Genetic analysis of fat content based on nested populations in peanut (Arachis hypogaea L.)
HUANG Bing-Yan,1,**, SUN Zi-Qi1,**, LIU Hua1, FANG Yuan-Jin1, SHI Lei1, MIAO Li-Juan1, ZHANG Mao-Ning1, ZHANG Zhong-Xin1, XU Jing1, ZHANG Meng-Yuan2, DONG Wen-Zhao1, ZHANG Xin-You,1,*1Henan Academy of Crops Molecular Breeding, Henan Academy of Agricultural Sciences/Graduate R & T Base of Zhengzhou University/Key Laboratory of Oil Crops in Huang-Huai-Hai Plans, Ministry of Agriculture and Rural Affairs/Henan Provincial Key Laboratory for Oil Crops Improvement, Zhengzhou 450002, Henan, China 2College of Agriculture, Henan University of Science and Technology, Luoyang 471023, Henan, China
First author contact:**Contributed equally to this work Received:2020-06-24Accepted:2020-12-1Online:2021-06-12
Fund supported:
The China Agricultural Research System.CARS-13 The Henan Agricultural Research System.2016-05 The Key Scientific and Technological Project of Henan Province.201300111000
Abstract Nested populations can be used to dissect the heredity of complex traits. The genetic models of fat content of F2:3 families in nested combinations with one common parent and six founder parents were analyzed, aiming to detect the genetic differences among the founders and to provide bases for breeding strategy for fat content improvement in peanut kernels. The common parent was Yuhua 15, an irregular-type variety with high fat content, and the other six founder parents were different botanical varieties with different fat contents. The results showed that the genetic model of fat content traits was different in different combinations. Six crosses were in accordance with three genetic patterns, including none major gene model, one major gene model with additive and dominant effect, and two major genes model with equal additive and dominant effect. The estimated values of various genetic effects were also different. The heritability of the main genes ranged from 32% to 80%, indicating that the gene loci controlling the fat content and their segregation patterns were different in different F2:3 populations. There were more individuals with high fat content in the offspring from combinations with both parents of high fat content. However, the heritability was low and phenotypic selections for fat content were not suggested in the early generations in such combinations. The offspring from combinations with parents of significantly different fat content had a larger variation range in fat content, and phenotypes with variable fat content were available. In this study, the large variances in the nested populations demonstrated the genetic complexity of fat content and the characteristics of multi major gene regulation. These results provide a comprehensive base for understanding the genetics and regulation of fat content, and the nested populations will be helpful in further QTL detection of fat content in peanut. Keywords:peanut (Arachis hypogaea L.);nested populations;fat content;F2:3 family;inheritance model
PDF (350KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄冰艳, 孙子淇, 刘华, 房元瑾, 石磊, 苗利娟, 张毛宁, 张忠信, 徐静, 张梦圆, 董文召, 张新友. 花生巢式群体的脂肪含量遗传分析[J]. 作物学报, 2021, 47(6): 1100-1108. doi:10.3724/SP.J.1006.2021.04138 HUANG Bing-Yan, SUN Zi-Qi, LIU Hua, FANG Yuan-Jin, SHI Lei, MIAO Li-Juan, ZHANG Mao-Ning, ZHANG Zhong-Xin, XU Jing, ZHANG Meng-Yuan, DONG Wen-Zhao, ZHANG Xin-You. Genetic analysis of fat content based on nested populations in peanut (Arachis hypogaea L.)[J]. Acta Agronomica Sinica, 2021, 47(6): 1100-1108. doi:10.3724/SP.J.1006.2021.04138
Table 4 表4 表4脂肪含量遗传参数估计 Table 4Estimation of genetic parameters of fat content
组合 Crosses
模型 Models
m
da
db
ha
hb
Var.mg
hmg
AC
1对主基因加性显性 1MG-AD
57.5201
1.1342
-2.3266
1.0357
31.9936
无主基因 0MG
1对主基因负向完全显性 1MG-NCD
57.2576
1.2970
0.9789
30.2396
2对主基因等加性 2MG-EA
56.9106
0.2372
0.0660
2.0382
BC
2对主基因等显性 2MG-EAD
55.5452
0.9807
1.1551
47.1385
1对主基因加性显性 1MG-AD
55.4744
1.0402
2.1944
0.8997
36.7188
无主基因 0MG
1对主基因完全显性 1MG-EAD
55.7181
1.2259
0.8773
35.8046
CE
2对主基因等显性 2MG-EAD
54.0134
1.0075
1.2100
52.9489
1对主基因加性显性 1MG-AD
53.9686
1.0950
2.1686
0.9448
41.3432
2对主基因完全显性 2MG-CD
54.1576
1.2797
0.1270
0.9596
41.9920
1对主基因完全显性 1MG-EAD
54.1894
1.2796
0.9498
41.5652
CF
无主基因 0MG
2对主基因等显性 2MG-EAD
53.6370
0.7674
0.7028
31.3534
2对主基因等加性 2MG-EA
54.0267
0.2282
0.0604
2.6958
1对主基因加性 1MG-A
54.0290
0.2888
0.0494
2.2052
组合 Crosses
模型 Models
m
da
db
ha
hb
Var.mg
hmg
CG
2对主基因等显性 2MG-EAD
51.6018
1.6437
3.6377
79.7548
1对主基因加性显性 1MG-AD
51.8983
2.3225
1.5772
2.9822
65.3833
2对主基因完全显性 2MG-CD
51.7690
2.1335
0.1047
2.7537
60.3730
1对主基因完全显性 1MG-EAD
51.7952
2.1334
2.7468
60.2228
CH
无主基因 0MG
2对主基因等显性 2MG-EAD
53.8575
0.7166
0.6290
16.0300
2对主基因等加性 2MG-EA
54.2152
0.3550
0.1469
3.7446
1对主基因加性 1MG-A
54.2168
0.4650
0.1280
3.2625
A: 远杂9102; B: 中花6号; C: 豫花15号; E: 粤油20; F: 四粒红; G: 伏花生; H: NC94022。m: 群体平均数; da: 第1对主基因加性效应; db: 第2对主基因加性效应; ha: 第1对主基因显性效应; hb: 第2对主基因显性效应; Var.mg: 主基因方差; hmg: 主基因遗传力。 A: Yuanza 9102; B: Zhonghua 6; C: Yuhua 15; E: Yueyou 20; F: Silihong; G: Fuhuasheng; H: NC94022. m:mean of population; da: additive effect of the first major gene; db: additive effect of the second major gene; ha: dominant effect of the first major gene; hb: dominant effect of the second major gene; Var.mg: major gene variance; hmg: heritability of major gene. MG: major gene; AD: additive and dominant; NCD: negative complete dominant; EA: equal additive; EAD: equal dominant.
Liao BS, LeiY, Wang SY, LiD, Huang JQ, Jiang HF, Ren XP. Genetic diversity of peanut RILs and enhancement for high oil genotypes Acta Agron Sin, 2008,34:999-1004 (in Chinese with English abstract). [本文引用: 1]
Yu SL, Yang QL, Pan LJ, Bo WN. Genetic analysis for oil content of peanut seeds J Plant Genet Resour, 2009,10:453-456 (in Chinese with English abstract). [本文引用: 2]
Chen SL, Li YR, Cheng ZS, Liao BS, LeiY, Liu JS. Heterosis and genetic analysis of oil content in peanut using mixed model of major gene and polygene Sci Agric Sin, 2009,42:3048-3057 (in Chinese with English abstract). [本文引用: 1]
ZhangX, ZhuS, HanS, XuJ, LiuH, TangF, DongW, ZangX, Zhang ZX. Inheritance of fat and fatty acid compositions in peanut (Arachis hypogaea L.) Agric Sci Technol, 2011,12:943-946. [本文引用: 2]
LiuH, Zhang XY, Cui DX, Tang FS, Dong WZ, Han SY, XuJ, Zang XW, Zhang ZX. Genetic analysis of protein and fat content using major gene plus polygene methods in peanut (Arachis hypogaea L.) Jiangsu Agric Sci, 2011,39(2):127-130 (in Chinese with English abstract). [本文引用: 1]
LiuH, QinL, Zhang XY, Han SY, DuP, Zhang ZX, Sun ZQ, Qi FY, Dong WZ, Huang BY. Genetic models of peanut fat and fatty acid compositions based on interspecific RIL population Chin J Oil Crop Sci, 2016,38:172-178 (in Chinese with English abstract). [本文引用: 2]
Wang JK, Gai JY. Identification of major gene and polygene mixed inheritance model and estimation of genetic parameters of a quantitative trait from F2 progeny Act Genet Sin, 1997,24:432-440 (in Chinese with English abstract). [本文引用: 1]
MengL, LiH, ZhangL, WangJ. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in bi-parental populations Crop J, 2015,3:269-283. [本文引用: 1]
Gan SM, Cao YL, Wei JX, Cui WF, Gu SY, Liu FS. Combining ability analysis of quatitative traits in peanut Chin J Oil Crop Sci, 1981, ( 3):33-45 (in Chinese with English abstract). [本文引用: 1]
Isleib TG, Pattee HE, Giesbrecht FG. Oil, sugar, and starch characteristics in peanut breeding lines selected for low and high oil content and their combining ability J Agric Food Chem, , 200452:3165-3168. [本文引用: 1]
Xu YM, Gan XM, Cao YL, Gu SY, Liu FS. studies on combining ability of major nutritional quality characters and agronomic characters in peanut Sci Agric Sin, 1995,28(2):15-23 (in Chinese with English abstract) . [本文引用: 1]
Liang XQ, Zheng GR, Xiang RY, Li XY. Combining ability analysis of yield and oil content of Spanish type peanut Sci Technol Peanut, 1991, ( 3):11-14 (in Chinese with English abstract). [本文引用: 1]
Liang XQ, Zheng GR, Xiang RY, Li XY. A diallel analysis of yield and oil content of spanish type peanut Chin Oil Crop, 1992, ( 1):19-23 (in Chinese with English abstract). [本文引用: 1]
Fu SX, Qi CK. Major gene plus polygene inheritance of oil content in Brassica napus L Jiangsu J Agric Sci, 2009,25:731-736 (in Chinese with English abstract). [本文引用: 1]
Zhang SF, Ma CZ, Zhu JC, Wang JP, Wen YC, Fu TD. Genetic analysis of oil content in Brassica napus L. using mixed model of major gene and polygene Acta Genet Sin, 2006,2:171-180. [本文引用: 1]
Jin MY, Li JN, Fu FY, Zhang ZS, Zhang XK, Liu LZ. QTL analysis of oil and hull content in Brassica napus L Sci Agric Sin, 2007,40:677-684 (in Chinese with English abstract). [本文引用: 1]
Zhang JF, Qi CK, Pu HM, ChenS, ChenF, Gao JQ, Chen XJ, GuH, Fu SZ. Inheritance and QTL identification of oil content in rapeseed (Brassica napus L.) Acta Agron Sin, 2007,33:1495-1501 (in Chinese with English abstract). [本文引用: 1]
Zhang XY, Han SY, XuJ, YanM, LiuH, Tang FS, Dong WZ, Huang BY. Identification of QTLs for important quality traits in cultivated peanut (Arachis hypogaea L.) Chin J Oil Crop Sci, 2012,34:311-315 (in Chinese with English abstract). [本文引用: 1]
HuangL, Zhao XY, Zhang WH, Fan ZM, Ren XP, Liao BS, Jiang HF, Chen YN. Identification of SSR markers linked to oil content in peanut (Arachis hypogaea L.) through RIL population and natural population Acta Agron Sin, 2011,37:1967-1974 (in Chinese with English abstract). [本文引用: 1]
Pandey MK, Wang ML, Qiao LX, Feng SP, KheraP, WangH, TonnisB, Barkley NA, Wang JP, Holbrook CC, Culbreath AK, Varshney RK, Guo BZ. Identification of QTLs associated with oil content and mapping FAD2 genes and their relative contribution to oil quality in peanut (Arachis hypogaea L.) BMC Genet, 2014,15:133. [本文引用: 1]
Wei DY, Cui YX, Mei JQ, Tang QL, Li JN, QianW. Genome-wide association study on seed oil content in rapeseed and construction of integration system for oil content loci Acta Agron Sin, 2018,44:1311-1319 (in Chinese with English abstract). [本文引用: 1]
Sun FM, LiuJ, HuaW, Sun XC, Wang XF, Wang HZ. Identification of stable QTLs for seed oil content by combined linkage and association mapping in Brassica napus Plant Sci, 2016,252:388-399. URLPMID:27717475 [本文引用: 1]
YuJ, Holland JB, McMullen MD, Buckler ES. Genetic design and statistical power of nested association mapping in maize Genetics, 2008,178:539-551. [本文引用: 1]
LiH, BradburyP, ErsozE, Buckler ES, WangJ. Joint QTL linkage mapping for multiple-cross mating design sharing one common parent PLoS One, 2011,6:e17573. [本文引用: 1]
XavierA, XuS, Muir WM, Rainey KM. NAM: association studies in multiple populations Bioinformatics, 2015,31:3862-3864. [本文引用: 1]
BouchetS, Olatoye MO, MarlaSR, PerumalR, TessoT, Yu JM, TuinstraM, Morris GP. Increased power to dissect adaptive traits in global sorghum diversity using a nested association mapping population Genetics, 2017,206:573-585. [本文引用: 1]
SchnaithmannF, KopahnkeD, PillenK. A first step toward the development of a barley NAM population and its utilization to detect QTLs conferring leaf rust seedling resistance Theor Appl Genet, 2014,127:1513-1525. [本文引用: 1]
XavierA, JarquinD, HowardR, RamasubramanianV, Specht JE, Graef GL, Beavis WD, Diers BW, SongQ, Cregan PB, NelsonR, MianR, Shannon JG, McHaleL, WangD, SchapaughW, Lorenz AJ, XuS, Muir WM., Rainey KM. Genome-wide analysis of grain yield stability and environmental interactions in a multiparental soybean population G3: Genes Genom Genet, 2018,8:519-529. [本文引用: 1]
Khan MA, TongF, WangW, HeJ, ZhaoT, GaiJ. Analysis of QTL-allele system conferring drought tolerance at seedling stage in a nested association mapping population of soybean [ Glycine max (L.) Merr.] using a novel GWAS procedure Planta, 2018,248:947-962. [本文引用: 1]
ChenQ, Yang CJ, York AM, XueW, Daskalska LL, DeValk CA, Krueger KW, Lawton SB, Spiegelberg BG, Schnell JM, Neumeyer MA, Perry JS, Peterson AC, KimB, BergstromL, YangL, Barber IC, TianF, Doebley JF. TeoNAM: a nested association mapping population for domestication and agronomic trait analysis in maize Genetics, 2019,213:1065-1078. [本文引用: 1]
BecheE, Gillman JD, SongQ, NelsonR, BeissingerT, DeckerJ, ShannonG, Scaboo AM. Nested association mapping of important agronomic traits in three interspecific soybean populations Theor Appl Genet, 2020,133:1039-1054. [本文引用: 1]
GarinV, WimmerV, MezmoukS, MalosettiM, EeuwijkF. How do the type of QTL effect and the form of the residual term influence QTL detection in multi-parent populations? A case study in the maize EU-NAM population Theor Appl Genet, 2017,130:1753-1764. URLPMID:28547012 [本文引用: 1]
Monir MM, ZhuJ. Dominance and epistasis interactions revealed as important variants for leaf traits of maize NAM population Front Plant Sci, 2018,9:627. [本文引用: 1]
HuJ, GuoC, WangB, YeJ, LiuM, WuZ, XiaoY, ZhangQ, LiH, King GJ, LiuK. Genetic properties of a nested association mapping population constructed with semi-winter and spring oilseed rapes Front Plant Sci, 2018,9:1740. URLPMID:30534135 [本文引用: 1]
Jordan KW, WangS, HeF, ChaoS, LunY, PauxE, SourdilleP, ShermanJ, AkhunovaA, Blake NK, Pumphrey MO, GloverK, DubcovskyJ, TalbertL, Akhunov ED. The genetic architecture of genome-wide recombination rate variation in allopolyploid wheat revealed by nested association mapping Plant J, 2018,95:1039-1054. [本文引用: 1]
Kidane YG, Gesesse C A. Hailemariam B N, Desta EA, Mengistu DK, FaddaC, Pè ME, Dell’AcquaM . A large nested association mapping population for breeding and quantitative trait locus mapping in Ethiopian durum wheat Plant Biotechnol J, 2019,17:1380-1393. [本文引用: 1]
Marla SR., BurowG, ChopraR, HayesC, Olatoye MO, FelderhoffT, HuZ, RaymundoR, PerumalR, Morris GP. Genetic architecture of chilling tolerance in sorghum dissected with a nested association mapping population G3: Genes Genom Genet, 2019,9:4045-4057. [本文引用: 1]
 ,1,**, 孙子淇1,**, 刘华1, 房元瑾1, 石磊1, 苗利娟1, 张毛宁1, 张忠信1, 徐静1, 张梦圆2, 董文召1, 张新友
,1,**, 孙子淇1,**, 刘华1, 房元瑾1, 石磊1, 苗利娟1, 张毛宁1, 张忠信1, 徐静1, 张梦圆2, 董文召1, 张新友
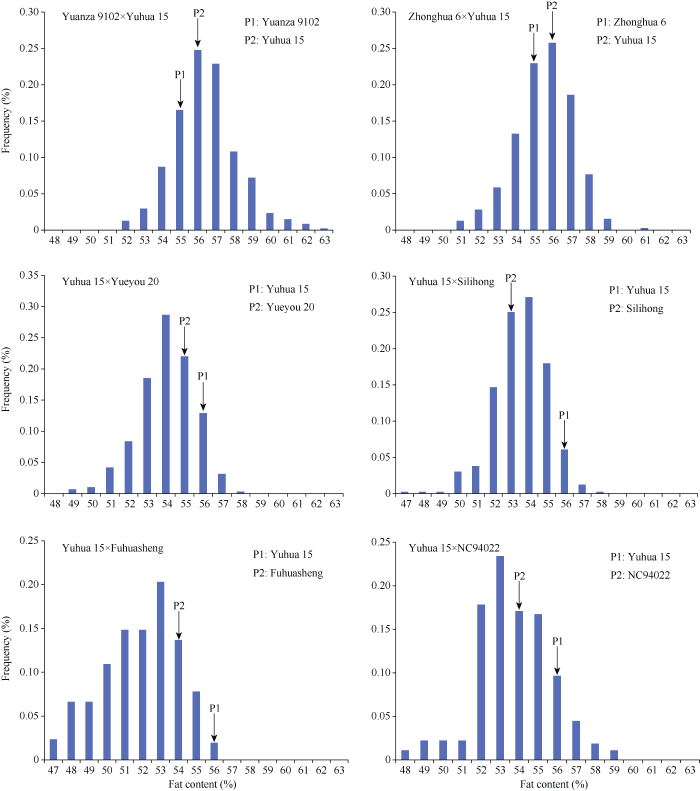 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}