Regulation of ACC treatment on nitrogen supply response of maize seedlings with different genotypes
WU Bing-Hui,**, WANG Gui-Ping,**, WANG Yu-Bin, LI Zhao-Hu, ZHANG Ming-Cai,*Engineering Research Center of Plant Growth Regulator, Ministry of Education / College of Agronomy and Biotechnology, China Agricultural University, Beijing 10093, China
Abstract Ethylene plays an important role in the growth and morphological formation of maize, while little research is involved in the regulatory effects of ethylene on the nitrogen absorption and accumulation in maize, which limits the application of ethylene in high yield and efficient cultivation of maize production. In this study, three maize varieties (Zhengdan 958, Ruifuer 1, and Demeiya 3) were used as experimental materials to study the response of maize varieties with different nitrogen absorption efficiency under different nitrogen levels condition. Combined with the addition of the precursor of ethylene synthesize 1-aminocyclopropane-1-carboxylic acid (ACC), the regulation effects of ethylene on nitrogen uptake of different genotypes were analyzed in maize. The results showed that nitrogen-sensitive varieties (Ruifuer 1 and Demeiya 3) had more obvious phenotype of nitrogen deficiency than Zhengdan 958 under low nitrogen condition, and were more sensitive to ACC treatment. Moreover, ACC treatment repressed the growth and dry matter accumulation in the shoot and root of maize plants. ACC treatment decreased the chlorophyll content of leaves under low nitrogen, reduced the accumulation of soluble protein in leaves, and promoted the premature aging of maize leaves. Among them, the contents of chlorophyll and soluble protein in Zhengdan 958 leaves with ACC treatment were significantly higher than those of Ruifuer 1 and Demeiya 3. Furthermore, low nitrogen treatment inhibited the expression of key enzymes (ZmACS7 and ZmACO15) in ethylene synthesis, while decreased the ethylene release rate. Thus, ACC treatment promoted the expression of ZmACS7 and ZmACO15 and enhanced ethylene release rate under low nitrogen treatments. Low nitrogen treatment inhibited the expression of ZmNRT2.1 in maize roots, but ACC treatment promoted the expression of ZmNRT2.1 in maize roots. In addition, the expression of ZmNRT2.1 in roots of Zhengdan 958 was significantly higher than those of Ruifuer 1 and Demeiya 3 under low nitrogen treatments. The results showed that ethylene regulated the uptake and distribution of nitrogen by regulating the key enzyme genes of ethylene synthesis and the expression of ZmNRT2.1 in maize which affected plants growth , and nitrogen-sensitive varieties were more sensitive to ethylene than green-holding variety. Keywords:maize;genotype;nitrogen absorption;nitrate nitrogen;1-aminocyclopropane-1-carboxylic acid
PDF (546KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 吴冰卉, 王桂萍, 王玉斌, 李召虎, 张明才. ACC处理对不同基因型玉米幼苗响应氮素供给的调控效应[J]. 作物学报, 2021, 47(5): 799-806. doi:10.3724/SP.J.1006.2021.03047 WU Bing-Hui, WANG Gui-Ping, WANG Yu-Bin, LI Zhao-Hu, ZHANG Ming-Cai. Regulation of ACC treatment on nitrogen supply response of maize seedlings with different genotypes[J]. Acta Agronomica Sinica, 2021, 47(5): 799-806. doi:10.3724/SP.J.1006.2021.03047
A和B分别为ACC处理7 d后不同基因型玉米植株在不同氮素条件下根系和地上部的干重; C为ACC处理7 d后不同基因型玉米植株在不同氮素条件下的根冠比。 Fig. 1Effects of ACC on the dry weight and the ratio of root and shoot in different maize genotypes under different nitrogen treatments
A and B indicate the dry weight of roots and shoots of different maize genotypes under the different nitrogen conditions after 7 days of ACC treatment, respectively; C indicates the root-to-shoot ratio of different maize genotypes under different nitrogen conditions after 7 days with ACC treatment. LN: low nitrogen level; SN: sufficient nitrogen level.
Cui AM, Zhang JG, ZhangH, ShanH, ChenW. Preliminary exploration on current situation and development of maize production in China J Agric Sci Technol, 2020,22(7):10-19 (in Chinese with English abstract). [本文引用: 1]
Hou YP, Kong LL, YangJ, Yin CX, Qin YB, LiQ, YuL, Wang LC, Xie JG. Effects of nitrogen fertilizer management on yield, nutrient absorption and translocation of spring maize J Maize Sci, 2016,24(4):137-143 (in Chinese with English abstract). [本文引用: 1]
Yin MH, Li YN, LiH, Xu YB, Zhou CM, Zhang TL. Effects of nitrogen application rates on root growth and nitrogen use of summer maize Trans CSAM, 2016,47(6):129-138 (in Chinese with English abstract). [本文引用: 1]
Zhang LL, QiH, FanY, Yang HL, FuJ, Xu SJ, Jing XQ, WangP. Effects of postponing N application on dry matter accumulation and nitrogen utilization in maize canopy J Maize Sci, 2017,25(1):127-132 (in Chinese with English abstract). [本文引用: 1]
Ciampitti IA, Vyn TJ. Grain nitrogen source changes over time in maize: a review Crop Sci, 2013,53:366-377. [本文引用: 1]
Chen YL, Xiao CX, Chen XC, LiQ, ZhangJ, Chen FJ, Yuan LX, Mi GH. Characterization of the plant traits contributed to high grain yield and high grain nitrogen concentration in maize Field Crops Res, 2014,159:1-9. [本文引用: 1]
Chen YL, Xiao CX, Wu DL, Xia TT, Chen QW, Chen FJ, Yuan LX, Mi GH. Effects of nitrogen application rate on grain yield and grain nitrogen concentration in two maize hybrids with contrasting nitrogen remobilization efficiency Eur J Agron, 2015,62:79-89. [本文引用: 1]
GuoS, Sun WY, Gu RL, Wang ZK, Chen FJ, Zhao BQ, Yuan LX, Mi GH. Differences in leaf nitrogen remobilization efficiency and related gene expression during grain filling stage of two maize hybrids Plant Nutr Fert Sci, 2018,24:1149-1157 (in Chinese with English abstract). [本文引用: 1]
Khan M IR, TrivelliniA, FatmaM, MasoodA, FranciniA, IqbalN, FerranteA, Khan NA. Role of ethylene in responses of plants to nitrogen availability Front Plant Sci, 2015,6:1-20. [本文引用: 2]
ShekoofaA, EmamY. Plant growth regulator (ethephon) alters maize (Zea mays L.) growth, water use and grain yield under water stress J Agron, 2008,7:41-48. [本文引用: 1]
Ye DL, Zhang YS, Al-KaisiM M, DuanL S, ZhangM C, Li ZH. Ethephon improved stalk strength associated with summer maize adaptations to environments differing in nitrogen availability in the North China Plain J Agric Sci, 2016,154:960-977. [本文引用: 1]
YuY, LuS, Song HX, Guan CY, Chen KH, Zhang ZH. Regulation mechanisms of NO3- re-utilization by ACC at later growth stages of Brassica napus Plant Nutr Fert Sci, 2017,23:1378-1386 (in Chinese with English abstract). [本文引用: 1]
LeblancA, RenaultH, LecourtJ, EtienneP, DeleuC, Le DE. Elongation changes of exploratory and root hair systems induced by aminocyclopropane carboxylic acid and aminoethoxyvinylglycine affect nitrate uptake and BnNrt2.1 and BnNrt1.1 gene expression in oil rape Plant Physiol, 2008,146:1928-1940. [本文引用: 1]
LeblancA, SeguraR, DeleuC, Le DE. In low transpiring conditions, uncoupling the BnNrt2.1 and BnNrt1.1 NO3- transporters by glutamate treatment reveals the essential role of BnNRT2.1 for nitrate uptake and the nitrate-signalling cascade during growth Plant Signal Behav, 2013,8:e22904. [本文引用: 1]
IqbalN, NazarR, SyeedS, MasoodA, Khan NA. Exogenously-sourced ethylene increases stomatal conductance, photosynthesis, and growth under optimal and deficient nitrogen fertilization in mustard J Exp Bot, 2011,62:4955-4963. [本文引用: 1]
Ye DL, Wang YB, ZhouL, Li JM, Duan LS, Zhang MC, Li ZH. Effect of ethephon and nitrogen fertilizer on nitrogen uptake, nitrogen use efficiency and yield of summer maize Acta Agron Sin, 2015,41:1701-1710 (in Chinese with English abstract). [本文引用: 1]
KurtykaR, Ma?kowskiE, KitaA, KarczW. Effect of calcium and cadmium on growth and accumulation of cadmium, calcium, potassium and sodium in maize seedlings Pol J Environ Stud, 2008,17:51-56. [本文引用: 1]
Bremner JM, Mulvaney CS. Methods of Soil Analysis, Part 2 Chemical and Microbiological Properties. Madison, WI: Soil Science Society of America, Inc., 1982. pp 595-624. [本文引用: 1]
Abdel-Ghani AH, KumarB, Reyes-MatamorosJ, Gonzalez- Portilla PJ, JansenC, Martin J PS, LeeM, LübberstedtT. Genotypic variation and relationships between seedling and adult plant traits in maize ( Zea mays L.) inbred lines grown under contrasting nitrogen levels Euphytica, 2013,189:123-133. [本文引用: 1]
SchlüterU, MascherM, ColmseeC, ScholzU, Br?utigamA, FahnenstichH, SonnewaldU. Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis Plant Physiol, 2012,160:1384-1406. [本文引用: 2]
CoqueM, MartinA, Veyrieras JB, HirelB, GallaisA. Genetic variation for N-remobilization and postsilking N-uptake in a set of maize recombinant inbred lines: 3. QTL detection and coincidences Theor Appl Genet, 2008,117:729-747. [本文引用: 1]
Lee RB, Drew MC. Nitrogen-13 studies of nitrate fluxes in barley roots J Exp Bot, 1986,185:1768-1779. [本文引用: 1]
Tsay YF, Chiu CC, Tsai CB, Ho CH, Hsu PK. Nitrate transporters and peptide transporters FEBS Lett, 2007,581:2290-2300. [本文引用: 1]
LiW, WangY, OkamotoM, Crawford NM, Siddiqi MY, Glass A DM. Dissection of the AtNRT2.1: AtNRT2.2 inducible high-affinity nitrate transporter gene cluster Plant Physiol, 2007,143:425-433. [本文引用: 1]
TaulemesseF, Le GouisJ, GouacheD, GibonY, AllardV. Post-flowering nitrate uptake in wheat is controlled by N status at flowering, with a putative major role of root nitrate transporter NRT2.1 PLoS One, 2015,10:e0120291. [本文引用: 1]
LiY, Li JQ, YanY, Liu WQ, Zhang WN, Gao LH, Tian YQ. Knock-down of csNRT2.1, a cucumber nitrate transporter, reduces nitrate uptake, root length, and lateral root number at low external nitrate concentration Front Plant Sci, 2018,9:1-13. [本文引用: 1]
TrevisanS, BorsaP, BottonA, VarottoS, MalagoliM, RupertiB, QuaggiottiS. Expression of two maize putative nitrate transporters in response to nitrate and sugar availability Plant Biol (Stuttg), 2008,10:462-475. [本文引用: 1]
Tian QY, SunP, Zhang WH. Ethylene is involved in nitrate-dependent root growth and branching in Arabidopsis thaliana New Phytol, 2009,184:918-931. [本文引用: 2]
Zheng DC, HanX, AnY, Guo HW, Xia XL, YinW. The nitrate transporter NRT2.1 functions in the ethylene response to nitrate deficiency in Arabidopsis Plant Cell Environ, 2013,36:1328-1337. [本文引用: 1]
Johnson PR, Ecker JR. The ethylene gas signal transduction pathway: a molecular perspective Annu Rev Genet, 1998,32:227-254. [本文引用: 1]
TsuchisakaA, TheologisA. Unique and overlapping expression patterns among the Arabidopsis 1-amino-cyclopropane-1-carboxylate synthase gene family members Plant Physiol, 2004,136:2982-3000. [本文引用: 1]
YangS, HoffmanN. Ethylene biosynthesis and its regulation in higher plants Annu Rev Plant Physiol, 1984,35:155-189. [本文引用: 1]
 ,**, 王桂萍
,**, 王桂萍
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT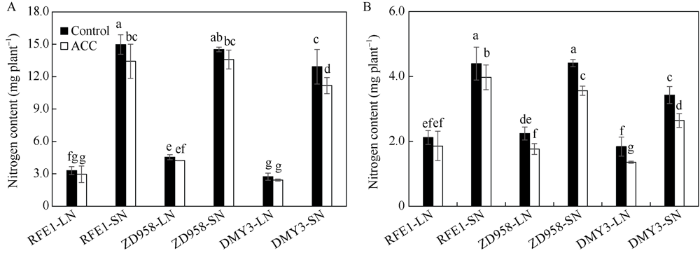 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT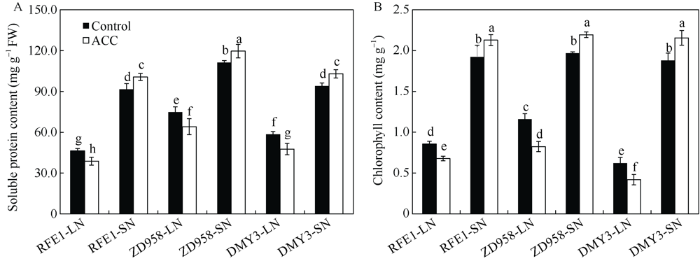 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT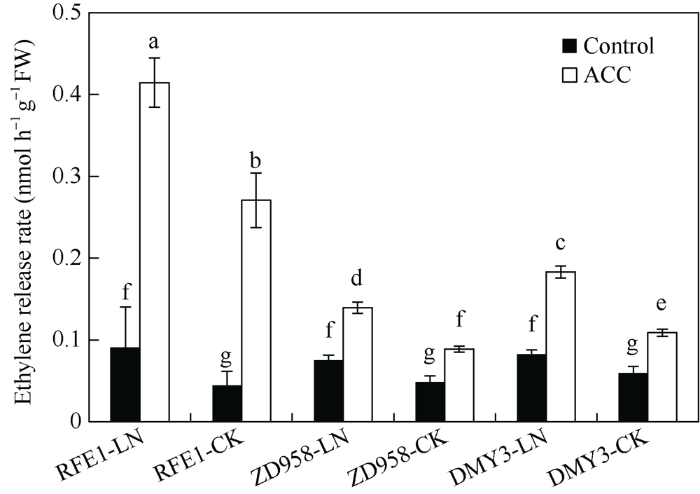 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT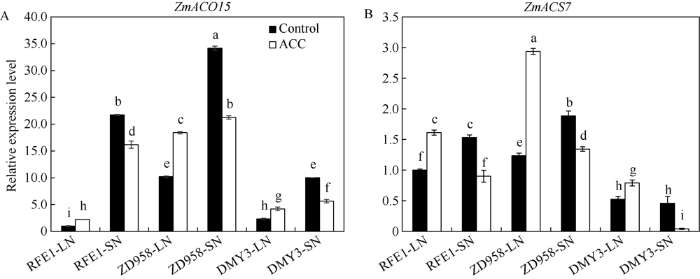 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT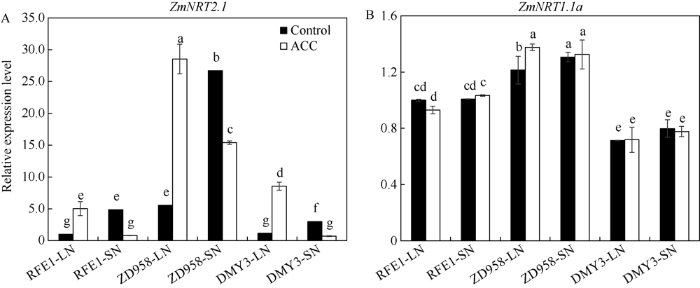 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}