺性状的影响
Table 2
| 株系 Line | 2011 | 2012 | ||||
|---|---|---|---|---|---|---|
| 株高 Plant height (cm) | 果枝始节 First node producing fruiting branch | 果枝数 Number of fruiting branch | 株高 Plant height (cm) | 果枝始节 First node producing fruiting branch | 果枝数 Number of fruiting branch | |
| 冀合321 Jihe 321 | 103.3 a | 6.3 b | 14.0 a | 134.5 a | 7.6 b | 16.9 a |
| OE-30 | 91.7 bc | 8.0 a | 12.3 a | 127.1 a | 9.5 a | 16.5 a |
| OE-37 | 88.0 c | 8.0 a | 12.0 a | 129.8 a | 8.3 a | 16.2 a |
| OE-38 | 100.3 b | 8.3 a | 13.0 a | 123.4 a | 9.6 a | 15.0 a |
新窗口打开|下载CSV
2.6 转PSAG12-IPT基因对棉花产量和纤维品质的影响
由表3可知, 抗早衰能力较弱的OE-30的籽棉产量与野生型无显著差异, 但抗早衰能力中等和较强的OE-37和OE-38的籽棉产量显著低于野生型, 其中2011年分别降低35.2%和68.1%、2012年降低9.6%和14.1%。OE-37和OE-38的单株成铃数(直径≥2 cm)未减少, OE-38在2011年的单株成铃数显著多于野生型, 其减产的主要原因在于晚熟, 2011年10月8日第一次收获时OE-37和OE-38的吐絮率较野生型分别降低了43.6%和59.9%, 2012年10月11日第一次收获时二者的吐絮率较野生型降低了54.5%和82.8%, 这导致OE-37和OE-38的有效收获铃数大幅减少。此外, OE-37和OE-38的铃重也有低于野生型的趋势, 这可能也是产量降低的原因之一。IPT过表达株系的衣分与野生型无显著差异。Table 3
表3
表3PSAG12-IPT基因导入对棉花产量和产量构成因素的影响
Table 3
| 年份 Year | 株系 Line | 单株成铃数 Boll number per plant | 铃重 Boll weight (g) | 衣分 Lint percentage (%) | 吐絮率 Boll opening percentage (%) | 籽棉产量 Seed cotton yield (kg hm-2) |
|---|---|---|---|---|---|---|
| 2011 | 冀合321 Jihe 321 | 12.4 b | 4.2 ab | 37 a | 94.4 a | 1257.0 a |
| OE-30 | 18.6 a | 4.6 a | 36 a | 71.5 b | 1364.8 a | |
| OE-37 | 12.3 b | 4.0 b | 40 a | 53.2 bc | 814.9 b | |
| OE-38 | 17.7 a | 3.8 b | 34 a | 37.9 c | 401.3 c | |
| 2012 | 冀合321 Jihe 321 | 24.2 a | 4.3 a | 35 a | 49.9 a | 1554.9 a |
| OE-30 | 24.0 a | 4.2 a | 33 a | 30.0 b | 1605.9 a | |
| OE-37 | 21.0 a | 4.0 a | 33 a | 22.7 c | 1405.1 b | |
| OE-38 | 23.6 a | 3.7 a | 32 a | 8.6 d | 1335.5 b |
新窗口打开|下载CSV
由纤维品质2年均值(表4)可以看出, 与野生型相比, IPT过表达株系的纤维长度和断裂比强度呈增加趋势但差异不显著, 整齐度指数则显著提高; 马克隆值与野生型持平或显著增加(OE-38), 断裂伸长率变化不大。
Table 4
表4
表4PSAG12-IPT基因导入对棉花品质的影响
Table 4
| 株系 Line | 纤维长度 Fiber length (mm) | 整齐度指数 Uniformity index (%) | 马克隆值 Micronaire value | 断裂伸长率 Breaking elongation (%) | 断裂比强度 Breaking strength (cN tex-1) |
|---|---|---|---|---|---|
| 冀合321 Jihe 321 | 29.2 a | 82.9 c | 4.1 b | 6.5 a | 29.5 a |
| OE-30 | 31.6 a | 84.7 b | 4.4 ab | 6.2 a | 30.6 a |
| OE-37 | 30.5 a | 84.0 b | 4.3 b | 6.4 a | 30.5 a |
| OE-38 | 29.5 a | 85.3 a | 4.7 a | 6.3 a | 29.8 a |
新窗口打开|下载CSV
3 讨论
3.1 转PSAG12-IPT基因延缓了棉花叶片衰老
叶片衰老伴随着叶绿素、蛋白质和核酸等的降解及光合速率下降。细胞分裂素(CTKs)对叶片衰老有延缓作用[22], PSAG12-IPT嵌合基因只在叶片开始衰老时才被诱导表达并具有自动调控功能[10], 可以避免因组成型表达在全株范围内持续形成高浓度CTKs而影响植物生长发育和产量形成[23,24]。在本研究中, 与野生型冀合321相比, 转PSAG12-IPT基因棉花株系在叶片衰老阶段IPT基因表达量大幅上调, Z+ZR类CTKs含量显著升高, CTKs调节编码感光叶绿素结合蛋白的基因的活性, 并通过刺激衰老质体向叶绿体再分化的基因的表达, 延缓叶绿素降解[25,26], 衰老叶片的蛋白质被有效地转移至幼嫩组织和生殖器官[11], 从而延迟叶片衰老。这与前人在棉花[17]、小麦[13]和玉米[27]中的报道一致。此外, 本研究获得的转PSAG12-IPT基因棉花株系抗衰老能力不同, 这可能与外源IPT基因的插入位点[28,29]、外源基因与受体基因的互作有关[30], 也不排除转基因过程自身所导致的差异。3.2 转PSAG12-IPT基因延迟了棉花的成熟
花芽分化时间是影响棉花熟性的重要因素之一[31,32]。本研究中3个转PSAG12-IPT基因株系(OE-30、OE-37和OE-38)的果枝始节均显著高于野生型, 2年平均较野生型提高1~2个节位, 说明转基因株系的花芽分化开始较晚。研究表明, 在转IPT基因玉米中, 衰老激活IPT基因表达, CTKs的产生导致开花明显延迟[27]。当植株内CTKs、ABA和GAs含量较高或IAA含量较低时都可以促进棉花的花芽分化, 反之抑制花芽分化, 植物激素的含量及其动态平衡对花芽分化起着重要的调控作用[33,34]。本研究的IPT过表达株系在幼苗阶段可能也会影响生长点的CTKs含量及与其他植物激素的互作, 从而打破了棉花花芽分化阶段植物激素的固有平衡, 延迟了花芽分化。此外, 虽然3个株系(OE-30、OE-37和OE-38)的果枝始节无显著差异(表2), 但OE-37和OE-38在10月10日左右的吐絮率低于OE-30, 晚熟现象更为严重(表3)。株系间成熟阶段熟性的差异可能与棉铃的时空分布有关, OE-37和OE-38株系的叶绿素、蛋白质和CTKs含量高于OE-30, 表明其营养生长势较强, 这可能导致同化物向库器官的转移减少[13], 从而加重中下部和内围果节产量器官的脱落, 使上部果枝和外围果节棉铃占总铃数的比例提高引起晚熟。
3.3 转PSAG12-IPT基因对棉花产量和纤维品质的影响
通过过表达IPT基因[35,36]、抑制CTKs降解酶(CKX)[37]基因, 或外源施加CTKs类调节剂激动素(KT, 6-furfurylaminopurine)[38]提高植株CTKs含量, 可以增加水稻和棉花的产量。本研究结果与此不同, OE-30的籽棉产量与野生型持平, 抗衰老能力强的OE-37和OE-38株系的籽棉产量反而显著低于野生型。从产量构成因素可知, OE-37和OE-38的铃数和铃重与野生型相比并不低(表3), 其减产的原因主要由于晚熟导致部分棉铃在本试验条件下不能正常成熟形成有效产量。早期报道中高浓度的CTKs抑制纤维的伸长[15]; Zhao等[37]研究表明, 适度提高CTKs水平, 棉花纤维长度与野生型对照相比差异并不显著。本试验中导入PSAG12-IPT基因后, 纤维长度整齐度指数显著提高, 马克隆值和断裂比强度也表现出高于野生型的趋势, 这可能与转基因株系在生育后期叶片功能较强、合成的同化物较多有关, 也不排除纤维中的CTKs也得到提高后直接调控纤维发育过程。
综上, PSAG12-IPT嵌合基因的导入提高了棉花的抗衰老能力, 但同时延缓了棉花的成熟。应用该基因对生产上有早衰缺陷的品种进行遗传改良时, 需注意协调抗衰与早熟的关系, 以同时达到早熟高产优质的生产目的。
4 结论
PSAG12-IPT基因的导入提高了棉花叶片衰老阶段IPT基因的表达量及叶片叶绿素、蛋白质和细胞分裂素(CTKs)含量、延迟了叶片的衰老。转PSAG12- IPT基因延迟了棉花的花芽分化, 且成熟阶段转基因株系的抗衰老能力越强、早熟性越差, 抗衰老能力中等和较强的转基因株系因晚熟而降低产量。PSAG12-IPT基因导入在一定程度上改善了棉花纤维品质。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
Berlin: Springer Verlag Press,
[本文引用: 1]
PMID [本文引用: 1]

Cytokinins (CKs) coming from the roots via the xylem are known to delay leaf senescence, and their decline may be important in the senescence of soybean (Glycine max) plants during pod development (monocarpic senescence). Therefore, using radioimmunoassay of highly purified CKs, we quantified the zeatin (Z), zeatin riboside (ZR), the dihydro derivatives (DZ, DZR), the O-glucosides, and DZ nucleotide in xylem sap collected from root stocks under pressure at various stages of pod development. Z, ZR, DZ, and DZR dropped sharply during early pod development to levels below those expected to retard senescence. Pod removal at full extension, which delayed leaf senescence, caused an increase in xylem sap CKs (particularly ZR and DZR), while depodding at late podfill, which did not delay senescence, likewise did not increase the CK levels greatly. The levels of the O-glucosides and the DZ nucleotide were relatively low, and they showed less change with senescence or depodding. The differences in the responses of individual CKs to senescence and depodding suggest differences in their metabolism. Judging from their activity, concentrations and response to depodding, DZR and ZR may be the most important senescence retardants in soybean xylem sap. These data also suggest that the pods can depress CK production by the roots at an early stage and this decrease in CK production is required for monocarpic senescence in soybean.
DOIPMID [本文引用: 1]
The cytokinin content of root exudate of sunflowers increases during the exponential growth phase of the plants. The concentration of cytokinins drops, however, by a factor of ten when the plants have reached their final size. The reduced supply of cytokinins from the root to the shoot is regarded as one of the factors bringing about shoot senescence.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 3]
Controlling expression of IPT, a gene encoding isopentenyltransferase (the enzyme that catalyzes the rate-limiting step in cytokinin biosynthesis), with a senescence-specific promoter results in the suppression of leaf senescence. Transgenic tobacco plants expressing this chimeric gene do not exhibit the developmental abnormalities usually associated with IPT expression because the system is autoregulatory. Because sufficient cytokinin is produced to retard senescence, the activity of the senescence-specific promoter is attenulated. Senescence-retarded leaves exhibit a prolonged, photosythetically active life-span. This result demonstrates that endogenously produced cytokinin can regulate senescence and provides a system to specifically manipulate the senescence program.
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
PMID [本文引用: 1]
The two most commonly used methods to analyze data from real-time, quantitative PCR experiments are absolute quantification and relative quantification. Absolute quantification determines the input copy number, usually by relating the PCR signal to a standard curve. Relative quantification relates the PCR signal of the target transcript in a treatment group to that of another sample such as an untreated control. The 2(-Delta Delta C(T)) method is a convenient way to analyze the relative changes in gene expression from real-time quantitative PCR experiments. The purpose of this report is to present the derivation, assumptions, and applications of the 2(-Delta Delta C(T)) method. In addition, we present the derivation and applications of two variations of the 2(-Delta Delta C(T)) method that may be useful in the analysis of real-time, quantitative PCR data.Copyright 2001 Elsevier Science (USA).
[本文引用: 1]
[本文引用: 1]
PMID [本文引用: 1]
PMID [本文引用: 1]
PMID [本文引用: 1]
Answers to long-standing questions concerning the molecular mechanism of auxin action and auxin's exact functions in plant growth and development are beginning to be uncovered through studies using mutant and transgenic plants. We review recent work in this area in vascular plants. A number of conclusions can be drawn from these studies. First, auxin appears essential for cell division and viability, as auxin auxotrophs isolated in tissue culture are dependent on auxin for growth and cannot be regenerated into plants even when auxin is supplied exogenously. Secondly, plants with transgenes that alter auxin levels are able to regulate cellular auxin concentrations by synthesis and conjugation; wild-type plants are probably also capable of such regulation. Thirdly, the phenotypes of transgenic plants with altered auxin levels and of mutant plants with altered sensitivity to auxin confirm earlier physiological studies which indicated a role for auxin in regulation of apical dominance, in development of roots and vascular tissue, and in the gravitropic response. Finally, the cloning of a mutationally identified gene important for auxin action, along with accumulating biochemical evidence, hints at a major role for protein degradation in the auxin response pathway.
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 2]
We have genetically modified maize plants to delay leaf senescence. A senescence-enhanced promoter from maize (P(SEE1)) was used to drive expression of the Agrobacterium cytokinin biosynthesis gene IPT in senescing leaf tissue. Three maize lines expressing IPT from P(SEE1), Sg1, Sg2 and Sg3, were analysed in detail, representing mild, intermediate and extreme expression, respectively, of the delayed-senescence phenotype. Backcross populations segregating for the presence or absence of the P(SEE1Xba)IPTNOS transgene also simultaneously segregated for the senescence phenotype. At the time of ear leaf emergence, individuals of lines Sg1 and Sg2 segregating for the presence of the transgene carried about three fewer senescing leaves than control (transgene-minus) segregants, and IPT transcript levels were higher in leaves at incipient senescence than in young leaves. Leaves of transgenic Sg3 plants were significantly greener than controls and progressed directly from fully green to bleached and dead without an intervening yellowing phase. IPT transcript abundance in this line was not related to the initiation of senescence. Extended greenness was accompanied by a delay in the loss of photosynthetic capacity with leaf age. The delayed-senescence trait was associated with relatively minor changes in morphology and development. The phenotype was particularly emphasized in plants grown in low soil nitrogen. The reduced ability of the extreme transgenic line Sg3 to recycle internal nitrogen from senescing lower leaves accounted for significant chlorosis in emerging younger leaves when plants were grown in low nutrient conditions. This study demonstrates that the agronomically important delayed-senescence ('stay-green') trait can be engineered into a monocot crop, and is the first example outside Arabidopsis of senescence modification using a homologous senescence-enhanced promoter.
PMID [本文引用: 1]
Seven transgenic lines of wheat have been evaluated under field conditions during 2 agonomic years. Four lines contained the transgenes for beta-glucuronidase ( uidA), herbicide resistance ( bar) and for one high-molecular-weight (HMW) subunit, and three lines contained only one transgene for one HMW glutenin subunit and no marker genes. Agronomic traits and yield components were studied in transgenic lines and compared with the non-transgenic parent and null segregant lines. Although phenotypic differences for many traits have been found, only heading date and the number of spikelets per spike showed clear genotypic differences for both field trials. All transgenic lines had a longer heading date than parent lines whereas the number of spikelets per spike in transgenic lines was around that for L88-31 and higher for L88-6 than the corresponding parent lines. No differences were found between lines constitutively expressing the uidA and bar genes from those which only expressed the HMW genes. We conclude that differences between transgenic lines and their parents are small, and could be eliminated by backcrossing transgenic lines with their parents and selecting for the wanted genotype.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
PMID [本文引用: 1]
Two field experiments in 1993 and 1994 as well as a laboratory germination experiment were conducted on the Egyptian cotton cultivar Giza 75 (Gossypium barbadense) to determine the effect of six concentrations of kinetin (6-furfurylaminopurine) ranging from 0 to 10.0 mg l(-1) and three different methods of application: (A) seeds were soaked for 24 h before germination (laboratory experiment) or sowing (field experiment) in solutions of different kinetin concentration, (B) cotton plants were sprayed twice with different kinetin concentrations at 60 and 75 days after sowing (DAS) during the square initiation and the beginning of bolling stages, at volume solution of 480 l ha(-1). (C) Seeds were soaked in kinetin solutions before sowing as method A. In addition cotton plants were sprayed twice as method B with the same kinetin concentrations. Kinetin application improved seed viability and seedling vigour as shown by lengths of the hypocotyl, radicle and the entire seedling, as well as seedling fresh weight. Moreover, significant increases were recorded in the number of open bolls/plant, boll weight, lint and seed indices, seed cotton yield/plant, and seed cotton and lint yields/plot. The highest means were obtained at 5 mg kinetin l(-1) concentration and under method C of application. Treatments generally, had no significant effects on lint percentage, yield earliness and fiber properties. These results show that, the use of kinetin at 5 mg l(-1) for pre-soaking seeds before planting and spraying cotton plants at 60 and 75 DAS with the same concentration could improve cotton germination, seed cotton and lint yields.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}