Identification and expression of PAL genes in sisal
HUANG Xing,1, XI Jin-Gen1, CHEN Tao2, QIN Xu2, TAN Shi-Bei1, CHEN He-Long3, YI Ke-Xian,1,*1Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences/Key Laboratory of Integrated Pest Management on Tropical Crops, Ministry of Agriculture and Rural Affairs/Hainan Key Laboratory for Monitoring and Control of Tropical Agricultural Pests, Haikou 571101, Hainan, China 2Guangxi Subtropical Crops Research Institute, Nanning 530001, Guangxi, China 3Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, Hainan, China
The National Key Research and Development Program of China.2018YFD0201100 The China Agriculture Research System.CARS-16 The Hainan Provincial Natural Science Foundation of China.319QN275 The Hainan Provincial Natural Science Foundation of China.320RC698 The Guangxi Key Research and Development Program.桂科AB18221105 The Belt and Road Tropical Project.BARTP-08
Abstract Sisal is an important fiber crop in tropical areas, but its research foundation of molecular biology is relatively weak, and the mechanism of fiber development still remains unclear. Phenylalanine ammonia-lyase (PAL) is the first enzyme of lignin bio-synthesis, which is an important component of fiber. According to published transcriptome data, two sisal PAL genes with complete coding sequences were successfully identified. Their expression patterns during sisal leaf development were consistent with previously reported PAL activity changes during fiber development, indicating that PAL was closely related to lignin bio-synthesis. Phylogenetic analysis showed that sisal PALs were closely related with Agave americana. Selection pressure analysis showed similar selection pressure of PALs in sisal and A. americana, which were higher than those in A. tequilana. This might be caused by the convergent evolution of fiber-related traits in sisal and A. americana. In addition, sisal PALs were not significantly expressed under neither copper nor lead stress, which might be caused by post-transcriptional regulation under heavy metal stresses. It was worth noting that the expression of sisal PALs was highly up-regulated after Phytophthora nicotianae Breda inoculation. Sisal PALs might participate in the bio-synthesis of disease resistance-related secondary metabolites in phenylpropanoid pathway, as well as plant cell-wall mediated immunity. Therefore, functional characterization of sisal PALs could improve the understanding of mechanisms in fiber development and disease resistance, which is of great importance for breeding new sisal varieties with high yield, high quality and multiple resistance. Keywords:sisal;PAL gene;phylogenetic analysis;selection pressure;expression pattern;adverse stress
PDF (597KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄兴, 习金根, 陈涛, 覃旭, 谭施北, 陈河龙, 易克贤. 剑麻苯丙氨酸裂解酶基因的鉴定及表达分析[J]. 作物学报, 2021, 47(6): 1082-1089. doi:10.3724/SP.J.1006.2021.04116 HUANG Xing, XI Jin-Gen, CHEN Tao, QIN Xu, TAN Shi-Bei, CHEN He-Long, YI Ke-Xian. Identification and expression of PAL genes in sisal[J]. Acta Agronomica Sinica, 2021, 47(6): 1082-1089. doi:10.3724/SP.J.1006.2021.04116
物种缩写同图1。使用DnaSP进行滑窗法分析, 窗口长度为30 bp, 步移长度为6 bp。 Fig. 2Selection pressure analysis of PALs in Agave
Abbreviations of species are the same as those given in Fig. 1. Sliding window analysis was conducted by DnaSP with a window size of 30 bp and a step size of six bp.
Xu NK, Yu RS, Sun GM. Effects of different levels of NPKCaMg fertilizers on yield, fibre quality and nutrient content of Agave H.11648 Chin J Trop Crops, 1994,15(1):39-45 (in Chinese with English abstract). [本文引用: 1]
HuangX, ChenT, Xi JG, He CP, Wu WH, Liang YQ, Zheng JL, LiR, Yi KX. The Correlation between single leaf traits and fresh yield of sisal Plant Fiber Sci China, 2018,40(2):70-74 (in Chinese with English abstract). [本文引用: 1]
HuangX, XiaoM, XiJ, HeC, ZhengJ, ChenH, GaoJ, ZhangS, WuW, LiangY, XieL, YiK. De novo transcriptome assembly of Agave H11648 by Illumina sequencing and identification of cellulose synthase genes in Agave species Genes, 2019,10:103. [本文引用: 6]
DengG, HuangX, XieL, TanS, Gbokie TJ, BaoY, XieZ, YiK. Identification and expression of SAUR genes in the CAM plant agave Genes, 2019,10:555. [本文引用: 3]
HuangX, WangB, XiJ, ZhangY, HeC, ZhengJ, GaoJ, ChenH, ZhangS, WuW, LiangY, YiK. Transcriptome comparison reveals distinct selection patterns in domesticated and wild Agave species, the important CAM plants Int J Genomics, 2018,2018:5716518. [本文引用: 5]
Li LB, LiuL, He CF, Dong YM, Peng ZH. Research progresses on the genes encoding the key enzymes in biosynthetic pathway of lignin Mol Plant Breed, 2007,5(S1):45-51 (in Chinese with English abstract). [本文引用: 2]
Shi HY, Zhang YX. Molecular characterization of key enzyme genes related to the pathway of lignin biosynthesis Chin Agric Sci Bull, 2011,27(5):288-291 (in Chinese with English abstract). [本文引用: 1]
Wanner LA, LiG, WareD, Somssich IE, Davis KR. The phenylalanine ammonia-lyase gene family in Arabidopsis thaliana Plant Mol Biol, 1995,27:327-338. [本文引用: 1]
Olsen KM, Lea US, SlimestadR, VerheulM, LilloC. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J Plant Physiol, 2008,165:1491-1499. [本文引用: 1]
RaesJ, RohdeA, Christensen JH, Van YD P, BoerjanW. Genome-wide characterization of the lignification toolbox in Arabidopsis Plant Physiol, 2003,133:1051-1071. [本文引用: 1]
Yu XZ, Fan WJ, Lin YJ, Zhang FF, Gupta DK. Differential expression of the PAL gene family in rice seedlings exposed to chromium by microarray analysis Ecotoxicology, 2018,27:325-335. [本文引用: 2]
JaillonO, Aury JM, NoelB, PolicritiA, ClepetC, CasagrandeA, ChoisneN, AubourgS, VituloN, JubinC. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla Nature, 2007,449:463. [本文引用: 1]
ShiR, Sun YH, LiQ, HeberS, SederoffR, Chiang VL. Towards a systems approach for lignin biosynthesis in Populus trichocarpa: transcript abundance and specificity of the monolignol biosynthetic genes Plant Cell Physiol, 2010,51:144-163. [本文引用: 1]
Wada KC, MizuuchiK, KoshioA, KanekoK, MitsuiT, TakenoK. Stress enhances the gene expression and enzyme activity of phenylalanine ammonia-lyase and the endogenous content of salicylic acid to induce flowering in pharbitis J Plant Physiol, 2014,171:895-902. [本文引用: 1]
Li FY, Zhang LM, Li XM, GuoB, Chen LY, Qi ZP. Sisal tolerance of cupreous and its accumulation preliminary explore Chin Agric Sci Bull, 2006,22(12):417-420 (in Chinese with English abstract). [本文引用: 1]
Chen LY, Zhang LM, Li FY, GuoB, Li XM, Liao XJ, Qi ZP. A primary research on sisal’s uptake property and the accumulation rule to Pb ions J Agro-Environ Sci, 2007,26:1879-1883 (in Chinese with English abstract). [本文引用: 1]
WangP, Gao JM, YangF, Zheng JL, Liu QL, Chen HL, Yi KX. Transcriptome of sisal leaf pretreated with Phytophthora nicotianae Breda Chin J Trop Crops, 2014,35:576-582 (in Chinese with English abstract). [本文引用: 1]
Rombel IT, Sykes KF, RaynerS, Johnston SA. ORF-FINDER: a vector for high-throughput gene identification Gene, 2002,282:33-41. [本文引用: 1]
Wilkins MR, GasteigerE, BairochA, Sanchez JC, Williams KL, Appel RD, Hochstrasser DF. Protein identification and analysis tools in the ExPASy server Methods Mol Biol, 1999,112:531-552. [本文引用: 1]
YuC, ChenY, LuC, HwangJ. Prediction of protein subcellular localization Proteins, 2006,64:643-651. [本文引用: 1]
Gross SM, Martin JA, SimpsonJ, Abraham-Juarez MJ, WangZ, ViselA. De novo transcriptome assembly of drought tolerant CAM plants,Agave deserti and Agave tequilana BMC Genomics, 2013,14:563. [本文引用: 2]
Abraham PE, YinH, Borland AM, WeighillD, Lim SD, De PaoliH C, EngleN, Jones PC, AghR, Weston DJ, Wullschleger SD, TschaplinskiT, JacobsonD, Cushman JC, Hettich RL, Tuskan GA, YangX. Transcript, protein and metabolite temporal dynamics in the CAM plant Agave Nat Plants, 2016,2:16178. [本文引用: 2]
Larkin MA, BlackshieldsG, Brown NP, ChennaR, McGettigan PA, McWilliamH, ValentinF, Wallace IM, WilmA, LopezR, Thompson JD, Gibson TJ, Higgins DG. Clustal W and Clustal X version 2.0 Bioinformatics, 2007,23:2947-2948. [本文引用: 1]
LibradoP, RozasJ. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data Bioinformatics, 2009,25:1451-1452. [本文引用: 1]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method Methods, 2001,25:402-408. [本文引用: 1]
HuangJ, GuM, LaiZ, FanB, ShiK, Zhou YH, Yu JQ, ChenZ. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress Plant Physiol, 2010,153:1526-1538. [本文引用: 1]
YanF, Li HZ, ZhaoP. Genome-wide identification and transcriptional expression of the PAL gene family in common walnut(Juglans regia L.) Genes, 2019,10:46. [本文引用: 2]
Li JJ, Guo QQ, Chen JR. Study on related enzymes of lignin synthesis in ramie Plant Fiber Sci China, 2009,31(2):119-124 (in Chinese with English abstract). [本文引用: 1]
PicherskyE, LewinsohnE. Convergent evolution in plant specialized metabolism Annu Rev Plant Biol, 2011,62:549-566. [本文引用: 1]
SghaierA, ChaabouniY, MsahliS, SakliF. Morphological and crystalline characterization of NaOH and NaOCl treated Agave americana L. fiber Ind Crops Prod, 2012,36:257-266. [本文引用: 1]
Ková?ikJ, KlejdusB, HedbavnyJ, ZońJ. Copper uptake is differentially modulated by phenylalanine ammonia-lyase inhibition in diploid and tetraploid chamomile J Agric Food Chem, 2010,58:10270-10276. [本文引用: 1]
Pawlak-SpradaS, Arasimowicz-JelonekM, PodgórskaM, DeckertJ. Activation of phenylpropanoid pathway in legume plants exposed to heavy metals. Part I. Effects of cadmium and lead on phenylalanine ammonia-lyase gene expression, enzyme activity and lignin content Acta Biochim Pol, 2011,58:211-216. URLPMID:21503278 [本文引用: 1] Species-specific changes in expression of phenylalanine ammonia-lyase (PAL) and lignin content were detected in roots of soybean (Glycine max L.) and lupine (Lupinus luteus L.) seedlings treated with different concentrations of cadmium (Cd(2+), 0-25 mg/l) or lead (Pb(2+), 0-350 mg/l). The stimulatory effect of both metals was observed in mRNA coding for PAL in soybean. In the case of lupine, changes of PAL mRNA level were dependent on the metal used: Cd(2+) caused a decrease, whereas Pb(2+) an increase of PAL transcript level. The activity of PAL was enhanced in both plant species at higher metal concentrations (15-25 mg/l of Cd(2+) or 150-350 mg/l of Pb(2+)); however it was not directly correlated with PAL mRNA. This suggests a transcriptional and posttranscriptional control of PAL expression under heavy metals stress. In soybean, Cd(2+) or Pb(2+) treatment increased lignin content, while in lupine the effect was opposite. The decreased lignin accumulation in lupine roots in response to heavy metals, despite an increased PAL activity, suggests that the activated phenylpropanoid pathway was involved in the synthesis of secondary metabolites other than lignin.
WangR, Wang GL, NingY. PALs: emerging key players in broad-spectrum disease resistance Trends Plant Sci, 2019,24:785-787. [本文引用: 1]
DuanL, Liu HB, Li XH, Xiao JH, Wang SP. Multiple phytohormones and phytoalexins are involved in disease resistance to Magnaporthe oryzae invaded from roots in rice Physiol Plant, 2014,152:486-500. URLPMID:24684436 [本文引用: 1]
 ,1, 习金根1, 陈涛2, 覃旭2, 谭施北1, 陈河龙3, 易克贤
,1, 习金根1, 陈涛2, 覃旭2, 谭施北1, 陈河龙3, 易克贤
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT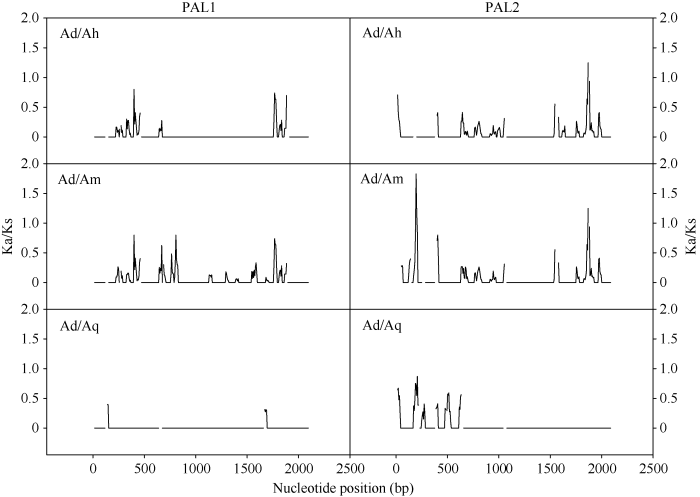 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}