 ,1,2,*
,1,2,*Identification of resistance to wheat and molecular detection of resistance genes to wheat stripe rust of 78 wheat cultivars (lines) in Sichuan province
XI Ling1,2, WANG Yu-Qi1,2, ZHU Wei1,2, WANG Yi1,2, CHEN Guo-Yue1,2, PU Zong-Jun3, ZHOU Yong-Hong1,2, KANG Hou-Yang,1,2,*通讯作者:
收稿日期:2020-08-2接受日期:2020-11-13网络出版日期:2020-12-22
| 基金资助: |
Received:2020-08-2Accepted:2020-11-13Online:2020-12-22
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (849KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
习玲, 王昱琦, 朱微, 王益, 陈国跃, 蒲宗君, 周永红, 康厚扬. 78份四川小麦育成品种(系)条锈病抗性鉴定与抗条锈病基因分子检测[J]. 作物学报, 2020, 47(7): 1309-1323. doi:10.3724/SP.J.1006.2021.01061
XI Ling, WANG Yu-Qi, ZHU Wei, WANG Yi, CHEN Guo-Yue, PU Zong-Jun, ZHOU Yong-Hong, KANG Hou-Yang.
小麦条锈病是由条形柄锈菌小麦专化型(Puccinia striiformis f. sp. tritici, Pst)引起的真菌病害, 严重影响小麦的产量[1]。条锈菌主要随风远距离传播, 引起大规模的流行发生, 从而造成巨大的产量和经济损失。由于病原菌的快速进化和新毒力小种的出现, 常常导致小麦生产上广泛应用的品种丧失抗条锈性[2]。迄止, 中国小麦主产区已发生过多次条锈病大流行, 1950年、1964年、1990年、2002年和2017年的条锈病流行分别导致小麦产量损失60亿、32亿、18亿、13亿和15亿千克, 严重影响小麦生产安全[3]。
研究发现, 导致小麦品种丧失抗条锈性的新毒性小种主要来源于甘肃南部和四川西北部等地区[4,5]。四川省是中国重要的小麦主产区, 也是条锈菌冬繁区, 四川盆地海拔300~700 m, 属于亚热带季风性湿润气候, 冬季气候温和, 平均气温6~8℃, 为条锈菌的繁殖提供了适宜环境, 病原菌侵染秋苗并越冬繁殖, 翌春, 病原菌随着高空气流传播至中、东部的黄淮海麦区以及长江中下游麦区, 导致条锈病大规模发生[6,7,8]。2009年首次在四川省发现以条中34 (CYR34)为代表的贵农22致病类群(对Yr26致病, Yr26=Yr24), 几年内扩展至全国小麦主产区。CYR34对小麦生产上应用较广的重要抗条锈病基因Yr10、Yr24 (=Yr26)、YrCH42产生毒性, 成为当前的主流小种, 导致抗条锈病的川麦42、绵麦37、内麦8号等主推品种丧失抗性[9,10], 而川麦42是由四川省农业科学院利用国际玉米小麦改良中心(CIMMYT)引进的一批硬粒小麦—节节麦人工合成种与四川生产上推广的川麦30杂交、回交培育而成的, 在CYR34出现之前具有高产、抗病等优良性状[11,12]。
到目前为止, 四川省与CIMMYT进行种质资源交换已有30多年历史, CIMMYT种质已成为四川小麦育种的重要亲本资源[13]。在四川麦区, 通过与CIMMYT选育得到了突破性品种繁6及衍生系绵阳11, 以及后来具有CIMMYT小麦种质血缘的川麦18、川麦30、川麦32、川麦36、川麦39和人工合成种后代川麦42、川麦43, 这些品种在条锈病新抗源, 高产强筋上都具有较大突破[14]。CIMMYT小麦种质的引入极大地促进了四川小麦品种改良, 特别在其抗病性和广适性等方面[15]。四川省的小麦条锈病新毒性小种变异快, 利用分子标记辅助选择(marker-assisted selection, MAS)将CIMMYT的外源优异抗性基因导入本地品种中, 以培育持久抗性新品种[13]。
实践证明, 不断培育和利用抗病品种是防治条锈病最具生态效益和经济安全的方法。目前已正式命名的抗条锈病基因有83 (Yr1~Yr83)个, 但是许多小种特异性抗病基因已被毒性小种所克服, 若长期大面积单一使用某一抗性基因(如Yr10、Yr24/Yr26、YrCH42), 导致病原菌与抗源协同进化使条锈菌产生变异, 则会出现条锈病大面积流行[16], 因此需要不断挖掘小麦抗条锈病基因或QTL (quantitative trait locus), 加强四川小麦与CIMMYT的种质交流。Ye等[17]在244份四川小麦地方品种和小麦育成品种中鉴定出52份在6种环境中表现稳定抗条锈病的材料, 并定位到两个新的抗性QTL (Qyrsicau-5AS和Qyrsicau-5AL)。周露[18]、王健维[19]、刘涛[20]分别在四川小麦地方品种古宋大龙须、仪陇坨麦和合江移栽麦鉴定出显性抗条锈病基因, 分别命名为Yrgsdlx、Yryltm和YrHY。王相权等[21]利用CYR32、CYR33和CYR34的混合菌从1949—2016年四川省审(认)定的170份冬小麦品种(系), 筛选出川农30、蜀麦126、川麦55等43份品种(系)抗条锈病, 这些材料可在小麦生产上继续使用或作为亲本资源。曹世勤等[22]利用CYR32、CYR33、CYR34及G22-14、中4-1和混合菌对331份四川小麦品种(系)在甘肃陇南的抗条锈病鉴定, 表明成株期表现抗性的有72份, 仅11份材料对条锈菌具有全生育期抗性(all stage resistance, ASR)。以上研究结果仅对四川小麦品种(系)的条锈病抗性进行了鉴定和评价, 但是未涉及抗条锈病基因的系统研究。因此, 本研究对2016年以来四川省78份小麦育成品种(系)进行苗期和成株期抗条锈病鉴定, 抗条锈病基因的分子检测, 明确这些小麦品种(系)的条锈病抗性水平和抗条锈病基因的利用, 为抗病品种的合理布局防控小麦条锈病提供理论依据。
1 材料与方法
1.1 供试材料
78份四川小麦育成品种(系), 由四川省农业科学院作物所蒲宗君博士、绵阳市农业科学院任勇博士、西南科技大学杨随庄博士、四川农业大学谭飞泉博士、代寿芬博士提供。铭贤169用作苗期条锈病感病对照和诱发条锈病小麦品种, SY95-71和台长29用作成株期诱发条锈病小麦材料, 15个已知抗条锈病基因载体材料: Triticum. spelta album (Yr5)、Yr10/6*Avocet S、Yr15/6*Avocet S、Yr17/6*Avocet S、Yr18/6*Avocet S、Yr26/6*Avocet S、蜀麦1675 (Yr28)、川麦55 (Yr29)、Opata 85 (Yr30)、RSL65 (Yr36)、PI 566596 (Yr39)、川农19 (Yr41)、PI 610750 (Yr48)、PI 480016 (Yr65)和C591 (Yr67), 感病材料Avocet S作为阴性对照。对照材料均由四川农业大学小麦研究所提供。苗期鉴定在甘肃省农业科学院温室进行, 采用当前流行小种CYR32 (Avirulence/Virulence: Yr5、Yr10、Yr15、Yr19、Yr24、Yr26、Yr32、YrTr1/Yr1、Yr2、Yr3、Yr4、Yr6、Yr7、Yr8、Yr9、Yr11、Yr12、Yr13、Yr14、Yr16、Yr17、Yr18、Yr25、Yr27、Yr28、Yr29、Yr30、Yr31、Yr32、Yr43、Yr44、YrExp2、YrA、YrSk、YrSp)[23,24,25]和CYR34 (Avirulence/Virulence: Yr5、Yr15、Yr32、YrTr1/Yr1、Yr2、Yr3、Yr4、Yr6、Yr7、Yr8、Yr9、Yr10、Yr11、Yr12、Yr13、Yr14、Yr16、Yr17、Yr18、Yr19、Yr25、Yr24、Yr26、Yr27、Yr28、Yr29、Yr30、Yr31、Yr32、Yr43、Yr44、YrExp2、YrA、YrSk、YrSp)[24,25,26]。成株期抗条锈病鉴定在四川温江基地, 通过人工接种CYR32、CYR33、CYR34及贵22-14、水源类型、贵农致病类群的等量混合菌, 供试菌种均由甘肃省农业科学院植物保护研究所贾秋珍研究员提供。1.2 试验方法
1.2.1 苗期抗病性鉴定 在甘肃省农业科学院植物保护研究所温室对78份供试材料进行苗期抗性鉴定。感病对照为铭贤169, 所有材料分别选取10~15粒种子播种, 待长到一叶一心期时用于接种。小种 CYR32和CYR34各25 mg新鲜夏孢子加在1 mL的吐温水溶液中制备条锈菌悬浮液, 用喷雾装置将孢子悬浮液分小种均匀喷洒在植株叶片上, 静置30 min晾干, 将接种植株置于湿度>95%的10℃保湿间黑暗保湿24 h, 后置于光照培养箱内培养, 接种后18~20 d, 待铭贤169充分感病时, 调查反应型(infection type, IT), 反应型按照0~9级[27]调查标准进行记录, 高抗(ITs 0~3)、中抗(ITs 4~6)、高感(ITs 7~9)[28]。1.2.2 成株期人工诱发抗病性鉴定 78份四川小麦育成品种(系)于2018—2019年在四川农业大学温江基地进行成株期抗条锈病鉴定。采用随机区组设计, 行长1.5 m、行距0.3 m, 每个材料单粒播种, 每行定植15株, 垂直于供试品种种植行一端种植1行感病对照SY95-71作为条锈病诱发行。在小麦分蘖期时用等量的混合菌与滑石粉1︰50 (m/v)的比例混合均匀, 采用涂抹法[29]进行人工接种, 待SY95-71严重度达80%以上时, 对供试材料进行成株期抗病性调查, 每行随机选取单株6株, 调查其病害严重度(disease severity, DS)和普遍率, 每隔7 d调查一次, 共3次, 最后一次调查反应型, 计算其平均病害严重度(mean disease severity, MDS)各平均普遍率。反应型鉴定标准与苗期的一致, 严重度采用8个等级(0、5%、10%、20%、40%、60%、80%和100%)[27]。
1.2.3 抗条锈病基因的分子检测 取小麦叶片组织, 采用Hill-Ambroz等[30]改良的CTAB提取方法提取供试材料基因组DNA, 使用分光光度计(NanoDropone, 赛默飞世尔科技(中国)有限公司, 美国)测定DNA浓度, 并用1×TE溶液将DNA稀释至100 ng μL-1, 于-20℃冰箱中储存备用。选用与QYr.nwafu-4BL、Yr5、Yr10、Yr15、Yr17、Yr18、Yr26、Yr28、Yr29、Yr30、Yr36、Yr39、Yr41、Yr48、Yr65、Yr67、Yr78、Yr80和Yr81[31]紧密连锁的Simple Sequence Repeats (SSR)、Cleavage Amplification Polymorphism Sequence (CAPS)、Sequence Tagged Sites (STS)和Kompetitive Allele-specific PCR (KASP)等标记或功能标记, 对供试材料基因组DNA进行PCR扩增, 扩增程序使用降落式PCR (Touchdown PCR), 扩增产物在1.5%~2.0% (m/v)的琼脂糖凝胶电泳(视PCR扩增产物大小而定)、毛细管电泳、荧光定量PCR中扩增检测。分子标记的所有引物序列均由上海生工生物工程有限公司合成(表1)。
Table 1
表1
表1抗条锈病基因的分子标记及引物序列
Table 1
| Yr基因 Yr gene | 类别a Type a | 分子标记 Molecular marker | 引物序列 Primer sequence (5′?3′) | 参考文献 Reference |
|---|---|---|---|---|
| QYr.nwafu-4BL | KASP | AX-110963704 | GAAGGTGACCAAGTTCATGCTTCGTTTGCCTATGGTTCAAAAC | [31] |
| GAAGGTCGGAGTCAACGGATTTCGTTTGCCTATGGTTCAAAAG | ||||
| TCTCCATGACACGGGAGGAT | ||||
| Yr5 | DM | Yr5_B | GGGAACACTTCACGATCA | [32] |
| AATTCCTTCATGCCTTCC | ||||
| DM | Yr5_A | CGCTTAATTCCCCTTCCTTC | [32] | |
| TGGCTCCTTATTCGTTCTCTTTC | ||||
| KASP | Yr5F | GAAGGTGACCAAGTTCATGCTGCGCCCCTTTTCGAAAAAATA | [32] | |
| Yr5H | GAAGGTCGGAGTCAACGGATTCTAGCATCAAACAAGCTAAATA | |||
| Yr5R | ATGTCGAAATATTGCATAACATGG | |||
| Yr10 | AFLP | SC200 | CTGCAGAGTGACATCATACA | [33] |
| TCGAACTAGTAGATGCTGGC | ||||
| DM | Yr10-5 | GGAAATGTGGCGGAGTACCA | [34] | |
| CGGAAGGGAGAACCACTGTC | ||||
| DM | Yr10-6 | CAGCTTGACAAGGGCGAGTA | [34] | |
| CGCTTGTCGCCAATTCCAAA | ||||
| Yr15 | DM | Y15K1_F2 | GGAGATAGAGCACATTACAGAC | [35] |
| uhw301R | GGAGATAGAGCACATTACAGAC | |||
| DM | WJL2F | CCTTGTGTGCTACCAGGGTT | [36] | |
| WJL2R | GGAACTCAAGCCCTTCTGCT | |||
| DM | WJL3F | AAAAGAGCTCGCCTCCTACG | [37] | |
| WJL3R | GCCATGATGAGATCGGGAGG | |||
| Yr17 | SCAR | SC2372 | AGGGGCTACTGACCAAGGCT | [38] |
| TGCAGCTACAGCAGTATGTACACAAAA | ||||
| Yr18 | STS | csLV34 | GTTGGTTAAGACTGGTGATGG | [39] |
| TGCTTGCTATTGCTGAATAGT | ||||
| Cssfr1 | L34DINT9F | TTGATGAAACCAGTTTTTTTTCTA | [39] | |
| L34PLUSR | GCCATTTAACATAATCATGATGGA | |||
| KASP | Lr34-KASP-E11 | GAAGGTGACCAAGTTCATGCTGGGAGCATTATTTTTTTCCATCA | [40] | |
| GAAGGTCGGAGTCAACGGATTGGGAGCATTATTTTTTTCCATCT | ||||
| AGCGAATCCAGTATGGAAAT | ||||
| Yr26 | STS | Xwe173 | GGGACAAGGGGAGTTGAAGC | [41] |
| GAGAGTTCCAAGCAGAACAC | ||||
| STS | Xwe33 | TAAACCAAGTCCCCCAAA | [41] | |
| GGAGTCCATCTTCACCGA | ||||
| Yr28 | InDel | ZCZp343F | TGTGTCATGTTTGGTCGATAGG | [42] |
| ZCZp342R | TCCTCCCTTGTAGCTTCACG |
新窗口打开|下载CSV
(续表1)
(续表1)
| Yr基因 Yr gene | 类别a Type a | 分子标记 Molecular marker | 引物序列 Primer sequence (5′?3′) | 参考文献 Reference |
|---|---|---|---|---|
| Yr28 | DM | P175 | GCACCGTCCTTCATCTCAGT | [42] |
| P176 | TGCTTTTCCCCGTATCCCTT | |||
| Yr29 | CAPS | csLV46 | CGAGACGTCGTCTTCTCTAAC | [43] |
| GTGTATGTGTTGATTCTCCTCG | ||||
| Yr30 | SSR | Xgwm533 | GTTGCTTTAGGGGAAAAGCC | [44] |
| AAGGCGAATCAAACGGAATA | ||||
| Yr36 | DM | WKS1_150F | ATGGAGCTCCCACGAAACAAAC | [45] |
| WKS1_620R | ACCTCCATGTTGCTCGCATTTGCT | |||
| DM | WKS1_182F | TAGCTCTTCGTGGCCAAAGG | [45] | |
| WKS1_150R | TGGCCACGAAGAGCTAAAGG | |||
| Yr39 | SSR | Xgwm131 | AATCCCCACCGATTCTTCTC | [46] |
| AGTTCGTGGGTCTCTGATGG | ||||
| Yr41 | SSR | Xgwm410 | GCTTGAGACCGGCACAGT | [47] |
| CGAGACCTTGAGGGTCTAGA | ||||
| SSR | Xgwm374 | ATAGTGTGTTGCATGCTGTGTG | [47] | |
| TCTAATTAGCGTTGGCTGCC | ||||
| Yr48 | SSR | Xwmc727 | CATAATCAGGACAGCCGCAC | [48] |
| TAGTGGCCTGATGTATCTAGTTGG | ||||
| SSR | Xwms291 | CATCCCTACGCCACTCTGC | [48] | |
| AATGGTATCTATTCCGACCCG | ||||
| Yr65 | SSR | Xgwm18 | GGTTGCTGAAGAACCTTATTTAGG | [49] |
| TGGCGCCATGATTGCATTATCTTC | ||||
| Yr67 | SSR | Xbarc182 | CCATGGCCAACAGCTCAAGGTCTC | [50] |
| CGCAAAACCGCATCAGGGAAGCACCAAT | ||||
| SSR | Xcfa2040 | TCAAATGATTTCAGGTAACCACTA | [50] | |
| TTCCTGATCCCACCAAACAT | ||||
| Yr78 | KASP | IWA7257 | GAAGGTGACCAAGTTCATGCTAGACCCTACGACGTTAGCGC | [51] |
| GAAGGTCGGAGTCAACGGATTAGACCCTACGACGTTAGCGA | ||||
| ATTGGAATCAGCTGGGTCAT | ||||
| Yr80 | KASP | KASP_53113 | GAAGGTGACCAAGTTCATGCTTGTACAATGACTCCTCGACTAACA | [52] |
| GAAGGTCGGAGTCAACGGATTTGTACAATGACTCCTCGACTAACG | ||||
| GCCACGCAATATCACCATCG | ||||
| Yr81 | KASP | KASP_3077 | GAAGGTGACCAAGTTCATGCTATTCCAAAGTAATTGGCAACAGGTTCA | [53] |
| GAAGGTCGGAGTCAACGGATTCCAAAGTAATTGGCAACAGGTTCG | ||||
| TGTGGAGCGTGACAATGAGGAAGTT |
新窗口打开|下载CSV
2 结果与分析
2.1 苗期抗条锈性
条锈菌小种CYR32和CYR34对78份四川小麦育成品种(系)的苗期抗病性鉴定结果表明, 绵麦161、西科麦12、川农30等60份品种(系)对CYR32表现出苗期抗性(ITs 0-6), 占76.92%。绵麦827、蜀 麦1783、川麦608等40份品种(系)对CYR34表现出苗期抗性, 占51.28% (表2)。绵麦902、蜀麦1743、川麦611等36份品种(系)对2个小种均表现苗期抗病性, 占46.15%, 其中蜀麦1613、蜀麦1829、蜀麦1868、川麦605和川麦1747对CYR32和CYR34都表现为高抗(ITs 0-3) (表2)。对比2个小种抗病情况发现, 对CYR32表现感病的有18份, 对CYR34表现感病的有38份。其中凡37123、西科麦12和川麦613等24份种质对CYR32表现抗病, 但对CYR34感病; 而西科麦475、川农32、蜀麦1843和川麦1694对CYR32感病, 但对CYR34抗病, 说明在新小种CYR34出现后对一部分抗源已产生毒性, 以后应加强锈病监测。综合而言, 四川小麦育成品种(系)在苗期对当前2个流行的条锈菌生理小种具有较好的抗病性, 抗性水平整体偏高。图1
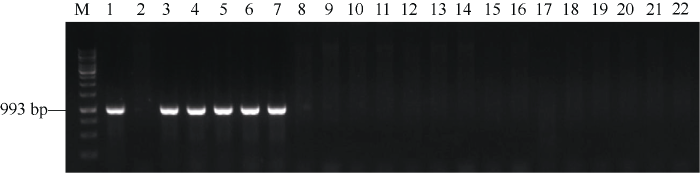 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1利用分子标记Y15K1_F2/uhw301R检测部分四川小麦育成品种(系)中Yr15基因
M: Molecular weight marker (1 kb); 1: Yr15/6*AvS; 2: Avocet S; 3: 绵麦835; 4: 西科麦557; 5: 蜀麦1829; 6: 蜀麦1868; 7: 川麦1747; 8: 凡37123; 9: 31966; 10: 绵麦161; 11: 绵麦827; 12: 绵麦906; 13: 西科麦12; 14: 川农27; 15: 川农41; 16: 蜀麦1613; 17: 蜀麦1821; 18: 蜀麦1871; 19: 川麦607; 20: 川麦611; 21: 川麦802; 22: 川麦803。
Fig. 1Agarose gel result of Yr15 gene by molecular marker Y15K1_F2/uhw301R in the part of Sichuan wheat cultivars (lines)
M: molecular weight marker (1 kb); 1: Yr15/6*AvS; 2: Avocet S; 3: Mianmai 835; 4: Xikemai 557; 5: Shumai 1829; 6: Shumai 1868; 7: Chuanmai 1747; 8: Fan 37123; 9: 31966; 10: Mianmai 161; 11: Mianmai 827; 12: Mianmai 906; 13: Xikemai 12; 14: Chuannong 27; 15: Chuannong 41; 16: Shumai 1613; 17: Shumai 1821; 18: Shumai 1871; 19: Chuanmai 607; 20: Chuanmai 611; 21: Chuanmai 802; 22: Chuanmai 803.
2.2 成株期抗条锈性
根据2019年温江(2019WJ)的田间条锈病鉴定结果, 78份四川小麦育成品种(系)在成株期对当前供试的条锈菌均表现抗病, 其中高抗条锈病(ITs 0-3, MDS<20%)的有76份, 占97.44%, 绵麦835、蜀麦1743、蜀麦1829和蜀麦1868在成株期对供试条锈菌表现为免疫水平(IT 0、MDS=0), 仅蜀麦1820和川麦1546表现为中抗条锈病(IT 4)(表2)。2.3 抗条锈病评价
根据78份四川小麦育成品种(系)的苗期和成株期抗病性鉴定结果, 利用成株期抗性(adult plant resistance, APR)和全生育期抗性(all stage resistance, ASR)确定供试材料的抗病性类型。所有供试材料中有36份表现为全生育期抗性, 占46.15%; 42份表现为成株期抗性, 占53,85%; 苗期和成株期均高抗(ITs 0-3)条锈病的有蜀麦1613、蜀麦1829、蜀麦1868、川麦605和川麦1747 (表2)。结果表明78份四川小麦品种(系)对条锈病具有较好的抗病性, 可以继续投入小麦生产使用中, 也可作为小麦育种的亲本资源。Table 2
表2
表278份四川小麦育成品种(系)抗条锈病评价及分子检测结果
Table 2
| 序号 Number | 品种 Variety | 苗期反应型 Infection type in seedling stage | 成株期2019WJ a 2019WJ a in adult-plant stage | 抗性类型b Resistance type b | Yr基因 Yr gene | ||
|---|---|---|---|---|---|---|---|
| CYR32 | CYR34 | IT | DS | ||||
| 1 | 凡37123 Fan 37123 | 5 | 7 | 1 | 0 | APR | Yr17+Yr81 |
| 2 | 31966 | 1 | 7 | 1 | 0 | APR | Yr17+Yr81 |
| 3 | 绵麦161 Mianmai 161 | 5 | 5 | 1 | 0 | ASR | Yr17+Yr26 |
| 4 | 绵麦319 Mianmai 319 | 5 | 5 | 2 | 5 | ASR | QYr.nwafu-4BL+Yr17+Yr26+Yr65 |
| 5 | 绵麦827 Mianmai 827 | 5 | 5 | 2 | 5 | ASR | Yr26+Yr30+Yr65+Yr78+Yr81 |
| 6 | 绵麦835 Mianmai 835 | 4 | 5 | 0 | 0 | ASR | Yr15 |
| 7 | 绵麦902 Mianmai 902 | 6 | 6 | 3 | 5 | ASR | Yr17+Yr78+Yr81 |
| 8 | 绵麦903 Mianmai 903 | 5 | 5 | 2 | 5 | ASR | Yr17+Yr29+Yr30+Yr78+Yr80 |
| 9 | 绵麦905 Mianmai 905 | 6 | 0 | 1 | 0 | ASR | Yr17+Yr28 |
| 10 | 绵麦906 Mianmai 906 | 2 | 6 | 1 | 0 | ASR | QYr.nwafu-4BL+Yr17+Yr26+Yr39+Yr80 |
| 11 | 绵麦1419 Mianmai 1419 | 5 | 7 | 1 | 0 | APR | Yr26+Yr30+Yr67+Yr78 |
| 12 | 绵麦1501 Mianmai 1501 | 5 | 7 | 3 | 10 | APR | Yr17+Yr26+Yr29+Yr78+Yr81 |
| 13 | 绵麦5658 Mianmai 5658 | 1 | 7 | 1 | 0 | APR | Yr17+Yr26+Yr30+Yr78+Yr80 |
| 14 | 绵麦5695 Mianmai 5695 | 1 | 4 | 2 | 5 | ASR | Yr26+Yr78+Yr81 |
| 15 | 西科麦11 Xikemai 11 | 5 | 7 | 1 | 0 | APR | Yr17+Yr26+Yr81 |
| 16 | 西科麦12 Xikemai 12 | 5 | 7 | 1 | 0 | APR | Yr29+Yr30 |
| 17 | 西科麦18 Xikemai 18 | 5 | 7 | 1 | 0 | APR | — |
| 18 | 西科麦475 Xikemai 475 | 7 | 5 | 2 | 5 | APR | Yr17+Yr26+Yr29+Yr30+Yr81 |
| 19 | 西科麦497 Xikemai 497 | 5 | 4 | 2 | 5 | ASR | Yr29+Yr78 |
| 20 | 西科麦518 Xikemai 518 | 7 | 7 | 1 | 0 | APR | Yr17+Yr29+Yr67 |
| 21 | 西科麦546 Xikemai 546 | 7 | 7 | 2 | 5 | APR | Yr17+Yr29+Yr78 |
| 22 | 西科麦557 Xikemai 557 | 6 | 0 | 2 | 5 | ASR | Yr15+Yr29 |
| 23 | 川农27 Chuannong 27 | 7 | 7 | 2 | 5 | APR | Yr17+Yr39+Yr81 |
| 24 | 川农30 Chuannong 30 | 5 | 5 | 2 | 5 | ASR | Yr17+Yr80 |
| 25 | 川农32 Chuannong 32 | 7 | 1 | 2 | 5 | APR | Yr17+Yr39+Yr80+Yr81 |
| 26 | 川农38 Chuannong 38 | 5 | 7 | 1 | 0 | APR | Yr17+Yr81 |
| 27 | 川农39 Chuannong 39 | 5 | 7 | 1 | 0 | APR | Yr17+Yr30+Yr67 |
| 28 | 川农41 Chuannong 41 | 5 | 7 | 1 | 0 | APR | Yr17+Yr81 |
| 29 | 川农42 Chuannong 42 | 7 | 7 | 1 | 0 | APR | Yr17+Yr81 |
| 30 | 蜀麦691 Shumai 691 | 8 | 7 | 2 | 5 | APR | Yr17+Yr29+Yr30+Yr67+Yr81 |
| 31 | 蜀麦1613 Shumai 1613 | 3 | 0 | 3 | 5 | ASR | Yr17+Yr29+Yr67 |
| 32 | 蜀麦1671 Shumai 1671 | 5 | 7 | 2 | 5 | APR | Yr26+Yr30+Yr65+Yr78 |
| 33 | 蜀麦1675 Shumai 1675 | 5 | 7 | 1 | 0 | APR | Yr28+Yr29+Yr30+Yr81 |
| 34 | 蜀麦1743 Shumai 1743 | 5 | 5 | 0 | 0 | ASR | Yr17+Yr29 |
| 35 | 蜀麦1783 Shumai 1783 | 5 | 1 | 3 | 5 | ASR | Yr17+Yr29+Yr67 |
| 36 | 蜀麦1812 Shumai 1812 | 7 | 7 | 2 | 5 | APR | Yr17+Yr81 |
| 37 | 蜀麦1820 Shumai 1820 | 7 | 8 | 4 | 10 | APR | Yr17+Yr29+Yr78 |
| 38 | 蜀麦1821 Shumai 1821 | 7 | 7 | 1 | 0 | APR | Yr26+Yr30+Yr78 |
| 39 | 蜀麦1829 Shumai 1829 | 0 | 1 | 0 | 0 | ASR | Yr15+Yr17 |
| 40 | 蜀麦1843 Shumai 1843 | 7 | 6 | 1 | 0 | APR | QYr.nwafu-4BL+Yr17+Yr29 |
| 41 | 蜀麦1862 Shumai 1862 | 7 | 7 | 1 | 0 | APR | Yr26+Yr30+Yr65+Yr78 |
| 42 | 蜀麦1868 Shumai 1868 | 2 | 0 | 0 | 0 | ASR | Yr15+Yr17+Yr30+Yr41+Yr78 |
| 43 | 蜀麦1871 Shumai 1871 | 5 | 5 | 1 | 0 | ASR | Yr17+Yr29+Yr30+Yr67+Yr81 |
| 44 | 蜀麦1885 Shumai 1885 | 6 | 5 | 3 | 10 | ASR | Yr17+Yr29+Yr67 |
| 45 | 川麦83 Chuanmai 83 | 5 | 0 | 2 | 5 | ASR | Yr17+Yr29 |
| 46 | 川麦84 Chuanmai 84 | 5 | 7 | 2 | 5 | APR | QYr.nwafu-4BL+Yr30+Yr65+Yr78 |
| 47 | 川麦86 Chuanmai 86 | 5 | 7 | 1 | 0 | APR | Yr26+Yr29+Yr30+Yr78 |
| 48 | 川麦87 Chuanmai 87 | 7 | 7 | 2 | 5 | APR | Yr17+Yr30+Yr80+Yr81 |
| 49 | 川麦88 Chuanmai 88 | 7 | 7 | 2 | 5 | APR | Yr30 |
| 50 | 川麦93 Chuanmai 93 | 6 | 7 | 3 | 5 | APR | Yr26+Yr28+Yr29+Yr30+Yr65+Yr81 |
| 51 | 川麦98 Chuanmai 98 | 5 | 5 | 1 | 0 | ASR | Yr17+Yr28+Yr29+Yr81 |
| 52 | 川麦603 Chuanmai 603 | 4 | 7 | 1 | 0 | APR | Yr26+Yr29+Yr78 |
| 53 | 川麦604 Chuanmai 604 | 5 | 7 | 1 | 0 | APR | Yr17+Yr26+Yr30+Yr65 |
| 54 | 川麦605 Chuanmai 605 | 0 | 1 | 2 | 5 | ASR | QYr.nwafu-4BL+Yr26+Yr29+Yr30+Yr65 |
| 55 | 川麦606 Chuanmai 606 | 5 | 7 | 1 | 0 | APR | Yr30+Yr78 |
| 56 | 川麦607 Chuanmai 607 | 7 | 7 | 1 | 0 | APR | Yr17+Yr26+Yr78 |
| 57 | 川麦608 Chuanmai 608 | 5 | 5 | 1 | 0 | ASR | Yr17+Yr26+Yr29+Yr65+Yr78 |
| 58 | 川麦609 Chuanmai 609 | 5 | 7 | 1 | 0 | APR | Yr17+Yr26+Yr29+Yr65+Yr78 |
| 59 | 川麦610 Chuanmai 610 | 1 | 4 | 1 | 0 | ASR | Yr30+Yr65 |
| 60 | 川麦611 Chuanmai 611 | 5 | 0 | 1 | 0 | ASR | Yr29 |
| 61 | 川麦613 Chuanmai 613 | 2 | 7 | 1 | 5 | APR | Yr17+Yr29+Yr81 |
| 62 | 川麦801 Chuanmai 801 | 5 | 7 | 1 | 0 | APR | Yr17+Yr18+Yr26+Yr30+Yr65+Yr81 |
| 63 | 川麦802 Chuanmai 802 | 5 | 5 | 1 | 0 | ASR | Yr17+Yr30+Yr81 |
| 64 | 川麦803 Chuanmai 803 | 5 | 7 | 1 | 5 | APR | Yr17+Yr26+Yr30+Yr65+Yr78 |
| 65 | 川麦901 Chuanmai 901 | 7 | 7 | 2 | 5 | APR | Yr29+Yr65 |
| 66 | 川麦905 Chuanmai 905 | 4 | 5 | 3 | 5 | ASR | Yr26+Yr30+Yr65 |
| 67 | 川麦906 Chuanmai 906 | 5 | 5 | 2 | 5 | ASR | Yr26+Yr28+Yr29+Yr30+Yr65+Yr81 |
| 68 | 川麦907 Chuanmai 907 | 5 | 5 | 1 | 0 | ASR | Yr30+Yr65 |
| 69 | 川麦1456 Chuanmai 1456 | 6 | 5 | 2 | 5 | ASR | Yr18+Yr26+Yr30+Yr41+Yr78 |
| 70 | 川麦1546 Chuanmai 1546 | 5 | 5 | 4 | 10 | ASR | Yr30+Yr65 |
| 71 | 川麦1557 Chuanmai 1557 | 7 | 7 | 2 | 5 | APR | Yr30+Yr65 |
| 72 | 川麦1580 Chuanmai 1580 | 5 | 5 | 1 | 0 | ASR | Yr30+Yr65 |
| 73 | 川麦1603 Chuanmai 1603 | 5 | 5 | 1 | 0 | ASR | Yr30+Yr65 |
| 74 | 川麦1648 Chuanmai 1648 | 5 | 5 | 1 | 0 | ASR | Yr17+Yr26+Yr30 |
| 75 | 川麦1650 Chuanmai 1650 | 5 | 7 | 1 | 0 | APR | Yr17+Yr26+Yr30 |
| 76 | 川麦1694 Chuanmai 1694 | 7 | 5 | 1 | 0 | APR | Yr26+Yr30 |
| 77 | 川麦1699 Chuanmai 1699 | 5 | 5 | 2 | 5 | ASR | Yr26+Yr30+Yr65 |
| 78 | 川麦1747 Chuanmai 1747 | 0 | 0 | 1 | 0 | ASR | Yr15+Yr29+Yr30 |
新窗口打开|下载CSV
2.4 抗条锈病基因分析
根据19个抗条锈病基因QYr.nwafu-4BL、Yr5、Yr10、Yr15、Yr17、Yr18、Yr26、Yr28、Yr29、Yr30、Yr36、Yr39、Yr41、Yr48、Yr65、Yr67、Yr78、Yr80和Yr81的分子鉴定结果(表2), 绵麦319、绵麦906和蜀麦1843等5份材料可能携带QYr.nwafu-4BL; 西科麦557、蜀麦1829、绵麦835等5份材料可能携带Yr15 (图1), 占6.41%; 凡37123、31966和西科麦475等45份材料可能携带Yr17; 与Yr18共分离的基因特异性标记Cssfr1、STS标记csLV34以及KASP标记Lr34-KASP-E11的检测表明, 仅川麦801和川麦1456 可能携带Yr18, 占2.56%; 与Yr26紧密连锁的STS标记Xwe173和Xwe33检测出绵麦1419、川麦607、川麦1699等30份材料可能携带该基因; 绵麦905、蜀麦1675、川麦93等5份材料可能携带Yr28, 占6.41%; 利用CAPS标记csLV46检测结果, 发现西科麦497、蜀麦1743、川麦83等30份材料可能携带Yr29, 占38.46%; 西科麦12、蜀麦1821、川麦1650等39份材料可能携带Yr30 (图2), 占50%; 绵麦906、川农27和川农32等3份材料可能含有Yr39, 占3.85%; 利用Yr41的紧密连锁标记Xgwm410和Xgwm374, 均扩增出目标条带的有蜀麦1868和川麦1456, 占2.56%; 可能携带Yr65的包括绵麦827、川麦605、川麦901等22份材料, 占28.21%; 川农39、西科麦518、蜀麦1885等8份材料可能携带Yr67, 占10.26%; 与Yr78连锁的KASP标记IWA7257检测出绵麦1501、西科麦546和川麦607等23份材料可能携带该基因; 根据KASP标记KASP_53113检测结果显示, 绵麦903、川农30、川麦87等6份材料可能携带Yr80 (图3), 占7.69%; 与Yr81连锁的KASP标记KASP_3077检测出凡37123、31966、西科麦11等24份材料可能含有该基因, 占30.77%。所有供试材料均未检测到Yr5、Yr10、Yr36和Yr48。仅西科麦18未检测到以上19个Yr基因, 推测其可能携带未检测到的其他已知或新的抗条锈基因。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2利用分子标记Xgwm533分子检测部分四川小麦育成品种(系)中Yr30 基因
M: Molecular olecular weight marker (35~5000 bp); 1: Opata85(Yr30); 2: Avocet S; 3: 绵麦827; 4: 绵麦1419; 5: 西科麦12; 6: 西科麦475; 7: 川农39; 8: 蜀麦1671; 9: 蜀麦1675; 10: 蜀麦1871; 11: 川麦86; 12: 川麦87; 13: 川麦802; 14: 川麦905; 15: 川麦906。
Fig. 2Molecular detection of Yr30 using marker Xgwm533 in the part of Sichuan wheat cultivars (lines)
M: molecular weight marker (35-5000 bp); 1: Opata 85 (Yr30); 2: Avocet S; 3: Mianmai 827; 4: Mianmai 1419; 5: Xikemai 12; 6: Xikemai 475; 7: Chuannong 39; 8: Shumai 1671; 9: Shumai 1675; 10: Shumai 1871; 11: Chuanmai 86; 12: Chuanmai 87; 13: Chuanmai 802; 14: Chuanmai 905; 15: Chuanmai 906.
2.5 Yr基因聚合的分子检测
由表2可知, 四川小麦育成品种(系)中含多个Yr基因的材料共74份, 占供试材料的94.87%, 同时携带2~6个Yr基因的材料分别为24、22、11、14和3份。携带2个Yr基因的西科麦12 (Yr29+Yr30)、川农41 (Yr17+Yr81)和川麦606 (Yr30+Yr78)等11份材料表现成株期抗性; 携带3个Yr基因的22份材料中, 西科麦518 (Yr17+Yr29+Yr67)、蜀麦1821 (Yr26+Yr30+Yr78)和蜀麦1843 (QYr.nwafu-4BL+ Yr17+Yr29)等12份材料表现出成株期抗性; 携带4个Yr基因的材料中仅绵麦319 (QYr.nwafu-4BL+ Yr17+Yr26+Yr65)和川麦98 (Yr17+Yr28+Yr29+Yr81)表现全生育期抗性, 其余均为成株期抗性材料; 携带5个Yr基因的绵麦827 (Yr26+Yr30+Yr65+Yr78 +Yr81)、蜀麦1868 (Yr15+Yr17+Yr30+Yr41+Yr78)和川麦1456 (Yr18+Yr26+Yr30+Yr41+Yr78)等8份材料表现全生育期抗性。携带6个Yr基因的川麦93 (Yr26+Yr28+Yr29+Yr30+Yr65+Yr81)和川麦801 (Yr17+Yr18+Yr26+Yr30+Yr65+Yr81)为成株期抗性材料, 川麦906 (Yr26+Yr28+Yr29+Yr30+Yr65+Yr81)表现全生育期抗性。这些携带多个Yr基因的种质在田间对条锈病均表现出高抗或免疫水平。图3
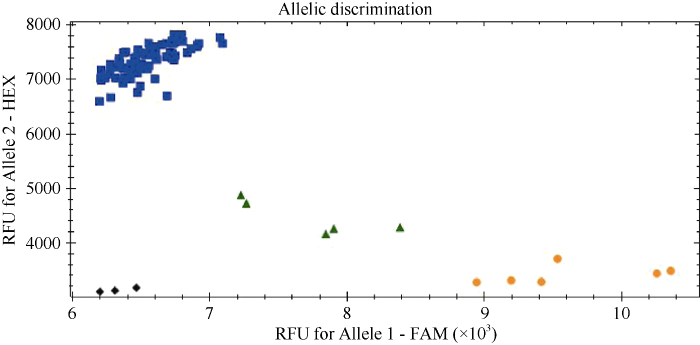 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3利用分子标记KASP_53113荧光定量PCR检测部分四川小麦育成品种(系)中Yr80基因
黄色: 携带Yr80的绵麦903、绵麦906、绵麦5658、川农30、川农32和川麦87。蓝色: 未携带Yr80的绵麦319、绵麦835、西科麦12、西科麦518、蜀麦1675和川麦605。绿色: Yr多基因品种(系)绵麦1501、蜀麦1871和川麦906。黑色: 空白对照H2O。
Fig. 3Detection of Yr80 based on real-time PCR using molecular marker KASP_53113 in the part of Sichuan wheat cultivars (lines)
Yellow: wheat cultivars carrying Yr80, Mianmai 903, Mianmai 906, Mianmai 5658, Chuannong 30, Chuannong 32, and Chuanmai 87. Blue: wheat cultivars without carrying Yr80, Mianmai 319, Mianmai 835, Xikemai 12, Xikemai 518, Shumai 1675, and Chuanmai 605. Green: Yr multigenic wheat cultivars (lines), Mianmai 1501, Shumai 1871, and Chuanmai 906. Black: H2O as blank control.
3 讨论
3.1 四川盆地小麦品种(系)条锈病抗性研究
四川盆地在我国西南麦区占有重要地位, 从1930年以来, 四川小麦育种已有80多年历史, 选育和推广具有优良农艺性状以及抗病抗虫的高产优质小麦品种是小麦育种的目标, 小麦育成品种的育种措施主要是杂交育种、诱变育种和杂交诱变相结合, 其中杂交育种是目前四川省品种选育的主要方式[54]。四川小麦通过与CIMMYT的冬春杂交穿梭育种培育出一大批高产, 优质和具有抗性的小麦中间材料或新品种, 使优异的抗性基因得到合理利用, 被广泛应用于小麦生产上[13]。还有一部分小麦种质由国内外育种专家利用小麦与外缘属种黑麦(Secale cereale L.)、簇毛麦(Haynaldia villosa L.)、粗山羊草(Aegilops tauschii)等创制的易位系进行育种, 如利用小麦-黑麦1RS?1BL易位系[55]作为亲本来源培育出的川农27、川农30和川农32等材料; 由小麦-簇毛麦6VS?6AL易位系选育而成的92R系列[56], 培育出蜀麦482、川麦91和川麦92等小麦品种; 以及利用CIMMYT的Syn-CD769和Syn-CD786等[57]人工合成小麦培育出绵麦367、川麦601和川麦602等优异的种质资源, 目前作为四川小麦育种中的重要抗性资源[54]。小麦条锈病是一种气传性真菌病害, 由于抗病基因布局单一, 当前小麦生产上大面积推广和种植的抗病品种携带相同抗病基因, 而长期以来病原菌与寄主之间协同进化, 极易导致抗病基因失效, 为小麦育种带来严峻挑战[58]。例如, 自20世纪60年代, 携带Yr9的小麦引进中国并广泛应用于小麦育种计划中, 到20世纪80年代末, 80%以上的小麦品种都携带Yr9, 然而在1985年出现的新小种CYR29导致携带Yr9的小麦品种丧失抗性, 造成1990年小麦产量损失265万吨[2,59-60]。因此培育具有持久抗性的小麦品种对可持续控制小麦条锈病至关重要[61]。四川省育种家加强了对条锈病的防控意识。2000年后, 在小麦品种审定中实行“条锈病抗性一票否决制”, 使小麦条锈病得到有效控制[21]。朱光等[62]利用CYR31、CYR32、CYR33的混合菌对427份湖北省小麦品系进行条锈病的抗性鉴定, 抗条锈病的有173份, 参试品系对条锈病抗性介于中抗和中感之间, 综合抗性水平不高。刘敏捷等[63]利用CYR32、CYR33的混合菌对369份山西省小麦区试品种以及育种材料进行条锈病抗性分析, 发现对小麦条锈病表现抗病的仅44份, 占11.9%, 抗性水平整体偏低, 建议在该地区加强新抗源的筛选和鉴定, 不断提高抗病育种水平。张培禹等[64], 王相权等[21], 曹世勤等[22]对部分四川省小麦品种(系)进行条锈病抗性鉴定, 抗性种质分别占供试材料的47.76%、21.75%和25.29%, 抗性水平整体偏低。本研究鉴定78份近几年选育的四川小麦品种(系)对当前流行的条锈菌均有较强的抗性, 与往年相比, 抗性品种所占比例明显提高, 可继续在小麦生产中使用。
3.2 四川小麦品种(系)抗条锈病基因布局
中国历史上发生过多次小麦条锈病大流行, 导致小麦严重减产, 其根本原因是使用单一或少数抗源, 小麦品种较快丧失抗性。分子标记辅助选择(MAS)可快速准确对抗病基因标记位点进行检测, 再与田间抗病性鉴定相结合, 可大大缩短育种年限, 有效提高育种效率[54]。Huang等[65]在2000—2016年黄淮海麦区审定的66份小麦品种中筛选到10份对CYR32、CYR33、CYR34都具有抗性的种质, 在24个品种中检测到Yr9、Yr10、Yr26和Yr32, 其中Yr9频率最高, 占28.8%, 所有种质都未检测到Yr5、Yr15和Yr18。本研究分子检测结果显示, 携带QYr.nwafu-4BL、Yr15、Yr17、Yr18、Yr26、Yr28、Yr29、Yr30、Yr39、Yr41、Yr65、Yr67、Yr78、Yr80和Yr81的材料各有5、5、45、2、30、5、30、39、3、2、22、8、23、6和24份。其中Yr18、Yr30和Yr65主要存在于川麦系列。Yr15源自四倍体野生二粒小麦(T. dicoccoides), 定位于1BS上, 本研究中5份种质检测出Yr15, 目前Yr15对中国的主流小种具有较好的抗性, 今后可在小麦育种中加强利用[32]。来自偏凸山羊草(Aegilops ventricosa Tsusch)的Yr17基因片段目前仍具有抗性[66], 本研究鉴定得到45份携带该基因的材料, 占比较高。Yr18作为慢锈性基因, 可延缓病情发展的速度, Lr34/Yr18/Pm38基因簇对叶锈病、条锈病和白粉病等多种真菌病害具有部分或持久抗性[39], 本研究中仅有川麦801和川麦1456检测出Yr18。董娜等[67]在348份小麦育成品种(系)中仅检测出7份携带Yr18的种质, 表明Yr18在中国育成品种(系)中分布频率较低, 在今后抗性育种中也应加强对此类慢锈性基因的利用。Yr26来源于中国圆锥小麦(Triticum turgidum L.) γ80-1, 被定位到染色体1BS上[68], 本研究筛选得到30份可能携带该基因的材料, 但Yr26对CYR34已失去抗性, 今后应谨慎使用, 避免单个基因应用到育种中, 应聚合不同抗病类型的基因培育持久抗性的新品种。Yr29源自普通小麦Pavon 76, 位于染色体1BL上[43], 本研究30份种质可能携带该基因。Yr30来源于普通小麦Opata 85, 位于染色体3BS上[44], 本研究检测出39份可能携带该基因的种质。Yr78源自普通小麦PI 519805[51], 本研究检测出23份可能含该基因的材料。Yr81被鉴定来自小麦地方品种Aus27430的染色体6AS上[53], 本研究得到24份可能携带该抗性基因的材料。仅西科麦18未检测到本研究中的19个抗条锈病基因, 推测可能含有其他已知或未知的抗条锈病基因。本研究结果可为育种家培育抗病新品种提供理论基础。研究表明, 提高有效抗性基因的利用率, 要因地制宜, 合理布局不同抗性类型基因, 从而实现小麦生产品种抗病基因多样化, 以培育出具有多基因聚合的持久抗性新品种[69]。周军等[69,70]对来自贵州、四川、山东等省的242份小麦品种(系)进行抗条锈病评价以及抗条锈病基因Yr5、Yr10、Yr15、Yr18、Yr26的分子检测, 发现具有成株期抗性的有贵协2号、贵农25号等116份材料, 同时含有2、3、4个抗条锈病基因的分别有39、7和1份, 如Yr26+Yr5、Yr26+Yr5+Yr15和Yr26+Yr15+Yr5+Yr10, 供试材料中携带Yr26基因的频率最高(24.38%)。蔚睿等[72]对2017-2019年黄淮麦区区试的150份小麦品种(系)进行条锈病抗性鉴定以及Yr5、Yr7、Yr9、Yr10、Yr17、Yr18、Yr26和YrSP 的分子标记检测, 鉴定到47份具有成株期抗性, 31份具有慢锈性, 其中69.3%的品种(系)携带Yr9, 16份材料同时携带2个抗性基因(Yr9+Yr17、Yr9+Yr18等), 2份携带3个抗条锈病基因(Yr9+Yr17+Yr18), 该研究发现供试材料的抗性主要是由多个基因提供。Liu等[73]对2个已知抗条锈病基因的小麦品种进行杂交, 培育具有显著改善条锈病抗性的小麦品系, 研究发现Yr15、Yr62和Yr65对目前的条锈菌生理小种仍具有较好的抗性, 在Yr10+Yr26, Yr26+Yr48和Yr30+Yr48等基因组合中存在加性效应或上位效应, 同时抗性基因的数量与小麦的农艺性状显著性较小。本研究筛选出含多基因的材料共74份, 同时携带2~6个抗性基因的材料分别有24、22、11、14和3份, 田间表型均高抗条锈病。Yr26主要以基因聚合形式存在, 其中绵麦1501含5个抗性基因Yr17+Yr26+Yr29+Yr78+Yr81, 川麦93和川麦906同时携带6个抗性基因Yr26+Yr28+Yr29+Yr30+Yr65+Yr81, Yr26虽对CYR34失去抗性, 但在多基因聚合的材料中不影响小麦品种的条锈病抗性。当前四川省选育的小麦品种(系)整体抗性水平较高, 可为小麦育种提供抗性亲本资源。若抗病品种的抗性年限能从目前的3~5年延长至8年, 也许就能使抗锈育种跟上品种的更新换代, 就需要利用分子标记辅助选择培育多基因聚合的品种, 提高抗性基因丰富度, 以有效提高品种的持久抗病性[69]。
4 结论
2016年以来四川小麦育成品种(系)对当前小麦条锈病流行小种具有较高抗性, 本研究中78份供试材料, 76份高抗条锈病, 2份中抗条锈病, 分子标记检测出74份为多基因聚合种质, 其抗性可能由多个基因共同作用, 同时材料中也可能存在新的抗条锈病基因, 如西科麦18, 可进一步加强对抗源和抗性基因的挖掘, 结合分子标记辅助选择应用于小麦育种中。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-020-03534-yURLPMID:31955232 [本文引用: 1]

KEY MESSAGE: A physical map of Secale cereale chromosome 6R was constructed using deletion mapping, and a new stripe rust resistance gene Yr83 was mapped to the deletion bin of FL 0.73-1.00 of 6RL. Rye (Secale cereale L., RR) possesses valuable genes for wheat improvement. In the current study, we report a resistance gene conferring stripe rust resistance effective from seedling to adult plant stages located on chromosome 6R. This chromosome was derived from triticale line T-701 and also carries highly effective resistance to the cereal cyst nematode species Heterodera avenae Woll. A wheat-rye 6R(6D) disomic substitution line exhibited high levels of seedling resistance to Australian pathotypes of the stripe rust (Puccinia striiformis f. sp. tritici; Pst) pathogen and showed an even greater resistance to the Chinese Pst pathotypes in the field. Ten chromosome 6R deletion lines and five wheat-rye 6R translocation lines were developed earlier in the attempt to transfer the nematode resistance gene to wheat and used herein to map the stripe rust resistance gene. These lines were subsequently characterized by sequential multicolor fluorescence in situ hybridization (mc-FISH), genomic in situ hybridization (GISH), mc-GISH, PCR-based landmark unique gene (PLUG), and chromosome 6R-specific length amplified fragment sequencing (SLAF-Seq) marker analyses to physically map the stripe rust resistance gene. The new stripe rust resistance locus was located in a chromosomal bin with fraction length (FL) 0.73-1.00 on 6RL and was named Yr83. A wheat-rye translocation line T6RL (#5) carrying the stripe rust resistance gene will be useful as a new germplasm in breeding for resistance.
URLPMID:30991940 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
URLPMID:30743706 [本文引用: 2]
A bud rot disease, referred to as
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
DOI:10.1038/s41477-018-0236-4URLPMID:30150615 [本文引用: 3]
Crop diseases reduce wheat yields by ~25% globally and thus pose a major threat to global food security(1). Genetic resistance can reduce crop losses in the field and can be selected through the use of molecular markers. However, genetic resistance often breaks down following changes in pathogen virulence, as experienced with the wheat yellow (stripe) rust fungus Puccinia striiformis f. sp. tritici (Pst)(2). This highlights the need to (1) identify genes that, alone or in combination, provide broad-spectrum resistance, and (2) increase our understanding of the underlying molecular modes of action. Here we report the isolation and characterization of three major yellow rust resistance genes (Yr7, Yr5 and YrSP) from hexaploid wheat (Triticum aestivum), each having a distinct recognition specificity. We show that Yr5, which remains effective to a broad range of Pst isolates worldwide, is closely related yet distinct from Yr7, whereas YrSP is a truncated version of Yr5 with 99.8% sequence identity. All three Yr genes belong to a complex resistance gene cluster on chromosome 2B encoding nucleotide-binding and leucine-rich repeat proteins (NLRs) with a non-canonical N-terminal zinc-finger BED domain(3) that is distinct from those found in non-NLR wheat proteins. We developed diagnostic markers to accelerate haplotype analysis and for marker-assisted selection to expedite the stacking of the non-allelic Yr genes. Our results provide evidence that the BED-NLR gene architecture can provide effective field-based resistance to important fungal diseases such as wheat yellow rust.
[本文引用: 1]
[本文引用: 1]
URLPMID:25336565 [本文引用: 2]
DOI:10.1038/s41467-017-02599-6URLPMID:29330508 [本文引用: 1]
Physical properties of crystalline materials often manifest themselves as atomic displacements either away from symmetry positions or driven by external fields. Especially the origin of multiferroic or magnetoelectric effects may be hard to ascertain as the related displacements can reach the detection limit. Here we present a resonant X-ray crystal structure analysis technique that shows enhanced sensitivity to minute atomic displacements. It is applied to a recently found crystalline modification of strontium titanate that forms in single crystals under electric field due to oxygen vacancy migration. The phase has demonstrated unexpected properties, including piezoelectricity and pyroelectricity, which can only exist in non-centrosymmetric crystals. Apart from that, the atomic structure has remained elusive and could not be obtained by standard methods. Using resonant X-ray diffraction, we determine atomic displacements with sub-picometer precision and show that the modified structure of strontium titanate corresponds to that of well-known ferroelectrics such as lead titanate.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-009-1097-zURLPMID:19578829 [本文引用: 3]
The locus Lr34/Yr18/Pm38 confers partial and durable resistance against the devastating fungal pathogens leaf rust, stripe rust, and powdery mildew. In previous studies, this broad-spectrum resistance was shown to be controlled by a single gene which encodes a putative ATP-binding cassette transporter. Alleles of resistant and susceptible cultivars differed by only three sequence polymorphisms and the same resistance haplotype was found in the three independent breeding lineages of Lr34/Yr18/Pm38. Hence, we used these conserved sequence polymorphisms as templates to develop diagnostic molecular markers that will assist selection for durable multi-pathogen resistance in breeding programs. Five allele-specific markers (cssfr1-cssfr5) were developed based on a 3 bp deletion in exon 11 of the Lr34-gene, and one marker (cssfr6) was derived from a single nucleotide polymorphism in exon 12. Validation of reference genotypes, well characterized for the presence or absence of the Lr34/Yr18/Pm38 resistance locus, demonstrated perfect diagnostic values for the newly developed markers. By testing the new markers on a larger set of wheat cultivars, a third Lr34 haplotype, not described so far, was discovered in some European winter wheat and spelt material. Some cultivars with uncertain Lr34 status were re-assessed using the newly derived markers. Unambiguous identification of the Lr34 gene aided by the new markers has revealed that some wheat cultivars incorrectly postulated as having Lr34 may possess as yet uncharacterised loci for adult plant leaf and stripe rust resistance.
DOI:10.1007/s00122-020-03589-xURLPMID:32281004 [本文引用: 1]
KEY MESSAGE: Heterogeneous Lr34 genes for leaf rust in winter wheat cultivar 'Duster' and KASP markers for allelic variation in exon 11 and exon 22 of Lr34. Wheat, Triticum aestivum (2n = 6x = 42, AABBDD), is a hexaploid species, and each of three homoeologous genomes A, B, and D should have one copy for a gene in its ancestral form if the gene has no duplication. Previously reported leaf rust resistance gene Lr34 has one copy on the short arm of chromosome 7D in hexaploid wheat, and allelic variation in Lr34 is in intron 4, exon 11, exon 12, or exon 22. In this study, we discovered that Oklahoma hard red winter wheat cultivar 'Duster' (PI 644,016) has two copies of the Lr34 gene, the resistance allele Lr34a and the susceptibility allele Lr34b. Both Lr34a and Lr34b were mapped in the same linkage group on chromosome 7D in a doubled-haploid population generated from a cross between Duster and a winter wheat cultivar 'Billings' which carries the susceptibility allele Lr34c. A chromosomal fragment including Lr34 and at least two neighboring genes on its proximal side but excluding genes on its distal side was duplicated in Duster. The Duster Lr34ab allele was associated with tip necrosis and increased resistance against leaf rust at adult plants in the Duster x Billings DH population tested in the field, demonstrating the function of the Duster Lr34ab allele in wheat. We have developed KASP markers for allelic variation in exon 11 and exon 22 of Lr34 in wheat. These markers can be utilized to accelerate the selection of Lr34 in wheat.
[本文引用: 2]
[本文引用: 2]
DOI:10.1094/PDIS-06-16-0890-REURLPMID:30677352 [本文引用: 2]
Leaf rust (LR) and stripe rust (YR) are important diseases of wheat worldwide. We used 148 recombinant inbred lines (RIL) from the cross of Avocet x Kundan for determining and mapping the genetic basis of adult plant resistance (APR) loci. The population was phenotyped LR and YR for three seasons in field trials conducted in Mexico and genotyped with the diversity arrays technology sequencing (DArT-Seq) and simple sequence repeat markers. The final genetic map was constructed using 2,937 polymorphic markers with an average distance of 1.29 centimorgans between markers. Inclusive composite interval mapping identified two co-located APR quantitative trait loci (QTL) for LR and YR, two LR QTL, and three YR QTL. The co-located resistance QTL on chromosome 1BL corresponded to the pleiotropic APR gene Lr46/Yr29. QLr.cim-2BL, QYr.cim-2AL, and QYr.cim-5AS could be identified as new resistance loci in this population. Lr46/Yr29 contributed 49.5 to 65.1 and 49.2 to 66.1% of LR and YR variations, respectively. The additive interaction between detected QTL showed that LR severities for RIL combining four QTL ranged between 5.3 and 25.8%, whereas the lowest YR severities were for RIL carrying QTL on chromosomes 1BL + 2AL + 6AL. The high-density DArT-Seq markers across chromosomes can be used in fine mapping of the targeted loci and development SNP markers.
DOI:10.1007/s00122-004-1787-5URLPMID:15340687 [本文引用: 2]
The stem rust resistance gene Sr2 has provided durable broad-spectrum, adult-plant resistance to the fungal pathogen Puccinia graminis Pers. f. sp. tritici throughout wheat-growing regions of the world for more than 50 years. The ability to select for Sr2 in wheat breeding programs was recently improved by the identification of a tightly linked microsatellite marker gwm533. This marker typically amplifies a 120-bp polymerase chain reaction fragment from wheat lines carrying Sr2. In instances where the 120-bp fragment is not associated with the presence of Sr2, DNA sequence analysis has shown that a second allele was amplified, differing in the structure of the microsatellite repeat. To discriminate this allelic homoplasy (alleles identical in size, but not identical by descent), sequence-tagged microsatellites (STM) markers were developed for the Xgwm533 locus. These markers were shown to be diagnostic for the presence of Sr2 in a wide range of germplasm, representative of all major wheat varieties historically grown in Australia. The STMs will be particularly useful for marker-assisted selection in Southern Australian breeding programs, where the use of the marker gwm533 is often precluded by the presence of the non- Sr2-associated 120-bp allele in the pedigree of current breeding germplasm. The STMs also revealed a high incidence of previously undetected allelic homoplasy at the Xgwm533 locus and may have broader utility in genetic research and breeding, as this locus is also reported to be strongly associated with a major gene conferring resistance to Fusarium head blight.
DOI:10.1007/s00122-016-2672-8URLPMID:26847646 [本文引用: 2]
KEY MESSAGE: The wheat stripe rust resistance gene Yr36 ( WKS1 ) with a unique kinase-START domain architecture is highly conserved in wild emmer wheat natural populations. Wild emmer wheat (Triticum dicoccoides) populations have developed various resistance strategies against the stripe rust pathogen Puccinia striiformis f. sp. tritici (Pst). The wild emmer gene, Yr36 (WKS1), which confers partial resistance to a broad spectrum of Pst races, is composed of a kinase and a START lipid-binding domain, a unique gene architecture found only in the Triticeae tribe. The analysis of 435 wild emmer accessions from a broad range of natural habitats revealed that WKS1 and its paralogue WKS2 are present only in the southern distribution range of wild emmer in the Fertile Crescent, supporting the idea that wheat domestication occurred in the northern populations. An analysis of full-length WKS1 sequence from 54 accessions identified 15 different haplotypes and very low-nucleotide diversity (pi = 0.00019). The high level of WKS1 sequence conservation among wild emmer populations is in contrast to the high level of diversity previously observed in NB-LRR genes (e.g., Lr10 and Pm3). This phenomenon may reflect the different resistance mechanisms and different evolutionary pathways that shaped these genes, and may shed light on the evolution of genes that confer partial resistance to stripe rust. Only five WKS1 coding sequence haplotypes were revealed among all tested accessions, encoding four different putative WKS1 proteins (designated P0, P1, P2, and P3). Infection tests showed that P0, P1, and P3 haplotypes display a resistance response, while P2 displayed a susceptible response. These results show that the WKS1 proteins (P0, P1, and P3) can be useful to improve wheat resistance to stripe rust.
DOI:10.1007/s00122-007-0518-0URLPMID:17318493 [本文引用: 1]
Stripe rust, caused by Puccinia striiformis f. sp. tritici, is one of the most widespread and destructive wheat diseases worldwide. Growing resistant cultivars is the preferred control of the disease. The spring wheat cultivar 'Alpowa' has both race-specific, all-stage resistance and non-race-specific, high-temperature adult-plant (HTAP) resistances to stripe rust. To identify genes for the stripe rust resistances, Alpowa was crossed with 'Avocet Susceptible' (AVS). Seedlings of the parents, and F(1), F(2) and F(3) progeny were tested with races PST-1 and PST-21 of P. striiformis f. sp. tritici under controlled greenhouse conditions. Alpowa has a single partially dominant gene, designated as YrAlp, conferring all-stage resistance. Resistance gene analog polymorphism (RGAP) and simple sequence repeat (SSR) techniques were used to identify molecular markers linked to YrAlp. A linkage group of five RGAP markers and two SSR markers was constructed for YrAlp using 136 F(3) lines. Amplification of a set of nulli-tetrasomic Chinese Spring lines with RGAP markers Xwgp47 and Xwgp48 and the two SSR markers indicated that YrAlp is located on the short arm of chromosome 1B. To map quantitative trait loci (QTLs) for the non-race-specific HTAP resistance, the parents and 136 F(3) lines were tested at two sites near Pullman and one site near Mount Vernon, Washington, under naturally infected conditions. A major HTAP QTL was consistently detected across environments and was located on chromosome 7BL. Because of its chromosomal location and the non-race-specific nature of the HTAP resistance, this gene is different from previously described genes for adult-plant resistance, and is therefore designated Yr39. The gene contributed to 64.2% of the total variation of relative area under disease progress curve (AUDPC) data and 59.1% of the total variation of infection type data recorded at the heading-flowering stages. Two RGAP markers, Xwgp36 and Xwgp45 with the highest R (2) values were closely linked to Yr39, should be useful for incorporation of the non-race-specific resistance gene into new cultivars and for combining Yr39 with other genes for durable and high-level resistance.
URLPMID:18956025 [本文引用: 2]
DOI:10.1007/s00122-011-1573-0URLPMID:21455722 [本文引用: 2]
A mapping population of 186 recombinant inbred lines developed from a cross between UC1110, an adapted California spring wheat, and PI610750, a synthetic derivative from CIMMYT's Wide Cross Program, was evaluated for its response to current California races of stripe rust (Puccinia striiformis f. sp. tritici) in replicated field trials over four seasons (2007-2010) in the northern Sacramento Valley. A genetic map was constructed consisting of 1,494 polymorphic probes (SSRs, DArTs, and ESTs) mapped to 558 unique loci, and QTL analysis revealed the presence of four stripe rust resistance QTL segregating in this population, two from UC1110 (on chromosomes 3BS and 2BS) and two from PI610750 (5AL and 2AS). The two QTL of largest effects (on 3BS and 5AL) were validated in independent populations and their intervals narrowed to 2.5 and 5.3 cM, respectively. The 3BS QTL was shown, by allelism test and genotype, to carry a gene different from the Yr30/Sr2 complex. Mapped position also suggests that the 3BS QTL is associated with a gene different from either Yrns-B1 or YrRub, two stripe rust resistance genes mapped to this region in other studies. The 5AL QTL carries a previously unreported partial stripe rust resistance gene, designated here as Yr48. This paper discusses the individual contributions to resistance of these four QTL, their epistatic interactions, and their potential in durable resistance breeding strategies based on combinations of partial resistance genes.
DOI:10.1007/s00122-014-2378-8URLPMID:25142874 [本文引用: 1]
KEY MESSAGE: This manuscript reports two new genes ( Yr64 and Yr65 ) for effective resistance to stripe rust and usefulness of their flanking SSR markers for marker-assisted selection. Stripe rust, caused by Puccinia striiformis f. sp. tritici (Pst), is one of the most important diseases of wheat worldwide and resistance is the best control strategy. Durum wheat accessions PI 331260 and PI 480016 were resistant to all tested Pst races. To transfer the resistance genes to common wheat and map them to wheat chromosomes, both accessions were crossed with the stripe rust-susceptible spring wheat 'Avocet S'. Resistant F3 plants with 42 chromosomes were selected cytologically and by rust phenotype. A single dominant gene for resistance was identified in segregating F4 lines from each cross. F6 populations for each cross were developed from single F5 plants and used for genetic mapping. Different genes from PI 331260 and PI 480016 were mapped to different loci in chromosome 1BS using simple sequence repeat markers. The gene from PI 331260 was flanked by Xgwm413 and Xgdm33 in bin 1BS9-0.84-1.06 at genetic distances of 3.5 and 2.0 cM; and the gene from PI 480016 was flanked by Xgwm18 and Xgwm11 in chromosome bin C-1BS10-0.50 at 1.2 and 2.1 cM, respectively. Chromosomal locations and race and allelism tests indicated that the two genes are different from previously reported stripe rust resistance genes, and therefore are named as Yr64 from PI 331260 and Yr65 from PI 480016. These genes and their flanking markers, and selected common wheat lines with the genes should be valuable for diversifying resistance genes used in breeding wheat cultivars with stripe rust resistance.
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s00122-018-3090-xURLPMID:29560515 [本文引用: 1]
KEY MESSAGE: A new adult plant stripe rust resistance gene, Yr80, was identified in a common wheat landrace Aus27284. Linked markers were developed and validated for their utility in marker-assisted selection. Stripe rust, caused by Puccinia striiformis f. sp. tritici (Pst), is among the most important constraints to global wheat production. The identification and characterisation of new sources of host plant resistance enrich the gene pool and underpin deployment of resistance gene pyramids in new cultivars. Aus27284 exhibited resistance at the adult plant stage against predominant Pst pathotypes and was crossed with a susceptible genotype Avocet S. A recombinant inbred line (RIL) population comprising 121 lines was developed and tested in the field at three locations in 2016 and two in 2017 crop seasons. Monogenic segregation for adult plant stripe rust response was observed among the Aus27284/Avocet S RIL population and the underlying locus was temporarily designated YrAW11. Bulked-segregant analysis using the Infinium iSelect 90K SNP wheat array placed YrAW11 in chromosome 3B. Kompetitive allele specific PCR (KASP) primers were designed for the linked SNPs and YrAW11 was flanked by KASP_65624 and KASP_53292 (3 cM) proximally and KASP_53113 (4.9 cM) distally. A partial linkage map of the genomic region carrying YrAW11 comprised nine KASP and two SSR markers. The physical position of KASP markers in the pseudomolecule of chromosome 3B placed YrAW11 in the long arm and the location of markers gwm108 and gwm376 in the deletion bin 3BL2-0.22 supported this conclusion. As no other stripe rust resistance locus has been reported in chromosome 3BL, YrAW11 was formally designated Yr80. Marker KASP_ 53113 was polymorphic among 94% of 81 Australian wheat cultivars used for validation.
DOI:10.1094/PDIS-06-18-1055-REURLPMID:30998448 [本文引用: 2]
The deployment of diverse sources of resistance in new cultivars underpins durable control of rust diseases. Aus27430 exhibited a moderate level of stripe rust resistance against Puccinia striiformis f. sp. tritici (Pst) pathotypes currently prevalent in Australia. Aus27430 was crossed with the susceptible parent Avocet S (AvS) and subsequent filial generations were raised. Monogenic segregation observed among Aus27430/AvS F3 families was confirmed through stripe rust screening of an F6 recombinant inbred line (RIL) population, and the resistance locus was temporarily named YrAW5. Selective genotyping using an Illumina iSelect 90K wheat SNP bead chip array located YrAW5 in chromosome 6A. Genetic mapping of the RIL population with linked 90K SNPs that were converted into PCR-based marker assays, as well as SSR markers previously mapped to chromosome 6A, confirmed the chromosomal assignment for YrAW5. Comparative analysis of other stripe rust resistance genes located in chromosome 6A led to the formal designation of YrAW5 as Yr81. Tests with a marker linked with Yr18 also demonstrated the presence of this gene in Aus27430. Yr18 interacted with Yr81 to produce stripe rust responses lower than those produced by RILs carrying these genes individually. Although gwm459 showed higher recombination with Yr81 compared with the other flanking marker KASP_3077, it amplified the AvS allele in 80 cultivars, whereas KASP_3077 amplified AvS allele in 67 cultivars. Both markers can be used in marker-assisted selection after confirming parental polymorphism.
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
DOI:10.1007/BF00223930URLPMID:24170007 [本文引用: 1]
Several Triticum aestivum L.-Haynaldia villosa disomic 6VS/6AL translocation lines with powdery mildew resistance were developed from the hybridization between common wheat cultivar Yangmai 5 and alien substitution line 6V(6A). Mitotic and meiotic C-banding analysis, aneuploid analysis with double ditelosomic stocks, in situ hybridization, as well as the phenotypic assessment of powdery mildew resistance, were used to characterize these lines. The same translocated chromosome, with breakpoints near the centromere, appears to be present in all the lines, despite variation among the lines in their morphology and agronomic characteristics. The resistance gene, conferred by H. villosa and designated as Pm21, is a new and promising source of powdery mildew resistance in wheat breeding.
DOI:10.1016/S1673-8527(08)60145-9URLPMID:19782955 [本文引用: 1]
Synthetic hexaploid wheat (Triticum turgidumxAegilops tauschii) was created to explore for novel genes from T. turgidum and Ae. tauschii that can be used for common wheat improvement. In the present paper, research advances on the utilization of synthetic hexaploid wheat for wheat genetic improvement in China are reviewed. Over 200 synthetic hexaploid wheat (SHW) accessions from the International Maize and Wheat Improvement Centre (CIMMYT) were introduced into China since 1995. Four cultivars derived from these, Chuanmai 38, Chuanmai 42, Chuanmai 43 and Chuanmai 47, have been released in China. Of these, Chuanmai 42, with large kernels and resistance to stripe rust, had the highest average yield (>6 t/ha) among all cultivars over two years in Sichuan provincial yield trials, outyielding the commercial check cultivar Chuanmai 107 by 22.7%. Meanwhile, by either artificial chromosome doubling via colchicine treatment or spontaneous chromosome doubling via a union of unreduced gametes (2n) from T. turgidum-Ae. tauschii hybrids, new SHW lines were produced in China. Mitotic-like meiosis might be the cytological mechanism of spontaneous chromosome doubling. SHW lines with genes for spontaneous chromosome doubling may be useful for producing new SHW-alien amphidiploids and double haploid in wheat genetic improvement.
[本文引用: 1]
DOI:10.1094/PDIS.2004.88.8.896URLPMID:30812521 [本文引用: 1]
In China, wheat stripe rust, caused by Puccinia striiformis f. sp. tritici, is one of the most destructive diseases of wheat and can cause severe yield losses when susceptible cultivars are grown and weather conditions are favorable for the disease. Wheat stripe rust most frequently affects the winter wheat growing areas in Northwest, Southwest, and North China, and the spring wheat growing areas in Northwest China. In the 2001-2002 growing season, a widespread stripe rust epidemic affected about 6.6 million hectares of wheat in 11 provinces: Si-chuan, Chongqing, eastern Gansu, southern and western Shaanxi, southern and central Ningxia, Yunnan, Guizhou, Hubei, Henan, southern and central Hebei, and Shandong. The epidemic could be attributed to relatively warm weather from November 2001 to March 2002, high frequencies of stripe rust races CYR31 and CYR32, and widely grown susceptible cultivars. Race CYR31 was virulent on the Chinese differential cultivars Trigo Eureka, Fulhard, Lutescens 128, Mentana, Virgilio, Abbondanza, Early Premium, Funo, Danish 1, Fengchan 3, Lovrin 13, Shui-yuan 11, Lovrin 10, and Hybrid 46. Race CYR32 had all the virulence factors of CYR31, plus virulences on Chinese differential cultivars Jubilejina 2 and Kangyin 655, i.e., CYR32 was virulent on all differential cultivars, except Zhong 4. When tested on the world and European differential and some other resistant genotypes, CYR32 was virulent on Chinese 166 (Yr1), Heines VII (Yr2, Yr25, and YrHVII), Vilmorin 23 (Yr3a and Yr4a), Heines Kolben (Yr6 and YrHK), Lee (Yr7, Yr22, and Yr23), Clement (Yr9, Yr25, YrCle), VPM1 (Yr17), Selkirk (Yr27), Anza (YrA), Carstens V (YrCV1, YrCV2, and YrCV3), Gaby (YrG), Strubes Dickkopf (Yr25), and Suwon 92/Omar (YrSO). Resistance genes in Triticum spelta album (Yr5), Zhong 4, and Moro (Yr10 and YrMor) were effective against all races identified.
DOI:10.1094/PDIS-93-11-1093URLPMID:30754577 [本文引用: 1]
Stripe (or yellow) rust caused by Puccinia striiformis f. sp. tritici is the most destructive foliar disease of wheat in China. The pathogen populations were analyzed for virulence evolution, complexity, phenotypic dynamics, and diversity on temporal and spatial bases. A total of 41 races were identified and characterized from 4,714 stripe rust isolates collected during 2003 through 2007 from wheat growing areas in 15 provinces in China. The races were based on avirulence/virulence patterns to 19 differential host genotypes. Chinese stripe rust population exhibited high diversity with a complex virulence structure. Comparisons using the relative Shannon's index indicated that some differences in the richness and evenness of races were present in pathogen populations within years and between regions despite a national tendency to reduced diversity over time. A noticeably increased frequency of race CYR33 (Chinese yellow rust 33) with virulence for YrSu was the major virulence change recorded in this study compared to the results on an annual basis. Isolates of Puccinia striiformis f. sp. tritici from different regions showed differences in the composition of races, distribution frequency, and diversity. The uneven distribution of major races and comparatively greater diversity in the Northwest and Southwest regions than that in the Huang-Huai-Hai region suggest that long-distance migrations of the pathogen occur from one or more over-summering areas eastward into over-wintering areas. This supports the hypothesis that southern Gansu and northwestern Sichuan comprises a
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1094/PDIS-10-19-2119-REURLPMID:32293996 [本文引用: 1]
Wheat stripe (yellow) rust, caused by Puccinia striiformis f. sp. tritici (Pst), is a serious fungal disease worldwide, especially in the Huang-Huai-Hai region, a main wheat production area in China. Gene postulation, molecular testing, and pedigree analysis were conducted to determine the presence of stripe rust resistance genes to 15 Pst races in 66 selected commercial wheat cultivars released from 2000 to 2016. In addition, races CYR32, CYR33, and CYR34 were used to evaluate resistance to Pst at the adult-plant stage of wheat in the field. Four Yr genes (Yr9, Yr10, Yr26, and Yr32) were postulated in 24 wheat cultivars either singly or in combination. Thirty-six cultivars might contain unknown Yr genes, whereas no identified Yr gene was postulated in six cultivars. Yr9 was detected at a frequency of 28.8%, and no cultivars carried Yr5, Yr15, or Yr18. Ten cultivars (15.2%) exhibited adult-plant resistance in the field tests with three predominant races. Three cultivars (Langyan 43, Xinong 889, and Yunfeng 139) had all-stage resistance. These results are useful to growers selecting cultivars and to breeders aiming to use more resistance genes to develop new cultivars with effective resistance in order to reduce stripe rust damage.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}