 ,, 冼俊霖, 孙超, 王春明, 石丽, 于为常,*深圳大学生命与海洋学院/广东省植物表观遗传学重点实验室, 广东深圳 518071
,, 冼俊霖, 孙超, 王春明, 石丽, 于为常,*深圳大学生命与海洋学院/广东省植物表观遗传学重点实验室, 广东深圳 518071Preliminary study of genome editing of peanut FAD2 genes by CRISPR/Cas9
ZHANG Wang,, XIAN Jun-Lin, SUN Chao, WANG Chun-Ming, SHI Li, YU Wei-Chang,*College of Life Sciences and Oceanography, Shenzhen University/Guangdong Provincial Key Laboratory for Plant Epigenetics, Shenzhen 518071, Guangdong, China通讯作者: * 于为常, E-mail:wyu@szu.edu.cn
收稿日期:2020-09-19接受日期:2021-01-13网络出版日期:2021-08-12
| 基金资助: |
Received:2020-09-19Accepted:2021-01-13Online:2021-08-12
| Fund supported: |
作者简介 About authors
E-mail:zahngwang@foxmail.com

摘要
油酸脱氢酶(Δ12FAD或FAD2)是催化油酸(OA)的C12位上脱氢生成双不饱和亚油酸(LA)的关键酶, 它控制油酸、亚油酸的含量及其比值(O/L)。研究表明, AhFAD2是油酸生成亚油酸的关键基因, 决定花生种子中油酸和亚油酸的相对含量。本研究根据AhFAD2基因序列, 设计了相应sgRNA序列, 并构建了旨在敲除FAD2A和FAD2B两个花生油酸脱氢酶的CRISPR/Cas9基因编辑载体。经花生基因转化后, 通过对转基因花生突变位点基因组序列分析找出基因突变体。对靶基因分析发现, 16株转基因花生含有29个FAD2A基因突变, 其中16个突变引起蛋白质序列变化; 11株转基因花生含有30个FAD2B基因突变, 其中17个突变引起蛋白质序列变化。FAD2A和FAD2B蛋白质序列的变化可影响花生油酸脱氢酶的活性, 改变油酸催化脱氢, 使亚油酸合成受阻, 油酸含量增加。本研究为研究花生脂肪酸合成及高油酸花生育种提供了宝贵的基因突变体材料。
关键词:
Abstract
Oleate dehydrogenase (Δ12FAD or FAD2) is the key enzyme catalyzing the dehydrogenation of oleic acid (OA) at the C12 position to produce diunsaturated linoleic acid (LA). It controls the contents and ratios (O/L) of oleic acid and linoleic acid in plants. Increasing evidences in molecular biology research indicate that AhFAD2 is the key gene for the conversion of oleic acid to linoleic acid, and determines the relative content of oleic acid and linoleic acid in peanut seeds. In this study, the corresponding sgRNA sequences were designed based on AhFAD2 gene sequences, and a CRISPR/Cas9 gene editing vector was constructed to mutate the peanut FAD2A and FAD2B genes. After peanut gene transformation, gene mutations were identified by genomic sequence analysis of transgenic peanut flanking the sgRNA target sites. Target gene analysis indicated that 29 mutations of FAD2A gene in 16 transgenic peanut plants were obtained, among which 16 mutations caused protein sequence changes; 30 mutations in 11 transgenic peanut plants contained mutations in FAD2B gene, among which 17 mutations caused changes in protein sequence. Changes in the protein sequences of the FAD2A and FAD2B genes might affect the enzyme activity, change the catalytic dehydrogenation of oleic acid, hinder the synthesis of linoleic acid, and thus increase the content of peanut oleic acid. These FAD2 gene mutants are valuable in the study of fatty acid metabolism and the breeding of high oleic peanuts.
Keywords:
PDF (3787KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张旺, 冼俊霖, 孙超, 王春明, 石丽, 于为常. CRISPR/Cas9编辑花生FAD2基因研究. 作物学报[J], 2021, 47(8): 1481-1490 DOI:10.3724/SP.J.1006.2021.04214
ZHANG Wang, XIAN Jun-Lin, SUN Chao, WANG Chun-Ming, SHI Li, YU Wei-Chang.
花生(Arachis hypogaea L.)又名落花生、长生果, 主要生长在半干旱和热带国家, 是我国产量丰富、食用广泛的一种坚果[1]。同时, 它是世界上重要的食用植物油和蛋白质来源, 在全球油脂生产中具有举足轻重的地位。目前, 我国花生产量约占油料总产量的一半, 是我国产值最大的油料作物[2,3]。营养学家将花生评为“A+”级别的农作物, 原因是花生包含40%~56%的油脂、20%~30%的蛋白质、10%~20%的碳水化合物、以及维生素E、烟酸、多酚、钙、镁、磷、锌、铁、钾、核黄素、硫胺素、植物甾醇和膳食纤维等多种营养成分, 对平衡膳食、改善人们的营养与健康状况具有重要作用[4,5]。
花生油主要由单不饱和脂肪酸油酸(oleic acid, C18:1, Δ9)、多不饱和脂肪酸亚油酸(linoleic acid, C18:2, Δ9, Δ12)、饱和脂肪酸棕榈酸(palmitic acid, C16:0)和硬脂酸(stearic acid, C18:0)组成。不饱和脂肪酸约为80%, 其中油酸36%~67%、亚油酸15%~43%, 饱和脂肪酸占20%, 其中棕榈酸6%~11%、硬脂酸2%~6%、花生酸5%~7%、山嵛酸2%~3%。脂肪酸的组成是决定花生油质量的重要因素, 也是衡量其品质优劣的重要指标[6]。其中油酸和亚油酸的含量及比值(油/亚比)对花生及其制品的稳定性起决定性作用。亚油酸属多不饱和脂肪酸, 其油酰残基易氧化而导致油脂腐败劣变, 严重影响油脂储存期, 而油酸分子结构只含有1个不饱和键, 化学结构相对比较稳定, 抗氧化能力强[7]。油酸的自氧化性比亚油酸稳定10倍, 油酸/亚油酸(O/L)比例高的花生更耐储存, 在精炼、储藏和煎炸时不易变质, 与普通花生相比, 高油酸含量的花生制品具有更长的氧化诱导期和货架期[8]。在食品健康越来越受重视的今天, 高油酸比例的食用油是众多食品加工和非饱和脂肪酸使用的一个良好选择, 其开发和应用的前景十分广阔。
油酸脱氢酶(Δ12FAD或FAD2)是催化油酸的C12位上脱氢生成双不饱和亚油酸的关键酶[9,10], 它控制油酸、亚油酸的含量及其比值(O/L)。在花生中该酶由AhFAD2基因编码。花生高油酸性状主要由AhFAD2基因突变产生, FAD2基因编码区序列变化所引起编码蛋白活性的差异与花生籽粒油酸、亚油酸含量密切相关, 它决定了花生仁中油酸和亚油酸的含量[11]。栽培种花生中AhFAD2由位于不同亚基因组上的2对非等位同源基因AhFAD2A和AhFAD2B共同编码。在普通油酸含量花生品种中这2个基因或者其中之一能正常表达, 而在高油酸品种中, AhFAD2A和AhFAD2B基因均突变造成功能缺陷, 共同导致高油酸性状的产生[11]。普通花生的不饱和脂肪酸中含有约45%的油酸和35%的亚油酸, 而含有AhFAD2A和AhFAd2B基因突变的高油酸花生中油酸含量可达80%, 而亚油酸的含量则减少到2%~3% [12]。因此, 高油酸花生育种的关键是油酸脱氢酶(FAD2)基因的突变。
基因组编辑技术的发展为创造高油酸花生突变体提供了新的途径。基因组编辑技术主要是利用序列特异性核酸酶 (sequence specific nucleases, SSNs)在特定基因位点产生DNA双链断裂, 借助编辑受体自身的DNA修复系统在非同源末端连接(non-homologous end joining, NHEJ)过程中产生随机的Indels (small insertions and deletions)或在同源重组(homologous recombination)修复过程中插入或替换相应的基因片段, 最终实现碱基突变、DNA片段缺失和插入等现象, 使原本的基因不能正常表达。CRISPR/Cas9基因组编辑系统具有突变效率高、准确度高、设计构建表达载体较为简单等特点, 成为当前主流的基因组编辑方法。目前该技术已成功应用于人类细胞、小鼠、斑马鱼、果蝇、线虫、大鼠、水稻、拟南芥、小麦、烟草等基因组精确修饰(包括碱基突变, 插入突变和缺失突变)[13,14]。本研究利用CRISPR/Cas9基因组编辑技术, 创造花生FAD2基因突变体材料, 旨在为研究花生脂肪酸合成及高油酸花生育种提供宝贵的种质资源。
1 材料与方法
1.1 试验材料
栽培花生(Arachis hypogaea L.)鲁花11, 2018— 2020年种植于深圳市龙华区深圳大学研究基地。1.2 试验方法
1.2.1 花生FAD2A和FAD2B基因的克隆 根据栽培花生delta-12 fatty acid desaturase (FAD) mRNA序列(GenBank: EF186911.1)设计引物: FAD2F: 5°-ATGGGAGCTGGAGGGCGTGTCACT-3′和FAD 2R: 5°-TCAGAACTTGTTCTTGTAC CAATA-3°。取1 g花生幼嫩叶片, 放液氮中快速冷冻并研磨, 用植物基因组DNA提取试剂盒(天根生化北京科技有限公司)提取基因组DNA, 通过高效高保真PCR扩增试剂盒(天根)扩增FAD2A和FAD2B基因。PCR反应体系包含5 μL 200 ng基因组DNA、引物1 (10 μmol L-1)和引物2 (10 μmol L-1)各2 μL、10 μL 5× buffer、1 μL Fast HiFidelity DNA聚合酶、2.5 μL 20× Fast PCR Enhancer, 加ddH2O至50 μL。PCR反应条件为94℃ 2 min; 94℃ 15 s, 60℃ 10 s, 68℃ 60 s, 35个循环; 68℃ 5 min。PCR产物利用Hieff Clone Plus一步法快速克隆试剂盒(翊圣生物科技(上海)有限公司)克隆后测序。1.2.2 CRISPR/Cas9基因组编辑载体的构建 通过NCBIblast比对找到花生FAD2A和FAD2B基因序列保守区域。利用CRISPR设计程序Cas-Designer根据上述FAD2A和FAD2B基因保守区域序列, 以野生花生(Arachis duranensis)基因组序列(PeantBase v1.0)为参考基因组, 设计2条sgRNA序列, 分别为T1-TCAAACCCTCCATTCAGTGT(TGG)和T2-TCA TATAACTGATACGCATG(TGG)。设计2条PCR引物, 分别为引物1: 5°-ATATATGGTCTCGATTGTCAAACCCTCCATTCAGTGTGTTTTAGAGCTAGAAATAGCAAGTT-3°; 引物2: 5°-ATTATTGGTCTCT AAACCATGCGTATCAGTTATATGACAATCTCTTAGTCGACTCTACCAAT-3°, 下画线部分为T1和T2 sgRNA靶点序列, 黑体部分为BSAI酶识别位点, 引物1和2的3°端分别与pCBC-DT1T2的gRNA-Sc和U6-29p配对(图1)。以pCBC-DT1T2为模板, 利用上述引物进行PCR扩增, PCR片段通过BSAI位点克隆到pSKE401载体[15], 构建含T1和T2两个靶点的CRISPR载体FAD2T1T2 (图1)。
图1
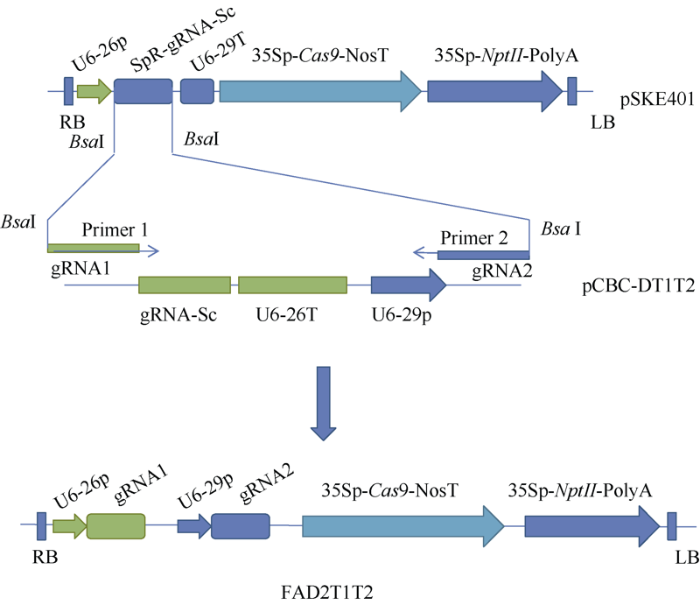 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1基因组编辑载体的构建
合成含有gRNA1和gRNA2序列的引物1和引物2, 以pCBC-DT1T2载体为模版合成sgRNA片段, 并通过引物1和引物2上的Bsa I酶切位点, 克隆到pSKE401载体, 获得含2个靶点的基因组编辑载体FAD2T1T2。LB、RB分别为农杆菌T-DNA的左右边界; U6-26p、U6-29p为拟南芥U6基因启动子; U6-29T、U6-26T为转录终止子; 35Sp-Cas9-NosT、35Sp-NptII-polyA分别为Cas9和NptII基因表达框。
Fig. 1Construction of genome editing vector
Primer 1 and primer 2 containing gRNA1 and gRNA2 sequences, are used to synthesize sgRNA by PCR with pCBC-DT1T2 as template. The synthesized sgRNA was cloned into pSKE401 plasmid by Bsa I restriction enzyme sites to obtain FAD2T1T1 genome editing construct. LB, RB denote T-DNA left and right border sequences; U6-26p, U6-29p are promoters while U6-29T and U6-26T are terminators of Arabidopsis U6 gene; 35Sp-Cas9-NosT, 35Sp-NptII-polyA are gene expression cassettes of Cas9 and NptII genes.
1.2.3 花生基因转化 花生的基因转化参考Livingstone和Birch的方法[16]。成熟花生种子用20% Clorox (含1% w/v次氯酸钠)表面消毒2次, 每次20 min, 无菌水冲洗3次。在超净台中切去花生胚芽, 去掉子叶及胚根, 放在MS5PG (4.4 g L-1 MS basal salts, 蔗糖30.0 g L-1, 谷氨酰胺1.0 g L-1, Picloram 5.0 mg L-1, 琼脂粉8 g L-1, pH 5.8)花生愈伤组织诱导培养基上, 26~28℃暗培养, 诱导胚性愈伤组织。胚性愈伤组织每2~3周继代1次, 4~6周后转入MS3PG (4.4 g L-1 MS basal salts, 蔗糖30.0 g L-1, 谷氨酰胺1.0 g L-1, Picloram 3.0 mg L-1, 琼脂粉8 g L-1, pH 5.8)花生愈伤组织继代培养基上, 进行胚状体的扩增及保持。
花生胚性愈伤组织在MS3PGO (MS3PG + 甘露醇0.2 mol L-1 + 山梨醇0.2 mol L-1)渗透压培养基上进行预处理4 h, 利用基因枪PDS-1000/HE进行转化, 金粉颗粒为0.6 μm, 每个转化质粒DNA用量为2 μg, 转化压力为650 PSI, 转化距离为6 cm。转化后在MS3PGO培养基上保留过夜, 然后重新转到MS3PG培养基上, 26~28℃暗培养7 d后转入筛选培养基MS3PGK50 (MS3PG + 卡那霉素50 mg L-1), 26~28℃暗培养8~10周, 每3~4周换1次培养基。
筛选8~10周后, 抗性愈伤组织转移到MSC (4.4 g L-1 MS basal salts, 蔗糖20.0 g L-1, 活性炭1.0 g L-1, 卡那霉素50 mg L-1, 琼脂粉8 g L-1, pH 5.8)分化培养基, 26~28℃光照培养(每日16 h光照) 4~6周, 胚状体转移到MS10 (4.4 g L-1 MS basal salts, 蔗糖20.0 g L-1, BAP 10 mg L-1, 卡那霉素50 mg L-1, 琼脂粉8 g L-1, pH 5.8)培养基, 26~28℃光照培养(每日16 h光照), 每3~4周继代1次, 直至长成小苗, 然后将小苗转入MSOR (4.4 g L-1 MS basal salts, 蔗糖20.0 g L-1, NAA 1.0 mg L-1, 卡那霉素50 mg L-1, 琼脂粉8 g L-1, pH 5.8)生根培养基, 26~28℃光照培养(每日16 h光照), 诱导生根, 三至四叶大的生根幼苗移栽到花盆土中, 在培养室中光照培养直至开花结果。
移栽成活的幼苗, 取幼苗叶片, 用植物基因组DNA提取试剂盒提取基因组DNA。根据pKSE401载体上Cas9基因序列, 设计PCR引物Cas9F: 5°-GACAAGAAGTACTCGATCGGCCTCG-3°和Cas9R: 5°-TCGTAGGGTACTTCTCGTGGTAGGC-3°。以花生幼苗基因组DNA为模板, 通过PCR进行转基因验证。
以FDA2T1T2转基因花生基因组DNA为模板, FAD2F和FAD2R为引物进行PCR扩增。将扩增PCR片段进行克隆, 然后挑选单克隆进行测序。每个基因转化植株挑选10个单克隆进行测序, 测序结果通过NCBI网站BLAST程序和野生型植株基因序列比对, 鉴定突变基因。
2 结果与分析
2.1 FAD2A和FAD2B基因序列分析
花生FAD2A和FAD2B基因编码序列皆为1140 bp, 各编码1个379氨基酸的蛋白。基因序列比对发现, FAD2B基因和EF186911.1序列完全一致, FAD2A基因和EF186911.1序列相比有10个SNP, 蛋白质序列有3个氨基酸的变化, 同源性均大于99%。分别在5°和3°端核酸序列同源区设计2个sgRNA靶点T1和T2 (图2), 用于CRISPR载体的构建。图2
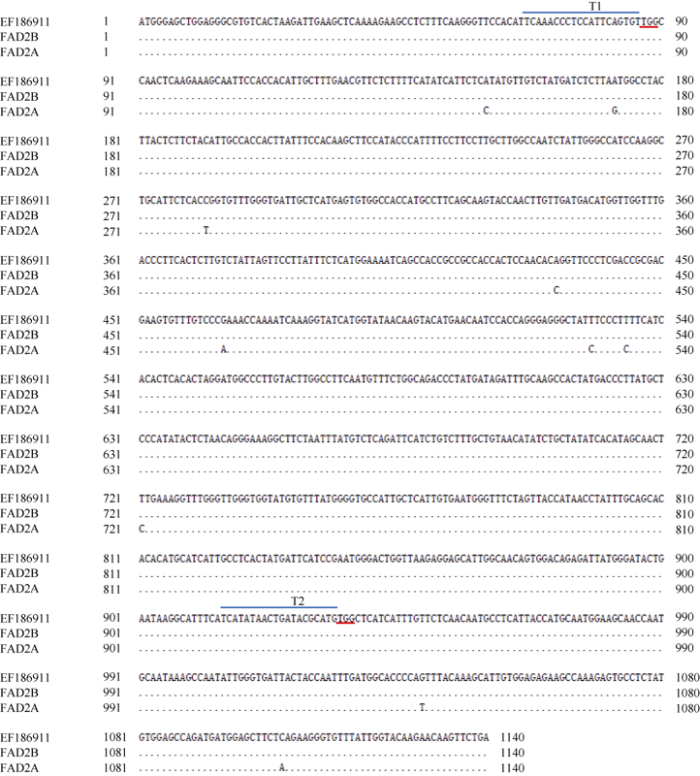 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2花生FAD2A, FAD2B基因序列分析及基因组编辑载体靶点的选择
黑点表示相同序列, T1和T2标记基因组编辑靶点位置, 红色下画线标记PAM (NGG)序列。
Fig. 2 FAD2A and FAD2B gene alignment and the selection of CRISPR target sites in peanut
Black dots denote identical nucleotides; T1 and T2 indicate the target sequences of genome editing; PAM sequences are underlined with red lines.
2.2 转基因花生分析
利用CRISPR载体FAD2T1T2 (图3-A)通过基因枪转化鲁花11胚性愈伤组织, 经过含卡那霉素的培养基筛选, 共获得16株抗性花生植株(图3- B)。取转基因花生幼叶提取基因组DNA, 利用CAS9序列引物, 通过PCR对转基因花生植株进行验证发现, 其中12株含有转基因成分(图3-C)。为研究基因组编辑载体对花生FAD2基因的编辑情况, 以12株转基因花生的基因组DNA为模板, FAD2F和FAD2R为引物进行PCR扩增, 并对克隆的PCR产物进行测序, 发现FAD2A和FAD2B基因的突变(图3-D, 表1, 表2)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3花生基因转化及转基因植株的分析
A: 转基因载体, T1、T2分别为gRNA1和gRNA2靶点序列; B: 转基因愈伤及再生植株, 左侧为抗性愈伤组织, 右侧为转基因植株; C: 转基因植株鉴定, 扩增条带为Cas9基因片段, 箭头指示400 bp PCR扩增产物; D: 突变体序列分析, 转基因植株通过测序检测基因编辑突变体。
Fig. 3Gene transformation and analysis of transgenic lines in peanut
A: gene transformation vector. T1 and T2 are gRNA1 and gRNA2 target sites; B: transgenic calli (left) and plantlets (right); C: PCR identification of transgenic plants. The amplified band is Cas9 gene fragment, and the arrow denotes 400 bp DNA marker; D: sequence analysis of genome editing mutants.
Table 1
表1
表1FAD2A基因突变
Table 1
| 突变位点 Mutation site | 核酸Nucleic acid | 密码子Codon | 氨基酸Amino acid | ||||
|---|---|---|---|---|---|---|---|
| 突变前Before mutation | 突变后 After mutation | 突变前Before mutation | 突变后 After mutation | 突变前Before mutation | 突变后 After mutation | ||
| 40 | A | G | AAG | GAG | K | E | |
| 150 | C | A | TCC | TCA | |||
| 172 | G | A | GTG | ATG | V | M | |
| 214 | A | C | AAG | CAG | K | Q | |
| 282 | T | C | ACT | ACC | |||
| 333 | C | T | TAC | TAT | |||
| 362 | C | T | ACC | ATC | T | I | |
| 384 | T | C | GTT | GTC | |||
| 416 | C | A | CGC | CAC | R | H | |
| 419 | A | T | CAC | CTC | H | L | |
| 432 | C | A | ACC | ACA | |||
| 448 | G | A | GAC | AAC | D | N | |
| 451 | G | T | GAA | TAA | E | 终止密码Stop codon | |
| 464, 465 | CA | TG | CCA | CTG | P | L | |
| 487 | T | C | TGG | CGG | W | R | |
| 515 | C | T | CCA | CTA | P | L | |
| 518 | G | A | GGG | GAG | G | E | |
| 528 | C | T | ATC | ATT | |||
| 534 | C | T | CTC | CTT | |||
| 668 | 插入序列7个碱基7 bp insertion (CTCAGGA) | 引起移码突变Frame shift mutation | |||||
| 721 | C | T | CTG | TTG | |||
| 800 | A | G | TAT | TGT | Y | C | |
| 807 | G | A | CAG | CAA | |||
| 930 | G | A | ACG | ACA | |||
| 963 | T | A | CCT | CCA | |||
| 1017 | C | T | TAC | TAT | |||
| 1039 | T | G | TTT | GTT | F | V | |
| 1047 | A | G | AAA | AAG | |||
| 1105 | A | C | AAG | CAG | K | Q | |
新窗口打开|下载CSV
Table 2
表2
表2FAD2B基因突变
Table 2
| 突变位点 Mutation site | 核酸Nucleic acid | 密码子Codon | 氨基酸Amino acid | |||
|---|---|---|---|---|---|---|
| 突变前 Before mutation | 突变后 After mutation | 突变前 Before mutation | 突变后 After mutation | 突变前 Before mutation | 突变后 After mutation | |
| 40 | A | G | AAG | GAG | K | E |
| 150 | A | C | TCA | TCC | ||
| 172 | A | G | ATG | GTG | M | V |
| 214 | A | C | AAG | CAG | K | Q |
| 282 | C | T | ACC | ACT | ||
| 333 | C | T | TAC | TAT | ||
| 362 | C | T | ACC | ATC | T | I |
| 384 | T | C | GTT | GTC | ||
| 416 | C | A | CGC | CAC | R | H |
| 419 | A | T | CAC | CTC | H | L |
| 432 | A | C | ACA | ACC | ||
| 451 | G | T | GAA | TAA | E | 终止密码Stop codon |
| 464 | C | T | CCG | CTG | P | L |
| 487 | T | C | TGG | CGG | W | R |
| 515 | C | T | CCA | CTA | P | L |
| 518 | G | A | GGG | GAG | G | E |
| 528 | T | C | ATT | ATC | ||
| 534 | T | C | CTT | CTC | ||
| 668 | 插入序列7个碱基 7 bp insertion (CTCAGGA) | 引起移码突变Frame shift mutation | ||||
| 721 | T | C | TTG | CTG | ||
| 800 | A | G | TAT | TGT | Y | C |
| 807 | G | A | CAG | CAA | ||
| 907 | G | A | GCA | ACA | A | T |
| 930 | G | A | ACG | ACA | ||
| 963 | T | A | CCT | CCA | ||
| 982 | G | T | GCA | TCA | A | S |
| 1017 | C | T | TAC | TAT | ||
| 1039 | G | T | GTT | TTT | V | F |
| 1047 | A | G | AAA | AAG | ||
| 1105 | C | A | CAG | AAG | Q | K |
新窗口打开|下载CSV
FAD2A基因共产生29个位点的突变, 16个引起蛋白质序列的变化。其中突变体12-6在第451位的碱基G突变为T, 使得原来编码谷氨酸(E)的密码子GAA变为终止密码子TAA, 使得FAD2A蛋白合成提前终止。另外在第668位插入7个碱基CTCAGGA, 使得该基因产生移码突变, 这些无义突变将产生FAD2A功能性缺失, 影响酶活性。其他单个氨基酸的突变包括已经报道的在第448位的单碱基替换(G→A), 使编码的蛋白质第150位氨基酸由天冬氨酸(D)转变为天冬酰胺(N), 而D150残基在所有FAD2中是绝对保守的, 是高油酸性状的关键位点, 突变导致AhFAD2A酶活性降低, 不能催化油酸向亚油酸转变的过程[17]。该突变在2株花生中被发现(148-1和149-3)。
FAD2B基因有30个突变。其中13个突变不造成编码蛋白质序列的变化, 17个突变引起蛋白质序列的变化(表2)。其中突变体12-2、13-5、148-2、149-6、148-10、149-4、150-12在451位的碱基由G变为T引起相应的密码子由GAA变为终止密码子TAA, 使蛋白质翻译提前终止, 产生150个氨基酸的蛋白质; 突变体12-2、12-5、13-5、148-2、148-6、148-9、148-10在第668位插入7个碱基CTCAGGA, 可引起蛋白质的移码突变。其他突变都为单个氨基酸突变, 但没有发现在448位的单碱基替换(G→A)。
3 讨论
Norden等[12]于1987年在佛罗里达育种计划中发现了第1个天然高油酸花生突变体F435, 从此打开高油酸花生育种和研究的序幕。F435种子油酸含量高达80%, 亚油酸含量仅为2%。美国大多数高油酸花生品种以F435为基础进行研究和发展。高油酸的性状由2个隐性突变基因(ol1和ol2)调控[3,12]。通过对普通油酸花生与F435序列比对发现, 在AhFAD2A的编码区从起始密码子起第448 bp碱基发现存在单碱基替换(G→A), 使编码的蛋白质第150位氨基酸由天冬氨酸(D)转变为天冬酰胺(N), 而D150残基在所有FAD2中是绝对保守的, 是高油酸性状的关键位点, 突变导致AhFAD2A酶活性降低, 不能催化油酸向亚油酸转变的过程[17]; 在AhFAD2B基因起始密码子起第441_442 bp处存在单碱基插入(442insA), 造成移码突变, 导致翻译的蛋白质序列提前终止, 产生无活性的蛋白[18,19]。随后, 通过化学诱变和物理诱变获得了一些高油酸花生材料, 如M2-225[20]、Mycogen-Flavo[21]、SPI098[22]和GAT2636M[23], 所有的这些突变都影响到了FAD2酶的活性。高油酸花生品种C458和M2-225在AhFAD2A自然突变(G448A)基础上, 诱变又使AhFAD2B基因起始密码子后第665 bp和997 bp处分别插入微型反向重复转座元件MITE (205 bp), 导致肽链提前终止, 使蛋白功能丧失。AhFAD2B的突变类型主要有2种: 441_442insA突变(F435及衍生系列、E16等)和MITE插入突变(C458、M2-225等)[24], 以上AhFAD2B突变基因均属于无义突变(nonsense mutation)[25], 使产生的蛋白不完全而丧失原基因的功能, 已成为花生高油酸育种的基因源。国际上除上述最基本F435高油酸基因材料外, 美国科学家又鉴定出2个新的高油酸花生自然突变体PI342664和PI342666, 其AhFAD2A基因编码区448位G→A突变与先前报道相同, 而AhFAD2B基因突变C301G导致氨基酸H101D改变属于新的突变位点, 最终使突变体油酸含量达到79%以上[26]。印度花生育种家Nadaf等[27]用EMS和γ射线处理GPBD-4, 获得2个高油酸系GM4-3和GM6-1, 通过克隆其AhFAD2B 基因和测序后发现了2个新的突变位点。GM6-1在第1085A→G和1111G→A位上发生碱基突变, GM4-3在1111G→A位上发生突变。1111G→A的突变使得AhFAD2B基因的第371位氨基酸由甘氨酸转变成丝氨酸, 1085A→G的突变使得AhFAD2B 基因的第362位氨基酸由丝氨酸转变为甘氨酸, 2个高油酸系油酸含量均在75%以上。国内花生研究者Fang等[28]利用EMS化学诱变获得AhFAD2B基因新的突变位点C313T, 引起氨基酸序列H105Y突变, 使油酸含量由44%提高到60%以上。庄伟建[29]等公布通过γ射线诱变处理获得一种花生高油酸隐性突变系Ahfad2a-1植株, 陈四龙[30]公布一种自然突变产生的高油酸花生突变体C814T材料, 在AhFAD2B基因第814位置包含1处有义碱基突变: 814C→T, 使得蛋白质序列272氨基酸残基由组氨酸(H)转变为酪氨酸(Y)。除上述F435为自然突变外, 其余高油酸品种均为人工诱变产生。这些高油酸花生的突变体为之后分子的遗传机理提供重要的材料, 促进了花生油酸基因的分离和性状遗传的探究。
随着生物技术不断发展和对高油酸遗传机理的研究深入, 很多生物学家尝试各种不同的方法来获得目标高油酸花生种子, 其中包括各种植物基因工程技术。徐霞[31]采用农杆菌介导法, 将反义FAD2基因导入花生, PCR检测和Southern杂交检测证实反义FAD2基因已经整合入花生基因组。Yin等[32]对获得的4份转FAD2基因的花生种子进行品质分析发现, 油酸含量由野生型的37.87%提高到59.62%~70.01%; 李桂民[33]应用双链RNA基因沉默技术使T1代转基因种子油酸含量由原来的38%提高到77%。Yuan等[34]最近报道了通过CRISPR/Cas9技术对花生FAD2基因的编辑, 并在原生质体及毛状根中获得基因突变, 但由于转基因花生原生质体及毛状根都无法再生植株, 因此没有获得可稳定遗传的花生材料。我们采用基因枪转化花生胚性愈伤组织, 获得转基因植株, 并鉴定到多个花生FAD2基因突变。基因枪转化是植物基因转化的常用方法之一, 由于其操作简单, 转化效率高, 对于一些农杆菌难浸染的植株尤其有效, 但基因枪转化也经常造成多个拷贝的转基因现象。在基因组编辑中, 多拷贝转基因有可能使得这些基因的表达提高, 从而有利于基因编辑, 增加突变效率。另一方面, 多拷贝转基因也为将来转基因成分的分离清除造成麻烦。本研究获得多个可以稳定遗传的材料, 为运用基因组编辑系统进行高油酸花生的育种研究奠定了基础。
4 结论
本研究通过CRISPR/Cas9基因组编辑技术实现了对栽培花生FAD2A和FAD2B基因的突变, 得到了可以稳定遗传的高油酸花生突变体, 为高油酸花生的培育带来了新的途径。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
PMID [本文引用: 1]

A high-oleate trait in the cultivated peanut ( Arachis hypogaea L.) was reported to rely on the allelic composition of the two homeologous, microsomal oleoyl-PC desaturase genes ( ahFAD2A or ahFAD2B). The enzyme activity of either ahFAD2A or ahFAD2B is sufficient for a normal oleate phenotype, and a significant reduction in the levels of ahFAD2B and a mutation in ahFAD2A were reported to be responsible for the high-oleate phenotype in one chemically induced mutant (M2-225) and one derived from a naturally occurring (8-2122) mutant. Here, we report an insertion of the same miniature inverted-repeat transposable element (MITE) in the ahFAD2B gene in another chemically induced mutant (Mycogen-Flavo) and the previously characterized M2-225 mutant. In both cases, this MITE insertion in ahFAD2B causes a frameshift, resulting in a putatively truncated protein sequence in both mutants. The insertion of this MITE in ahFAD2B, in addition to the point mutation in ahFAD2A, appears to be the cause of the high-oleate phenotype in Mycogen-Flavo and M2-225 mutants. Utilizing sequences of the MITE, we developed a DNA marker strategy to differentiate the two insertion-containing mutants from the normal oleate peanut variety (AT-108) and the naturally occurring, high-oleate mutant 8-2122. Reverse transcript-PCR/differential digestion results reveal the expression of both ahFAD2A and ahFAD2B genes in Mycogen-Flavo mutant. This result is in contrast to the observation that ahFAD2B transcripts are greatly reduced in the M2-225 mutant having the MITE insertion further 3' in ahFAD2B gene.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIPMID [本文引用: 1]
Increasing the content of oleic acid in peanut seeds is one of the major goals in peanut breeding due to consumer and industry benefits, such as anti-oxidation and long shelf-life. Homeologous ahFAD2A and ahFAD2B genes encode fatty acid desaturases, which are the key enzymes for converting oleic acid to linoleic acid that oxidizes readily. To date, all high oleic acid peanut varieties result from natural mutations occurred in both genes. A method to induce mutations in the genes of other elite cultivars could speed introgression of this valuable trait. The gene-editing approach utilizing CRISPR/Cas9 technology was employed to induce de novo mutations in the ahFAD2 genes using peanut protoplasts and hairy root cultures as models.The hot spot of natural mutation in these genes was selected as the target region. Appropriate sgRNAs were designed and cloned into a CRISPR/Cas9 expression plasmid. As a result of CRISPR/Cas9 activity, three mutations were identified - G448A in ahFAD2A, and 441_442insA and G451T in ahFAD2B. The G448A and 441_442insA mutations are the same as those seen in existing high oleate varieties and the G451T is new mutation. Because natural mutations appear more often in the ahFAD2A gene than in the ahFAD2B gene in subspecies A. hypogaea var. hypogaea, the mutations induced in ahFAD2B by gene editing may be useful in developing high oleate lines with many genetic backgrounds after validation of oleic acid content in the transformed lines. The appearance of the G448A mutation in ahFAD2A is a further benefit for high oleic acid oil content.Overall, these results showed that mutations were, for the first time, induced by CRISPR-based gene editing approach in peanut. This research demonstrated the potential application of gene editing for mutagenesis in peanut and suggested that CRISPR/Cas9 technology may be useful in the peanut breeding programs.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}