 ,1,2,4,**, 张佳蕾1,2,**, 郭峰1,2, 唐朝辉1,2, 杨莎1,2, 彭振英1,2, 孟静静1,2, 崔利3, 李新国,1,2,4,*, 万书波,1,2,4,*
,1,2,4,**, 张佳蕾1,2,**, 郭峰1,2, 唐朝辉1,2, 杨莎1,2, 彭振英1,2, 孟静静1,2, 崔利3, 李新国,1,2,4,*, 万书波,1,2,4,*Effects of interaction between calcium and nitrogen fertilizers on dry matter, nitrogen accumulation and distribution, and yield in peanut
WANG Jian-Guo,1,2,4,**, ZHANG Jia-Lei1,2,**, GUO Feng1,2, TANG Zhao-Hui1,2, YANG Sha1,2, PENG Zhen-Ying1,2, MENG Jing-Jing1,2, CUI Li3, LI Xin-Guo,1,2,4,*, WAN Shu-Bo,1,2,4,*通讯作者: * 李新国, E-mail:xinguol@163.com;万书波, E-mail:wanshubo2016@163.com
第一联系人:
收稿日期:2020-08-12接受日期:2021-01-21
| 基金资助: |
Corresponding authors: * E-mail:xinguol@163.com;E-mail:wanshubo2016@163.com
First author contact:
Received:2020-08-12Accepted:2021-01-21
| Fund supported: |
作者简介 About authors
王建国,E-mail:shanshanyilang@163.com

摘要
为探讨钙肥和氮肥施用量对花生干物质和氮素积累分配及产量的影响, 本研究以花育25为试验材料, 设置0和600 kg hm-2 (Ca0、Ca600) 2个钙肥水平, 0、75、150、225、300 kg hm-2 (N0、N75、N150、N225、N300) 5个氮肥水平, 研究不同试验样地增钙减氮对花生干物质积累和氮素积累与分配、产量及其构成因素的影响。结果表明, 与Ca 0相比, Ca600条件下花生干物质积累量显著升高, 济阳(JY)和饮马泉(YMQ)分别提高了13.5%和12.6%。与N0相比, 济阳(JY)各施氮处理花生植株干物质积累量分别提高了12.8%、17.7%、26.3%和21.0%, 饮马泉(YMQ)分别提高了16.7%、28.4%、24.9%和22.9%。花生干物质和氮素吸收积累动态曲线均符合Logistic模型, 济阳和饮马泉花生植株氮素最大累积量(Ym)分别在Ca600N225、Ca600N150处理下获得, 与平均值相比, 花生植株氮素最大累积速率(Vm)分别提高了12.4%和10.6%, 最大累积量分别提高了14.9%和13.7%, 快速累积持续时间(T)分别延长了2.3%和3.1%; 氮素快速积累期起始时期(t1)比干物质积累分别提早了7.5 d和9.4 d。济阳Ca600N225和饮马泉Ca600N150、Ca600N225处理花生荚果产量显著高于其他处理。钙氮互作效应对花生产量的影响显著。钙肥增施是提高氮肥利用效率, 增加花生结果数和百果重、促进稳产高产的重要途径。本试验区域减肥稳产增效栽培中最优施肥方案可采用钙肥600 kg hm-2 + 氮肥75 kg hm-2; 高产高效栽培中最优施肥方案可采用钙肥600 kg hm-2 + 氮肥150~225 kg hm-2。
关键词:
Abstract
The purpose of this study was to investigate the effects of calcium and nitrogen fertilizer on dry matter, nitrogen accumulation and distribution, and yield in peanut. A field experiment was conducted using Huayu 25 with two calcium (Ca) rates (0 and 600 kg hm-2), and five nitrogen (N) rates (0, 75, 150, 225, and 300 kg hm-2) in Jiyang (JY) and Yinmaquan (YMQ) in 2019. The results showed that compared with Ca0, the dry matter accumulation of peanuts under Ca600 treatment was significantly increased by 13.5% in the Jiyang, by 12.6% in Yinmaquant. Compared with N0, nitrogen fertilizer significantly promoted the accumulation of dry matter in peanut, and the N75, N150, N225, N300 treatments increased dry matter accumulation by 12.8%, 17.7%, 26.3%, 21.0% in Jiyang and 16.7%, 28.4%, 24.9%, 22.9% in Yinmaquan, respectively. The dynamic curve of peanut dry matter and nitrogen absorption accumulation conformed to the Logistic model, and the maximal nitrogen accumulation (Ym) was obtained under Ca600N225treatment in Jiyang and Ca600N150 treatment Yinmaquan. Compared to the average, the maximal speed of accumulation (Vm), the maximal biomass, duration of rapid accumulation (T), were increased at two plots by 12.4% and 10.6%, 14.9% and 13.7%, 2.3% and 3.1%, respectively. The starting date of rapid accumulation period (t1) was 7.5 days and 9.4 days earlier for nitrogen than for dry matter, indicated that the nutrient absorption of peanut was the premise of dry matter accumulation. The pod yield of peanut of Ca600N225 in Jiyang and Ca600N150 and Ca600N225 in Yinmaquan was higher than other treatments. Effects of interaction between calcium rates and nitrogen rates had a significant impact on peanut yield. Increasing calcium fertilizer application was an important way to improve nitrogen use efficiency, which increased pod number and 100-pod weight and promoted stably high yield in peanut. In this study, calcium fertilizer (600 kg hm-2) and nitrogen fertilizer (75 kg hm-2) were applied as the optimal fertilization scheme in the cultivation of less-fertilizer, stable-yield, and improved-efficiency treatment, while calcium fertilizer (600 kg hm-2) and nitrogen fertilizer (150-225 kg hm-2) as high-yield and high-efficiency treatment.
Keywords:
PDF (485KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王建国, 张佳蕾, 郭峰, 唐朝辉, 杨莎, 彭振英, 孟静静, 崔利, 李新国, 万书波. 钙与氮肥互作对花生干物质和氮素积累分配及产量的影响. 作物学报[J], 2021 , 47(9): 1666-1679 DOI:10.3724/SP.J.1006.2021.04186
WANG Jian-Guo, ZHANG Jia-Lei, GUO Feng, TANG Zhao-Hui, YANG Sha, PENG Zhen-Ying, MENG Jing-Jing, CUI Li, LI Xin-Guo, WAN Shu-Bo.
花生是我国重要的油料和经济作物, 种植面积约占世界的20%, 产量占世界总产约40% [1]。2018年全国花生种植面积为461.9万公顷、产量1733.2万吨, 其中山东省种植面积69.5万公顷、产量306.7万吨, 分别占全国的15.1%和17.7% (国家统计局), 均为全国第2位。山东省是我国花生重要主产区, 同时又是氮肥投入较多的省份, 花生平均施氮量为181.0 kg hm-2左右[2], 高产创建氮肥投入量为515.7 kg hm-2, 远高于花生氮肥需求量[3]。过量的氮肥施用带来了一系列的负面影响, 如生产成本增加、肥料利用率低、生态环境污染, 土壤可持续生产力降低[4,5,6,7]。因此, 如何在保障花生产量的同时, 减少氮肥施用量是目前亟待解决的一项新科学问题。
作物高产以生物量积累为前提, 而生物量累积是以养分吸收为基础[8]。钙肥和氮肥的施用是影响花生生长发育和产量形成的重要栽培措施[9,10,11,12]。氮素与花生植株生理代谢、产量及品质密切相关[13,14]。施氮显著改善花生叶片光合性能, 提高茎、叶及荚果干物质积累量[15,16,17,18]; 施氮量在0~150 kg hm-2时, 不同花生品种荚果产量均随施氮量的增加呈现先增加后减小的趋势[19]。施钙可促进花生干物质的积累, 减少花生空、秕果数, 增加饱果数和百果重, 进而提高荚果产量[11], 生产中推荐的钙肥施用量为210~600 kg hm-2 [10-11,20]。研究表明, 盆栽试验中高氮肥条件下, 基施钙肥(CaO 450 kg hm-2)显著促进了根系的生长发育, 并提高了荚果产量; 花生氮肥总量减施28.6%, 施肥方式为氮、钙肥基施+花针期追施氮肥, 其荚果产量小幅提高, 且荚果氮素积累量降低不明显[21]。随施钙量增加, 南方红壤旱地花生生殖器官氮素积累量和分配率提高[22]。豫南沙姜黑土区氮、钙肥互作能够影响花生产量和品质[20]。目前, 前人关于钙、氮肥对花生干物质和产量及氮素积累的影响已有大量研究, 但主要以单因素为主, 且研究多集中于盆栽试验和特定土壤类型, 少量钙、氮耦合研究也多是关注钙、氮常规用量配施下作物产量和品质的变化, 而单粒精播栽培中钙、氮互作对花生干物质、产量和氮素积累的影响鲜见报道。因此, 发挥钙肥和氮肥对花生生长最佳的互作效应, 实现以钙促氮、降低氮肥用量, 协调干物质积累与产量的关系, 在保障花生稳产、高产等方面具有重要的研究意义。本试验在前人研究的基础上采用单粒播种方式, 在不同试验样地设置不同的施钙量和施氮量, 研究钙肥和氮肥及其互作对花生干物质和氮素积累分配及荚果产量的影响, 探讨二者对花生产量构成的影响, 旨在为花生减氮协同增效栽培提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2019年在山东省农业科学院济阳综合试验示范基地和生物技术研究中心饮马泉试验基地进行。济阳基地位于山东省济南市济阳区太平镇(36°58′N, 116°58′E, 海拔18.1 m), 土壤质地为沙壤土, 含有机质16.7 g kg-1、碱解氮104.3 mg kg-1、有效磷53.2 mg kg-1、速效钾102.0 mg kg-1、交换性钙21.4 g kg-1, 土壤pH 8.0; 饮马泉基地位于山东省济南市历城区全福街道(36°45′N, 117°05′E, 海拔39 m), 土壤质地为壤土, 含有机质13.7 g kg-1、碱解氮80.2 mg kg-1、有效磷42.2 mg kg-1、速效钾238.5 mg kg-1、交换性钙5.2 g kg-1, 土壤pH 5.5。2个试验区均属温带季风气候。2019年济阳试验区花生播种后0~150 d内降雨量、平均气温、平均日照时数分别为316.4 mm、24.8℃、7.7 h; 饮马泉试验区分别为304.7 mm、27.8℃、8.2 h。1.2 试验设计
大田试验采用二因素裂区设计。设置钙肥水平为主区, 氮肥水平为副区。设0 (Ca0)、600 (Ca600) kg hm-2 2个钙肥水平, 0 (N0)、75 (N75)、150 (N150)、225 (N225)、300 (N300) kg hm-2 5个氮肥水平, 共10个处理, 每个处理3次重复, 小区面积为66.6 m2。钙肥为氧化钙(CaO含量99%), 氮肥为尿素(N含量46.7%)。磷钾肥为磷酸二氢钾, 用量为240 kg hm-2 (P2O5含量52%、K2O含量34%)。所有肥料均采用基肥一次性施用的方式。先施用氮肥、磷钾肥, 旋耕, 再施入钙肥, 旋耕起垄。花生垄距80 cm, 垄面宽50 cm, 垄高10 cm。单粒精播密度为25万株 hm-2, 穴距10 cm, 每穴播1粒种子, 覆膜栽培。济阳基地于2019年4月30日播种, 9月3日收获; 饮马泉基地于2019年5月20日播种, 9月18日收获。其他管理措施均按大田常规方式进行。1.3 测定项目与方法
1.3.1 产量及构成因素测定 于成熟期各处理分别选取代表性植株, 合计16株, 摘下所有荚果, 及时晾晒、考种、测产, 并测定晒干后的荚果含水量。1.3.2 干物质量和氮素测定 济阳取样时间分别为播种后32 d (苗期)、55 d (花针期)、74 d (结荚期)、98 d (饱果期)和127 d (成熟期); 饮马泉为22 d (苗期)、43 d (花针期)、67 d (结荚期)、88 d (饱果期)和122 d (成熟期)。按小区连续选取花生植株6株, 将每株分为根系、茎秆、叶片、果针和荚果, 105℃杀青30 min后, 于80℃烘干至恒重, 测定干物质。将烘干的花生植株各器官用高速万能粉碎机进行粉碎, 过80目筛, 采用H2SO4-H2O2消煮, 利用San++连续流动分析仪测定氮含量。
单位面积干物质积累量(kg hm-2)=不同时期单株干物质重(g 株-1)×种植密度(株 hm-2)×10-6
植株各器官氮素积累量(mg 株-1)=植株各器官干物质重(g 株-1)×植株各器官氮素含量(mg g-1)
单位面积氮素积累量(kg hm-2)=不同时期单株氮素积累量(mg 株-1)×种植密度(株 hm-2)×10-6
收获指数=成熟期荚果干物质量/植株干物质总量
利用Logistic曲线拟合花生干物质和氮素积累量。其中,
式中, Y为积累量(kg hm-2), K为理论最大积累量(kg hm-2), t为生长天数(d)[23], a、b为常数。分别对公式(1)求一阶、二阶导数, 得到Logistic生长曲线的最大积累速率出现时间(tm)、最大积累速率(Vm)、快速积累期起始时期(t1)、快速积累期终止时期(t2)[23,24]。
1.4 数据处理
用Microsoft Excel 2007整理数据并作图, 用IBM SPSS Statistics 21软件分析数据, 采用LSD法进行显著性分析。2 结果与分析
2.1 花生干物质积累动态
由表1可知, 增施钙肥处理花生干物质最大累积量(Ym)升高, 随着施氮量增加, 不同试验样地的变化不同。在济阳增施CaO 600 kg hm-2, 花生干物质最大积累速率(Vm)提高14.3%, 快速积累期起始时期(t1)推后, 快速积累持续期(T)缩短; 随着施氮量增加, 花生干物质最大累积量、最大积累速率和出现时间、快速积累期终止时期(t2)和快速积累持续期呈先升高后降低趋势, 快速积累期起始时期提前; 施钙量和施氮量互作下, 花生干物质最大累积量在Ca600N225获得最大值, 其最大累积速率为329.2 kg hm-2 d-1, 快速累积持续时间为33.7 d, 与各处理平均值相比, 花生干物质最大累积量提高了14.9%, 最大累积速率提高了12.4%, 快速累积持续时间延长了2.3%。在饮马泉增施CaO 600 kg hm-2, 花生干物质最大积累速率提高9.0%, 快速积累期起始时期提前, 快速积累期终止时期和快速积累持续期延长; 随着施氮量增加, 花生干物质最大累积量、最大积累速率和出现时间、快速积累期起始和终止时期和快速积累持续期呈先升高后降低趋势; 施钙量和施氮量互作下, 花生干物质最大累积量在Ca600N150获得最大值, 其最大累积速率为327.6 kg hm-2 d-1, 快速累积持续时间为36.1 d, 与各处理平均值相比, 花生干物质最大累积量提高了13.7%, 最大累积速率提高了10.6%, 快速累积持续时间延长了3.1%。Table 1
表1
表1不同施钙量和施氮量花生干物质累积动态特征值
Table 1
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ym (kg hm-2) | Vm (kg hm-2 d-1) | tm (d) | t1 (d) | t2 (d) | T (d) | Ym (kg hm-2) | Vm (kg hm-2 d-1) | tm (d) | t1 (d) | t2 (d) | T (d) | |
| Ca0 | 13,750 | 272.6 | 62.5 | 45.9 | 79.1 | 33.2 | 14,824 | 282.4 | 60.5 | 43.2 | 77.8 | 34.6 |
| Ca600 | 15,604 | 312.1 | 62.6 | 46.1 | 79.1 | 32.9 | 16,728 | 307.8 | 60.5 | 42.6 | 78.4 | 35.8 |
| N0 | 12,676 | 267.8 | 61.9 | 46.3 | 77.5 | 31.2 | 13,250 | 261.7 | 58.3 | 41.6 | 75.0 | 33.3 |
| N75 | 14,313 | 291.6 | 62.3 | 46.1 | 78.4 | 32.3 | 15,518 | 290.6 | 60.5 | 43.0 | 78.1 | 35.2 |
| N150 | 14,954 | 296.1 | 62.4 | 45.7 | 79.0 | 33.3 | 17,106 | 314.3 | 61.0 | 43.1 | 78.9 | 35.8 |
| N225 | 16,063 | 308.8 | 63.0 | 45.8 | 80.1 | 34.3 | 16,646 | 305.8 | 61.2 | 43.3 | 79.1 | 35.8 |
| N300 | 15,388 | 298.9 | 62.8 | 45.8 | 79.7 | 33.9 | 16,367 | 304.8 | 60.9 | 43.2 | 78.6 | 35.4 |
| Ca0×N0 | 11,732 | 250.8 | 61.6 | 46.2 | 77.0 | 30.8 | 11,844 | 250.8 | 55.8 | 40.2 | 71.3 | 31.1 |
| Ca0×N75 | 13,257 | 271.4 | 62.0 | 45.9 | 78.1 | 32.2 | 14,326 | 277.2 | 59.8 | 42.7 | 76.8 | 34.0 |
| Ca0×N150 | 13,923 | 278.1 | 62.1 | 45.6 | 78.5 | 33.0 | 16,262 | 301.3 | 61.6 | 43.8 | 79.4 | 35.5 |
| Ca0×N225 | 15,266 | 288.9 | 63.4 | 46.0 | 80.8 | 34.8 | 16,045 | 296.0 | 62.3 | 44.4 | 80.1 | 35.7 |
| Ca0×N300 | 14,582 | 277.4 | 63.1 | 45.8 | 80.4 | 34.6 | 15,663 | 294.5 | 61.6 | 44.1 | 79.1 | 35.0 |
| Ca600×N0 | 13,620 | 285.3 | 62.2 | 46.5 | 77.9 | 31.4 | 14,665 | 276.1 | 60.1 | 42.6 | 77.6 | 35.0 |
| Ca600×N75 | 15,369 | 312.0 | 63.2 | 47.0 | 79.4 | 32.4 | 16,716 | 304.7 | 61.2 | 43.1 | 79.2 | 36.1 |
| Ca600×N150 | 15,988 | 314.5 | 62.6 | 45.9 | 79.3 | 33.5 | 17,949 | 327.6 | 60.5 | 42.4 | 78.5 | 36.1 |
| Ca600×N225 | 16,860 | 329.2 | 62.5 | 45.7 | 79.4 | 33.7 | 17,243 | 316.4 | 60.1 | 42.2 | 78.0 | 35.9 |
| Ca600×N300 | 16,194 | 320.6 | 62.5 | 45.9 | 79.2 | 33.3 | 17,073 | 316.3 | 60.1 | 42.4 | 77.9 | 35.5 |
| 平均Average | 14,679 | 292.8 | 62.5 | 46.0 | 79.0 | 33.0 | 15,779 | 296.1 | 60.3 | 42.8 | 77.8 | 35.0 |
新窗口打开|下载CSV
由图1可知, 随着生育进程, 花生干物质积累量呈S型曲线变化, 随着施钙量和施氮量增加, 不同生育期的干物质积累量提高。在济阳相比不施钙肥处理(JYCa0), 增施钙肥处理(JYCa600)成熟期干物质积累 量提高13.5%。随着施氮量增加, 成熟期各施氮处理干物质积累量分别比N0提高了12.8%、17.7%、26.3%和21.0%; 干物质积累量在Ca600条件下N225时取得最大值。饮马泉与济阳表现相同, 增施钙肥处理(YMQCa600)成熟期干物质积累量提高了12.6%; 干物质积累量在Ca600条件下N150时取得最大值。随着施氮量增加, 成熟期各施氮处理干物质积累量分别比N0提高了16.7%、28.4%、24.9%和22.9%。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同施钙量和施氮量的花生干物质累积动态
JY: 济阳; YMQ: 饮马泉。处理同
Fig. 1Dry matter accumulation under different calcium and nitrogen fertilizer rates in peanut
JY: Jiyang; YMQ: Yinmaquan. Treatments are the same as those given in
2.2 花生氮素积累动态
由表2可知, 在济阳增施CaO 600 kg hm-2, 花生氮素最大积累量和速率分别提高16.5%和26.8%, 最大积累速率出现提前, 快速积累期起始时期推后, 快速积累终止时期和持续期缩短; 随着施氮量增加, 花生氮素最大累积量、最大积累速率和出现时间、快速积累期终止时期和快速积累持续期呈先升高后降低趋势, 快速积累期起始时期缩短, 不同施氮处理花生氮素积累在N225时获得最大; 施钙量和施氮量互作下, 花生氮素最大累积量在Ca600N225获得最大值, 其最大累积速率为7.9 kg hm-2 d-1, 快速累积持续时间为29.6 d, 与各处理平均值相比, 花生氮素累积量提高了22.1%, 最大累积速率提高了21.6%, 快速累积持续时间延长了0.3%。在饮马泉增施CaO 600 kg hm-2, 花生氮素最大积累量和速率分别提高16.2%和16.6%, 快速积累期起始时期提前, 快速积累期终止时期和快速积累持续期缩短; 随着施氮量增加, 花生氮素积累动态特征值变化趋势与济阳相似, 而不同施氮处理花生氮素积累在N150时获得最大; 施钙量和施氮量互作下, 花生氮素最大累积量在Ca600N150获得最大值, 其最大累积速率为7.1 kg hm-2 d-1, 快速累积持续时间为31.5 d, 与各处理平均值相比, 花生干物质最大累积量提高了14.9%, 最大累积速率提高了11.6%, 快速累积持续时间延长了3.3%。与济阳相比, 饮马泉花生氮素最快累积起始时间、结束时间和最大速率出现时间都提早, 而氮素快速累积持续期略长。与干物质积累相比, 花生氮素快速积累期起始时间提前7.5~9.4 d, 最大积累速率出现时间提前9.3~12.0 d, 花生群体氮素营养吸收高峰期的出现早于干物质积累, 表明氮素积累是干物质积累的基础。Table 2
表2
表2不同施钙量和施氮量花生氮素累积动态特征值
Table 2
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ym (kg hm-2) | Vm (kg hm-2 d-1) | tm (d) | t1 (d) | t2 (d) | T (d) | Ym (kg hm-2) | Vm (kg hm-2 d-1) | tm (d) | t1 (d) | t2 (d) | T (d) | |
| 施钙量 Ca rate (kg hm-2) | ||||||||||||
| Ca0 | 268 | 5.7 | 53.8 | 38.3 | 69.3 | 31.0 | 264 | 5.7 | 49.3 | 34.1 | 64.5 | 30.4 |
| Ca600 | 313 | 7.2 | 52.9 | 38.7 | 67.2 | 28.5 | 306 | 6.6 | 48.2 | 33.0 | 63.3 | 30.3 |
| 施氮量 N rate (kg hm-2) | ||||||||||||
| N0 | 225 | 5.5 | 52.2 | 38.8 | 65.5 | 26.7 | 225 | 5.5 | 45.8 | 32.3 | 59.2 | 26.9 |
| N75 | 280 | 6.4 | 53.1 | 38.7 | 67.6 | 28.9 | 275 | 6.3 | 47.4 | 33.1 | 61.7 | 28.6 |
| N150 | 302 | 6.7 | 53.3 | 38.5 | 68.1 | 29.6 | 318 | 6.6 | 50.0 | 34.2 | 65.8 | 31.6 |
| N225 | 334 | 7.0 | 54.0 | 38.2 | 69.8 | 31.7 | 309 | 6.4 | 49.7 | 33.9 | 65.5 | 31.5 |
| N300 | 311 | 6.7 | 53.6 | 38.3 | 68.8 | 30.5 | 297 | 6.2 | 49.4 | 33.7 | 65.1 | 31.4 |
| 施钙量×施氮量 Ca rate × N rate | ||||||||||||
| Ca0×N0 | 204 | 5.1 | 52.4 | 39.1 | 65.7 | 26.6 | 205 | 5.0 | 46.3 | 32.7 | 59.9 | 27.1 |
| Ca0×N75 | 256 | 5.7 | 53.1 | 38.3 | 67.9 | 29.6 | 252 | 6.0 | 47.1 | 33.2 | 61.1 | 27.9 |
| Ca0×N150 | 276 | 5.9 | 53.5 | 38.1 | 69.0 | 30.8 | 295 | 6.2 | 50.0 | 34.4 | 65.6 | 31.2 |
| Ca0×N225 | 314 | 6.1 | 54.6 | 37.6 | 71.6 | 33.9 | 290 | 5.9 | 51.0 | 34.8 | 67.1 | 32.3 |
| Ca0×N300 | 292 | 5.9 | 54.4 | 38.1 | 70.7 | 32.6 | 276 | 5.9 | 50.1 | 34.6 | 65.6 | 31.0 |
| Ca600×N0 | 245 | 6.0 | 52.0 | 38.7 | 65.4 | 26.7 | 244 | 6.0 | 45.3 | 32.0 | 58.6 | 26.6 |
| Ca600×N75 | 305 | 7.1 | 53.1 | 39.0 | 67.3 | 28.2 | 298 | 6.7 | 47.5 | 32.9 | 62.0 | 29.1 |
| Ca600×N150 | 329 | 7.5 | 53.2 | 38.7 | 67.6 | 28.9 | 342 | 7.1 | 49.5 | 33.8 | 65.3 | 31.5 |
| Ca600×N225 | 355 | 7.9 | 53.5 | 38.7 | 68.3 | 29.6 | 327 | 7.0 | 48.6 | 33.3 | 64.0 | 30.8 |
| Ca600×N300 | 330 | 7.7 | 52.4 | 38.3 | 66.5 | 28.2 | 319 | 6.9 | 47.2 | 32.0 | 62.3 | 30.3 |
| 平均Average | 291 | 6.5 | 53.2 | 38.5 | 68.0 | 29.5 | 285 | 6.3 | 48.3 | 33.4 | 63.2 | 29.8 |
新窗口打开|下载CSV
由图2可知, 随着生育进程, 花生氮素积累量呈S型曲线变化, 随着施钙量和施氮量增加, 花生不同生育期氮素积累量升高。在济阳成熟期JYCa600相比JYCa0, 其氮素积累量提高16.6%; 随着施氮量增加, 成熟期各施氮处理氮素积累量分别比N0提高了24.5%、34.4%、48.5%和38.2%。饮马泉氮素积累特征与济阳表现相似, 与YMQCa0处理相比, YMQCa600成熟期氮素积累量提高了16.2%; 不同施氮量处理间氮素积累量均在Ca600条件下N150时取得最大值。随着施氮量增加, 成熟期各施氮处理氮素积累量分别比N0提高了22.3%、41.5%、37.1%和32.2%。综合来看, 施钙条件下中高氮处理的氮素积累量较高。施钙有利于植株健壮, 促进了氮素吸收, 而中高施氮量即可提供充足的供氮能力, 又减缓氮阻遏现象, 促进了更多的氮素吸收。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同施钙量和施氮量花生氮素累积动态
JY: 济阳; YMQ: 饮马泉。处理同
Fig. 2Nitrogen accumulation under different calcium and nitrogen fertilizer rates in peanut
JY: Jiyang; YMQ: Yinmaquan. Treatments are the same as those given in
2.3 花生干物质分配
由表3可知, 不同施钙量和施氮量间生殖器官与营养器官干物质量均差异极显著, 其中饮马泉花生干物质分配到营养器官的量在施氮和施钙互作条件下差异极显著。增施钙肥促进干物质向生殖器官分配, 提高7.1%~7.3%。济阳随着施氮量增加, 干物质分配到营养器官的量呈先升高后降低的趋势, N225和N300显著高于其他处理, 其分配系数分别为55.6%、57.2%; 分配到生殖器官的量呈先升高后降低趋势, N225显著高于其他处理, 其分配系数为44.4%, N150、N300间差异不显著; 施钙量和施氮量互作下, 生殖器官与营养器官干物质量均以Ca600N225处理最高, 其分配系数分别为53.8%、46.2%。在不同施钙量和施氮量及其互作条件下, 饮马泉花生生殖器官与营养器官干物质量均在Ca600N150处理最高, 其分配系数分别为49.6%、50.4%, 更多的干物质分配到生殖器官, 为高产形成奠定物质基础。与济阳相比, 饮马泉各处理营养器官平均干物质积累减少1.3%, 而生殖器官干物质积累量提高15.2%。Table 3
表3
表3不同施钙量和施氮量对花生干物质分配的影响
Table 3
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | ||||||
|---|---|---|---|---|---|---|---|---|
| 营养器官VO (kg hm-2) | 营养器官 VO (%) | 生殖器官 GO (kg hm-2) | 生殖器官 GO (%) | 营养器官 VO (kg hm-2) | 营养器官 VO (%) | 生殖器官 GO (kg hm-2) | 生殖器官 GO (%) | |
| 施钙量 Ca rate (kg hm-2) | ||||||||
| Ca0 | 8065 b | 59.2 | 5593 b | 40.8 | 8054 b | 55.0 | 6622 b | 45.0 |
| Ca600 | 8709 a | 56.3 | 6806 a | 43.7 | 8502 a | 51.6 | 7992 a | 48.4 |
| 施氮量 N rate (kg hm-2) | ||||||||
| N0 | 7603 c | 60.4 | 5013 d | 39.6 | 7282 c | 55.5 | 5869 d | 44.5 |
| N75 | 8329 b | 58.6 | 5897 c | 41.4 | 8110 b | 53.0 | 7240 c | 47.0 |
| N150 | 8443 b | 56.8 | 6445 b | 43.2 | 8597 a | 51.0 | 8287 a | 49.0 |
| N225 | 8843 a | 55.6 | 7090 a | 44.4 | 8737 a | 53.2 | 7712 b | 46.8 |
| N300 | 8718 a | 57.2 | 6551 b | 42.8 | 8665 a | 53.9 | 7429 c | 46.1 |
| 施钙量×施氮量 Ca rate × N rate | ||||||||
| Ca0×N0 | 7170 e | 61.4 | 4503 g | 38.6 | 6717 e | 57.0 | 5067 h | 43.0 |
| Ca0×N75 | 7876 d | 59.8 | 5302 f | 40.2 | 7766 d | 54.7 | 6437 g | 45.3 |
| Ca0×N150 | 8077 d | 58.4 | 5763 d | 41.6 | 8418 c | 52.4 | 7647 d | 47.6 |
| Ca0×N225 | 8683 bc | 57.4 | 6450 c | 42.6 | 8743 a | 55.2 | 7101 f | 44.8 |
| Ca0×N300 | 8515 c | 58.9 | 5948 d | 41.1 | 8627 bc | 55.7 | 6858 ef | 44.3 |
| Ca600×N0 | 8036 d | 59.3 | 5523 e | 40.7 | 7848 d | 54.1 | 6668 fg | 45.9 |
| Ca600×N75 | 8780 ab | 57.5 | 6493 c | 42.5 | 8455 bc | 51.2 | 8043 c | 48.8 |
| Ca600×N150 | 8809 ab | 55.3 | 7127 b | 44.7 | 8775 a | 49.6 | 8927 a | 50.4 |
| Ca600×N225 | 9002 a | 53.8 | 7730 a | 46.2 | 8732 a | 51.2 | 8323 b | 48.8 |
| Ca600×N300 | 8920 ab | 55.5 | 7155 b | 44.5 | 8702 ab | 52.1 | 7999 c | 47.9 |
| 变异来源 Source of variation | ||||||||
| 施钙量 Calcium (Ca) | ** | ** | ** | ** | ||||
| 施氮量 Nitrogen (N) | ** | ** | ** | ** | ||||
| 施钙量×施氮量 Ca×N | ns | ns | ** | ns | ||||
新窗口打开|下载CSV
2.4 花生氮素分配
由表4可知, 不同施钙量和施氮量间生殖器官与营养器官的氮素积累量均差异极显著。增施钙肥促进氮素向生殖器官分配, 提高9.9%~12.0%。随着施氮量增加, 济阳花生氮素分配到生殖器官和营养器官的量呈先升高后降低的趋势, N225显著高于其他处理, 其分配系数分别为67.4%和32.6%; 施钙和施氮互作下, 生殖器官与营养器官氮素积累量均以Ca600N225处理最高, 其分配比例分别为69.3%和30.7%。Table 4
表4
表4不同施钙量和施氮量对花生氮素分配的影响
Table 4
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 营养器官 VO (kg hm-2) | 营养器官 VO (%) | 生殖器官 GO (kg hm-2) | 生殖器官 GO (%) | 营养器官 VO (kg hm-2) | 营养器官 VO (%) | 生殖器官 GO (kg hm-2) | 生殖器官 GO (%) | ||||||||||||
| 施钙量 Ca rate (kg hm-2) | |||||||||||||||||||
| Ca0 | 96.8 | 36.3 | 170.9 b | 63.7 | 98.8 b | 37.7 | 164.2 b | 62.3 | |||||||||||
| Ca600 | 100.7 a | 32.4 | 211.6 a | 67.6 | 104.3 a | 34.3 | 201.2 a | 65.7 | |||||||||||
| 施氮量 N rate (kg hm-2) | |||||||||||||||||||
| N0 | 81.5 e | 36.5 | 143.1 d | 63.5 | 86.5 c | 38.7 | 137.9 e | 61.3 | |||||||||||
| N75 | 96.9 d | 34.8 | 182.9 c | 65.2 | 98.8 b | 36.1 | 175.7 d | 63.9 | |||||||||||
| N150 | 100.9 c | 33.6 | 201.0 b | 66.4 | 107.0 a | 33.8 | 210.6 a | 66.2 | |||||||||||
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | |||||||||||||||||
| 营养器官 VO (kg hm-2) | 营养器官 VO (%) | 生殖器官 GO (kg hm-2) | 生殖器官 GO (%) | 营养器官 VO (kg hm-2) | 营养器官 VO (%) | 生殖器官 GO (kg hm-2) | 生殖器官 GO (%) | ||||||||||||
| N225 | 108.4 a | 32.6 | 225.2 a | 67.4 | 108.0 a | 35.2 | 199.8 b | 64.8 | |||||||||||
| N300 | 106.0 b | 34.3 | 204.3 b | 65.7 | 107.2 a | 36.3 | 189.4 c | 63.7 | |||||||||||
| 施钙量×施氮量 Ca rate × N rate | |||||||||||||||||||
| Ca0×N0 | 78.0 g | 38.2 | 126.2 g | 61.8 | 82.2 d | 40.1 | 122.6 h | 59.9 | |||||||||||
| Ca0×N75 | 94.1 e | 36.9 | 161.0 f | 63.1 | 95.1 c | 37.7 | 156.8 g | 62.3 | |||||||||||
| Ca0×N150 | 98.7 d | 35.9 | 176.6 e | 64.1 | 104.9 ab | 35.7 | 189.0 de | 64.3 | |||||||||||
| Ca0×N225 | 108.0 a | 34.5 | 204.8 c | 65.5 | 106.5 ab | 36.9 | 182.4 e | 63.1 | |||||||||||
| Ca0×N300 | 105.2 ab | 36.1 | 186.1 d | 63.9 | 105.2 ab | 38.2 | 170.0 f | 61.8 | |||||||||||
| Ca600×N0 | 85.0 f | 34.7 | 160.0 f | 65.3 | 90.9 c | 37.3 | 153.2 g | 62.7 | |||||||||||
| Ca600×N75 | 99.6 cd | 32.7 | 204.7 c | 67.3 | 102.6 b | 34.5 | 194.6 d | 65.5 | |||||||||||
| Ca600×N150 | 103.1 bc | 31.4 | 225.3 b | 68.6 | 109.2 a | 32.0 | 232.3 a | 68.0 | |||||||||||
| Ca600×N225 | 108.7 a | 30.7 | 245.5 a | 69.3 | 109.5 a | 33.5 | 217.2 b | 66.5 | |||||||||||
| Ca600×N300 | 106.9 ab | 32.4 | 222.6 b | 67.6 | 109.1 a | 34.3 | 208.9 c | 65.7 | |||||||||||
| 变异来源Source of variation | |||||||||||||||||||
| 施钙量 Calcium (Ca) | ** | ** | ** | ** | |||||||||||||||
| 施氮量 Nitrogen (N) | ** | ** | ** | ** | |||||||||||||||
| 施钙量×施氮量 Ca×N | ns | ns | ns | ns | |||||||||||||||
新窗口打开|下载CSV
随着施氮量增加, 饮马泉花生氮素分配到生殖器官和营养器官的量规律与济阳一致, 其中N150生殖器官氮素积累量显著高于其他处理, 其分配系数为66.2%。在不同施钙量和施氮量及其互作条件下, 生殖器官与营养器官氮素积累量均在Ca600N150处理最高, 其分配系数分别为68.0%和32.0%, 更多的氮素分配到生殖器官。
2.5 花生产量及产量构成
由表5可知, 不同试验样地花生产量存在差异。同一样地中的施钙量、施氮量及两者互作对花生荚果产量均有极显著影响, 其中百果重和结果数是差异的主要因素。增施钙肥, 荚果产量提高13.4%~15.7%, 结果数增加6.0%~7.0%。Table 5
表5
表5不同施钙量和施氮量对花生产量和产量构成的影响
Table 5
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 荚果产量 Pod yield (kg hm-2) | 总果数 Total pods (×104 pods hm-2) | 百果重 100-pod weight (g) | 百仁重 100-kernel weight (g) | 出仁率 Kernel rate (%) | 荚果产量 Pod yield (kg hm-2) | 总果数 Total pods (×104 pods hm-2) | 百果重 100-pod weight (g) | 百仁重 100-kernel weight (g) | 出仁率 Kernel rate (%) | ||||||||||||
| 钙量 Ca rate (kg hm-2) | |||||||||||||||||||||
| Ca0 | 5034 b | 330 b | 229.0 b | 94.6 b | 70.6 a | 5551 b | 345 b | 236.3 b | 97.3 a | 70.3 a | |||||||||||
| Ca600 | 5824 a | 351 a | 236.1 a | 95.9 a | 70.7 a | 62,942 a | 366 a | 242.7 a | 98.8 a | 70.5 a | |||||||||||
| 施氮量 N rate (kg hm-2) | |||||||||||||||||||||
| N0 | 4401 d | 312 d | 218.1 d | 92.7 d | 70.0 a | 5073 d | 330 d | 225.1 c | 95.5 c | 69.8 b | |||||||||||
| N75 | 5053 c | 329 c | 230.5 c | 95.2 c | 70.5 a | 5648 c | 344 c | 238.9 b | 98.0 b | 70.3 ab | |||||||||||
| N150 | 5712 b | 347 b | 237.0 bc | 96.1 c | 70.8 a | 6486 a | 375 a | 249.7 a | 100.3 a | 70.8 a | |||||||||||
| N225 | 6172 a | 356 a | 242.4 b | 97.4 b | 71.0 a | 6348 a | 367 b | 244.3 ab | 99.0 ab | 70.6 a | |||||||||||
| N300 | 5805 b | 352 a | 234.8 a | 94.8 a | 70.8 a | 6057 b | 363 b | 239.6 b | 97.5 b | 70.5 a | |||||||||||
| 施钙量×施氮量 Ca rate × N rate | |||||||||||||||||||||
| Ca0×N0 | 3903 g | 299 f | 210.8 f | 92.3 d | 69.8 c | 4686 f | 320 f | 219.2 e | 95.0 c | 69.7 c | |||||||||||
| Ca0×N75 | 4500 f | 313 e | 228.3 de | 94.4 c | 70.6 ab | 5219 e | 331 e | 235.4 c | 97.2 b | 70.0 bc | |||||||||||
| Ca0×N150 | 5379 d | 336 c | 233.6 cd | 95.2 b | 70.7 ab | 6102 c | 361 c | 248.7 a | 99.7 b | 70.7 ab | |||||||||||
| Ca0×N225 | 5876 c | 347 bc | 239.4 bc | 96.8 a | 71.0 a | 6044 c | 359 c | 241.5 b | 98.1 b | 70.6 ab | |||||||||||
| 处理 Treatment | 济阳Jiyang (JY) | 饮马泉Yinmaquan (YMQ) | |||||||||||||||||||
| 荚果产量 Pod yield (kg hm-2) | 总果数 Total pods (×104 pods hm-2) | 百果重 100-pod weight (g) | 百仁重 100-kernel weight (g) | 出仁率 Kernel rate (%) | 荚果产量 Pod yield (kg hm-2) | 总果数 Total pods (×104 pods hm-2) | 百果重 100-pod weight (g) | 百仁重 100-kernel weight (g) | 出仁率 Kernel rate (%) | ||||||||||||
| Ca0×N300 | 5510 cd | 343 c | 232.9 cd | 94.3 c | 70.7 ab | 5703 d | 355 c | 236.8 c | 96.7 bc | 70.3 abc | |||||||||||
| Ca600×N0 | 4900 e | 324 d | 225.3 e | 93.1 d | 70.2 bc | 5459 e | 340 d | 231.0 d | 95.9 c | 69.8 c | |||||||||||
| Ca600×N75 | 5606 c | 345 c | 232.7 cd | 96.0 ab | 70.5 ab | 6078 c | 357 c | 242.4 b | 98.9 ab | 70.7 ab | |||||||||||
| Ca600×N150 | 6046 b | 358 ab | 240.3 ab | 97.0 a | 70.9 ab | 6871 a | 388 a | 250.7 a | 100.9 a | 70.9 ab | |||||||||||
| Ca600×N225 | 6467 a | 366 a | 245.4 a | 98.0 a | 71.1 a | 6651 a | 374 b | 247.1 a | 99.9 ab | 70.7 a | |||||||||||
| Ca600×N300 | 6099 b | 360 a | 236.7 bc | 95.4 b | 70.9 ab | 6412 b | 370 b | 242.4 b | 98.3 b | 70.6 ab | |||||||||||
| 变异来源Source of variation | |||||||||||||||||||||
| 施钙量 Calcium (Ca) | ** | ** | ** | * | ns | ** | ** | ** | * | ns | |||||||||||
| 施氮量 Nitrogen (N) | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | |||||||||||
新窗口打开|下载CSV
随着施氮量增加, 济阳花生荚果产量呈现先升高后降低的趋势, 以N225花生荚果产量最高, N300与N150次之, 且差异不显著, 表明高施氮量对花生产量的提高效应与中施氮量相当。与N0相比, 各施氮处理间花生产量分别提高了14.8%、29.8%、40.2%和31.9%。施钙量和施氮量互作下, 花生荚果产量、结果数和百果重以Ca600N225最高, 而Ca0N150和Ca0N300、Ca600N150和Ca600N300处理间差异不显著。Ca600N75荚果产量高于Ca0N0、Ca0N75、Ca0N150、Ca0N300产量, 而相比Ca0N225产量降低4.8%, 但2个处理差异不显著(P>0.05), 可见, 在不追求花生荚果产量极高的条件下施氮量为75 kg hm-2, 且配施600 kg hm-2氧化钙, 可获得稳产。Ca600N150花生荚果产量高于不施钙(Ca0)处理N0~N300产量, 表明获得花生荚果高产可以通过增施钙肥来实现, 并且钙肥施用可作为氮肥减施后花生稳产高产的重要栽培措施。在饮马泉不同施钙量、施氮量处理间花生荚果产量的变化趋势与济阳相同。两者互作条件下, 荚果产量Ca600N150、Ca600N2250条件下均高产, 两处理间差异未达显著水平, 且均显著高于其他处理。与N0相比, 各施氮处理间花生产量分别提高了11.3%、27.9%、25.1%和19.4%。百果重、总果数在Ca600条件下显著高于Ca0, 同一钙肥水平下随施氮量增加呈先升高后降低的变化趋势。施氮量对百仁重和出仁率均有极显著影响, 而施钙量对百仁重有显著影响, 但是对出仁率无显著影响。与济阳相比, 饮马泉平均花生荚果产量高8.3%。
由图3可知, 不施钙肥条件下, 氮肥与对荚果产量的关系可用二次方程模拟y = -0.0311x2+ 14.302x+4192.4, 获得高产时最佳施氮量为229.9 kg hm-2; 施钙条件下, 氮肥与荚果产量的关系可用二次方程模拟y= -0.0344x2+13.875x+5001.9, 获得高产时最佳施氮量为201.8 kg hm-2。钙肥的施用促进了植株对氮素的吸收, 同时减少了相同荚果产量下氮肥的施用量。
图3
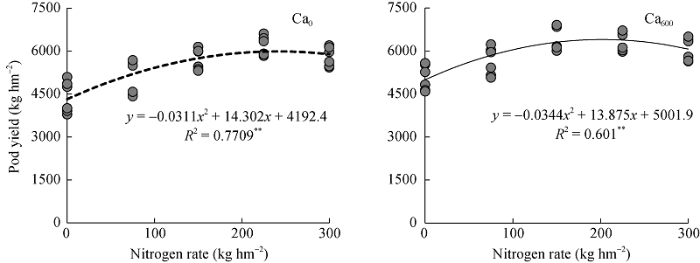 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3钙肥与氮肥互作对花生荚果产量的影响
**表示在0.01水平上显著相关。30个重复。
Fig. 3Effects of the interaction of calcium and nitrogen fertilizer on pod yield in peanut
** indicates significant correlation at the 0.01 probability level. Values are means with thirty repetitions.
3 讨论
3.1 施钙量和施氮量对花生干物质积累与分配的影响
肥料施用作为花生栽培中主要的调控措施, 对花生干物质积累与分配及产量有重大影响[6, 25-26]。钙是花生干物质积累及荚果发育等重要的调控措施[27,28]。氮是植物重要的营养元素, 在促进营养体的生长的同时决定着植物生殖体的发育。研究表明, 施氮显著改善花生叶片光合性能, 提高茎、叶及荚果干物质积累量[15,16,17,18]; 施氮量在0~135 kg hm-2时, 不同花生品种干物质量均随着施氮量的增加呈增加的趋势[29]。张智猛等[30]研究表明, 施氮量在0~180 kg hm-2时, 施氮量90 kg hm-2时植株总干物质较高、荚果干物质最高。过量施用氮肥(N>112.5 kg hm-2), 造成花生群体内透光条件变差, 群体叶面积系数和光合速率降低, 群体呼吸消耗所占比例增加, 不利于植株干物质的积累[17]。随施氮水平增加, 茎叶等干物质分配增加, 而荚果干物质分配减少, 收获指数降低[3]。本试验在前人研究基础上采取增施钙肥措施发现, 随着施钙量和施氮量增加, 济阳和饮马泉样地分别表现为Ca600N225、Ca600N150时花生干物质积累量最大、生殖器官的分配比例最高。与前人研究[29,30]相比, 本研究结果中的最佳施氮量(150~225 kg hm-2)偏高, 这可能与试验方法和试验条件有关。本试验中采用大田试验, 而与之前的相关研究均采用盆栽试验有所不同, 同时花生长季气象条件和土壤基础地力的影响也会引起不同区域试验结果上的差异。不施钙肥处理(Ca0N0~N225)、施钙高氮处理(Ca600N300)表现营养生长过旺、叶片熟相较差、贪青晚熟, 不利于后期花生荚果的进一步充实。钙肥增施可促进总干物质的积累、特别是生殖器官中干物质的积累; 氮肥减施可抑制“源”的冗余生长、促进“流”的通畅、进一步优化“库”容、提高生殖器官分配比例, 这些措施均为花生产量的提高提供了重要的物质基础。3.2 施钙量和施氮量对花生氮素积累与分配的影响
施钙是调节花生干物质和氮素积累分配的重要手段[11,31-32]。前人研究表明, 施硝酸钙促进花生对氮素营养的吸收及向“库”中的运输与转化[9]。随施钙量增加, 花生生殖器官氮素积累量和分配率提高, 氮素吸收效率提高[27]。戴良香等[12]研究表明, 施氮量为90 kg hm-2时, 花生籽仁干物重和氮素积累量最高。施氮量过高使花生营养体旺长倒伏, 产量和氮肥利用率降低[33]。本研究中花生干物质积累和氮素积累的变化规律一致, 增施钙肥显著提高花生植株氮素积累量, 随着施氮量的增加, 氮素积累量表现为先升高后降低的趋势。从文中的2个区域试验结果来看, 花生氮素快速积累期起始时间和最大积累速率较干物质积累分别提前7.5~9.4 d和9.3~ 12.0 d, 花生群体氮素营养吸收高峰期的出现早于干物质积累, 表明氮素营养的吸收和积累是干物质积累的基础。施钙和施氮互作下, 生殖器官与营养器官氮素积累量以Ca600N150和Ca600N225处理最高, 其生殖器官分配比例为68.0%~69.3%, 表明更多的氮素分配到生殖器官, 为提高群体质量建成打下物质基础。而施氮量过大导致更多的氮素分配到花生营养器官中, 地上部营养生长过盛, 可见根据不同的土壤肥力采取氮肥减施策略有利于氮素的高效利用。3.3 施钙量和施氮量对花生产量的影响
施钙和施氮是调节花生干物质和氮素积累分配的重要栽培措施[34,35,36,37]。有研究表明盐碱地胁迫下施用钙肥150 kg hm-2, 花生荚果产量提高达21.5% [38]; Rogers[39]研究发现, 沙壤土增施钙肥(白云质石灰石和钙质硅酸盐矿渣) 675 kg hm-2, 花生产量提高405~675 kg hm-2。山东酸性土壤施用钙肥为210 kg hm-2时, 花生产量增幅最大[34]; 湖南酸性红壤(pH为4.6)施钙量在750 kg hm-2时, 花生荚果和籽仁产量提高显著[27]。本试验中与济阳相比, 饮马泉样地平均花生荚果产量高8.3%, 可能原因是与济阳相比, 饮马泉样地土壤偏弱酸性、土质松软、水热条件适宜, 为花生干物质积累和荚果的充实奠定了良好的基础条件, 同时氮肥增产效率因土壤质地和氮素水平存在较大差异[19]。而不同花生产区对氮肥的投入量存在差异, 同时获得的产量水平也不同。辽宁省花生产区施氮量在75~105 kg hm-2时, 荚果产量为3114~7506 kg hm-2 [40]。湖北省农户花生平均氮肥施用量为114.7 kg hm-2, 平均产量为2968 kg hm-2 [41]。山东省花生平均施氮量在181.0 kg hm-2左右, 产量高达4256 kg hm-2 [2]。本研究发现中低产田花生增施钙肥600 kg hm-2、施氮量150~225 kg hm-2可得到高产, 高于山东省平均产量, 可解释为增施钙肥对碱性地块、酸性地块的花生产量都有增产作用[27,34,38]。而不追求花生荚果产量极高的条件下, 采取增施钙肥+减施氮肥栽培措施(氮肥施用量75 kg hm-2), 可获得稳产, 其增产增效最佳。产量构成因素中总果数、百果重是获得高产的主要因素, 减氮后增施钙肥促进结果数增加和荚果大而饱满, 是维持花生高产的重要保证。4 结论
增施钙肥和施氮可以显著提高花生生育前期和中期的氮素和干物质积累速率、增加氮素和干物质积累总量。两者互作可充分发挥弱碱性地块和弱酸性地块单粒精播花生群体增产潜力。通过增施钙肥提高氮肥利用效率, 来获得花生结果数和百果重的增加是稳产高产的重要途径。本试验区域花生生产中建议施钙量为600 kg hm-2, 施氮量75~225 kg hm-2。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 4]
[本文引用: 4]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
PMID [本文引用: 2]

Biological N(2) fixation represents the major source of N input in agricultural soils including those in arid regions. The major N(2)-fixing systems are the symbiotic systems, which can play a significant role in improving the fertility and productivity of low-N soils. The Rhizobium-legume symbioses have received most attention and have been examined extensively. The behavior of some N(2)-fixing systems under severe environmental conditions such as salt stress, drought stress, acidity, alkalinity, nutrient deficiency, fertilizers, heavy metals, and pesticides is reviewed. These major stress factors suppress the growth and symbiotic characteristics of most rhizobia; however, several strains, distributed among various species of rhizobia, are tolerant to stress effects. Some strains of rhizobia form effective (N(2)-fixing) symbioses with their host legumes under salt, heat, and acid stresses, and can sometimes do so under the effect of heavy metals. Reclamation and improvement of the fertility of arid lands by application of organic (manure and sewage sludge) and inorganic (synthetic) fertilizers are expensive and can be a source of pollution. The Rhizobium-legume (herb or tree) symbiosis is suggested to be the ideal solution to the improvement of soil fertility and the rehabilitation of arid lands and is an important direction for future research.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 4]
[本文引用: 4]
DOIPMID [本文引用: 1]
Calcium not only serves as a necessary nutrient for plant growth but also acts as a ubiquitous central hub in a large number of signaling pathways. Free Ca2+ deficiency in the soil may cause early embryo abortion, which eventually led to abnormal development of peanut pod during the harvest season. To understand the mechanisms of Ca2+ regulation in pod development, transcriptome analysis of peanut gynophores and pods was performed by comparing the treatments between free Ca2+ sufficiency and free Ca2+ deficiency using Illumina HiSeq (TM) 2000. 9,903,082,800 nt bases are generated totally. After assembly, the average length of 102,819 unigenes is 999 nt, N50 is 1,782 nt. RNA-seq based gene expression profilings showed a large number of genes at the transcriptional level changed significantly between the aerial pegs and underground swelling pods under free Ca2+ sufficienct or deficiency treatments, respectively. Genes encoding key members of Ca2+ signaling transduction pathway, enzymes for hormone metabolism, cell division and growth, transcriptional factor as well as embryo development were highlighted. This information provides useful information for our further study. The results of digital gene expression (DGE) indicated that exogenous calcium might contribute to the development of peanut pod through its signal transduction pathway, meanwhile, promote the normal transition of the gynophores to the reproductive development.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}