 ,1,2, 郑殿峰,1,4,*, 冯乃杰,1,4,*, 梁喜龙2,3, 项洪涛5, 冯胜杰1, 靳丹2, 刘美玲2, 牟保民2
,1,2, 郑殿峰,1,4,*, 冯乃杰,1,4,*, 梁喜龙2,3, 项洪涛5, 冯胜杰1, 靳丹2, 刘美玲2, 牟保民2Effects of uniconazole on physiological characteristics and microstructure under waterlogging stress at seedling stage in soybean
WANG Shi-Ya,1,2, ZHENG Dian-Feng,1,4,*, FENG Nai-Jie,1,4,*, LIANG Xi-Long2,3, XIANG Hong-Tao5, FENG Sheng-Jie1, JIN Dan2, LIU Mei-Ling2, MU Bao-Min2通讯作者: *郑殿峰, E-mail:zdffnj@163.com;冯乃杰, E-mail:byndfnj@126.com
收稿日期:2020-10-28接受日期:2021-01-13网络出版日期:2021-02-20
| 基金资助: |
Corresponding authors: *E-mail:zdffnj@163.com;E-mail:byndfnj@126.com
Received:2020-10-28Accepted:2021-01-13Published online:2021-02-20
| Fund supported: |
作者简介 About authors
E-mail:wsy1106ok@126.com

摘要
为探究苗期(V1期)淹水胁迫对大豆生理特性和显微结构的影响及烯效唑(S3307)的缓解效应, 以‘垦丰14’为材料, 于V1期进行叶面喷施S3307, 并于喷药后5 d进行淹水处理, 对淹水胁迫下大豆叶片和根系生理特性、下胚轴显微结构及S3307的调控效应进行了测定和分析。结果表明, 淹水胁迫会增加大豆下胚轴通气组织数量, 随淹水胁迫时间延长, 通气组织面积逐渐增大; S3307能提高大豆对淹水逆境的适应性, 增加通气组织数量和通气组织的面积, 以应对淹水胁迫对植株造成的缺氧胁迫。与对照(CK)相比, 淹水胁迫会增加叶片和根系中活性氧(ROS)和丙二醛(MDA)含量, 并随胁迫时间延长而逐渐升高。在淹水胁迫前期会诱导酶促抗氧化防御系统活性的增强, 引起渗透调节物质含量的增加, 随胁迫时间延长, 抗氧化酶活性和渗透调节物质含量均呈下降趋势。S3307可促进叶片和根系中抗氧化酶活性的提高, 抑制ROS和MDA含量的过量积累, 并始终维持较高的渗透调节物质含量, 减缓淹水胁迫造成的损伤。
关键词:
Abstract
To explore the effects of waterlogging stress on the physiological characteristics and microstructure of soybean and the mitigation effect of S3307, the physiological characteristics of soybean leaves and roots, the microstructure of hypocotyls and the regulation effect of S3307 were investigated using Kenfeng 14 as the experimental material with S3307 sprayed on the leaves of soybean at V1 stage and five days after waterlogging stress in a pot experiment. The results showed that waterlogging stress increased the number of ventilated tissues in soybean hypocotyl, and the area of ventilated tissues increased gradually with the prolongation of waterlogging stress. S3307 could improve soybean adaptability to waterlogging adversity, increase the number of ventilated tissue and cope with the hypoxia stress by waterlogging stress on plants. Compared with CK, waterlogging stress increased the accumulation of ROS and membrane lipid peroxidation in leaves and roots, and gradually increased with the prolongation of stress time. Meanwhile, the activity of enzymatic antioxidant defense system was enhanced at early stage under waterlogging stress, resulting in an increase in the content of osmotic regulatory substances, which decreased with the prolongation of stress time. S3307 can promote the activity of antioxidant enzyme in leaves and roots, inhibit the excessive accumulation of ROS and MDA content, maintain a high content of osmotic regulation substances, and alleviate the damage caused by waterlogging stress. In summary, there were differences in response to different organs under waterlogging stress. S3307 can alleviate the damage to soybean plants caused by waterlogging stress to a certain extent.
Keywords:
PDF (4970KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王诗雅, 郑殿峰, 冯乃杰, 梁喜龙, 项洪涛, 冯胜杰, 靳丹, 刘美玲, 牟保民. 植物生长调节剂S3307对苗期淹水胁迫下大豆生理特性和显微结构的影响[J]. 作物学报, 2021, 47(10): 1988-2000 DOI:10.3724/SP.J.1006.2021.04233
WANG Shi-Ya, ZHENG Dian-Feng, FENG Nai-Jie, LIANG Xi-Long, XIANG Hong-Tao, FENG Sheng-Jie, JIN Dan, LIU Mei-Ling, MU Bao-Min.
在全球变暖的背景下, 干旱胁迫、盐碱胁迫、温度胁迫和洪涝灾害等非生物胁迫发生频率增加, 严重影响农作物生产[1]。与其他非生物胁迫相比, 洪涝灾害危害程度最大, 约占受灾比例的60% [2]。IPCC AR5报告曾指出, 大气中温室气体含量不断增多将会导致极端降水事件发生的频次和强度增加, 提高洪涝灾害风险[3]。洪涝灾害是一种复杂的胁迫, 是指土壤被水分充分浸透, 影响植物的正常生物活动。淹水对植物生长施加了多种限制, 首先导致根系缺氧, 阻碍根系的生长发育, 致使根系从有氧呼吸变为无氧呼吸, 进而影响地上部的养分吸收、同化物运输等生理生化过程[4]。在淹水胁迫下植株体内积累过量的活性氧(reactive oxygen species, ROS)会对植株造成伤害, 而植物为防御淹水胁迫则会通过提高自身的抗氧化酶系统和非酶抗氧化剂进而有效清除胁迫下产生的活性氧[5], 但未能被及时清除的ROS则会导致细胞膜被破坏, 影响细胞膜的通透性, 加快膜质过氧化进程, 最终导致植株体内生理生化代谢紊乱, 影响正常生长发育[6]。
大豆(Glycine max L.), 是我国重要的粮食作物和经济作物, 富含丰富的蛋白质、植物油和多种植物化学物质[7]。黑龙江作为中国优质大豆的主产区, 种植面积可达全国的40%~44% [8]。但黑龙江是典型的气候变化敏感区和脆弱区, 大豆生长季易发生洪涝灾害, 使大豆生产风险加剧, 最终导致减产[9]。研究发现, 应用植物生长调节剂烯效唑(uniconazole, S3307)可有效提高逆境胁迫下植物的抗逆性[10]。在淹水胁迫条件下, 喷施S3307可抑制大豆株高生长, 缩短节间长, 同时增加基部茎粗, 从而提高大豆的抗倒伏性。于奇等[11]和张洪鹏等[12]研究指出, 叶面喷施S3307可提高大豆和绿豆的光合能力、促进抗氧化酶活性以抵御ROS的过量积累, 进而提高2种作物的抗涝性, 缓解淹水对产量造成的损失。S3307还可诱导逆境胁迫下超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)等保护酶活性的增加, 提高渗透调节物质含量, 降低丙二醛(malondialdehyde, MDA)的生成量和积累量, 提高细胞膜透性, 维持细胞膜的稳定性, 增强细胞膜抵御逆境的能力[13,14,15]。目前关于S3307缓解大豆淹水胁迫的研究集中于花期和鼓粒期, 对苗期的研究相对较少, 因此, 研究V1期淹水胁迫对大豆生理及显微结构的影响及S3307的缓解效应, 以期为化控技术在大豆抗逆生产上的推广提供理论依据。
1 材料与方法
1.1 试验材料
‘垦丰14’由黑龙江省农业科学院佳木斯分院提供; 植物生长调节剂选用烯效唑(S3307), 由黑龙江八一农垦大学植物生长调节剂工程技术研究中心提供。1.2 试验设计与处理
试验于2018—2019年在黑龙江八一农垦大学国家杂粮工程技术研究中心植物培养箱内进行, 采用底部打孔的注塑杯(上口径×底径×高 = 9.0 cm × 5.5 cm × 17.8 cm)进行试验。试验用土为腐殖土、沙子和蛭石按3∶1∶1体积比例混合组成, 每杯装入基质0.38 kg, 覆土0.02 kg。于4月20日选取大小一致、籽粒饱满的大豆种子进行播种, 试验共设3个处理, 4个单元为1个处理, 每杯播种4粒, 出苗后每杯定苗2株, 具体设计见表1。待植株生长至V1期时进行叶面喷施S3307 (浓度为50 mg L-1, 喷施量为225 L hm-2), 并于喷药后第5天进行淹水处理(套杯淹水, 淹水程度以水面高于土壤表面2~3 cm为准), 淹水持续7 d。Table 1
表1
表1试验设计方案
Table 1
| 处理编号 Treatment code | 药剂处理 Pharmaceutical treatment | 水分处理 Water treatment |
|---|---|---|
| CK | 清水喷施 Spray water | 适宜土壤水分 Suitable soil moisture |
| W | 清水喷施 Spray water | 淹水胁迫 Waterlogging stress |
| S | S3307喷施 Spray S3307 | 淹水胁迫 Waterlogging stress |
新窗口打开|下载CSV
1.3 测定项目及方法
1.3.1 形态和叶面积 于淹水胁迫后0、1、3、5、7 d统计株高和茎粗, 同时使用Yaxin-1241叶面积仪测定全株叶面积。1.3.2 逆境生理指标的测定 于淹水后0、1、3、5、7 d进行取样, 取叶片和根系并迅速至于液氮中速冻后置于-80℃中保存, 用于生理指标的测定。按照李合生等[16]方法测定SOD、POD、CAT的酶活性及MDA含量; 按照高俊凤[17]方法测定超氧阴离子产生速率(superoxide anion production rates, O2-); 采用碘化钾法[18]测定过氧化氢(hydrogen peroxide, H2O2)含量; 采用考马斯亮蓝G-250染色法[16]测定可溶性蛋白含量; 采用硫酸蒽酮比色法[19]测定可溶性糖含量; 采用茚三酮比色法[19]测定脯氨酸含量。
1.3.3 石蜡切片制作 于淹水后0、1、3、5、7 d进行取样, 取距离土壤表面1~2 cm处的大豆下胚轴(横切), 固定于FAA固定液中, 固定48 h以上后进行脱水、透明、浸蜡包埋等处理, 采用徕卡切片机(RM2016)制备100~140 mm厚度的切片。切片用番红-固绿染色, 用尼康正置光学显微镜(NIKON ECLIPSE E100)进行观察并使用尼康相机(NIKON DS-U3)进行图像采集分析。采用CaseViewer植物显微成像分析系统观察切片和测量各组织面积。
1.4 数据处理
采用Microsoft Excel 2013录入及整理数据, 用SPSS 25进行方差分析, 用Origin 2018软件绘图。2 结果与分析
2.1 S3307对V1期淹水胁迫下大豆形态指标的影响
由表2可知, 大豆在V1期受淹后株高和叶面积均有不同程度的降低。淹水胁迫处理3、5和7 d时, W处理的株高较CK分别降低15.83%、19.62%和20.94%; 淹水胁迫处理5 d和7 d时, W处理的叶面积较CK分别降低8.67%和11.05%。叶面喷施S3307后, S处理抑制了淹水胁迫下大豆株高生长, 5个时间点均与CK和W处理达显著差异水平, 同时减少了大豆叶面积, 在淹水胁迫处理3、5和7 d时, S处理叶面积较W处理分别显著降低10.00%、7.79%和5.47%。淹水胁迫可增加大豆茎粗, 在淹水胁迫处理3、5和7 d时, W处理较CK分别显著增加4.02%、4.85%和4.54%。叶面喷施S3307可有效增加大豆茎粗, 在淹水胁迫处理0、1和5 d时, S处理较W处理分别显著增加4.70%、3.33%和2.27%。Table 2
表2
表2S3307对V1期淹水胁迫下大豆地上部形态指标的影响
Table 2
| 指标 Trait | 处理 Treatment | 淹水天数 Waterlogging days | ||||
|---|---|---|---|---|---|---|
| 0 d | 1 d | 3 d | 5 d | 7 d | ||
| 株高 Plant height (cm) | CK | 16.32±0.502 a | 16.81±0.336 a | 20.09±0.360 a | 21.31±0.523 a | 21.82±0.666 a |
| W | 15.96±0.471 a | 16.81±0.756 a | 16.91±0.724 b | 17.13±0.535 b | 17.25±0.544 b | |
| S | 12.87±0.694 b | 13.22±0.305 b | 13.53±0.510 c | 13.80±0.317 c | 14.05±0.464 c | |
| 茎粗 Stem diameter (mm) | CK | 2.82±0.078 b | 2.86±0.067 b | 2.86±0.015 b | 2.87±0.031 c | 2.94±0.050 b |
| W | 2.84±0.075 b | 2.90±0.036 b | 2.98±0.042 a | 3.02±0.020 b | 3.08±0.015 a | |
| S | 2.98±0.030 a | 3.00±0.038 a | 3.04±0.040 a | 3.09±0.015 a | 3.11±0.025 a | |
| 叶面积 Leaf areas (mm2) | CK | 3598.50±239.881 a | 3689.56±199.213 a | 3576.06±208.030 a | 3837.35±67.895 a | 3956.45±55.776 a |
| W | 3289.21±143.851 ab | 3384.60±141.800 ab | 3542.72±81.976 a | 3505.03±45.032 b | 3519.19±33.713 b | |
| S | 3129.26±173.781 b | 3239.22±108.931 b | 3188.17±68.382 b | 3232.09±111.343 c | 3326.58±87.010 c | |
新窗口打开|下载CSV
2.2 S3307对V1期淹水胁迫下大豆下胚轴显微结构的影响
通气组织是植物茎内由充满空气的气腔组成, 从下胚轴横切结构可以看出, 淹水胁迫下W和S处理均刺激大豆下胚轴通气组织的形成。淹水胁迫0 d时, W处理和S处理从形态上看无明显变化(图2-1, 3), 从显微切片来看, W处理下胚轴中皮层细胞紧密排列(图2-2), S处理的下胚轴皮层细胞出现破裂(图2-4), S的通气组织数量和通气组织面积分别是W处理的1.53倍和1.54倍(图1-A)。随淹水胁迫天数延长, 2处理均生成不规则的溶生气腔, 并随处理时间延长, 气腔逐渐融合并扩大, 且呈无规则的崩解状。2个处理对淹水胁迫的响应不同, W处理在胁迫初期(1 d)已经形成大量的气腔(图2-6), 淹水胁迫中期(3 d), 气腔数量不断增多、面积逐渐扩大(图1-A, B), 随胁迫时间延长, 在淹水胁迫后期(5 d和7 d), 下胚轴结构出现破裂(图2-15, 16和图2-21, 22), 且相邻的多个气腔相互融合, 形成不规则的大气腔, 以致通气组织数量减少(图1-A, B)。而S处理在淹水胁迫1 d和3 d时, 虽有气腔形成, 但形成数量和面积明显小于W处理(图2-8, 14), 与W处理相比, 随胁迫时间延长, 下胚轴结构有明显的破裂, 气腔面积逐渐扩大(图2-20, 26)。图1
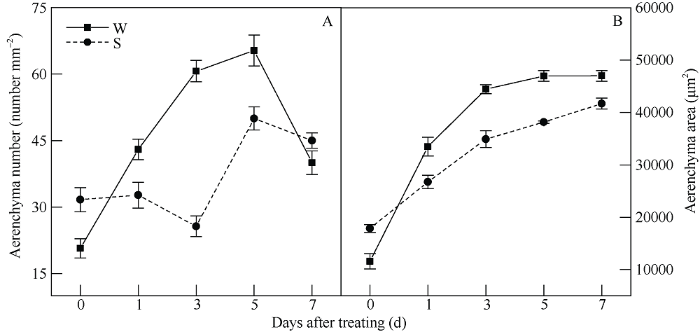 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1S3307对V1期淹水胁迫下大豆下胚轴通气组织数量和面积的影响
处理同
Fig. 1Effects of S3307 on aerenchyma number and area in leaves and roots under waterlogging stress at V1 stage in soybean
Treatments are the same as those given in
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2S3307对V1期淹水胁迫下大豆下胚轴显微结构的影响
1和2为淹水处理0 d的W处理; 3和4为淹水胁迫处理0 d的S处理; 5和6为淹水胁迫处理1 d的W处理; 7和8为淹水胁迫处理1 d的S处理; 9、10和11为淹水胁迫处理3 d的W处理; 12、13和14为淹水胁迫处理3 d的S处理; 15、16和17为淹水胁迫处理5 d的W处理; 18、19和20为淹水胁迫处理5 d的S处理; 21、22和23为淹水胁迫处理7 d的W处理; 24、25和26为淹水胁迫处理7 d的S处理。处理同
Fig. 2Effects of S3307 on microstructure of hypocotyl under waterlogging stress at V1 stage in soybean
1 and 2 show W treatment for 0 day of waterlogging treatment; 3 and 4 show S treatment for 0 day of waterlogging treatment; 5 and 6 show W treatment for 1 day of waterlogging treatment; 7 and 8 show S treatment for 1 day of waterlogging treatment; 9, 10 and 11 show W treatment for 3 days of waterlogging treatment; 12, 13, and 14 show S treatment for 3 days of waterlogging treatment; 15, 16, and 17 show W treatment for 5 days of waterlogging treatment; 18, 19, and 20 show S treatment for 5 days of waterlogging treatment; 21, 22, and 23 show W treatment for 7 days of waterlogging treatment; 24, 25, and 26 show S treatment for 7 days of waterlogging treatment. Treatments are the same as those given in
2.3 S3307对淹水胁迫下大豆叶片和根系膜质过氧化程度的影响
由图3可知, 随淹水胁迫时间延长, 大豆叶片和根系中MDA含量均呈逐渐上升趋势。淹水胁迫前期(0 d和1 d), W处理和S处理叶片中MDA含量与CK相比差异不显著, 在3、5和7 d时, 淹水胁迫显著增加了W处理的叶片中的MDA含量, 较CK分别增加45.91%、34.93%和44.24%; 叶面喷施S3307后, 可有效缓解淹水胁迫造成的伤害, S处理较W处理分别显著降低13.02%、18.59%和16.41%。图3
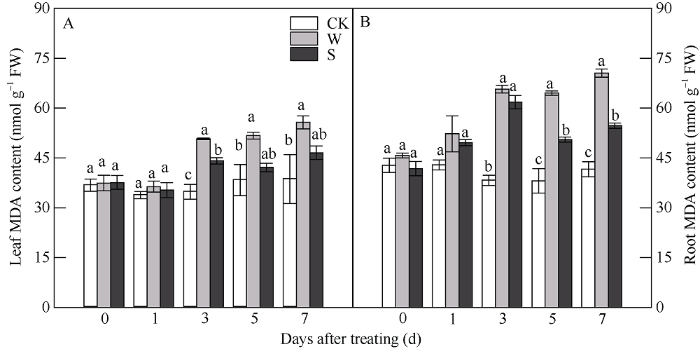 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3S3307对V1期淹水胁迫下大豆叶片和根系MDA含量的影响
处理同
Fig. 3Effects of S3307 on MDA content of leaves and roots under waterlogging stress at V1 stage in soybean
Treatments are the same as those given in
在淹水胁迫1 d时, W处理的大豆根系中MDA含量与CK和S相比有所增加, 但未达显著差异水平。淹水处理3、5和7 d时, W处理与CK处理相比根系中MDA含量分别增加71.56%、69.44%和69.51%, 达显著差异水平; 叶面喷施S3307后, S处理与CK相比虽有增加, 但明显抑制了淹水胁迫下大豆根系中MDA含量的积累, 在5 d和7 d时与W处理达显著差异水平, 较W处理分别显著降低21.74%和22.41%。
2.4 S3307对淹水胁迫下大豆叶片和根系O2-产生速率和H2O2含量的影响
淹水胁迫下, 大豆叶片和根系中O2-产生速率和H2O2含量呈逐渐上升趋势(图4)。在淹水胁迫初期(0 d和1 d), W处理与CK相比无明显变化, 但S处理显著抑制了叶片内O2-产生速率和H2O2含量的增加, 对于根系来说效果不明显。淹水胁迫3、5和7 d时, W处理导致叶片和根系中O2-产生速率和H2O2含量过量积累, 与CK相比差异显著, 叶面喷施S3307后缓解了上述指标的积累, S处理与W处理相比显著降低, 但仍显著高于CK。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4S3307对V1期淹水胁迫下大豆叶片和根系O2-产生速率和H2O2含量的影响
处理同
Fig. 4Effects of S3307 on O2- production rate and H2O2 content of leaves and roots under waterlogging stress at V1 stage in soybean
Treatments are the same as those given in
2.5 S3307对淹水胁迫下大豆叶片和根系酶促抗氧化防御系统的影响
由图5可知, 淹水胁迫下, 大豆叶片和根系中SOD、POD和CAT活性均呈先升高后下降趋势, 在3 d时达最大值, 随胁迫时间延长, 在淹水胁迫后期上述指标均有所降低。淹水胁迫5 d时, W处理的大豆叶片POD、CAT活性和根系的SOD活性与CK相比明显降低, 但未达显著差异水平; 淹水胁迫7 d时, W处理的大豆叶片SOD、POD活性和根系SOD活性显著低于CK, 其他处理与CK相比差异不显著。叶面喷施S3307后, 可进一步促进叶片和根系中上述3个指标的酶活性, 并始终维持较高水平。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5S3307对V1期淹水胁迫下大豆叶片和根系酶促抗氧化防御系统的影响
处理同
Fig. 5Effects of S3307 on enzymatic antioxidant system of leaves and roots under waterlogging stress at V1 stage in soybean
Treatments are the same as those given in
2.6 S3307对淹水胁迫下大豆叶片和根系渗透调节物质的影响
由图6可知, 淹水胁迫条件下, W处理的大豆叶片和根系中可溶性蛋白、脯氨酸和可溶性糖含量呈先上升后下降的趋势, 在淹水胁迫3 d时达最大值。随淹水胁迫时间延长, 淹水胁迫5 d时, W处理的叶片中可溶性蛋白、脯氨酸含量和根系中可溶性蛋白、脯氨酸、可溶性糖含量与CK相比有所降低, 其中叶片可溶性蛋白和根系中可溶性蛋白、脯氨酸、可溶性糖含量与CK相比达显著差异水平; 淹水胁迫7 d时, W处理的叶片和根系中上述3个指标均显著低于CK。叶面喷施S3307后, 有效增加了淹水胁迫下叶片和根系中可溶性蛋白、脯氨酸、可溶性糖含量, 在淹水胁迫0 d时, 除叶片中可溶性蛋白和可溶性糖外, 与CK相比均达显著差异水平, 且随胁迫时间延长, 维持较高水平, 在淹水胁迫处理7 d时, S处理各指标含量虽有降低, 但始终显著高于W处理。图6
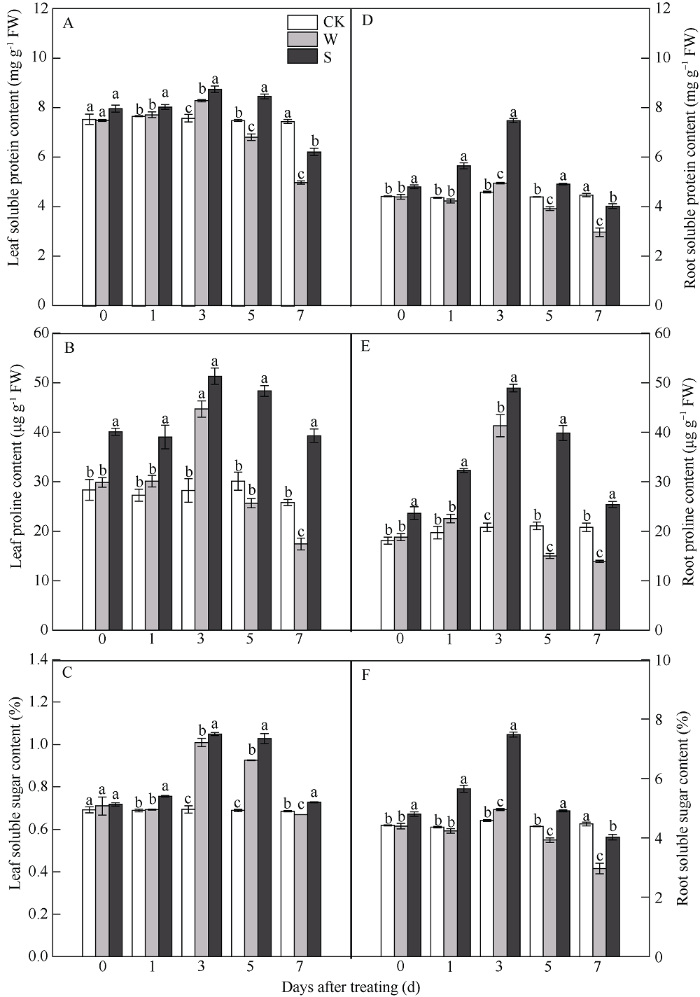 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6S3307对V1期淹水胁迫下大豆叶片和根系渗透调节物质的影响
处理同
Fig. 6Effects of S3307 on osmotic regulating substance of leaves and roots under waterlogging stress at V1 stage in soybean
Treatments are the same as those given in
3 讨论
3.1 V1期淹水胁迫及S3307处理对大豆地上部形态指标的影响
淹水是一种环境胁迫因子, 对大豆株型的影响非常明显[20]。相关研究曾指出, 淹水胁迫可抑制大豆植株的正常生长, 表现为降低大豆株高, 增加茎基部茎粗, 减少叶面积等[10,21], 这与本试验研究结果一致。淹水胁迫前期, 对大豆植株影响效果不明显, 随胁迫时间延长, 株高生长受到抑制, 进而影响地上部正常发育, 导致叶面积降低(表2)。在淹水胁迫下, 植物会根据所受环境刺激形成应激机制, 本试验结果表明, 淹水胁迫导致大豆植株茎部增粗, 在淹水胁迫3、5和7 d时增加效果显著。田一丹等[22]研究发现, 在淹水胁迫条件下, 大豆根与茎在土壤层上方和水面层之间的部位会明显变粗, 说明大豆在应对淹水胁迫时, 茎部形成了发达的通气组织, 促进了茎部增粗。S3307作为一种植物生长延缓剂, 会进一步降低淹水胁迫下植物株高和叶面积[20], 这一点在本试验中也可得到验证, 在整个胁迫期间, S3307处理的大豆植株株高、茎粗和叶面积与CK相比均达显著差异水平, 与淹水处理相比进一步降低大豆株高并增加茎粗, 有利于提高大豆植株的抗倒伏能力, 进而提高大豆的耐涝性。3.2 V1期淹水胁迫及S3307处理对大豆下胚轴显微结构的影响
对于旱田作物来说, 淹水胁迫通常导致植物缺氧, 并且严重抑制植物的正常生长发育, 植物为应对淹水胁迫, 已进化出多种应激特性, 通气组织的形成就是其中一种[23,24,25]。研究发现, 植物通气组织首先在根系与茎的连接部位形成, 随之向根系和茎上部扩展, 这可能是由于前体物质1-氨基环丙烷-1-羧酸(ACC)常在近地表处氧化生成乙烯, 而乙烯是通气组织形成过程中的信号物质, 因此植物下胚轴部位最先生成通气组织[26]。王寒等[27]研究发现, 植物融生气腔数量或通气组织的发达程度与植株耐涝性密切相关。本试验采用耐涝性较强的品种垦丰14 [12,28], 在淹水胁迫1 d时, 下胚轴部开始形成通气组织, 并随淹水胁迫时间延长, 通气组织数量和通气组织面积随胁迫时间延长逐渐增加, 从而促进根部向地上部氧气的供应, 这与Takahashi等[23]研究结果一致。叶面喷施S3307后, 可促进通气组织的形成, 使大豆更快适应淹水环境, 随胁迫时间延长, 通气组织面积虽然小于淹水胁迫处理的面积, 但多个气腔相互融合形成大气腔, 有助于提高氧气的扩散能力并促进养分的运输, 保证地上部的正常生长发育。3.3 V1期淹水胁迫及S3307处理对大豆叶片和根系膜质过氧化程度的影响
淹水胁迫导致植物膜质过氧化的过量积累常被认为是逆境胁迫下细胞膜损伤的重要表征[29], MDA是膜质过氧化产物, MDA含量的高低可直接逆境下细胞膜的稳定性, 从而间接反映植物质膜透性的受损程度[30,31]。本试验结果表明, 随淹水胁迫时间延长, 大豆叶片和根系中MDA含量呈逐渐上升趋势, 且根系中MDA含量的积累大于叶片, 可见根系的受抑制程度大于叶片, 这与于奇等[11]和齐玉军等[32]研究结果一致。在逆境条件下应用S3307可缓解胁迫对作物造成的伤害。研究曾指出, 叶面喷施S3307可提高红小豆抵御低温胁迫的能力[33,34], 外源喷施S3307可提高绿豆和大豆的耐涝性[10,11], 幼苗期喷施S3307可提高大豆的耐盐碱性[35]等, 上述研究虽与本试验喷施S3307时间不同, 但效果一致, 均能提高作物抵抗抗逆的能力, 缓解逆境对作物造成的伤害。本研究发现, 叶面喷施S3307后, 有效缓解了淹水胁迫下大豆叶片和根系中MDA含量的增加, 减轻淹水胁迫下大豆叶片和根系中膜质过氧化作用, 保证大豆在苗期的正常生长和生理代谢, 有利于提高苗期大豆的耐涝性。3.4 V1期淹水胁迫及S3307处理对大豆叶片和根系ROS代谢的影响
植物在正常条件下, 体内ROS的产生和清除处于动态平衡状态, 主要作为信号分子调节植物的生长发育[36]; 当植物受淹水胁迫时, 会导致氧化产生过量的ROS、O2?和过氧化氢H2O2等, ROS的过量增加会对细胞膜脂质过氧化、蛋白质和核糖核酸等造成氧化损伤, 从而导致细胞造成氧化损伤, 最终导致细胞死亡[37]。因此, ROS在植物体内具有双重功能, 是参与调节植物生长发育和响应逆境胁迫的重要指标。本试验结果表明, 淹水胁迫导致V1期大豆叶片和根系中O2?和H2O2含量显著增加, 随胁迫时间延长, 对大豆叶片和根系造成伤害越大, 上述指标的上升趋势越明显。S3307具有降低ROS积累、减轻细胞膜损伤, 提高作物在逆境下耐受性的作用[38]。本试验结果表明, S3307可有效减少淹水胁迫下V1期大豆叶片和根系内O2-产生速率和H2O2含量的积累, 说明S3307对维持ROS平衡具有积极作用。3.5 V1期淹水胁迫及S3307处理对大豆叶片和根系酶促抗氧化防御系统的影响
在非生物胁迫下, 植物均具有解毒能力, 在逆境下积累的ROS可通过抗氧化酶系统进行清除[37]。Garcia等[39]研究发现, 耐涝性强的植物体内抗氧化酶活性可清除淹水胁迫下积累的活性氧, 其中SOD、POD和CAT是主要的清除剂。SOD是清除ROS的第一道防线, 它可催化O2-生成H2O2和H2O。在本研究中, 叶片和根系中SOD活性呈先升高后降低的趋势, 在淹水胁迫3 d时达最大值, 随胁迫时间延长显著降低, 且根系中SOD活性下降趋势大于叶片, SOD活性的下降可能是由于O2-的过量积累, 促使SOD的清除效率降低, 从而导致SOD活性随胁迫时间延长而下降。而在徐涛等[40]研究中, 玉米在淹水胁迫1、3、5 d时SOD活性呈显著升高趋势, 在7 d时有所下降, 这表明不同作物在淹水胁迫下的耐受性不同。研究表明, 淹水胁迫下应用S3307可提高抗氧化酶活性[10], 这与本试验结果一致。POD和CAT具有清除逆境胁迫产生的H2O2的作用, 进而保持ROS代谢平衡, 减轻逆境对植物造成的伤害[41]。本试验研究发现, 淹水胁迫条件下, V1期大豆叶片和根系随淹水胁迫时间延长, POD和CAT活性呈先升高后下降趋势, 在淹水胁迫中期(3 d)时, POD和CAT活性达最大值, 说明POD和CAT在3 d时清除能力最强, 随淹水胁迫时间的延长, 积累的H2O2含量逐渐增加, 导致POD和CAT的清除能力减弱, 在淹水胁迫7 d时显著降低, 与CK相比达显著差异水平。研究曾指出, 适当浓度的S3307可提高作物体内的抗逆酶活性, 增强植株抗氧化能力[34], 本研究得出相同结果, 叶面喷施S3307后, 大豆叶片和根系中POD和CAT始终维持较高活性, 提高ROS的清除效率, 减轻氧化损伤。3.6 V1期淹水胁迫及S3307处理对大豆叶片和根系渗透调节物质的影响
渗透调节是作物适应逆境适应性调节的重要机制, 植物在逆境胁迫下主要通过可溶性糖、可溶性蛋白和脯氨酸3种渗透调节物质含量的变化来适应环境[42]。植物在逆境下, 可大量积累渗透调节物质, 使组织水势下降后维持细胞的正常状态, 进而保证植物生理生化等代谢活动的正常进行, 从而提高作物的抗逆性[43]。姜颖等[44]研究发现, 逆境胁迫下应用S3307可增加作物渗透调节物质的含量, 提高抵御逆境能力, 这与本试验研究结果类似。本研究发现, 淹水胁迫前期(1 d)大豆叶片造成伤害较小, 与CK相比无显著差异, 这与项洪涛等[45]研究结果有所不同, 其研究表明, 淹水胁迫1 d时, 小豆叶片脯氨酸、可溶性糖和可溶性蛋白含量显著增加, 这表明不同作物、不同时期对淹水胁迫的响应存在差别, 大豆在淹水胁迫前期的耐涝性可能强于小豆。随淹水胁迫时间延长, W处理促进了大豆叶片和根系内渗透调节物质含量的增加, 不同部位渗透调节物质积累量不同, 叶片中可溶性蛋白和脯氨酸含量高于根系, 根系可溶性糖含量大于叶片, 而叶片和根系中渗透调节物质含量均在淹水胁迫3 d时达最大值。叶面喷施S3307后可进一步促进了淹水胁迫下大豆叶片和根系中可溶性糖、可溶性蛋白和脯氨酸含量。但喷施S3307时间不同对作物产生的效果不同, 低温处理前1 d喷施S3307, 在低温胁迫3~4 d时可显著增加小豆的可溶性糖和可溶性蛋白含量[33], 而本试验中, 淹水胁迫处理5 d前喷施S3307, 有效促进了正常条件下大豆叶片内渗透调节物质含量的增加, 在淹水胁迫前期维持较高的渗透调节物质含量, 进而增强大豆在淹水胁迫下的抗涝性, 随淹水胁迫时间延长S处理虽有降低, 但显著高于W处理, 说明叶片喷施S3307可促进大豆在淹水胁迫下正常生长。4 结论
V1期淹水胁迫降低了大豆株高和叶面积, 增加了大豆茎粗、通气组织数量和面积, 导致ROS代谢紊乱, 产生氧化应激, 加剧膜质过氧化程度, 同时诱导了SOD、POD和CAT活性的增强, 并提高了渗透调节物质含量。叶面喷施S3307有助于提高大豆植株的抗倒伏能力, 表现为降低株高、减小叶面积和增加茎粗等, 叶喷S3307可增加通气组织面积, 促进气腔融合, 有助于氧气和养分的运输, 维持地上和地下的正常生长, 同时提高抗氧化酶活性、增加渗透调节物质含量以清除过量积累的ROS和膜质过氧化物, 缓解淹水胁迫对大豆植株造成的伤害, 提高大豆的抗涝性。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIPMID [本文引用: 1]

Reactive oxygen species (ROS) regulate plant growth and development. ROS are kept at low levels in cells to prevent oxidative damage, allowing them to be effective signaling molecules upon increased synthesis. In plants and animals, NADPH oxidase/respiratory burst oxidase homolog (RBOH) proteins provide localized ROS bursts to regulate growth, developmental processes, and stress responses. This review details ROS production via RBOH enzymes in the context of plant development and stress responses and defines the locations and tissues in which members of this family function in the model plant Arabidopsis thaliana. To ensure that these ROS signals do not reach damaging levels, plants use an array of antioxidant strategies. In addition to antioxidant machineries similar to those found in animals, plants also have a variety of specialized metabolites that scavenge ROS. These plant specialized metabolites exhibit immense structural diversity and have highly localized accumulation. This makes them important players in plant developmental processes and stress responses that use ROS-dependent signaling mechanisms. This review summarizes the unique properties of plant specialized metabolites, including carotenoids, ascorbate, tocochromanols (vitamin E), and flavonoids, in modulating ROS homeostasis. Flavonols, a subclass of flavonoids with potent antioxidant activity, are induced during stress and development, suggesting that they have a role in maintaining ROS homeostasis. Recent results using genetic approaches have shown how flavonols regulate development and stress responses through their action as antioxidants.
DOIPMID [本文引用: 1]
Soybean is the important crop with abundant protein, vegetable oil, and several phytochemicals. With such predominant values, soybean is cultivated with a long history. However, flooding and drought stresses exert deleterious effects on soybean growth. The present review summarizes the morphological changes and affected events in soybean exposed to such extreme-water conditions. Sensitive organ in stressed soybean at different-developmental stages is presented based on protein profiles. Protein quality control and calcium homeostasis in the endoplasmic reticulum are discussed in soybean under both stresses. In addition, the way of calcium homeostasis in mediating protein folding and energy metabolism is addressed. Finally, stress response to flooding and drought is systematically demonstrated. This review concludes the recent findings of plant response to flooding and drought stresses in soybean employed proteomic approaches.Soybean is considered as traditional-health food because of nutritional elements and pharmacological values. Flooding and drought exert deleterious effects to soybean growth. Proteomic approaches have been employed to elucidate stress response in soybean exposed to flooding and drought stresses. In this review, stress response is presented on organ-specific manner in the early-stage plant and soybean seedling exposed to combined stresses. The endoplasmic reticulum (ER) stress is induced by both stresses; and stress-response in the ER is addressed in the root tip of early-stage soybean. Moreover, calcium-response processes in stressed plant are described in the ER and in the cytosol. Additionally, stress-dependent response was discussed in flooded and drought-stressed plant. This review depicts stress response in the sensitive organ of stressed soybean and forms the basis to develop molecular markers related to plant defense under flooding and drought stresses.Copyright © 2017 Elsevier B.V. All rights reserved.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 4]
URL [本文引用: 3]
URL [本文引用: 3]
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}