 ,, 杜昕, 于高波, 金喜军, 张明聪, 任春元, 王孟雪,*, 张玉先,*黑龙江八一农垦大学农学院, 黑龙江大庆 163319
,, 杜昕, 于高波, 金喜军, 张明聪, 任春元, 王孟雪,*, 张玉先,*黑龙江八一农垦大学农学院, 黑龙江大庆 163319Regulation of carbon and nitrogen metabolism in leaf of soybean cultivar Suinong 26 at seed-filling stage under drought stress by exogenous melatonin
CAO Liang,, DU Xin, YU Gao-Bo, JIN Xi-Jun, ZHANG Ming-Cong, REN Chun-Yuan, WANG Meng-Xue,*, ZHANG Yu-Xian,*College of Agronomy, Heilongjiang Bayi Agricultural University, Daqing 163319, Heilongjiang, China通讯作者: * 张玉先, E-mail:zyx_lxy@126.com;王孟雪, E-mail:wangmengxue1978@163.com
收稿日期:2020-07-10接受日期:2021-01-21网络出版日期:2021-03-01
| 基金资助: |
Corresponding authors: * E-mail:zyx_lxy@126.com;E-mail:wangmengxue1978@163.com
Received:2020-07-10Accepted:2021-01-21Online:2021-03-01
| Fund supported: |
作者简介 About authors
E-mail:miss9877@126.com

摘要
鼓粒期是大豆碳氮代谢最复杂的阶段, 干旱胁迫必然限制鼓粒期大豆碳氮同化、分配和转移, 影响大豆产量的形成。在我们前期的研究中, 明确了外源褪黑素对干旱胁迫下鼓粒期大豆抗旱和碳氮代谢的生理调控效应。本研究通过转录组和代谢组分析来确定褪黑素对大豆干旱条件反应的一些重要的碳氮代谢基因和途径。转录组分析表明, 与干旱胁迫处理相比, 正常供水和干旱胁迫下喷施外源褪黑素处理的大豆叶片共同上调和下调的基因分别有37个和493个。上调的基因中存在着直接和间接参与碳氮代谢的功能基因, 包括正向调控的参与半胱氨酸合成、光合作用、碳水化合物代谢和葡萄糖代谢等途径关键基因。代谢组分析发现, 与干旱胁迫处理相比, 正常供水和干旱胁迫下喷施外源褪黑素处理的大豆叶片共同上调和下调的代谢物分别有17个和43个, 上调的代谢物中绝大部分(14/17)属于氨基酸、脂质、有机酸和碳水化合物, 进一步揭示了外源褪黑素能够提高大豆碳氮代谢与抗旱的能力。结合转录组和代谢组分析发现, 褪黑素通过调节氨基酸代谢和淀粉蔗糖代谢途径, 促进干旱胁迫下β-葡萄糖苷酶基因表达, 提高了L-天冬酰胺和6-磷酸葡萄糖代谢物的含量, 最终提高了大豆的抗旱性。
关键词:
Abstract
The grain-filling stage is the most complex stage of carbon and nitrogen metabolism. Drought stress inevitably inhibits the assimilation, distribution, and transition of carbon and nitrogen at grain-filling stage in soybean, resulting in less soybean yield. The objective of this study was to investigate the effects of exogenous melatonin on the carbon and nitrogen metabolism genes and pathways under drought stress in soybean. Transcriptome analysis showed that, compared with drought stress treatment, 37 and 493 genes were jointly up-regulated and down-regulated in soybean leaves treated with normal water supply and treated with exogenous melatonin under drought stress, respectively. The up-regulated genes included functional genes directly and indirectly involved in carbon and nitrogen metabolism, such as the key genes involved in the cysteine synthesis pathway, photosynthesis, carbohydrate metabolism, and glucose metabolism. Metabolomic analysis revealed that, compared with drought stress treatment, 17 and 43 metabolites were jointly up-regulated and down-regulated in soybean leaves treated with normal water supply and treated with exogenous melatonin under drought stress, respectively. Most (14/17) of up-regulated metabolites were amino acids, lipids, organic acids, and carbohydrates, which further indicated that exogenous melatonin could improve soybean carbon and nitrogen metabolism and drought resistance in soybean. Combined with transcriptome and metabolomic profile, melatonin promoted the relative expression level of β-D-Glucosidase gene due to regulate the pathway of amino acid metabolism and starch and sucrose metabolism, improved the contents of L-Asparagine and D-glucose-6P metabolites, and ultimately improves the ability of drought resistance in soybean.
Keywords:
PDF (6085KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
曹亮, 杜昕, 于高波, 金喜军, 张明聪, 任春元, 王孟雪, 张玉先. 外源褪黑素对干旱胁迫下绥农26大豆鼓粒期叶片碳氮代谢调控的途径分析. 作物学报[J], 2021, 47(9): 1779-1790 DOI:10.3724/SP.J.1006.2021.04151
CAO Liang, DU Xin, YU Gao-Bo, JIN Xi-Jun, ZHANG Ming-Cong, REN Chun-Yuan, WANG Meng-Xue, ZHANG Yu-Xian.
在全球气候变化的大环境背景下, 黑龙江作为我国大豆主要产区受到天气因素影响日益加剧, 干旱是影响黑龙江大豆产业最主要的环境因素[1,2]。根据全球降雨量和温度变化长期数据统计, 干旱胁迫发生的频率和强度每年都在增加, 由此产生的干旱地区也逐年增多[3,4]。已有研究表明, 当大豆遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降, 叶绿体结构被破坏, 造成光合损伤, 并最终抑制大豆的生长发育[5,6]。鼓粒期是碳氮代谢最旺盛、最复杂的阶段, 而干旱胁迫必然限制鼓粒期大豆生长, 影响鼓粒期大豆的碳氮同化, 改变碳氮积累、分配和转移规律[7]。
褪黑素在植物生长、发育和应激反应中是一种多样化的调节因子[8,9,10]。褪黑素能够增强干旱胁迫条件下大豆幼苗抗氧化酶活性, 减少ROS的过度积累, 提高渗透调节物质含量, 进而降低膜脂过氧化程度[11]; 可以减轻干旱胁迫对大豆光合系统造成的损伤, 提高光合特性和叶绿素荧光参数、最终促进大豆的生长[12]。进一步的转录组分析表明, 褪黑素通过调控光合作用、细胞循环、DNA复制、淀粉蔗糖代谢和脂质生物合成来发挥其功能[13]。虽然大量研究已经证实褪黑素能够改善干旱胁迫下大豆生长发育[14], 但大多研究仅从提高抗氧化胁迫能力、改善光合同化和分配的角度分析褪黑素的积极作用, 缺少从调节碳氮代谢角度深度解析干旱胁迫下褪黑素促进大豆发育的分子机制。
本研究通过转录组与代谢组联合分析外源褪黑素调控干旱胁迫下鼓粒期大豆碳氮代谢的可能调控机制, 以期为进一步鉴定其重要的相关基因和蛋白功能、利用基因工程进行遗传改良提供理论基础。
1 材料与方法
1.1 试验材料和设计
以干旱敏感型大豆品种绥农26为供试品种[12], 盆栽用桶直径30.0 cm, 高33.0 cm。为有效控制土壤含水量的同时避免土壤板结, 采用蛭石、珍珠岩和黑钙土按照1:3:12 (v/v/v)混合作为基质, 混合培养土含碱解氮72.1 mg kg-1、速效磷14.1 mg kg-1、速效钾173.0 mg kg-1、有机质3.9 mg kg-1, pH 7.1。施肥量为磷酸二铵300 kg hm-2、尿素75 kg hm-2、硫酸钾75 kg hm-2。播种前准备基质过程中, 测量每盆基质的质量和含水量, 计算出基质干重。并通过环刀法测量最大持水量, 以便后续控水过程中准确掌握田间持水量。试验于2018—2019年在国家杂粮工程技术研究中心(大庆)进行。为了避免雨水淋浇, 所有盆栽均放置在遮雨棚中。大豆种子采用75%的酒精处理2 min, 再用5%的次氯酸钠进行消毒5 min, 用无菌水洗净后播种。每盆播种6粒种子, 均匀排布。待子叶期(Cotyledon, VC)定苗3株, 去除过大或过小的幼苗, 保留长势适中的幼苗。
自播种至鼓粒始期, 采用称重法控制土壤含水量为田间持水量的80%。鼓粒始期记为第1天, 正常供水处理仍保持土壤含水量为田间持水量的80%, 干旱处理通过停止供水逐步使土壤含水量达到田间持水量的50%, 并维持此含水量至处理结束, 凋萎含水量为田间持水量的40%。每个处理100盆, 具体处理设置如下:
正常供水处理(WW), 保持土壤含水量为田间持水量的80%;
干旱胁迫处理(D), 停止供水并于每天18:00称重, 第10天达到田间持水量的50%, 而后补水以维持此含水量;
干旱胁迫喷施褪黑素处理(DM), 在干旱胁迫处理的基础上, 在第11、12、13天夜晚21点喷施浓度为100 μmol L -1褪黑素, 褪黑素见光易分解, 因此选择夜间喷施[15]。
用于转录组和代谢组检测的样品于褪黑素处理后第3天(即鼓粒初期后第16天)取样, 取大豆功能叶片(倒三叶), 液氮冷冻后快速转移至-80℃冰箱保存待分析用。
1.2 测定项目与方法
1.2.1 总RNA分离和转录组分析 本研究使用EASYspin Plus试剂盒, 按照说明提取在正常供水(WW)、干旱胁迫(D)和干旱胁迫下同时进行外源褪黑素处理(DM) 3种条件下, 3个生物学重复的大豆叶片RNA。制备RNA-Seq文库并测序。测序数据已提交至NCBI (1.2.2 代谢产物检测和数据分析 冷冻叶片用氧化锆珠磨样: 1.5 min, 30 Hz。将100 mg样品在4℃下用1 mL 70%的甲醇水溶液, 离心10,000×g, 10 min后, 吸收提取物并过滤, 后进行高效液相色谱—质谱分析。利用三重四极线性离子阱质谱仪的多反应监测分析, 利用API 4500 Q trap LC/MS/MS系统对代谢物进行定量。分析条件和详细的操作参数是按照之前的方法完成的。为了进行统计分析, 在分析前对各代谢物的相对峰度进行对数变换, 使其达到正态分布。使用R对规范化数据进行主成分分析。在正交偏最小二乘判别分析(OPLS-DA)的基础上, 根据不同样品中代谢物含量的倍数关系大于2选择差异代谢物(项目中的可变重要度, VIP≥1)。与转录组结果一致, 通过SoyBase (
1.2.3 实时荧光定量qRT-PCR验证 为进一步证实本研究的转录组结果, 使用实时荧光定量PCR(qRT-PCR)分析了所有样品。按照说明书, 使用PrimeScript RT试剂盒, 对转录组分析的RNA样本进行反转录。根据说明, 使用TB Green Premix Ex TaqII在实时PCR系统上执行qRT-PCR程序。特异基因引物序列列于附表1。采用2-ΔΔCt法选择基因的相对表达水平, 大豆基因GAPDH作为内参。
2 结果与分析
2.1 外源褪黑素诱导干旱胁迫下大豆叶片差异表达基因的初步分析
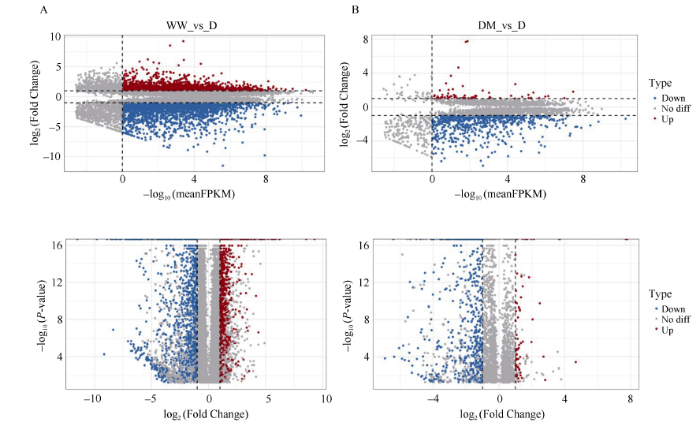
以干旱胁迫(D)为基准, 通过不同条件下大豆叶片转录水平的两两比较, 分析正常供水(WW)与干旱胁迫(D), 和干旱胁迫下同时进行褪黑素处理(DM)与干旱胁迫(D)之间的基因表达差异, 进而揭示外源褪黑素提高大豆抗旱能力的潜在机制。依据经验值, 本研究仅选择所有样品的平均表达值FPKM (每一百万个比对上的序列中能够比对到外显子的每一千碱基上的双端序列的个数)大于1, FDR小于0.05的表达基因。由图1可知, 与干旱胁迫(D)处理相比, 正常供水(WW)处理的大豆叶片有1604个基因上调, 2382个基因下调, 干旱胁迫下同时进行褪黑素处理(DM)的大豆叶片有68个基因上调, 600个基因下调。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1WW/D和DM/D比较中差异表达基因图
灰点代表无明显差异的表达基因, 红色和蓝色点分别表示在WW/D和DM/D比较中显著上调和显著下调的基因。WW为正常供水处理, D为干旱处理, DM为干旱条件下喷施褪黑素处理。
Fig. 1Comparisons of differentially expressed genes in WW/D and DM/D
Grey dots represent genes without significant differential expressions, red and blue dots denote significantly up-regulated and down-regulated genes in the WW/D and DM/D comparisons, respectively. WW is the normal water supply group, D is the drought stress group, and DM is the foliar application of melatonin under drought stress group.
为筛选出外源褪黑素提高大豆叶片抗旱能力的相关基因, 首先构建了WW/D与DM/D的韦恩图。与干旱胁迫(D)处理相比, 正常供水(WW)和干旱胁迫下同时进行褪黑素处理(DM)的大豆叶片共同上调和下调的基因分别有37个和493个(图2-A)。进一步对这530个差异表达基因进行无监督聚类分析(图2-B)发现, 与干旱胁迫(D)处理相比, 正常供水处理(WW)和干旱胁迫下同时进行褪黑素处理(DM)的大豆叶片转录组呈现相同的上调和下调趋势, 表明外源褪黑素对大豆叶片抗旱能力的提高与上述差异表达的基因相关。
图2
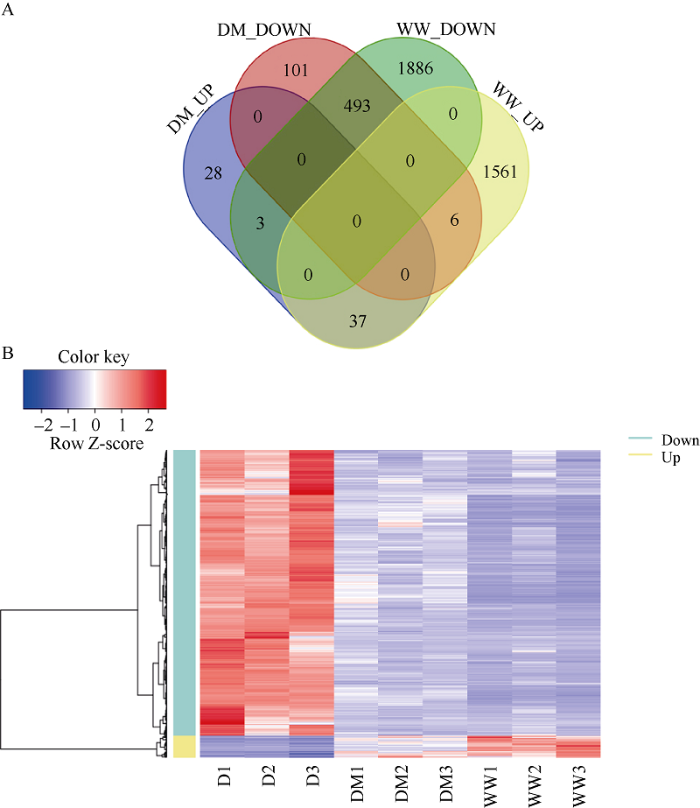 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2外源褪黑素对干旱胁迫下大豆叶片转录组的影响
A: 韦恩图显示了WW/D和DM/D比较之间的共同差异表达基因。B: WW/D和DM/D共同上调和下调表达的差异基因的热图。处理同
Fig. 2Effects of exogenous melatonin on leave transcriptome under drought stress in soybean
A: Venn diagram of the differentially expressed genes between WW/D and DM/D. B: the heat maps of the common differentially expressed genes between WW/D and DM/D. Treatments are the same as those given in
2.2 外源褪黑素诱导干旱胁迫下大豆叶片差异基因的富集分析
为进一步证明上调基因参与了大豆叶片碳氮代谢过程, 基于P值小于0.05对37个共同上调的基因进行GO富集结果的筛选, 主要包括丝氨酸合成代谢过程、含硫氨基酸合成代谢过程、戊糖代谢调控, RNA聚合酶Ⅱ启动子的调控过程, 发现这些上调的基因在生物学过程的P值更小, 进一步表明外源褪黑素能正向调控大豆叶片中碳代谢和氨基酸代谢过程。图3
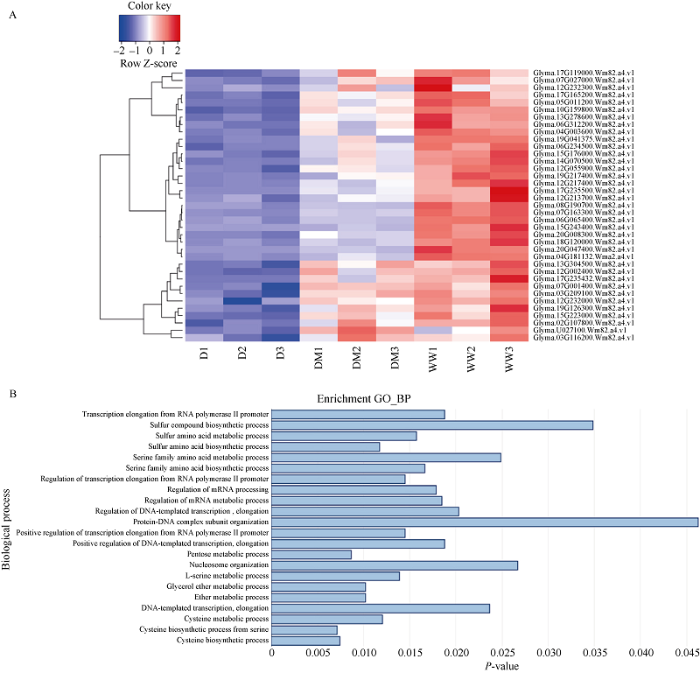 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3外源褪黑素上调干旱胁迫下大豆叶片基因的生物学过程富集分析
A: 热图显示了WW/D和DM/D比较之间的共同表达上调的基因。B: WW/D和DM/D比较之间的共同表达上调基因GO生物学过程富集分析。处理同
Fig. 3GO biological process enrichment of up-regulated differentially expressed genes under melatonin treatment in WW/D and DM/D
A: the heatmap of the up-regulated expressed genes between the WW/D and DM/D. B: GO biological process enrichment on common up-regulated expressed genes between the WW/D and DM/D. Treatments are the same as those given in
对这些候选基因进行KEGG代谢通路分析(图4)发现, 富集的通路上调的基因也主要集中在碳氮代谢方面, 包括淀粉蔗糖代谢、氰胺酸代谢、半胱氨酸和蛋氨酸代谢, 下调的基因主要包括脱落酸的生物合成、脂质氧化、活性氧产生和叶片衰老等相关基因, 表明褪黑素可以提高干旱胁迫下大豆碳氮代谢能力, 抑制脱落酸的合成和活性氧的产生, 延缓大豆叶片衰老, 进而表明这些基因能够从调节碳氮代谢角度深度解析干旱胁迫下褪黑素促进大豆发育和产量提高的机制。
图4
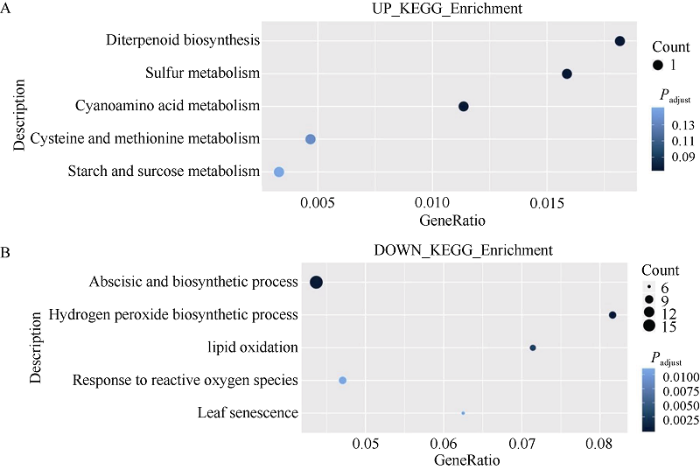 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4WW/D和DM/D共同上调与下调基因的KEGG富集图
A: 上调。B: 下调。富集因子的计算方法是: 差异表达基因数除以任何给定途径的基因数, 点的大小表示基因数, 点的颜色表示调整后的P值范围。处理同
Fig. 4KEGG pathway enrichment for differentially up-regulated and down-regulated expressed genes both in WW/D and DM/D
A: up-regulated. B: down-regulated. The rich factor is calculated as the differentially expressed genes number divided by the base number of any given pathway. Dot size denotes the number of genes and dot color denotes the range of adjusted P-value. Treatments are the same as those given in
2.3 关键差异表达基因的定量验证分析
利用qRT-PCR方法对转录组数据分析结果进行验证(图5)。随机选择10个差异表达基因, 包括参与光合作用(Glyma.19G261400、Glyma.18G120000)、碳水化合物和糖代谢(Glyma.U027100、Glyma.13G 278600)、半胱氨酸代谢(Glyma.10G159800)、脱落酸合成(Glyma.11G180800)、叶片衰老(Glyma.06G 082400)、脂质氧化(Glyma.13G030300)、过氧化氢合成(Glyma.01G031500)、活性氧生成(Glyma. 12G098900)。qRT-PCR结果分析表明所挑选的10个基因的表达水平变化趋势与转录组测序分析结果一致, 表明该转录组数据结果真实可信。图5
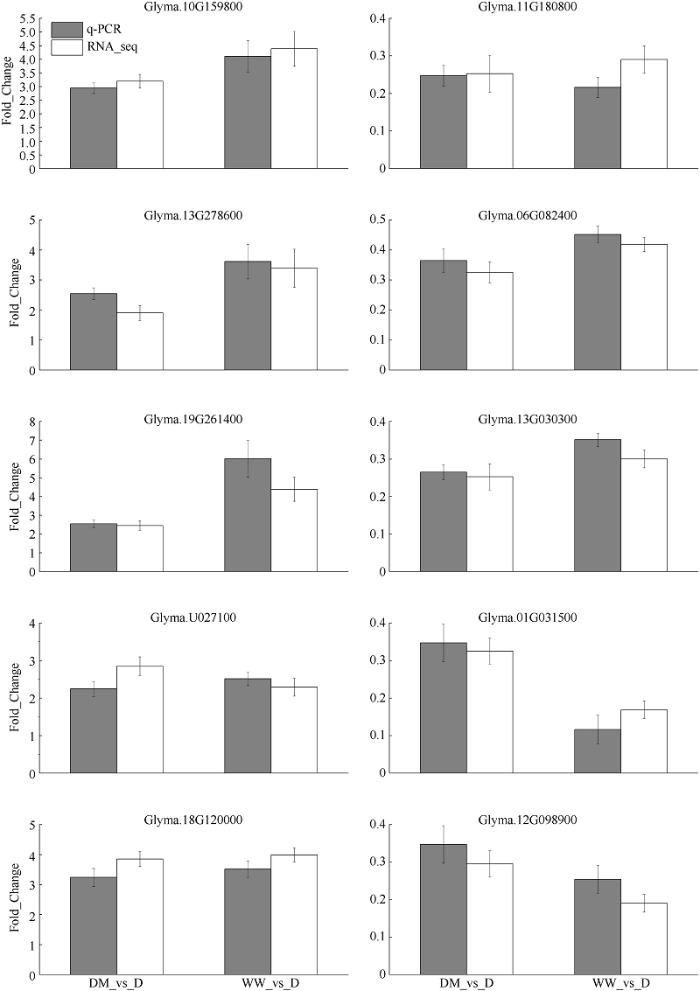 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5差异表达基因的 RT-qPCR 验证
白色的表示差异表达基因的转录组数据, 黑色的表示差异表达基因的qRT-PCR结果。处理同
Fig. 5RT-PCR validation of relative expression levels of DEGs
White bar signifies transcriptomic data of the differentially expressed genes, and black bar denotes qRT-PCR results of the differentially expressed genes. Treatments are the same as those given in
2.4 外源褪黑素诱导干旱胁迫下大豆叶片差异代谢物和KEGG分析
为研究干旱胁迫下大豆叶片代谢产物的变化以及外源褪黑素对干旱胁迫下大豆叶片代谢产物的影响, 首先对不同条件下(包括正常供水WW、干旱胁迫D和干旱胁迫下同时进行外源褪黑素处理DM), 3个生物学重复的大豆叶片代谢产物的表达进行主成分分析, 发现生物学重复的样品聚集在一起, 并且各组间差异显著, 表明了数据的可靠性。此外, WW处理与DM处理之间的距离比WW处理与D处理之间的距离更近(图6), 表明外源褪黑素能够通过大豆叶片的代谢产物变化而提高其抗旱能力。图6
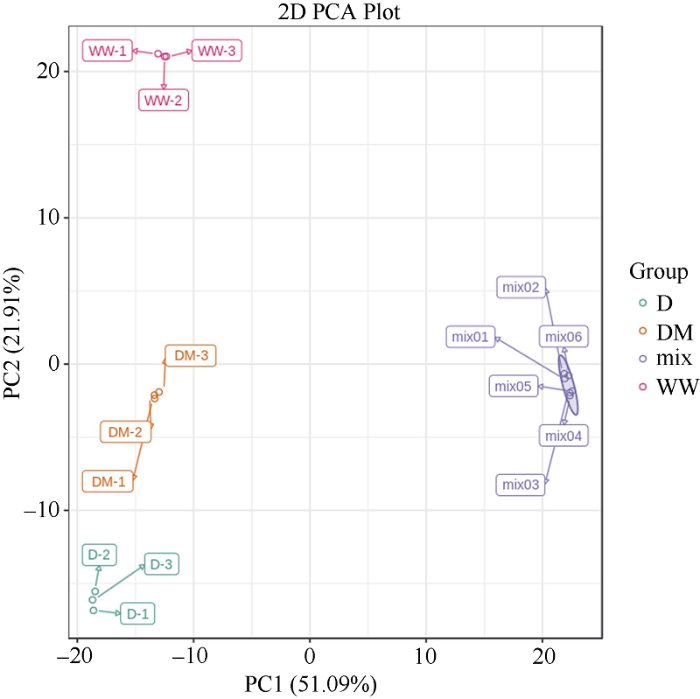 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6代谢组主成分分析
处理同
Fig. 6PCA analysis of metabolomics
Treatments are the same as those given in
考虑到存在试验过程引入的偏好性, 仅考虑在生物学重复样品之间的代谢产物含量比例小于2的化合物, 因此, 共检测到稳定表达的613种化合物, 其中与干旱胁迫(D)组相比, 正常供水(WW)和干旱胁迫下同时进行褪黑素处理(DM)的大豆叶片共同上调和下调的代谢物分别有17个和43个(图7-A)。即使下调的代谢物主要为黄酮类化合物, 包括柚皮素、鹰嘴豆芽素A、芒柄花黄素、L-2-氨基己二酸、山柰酚、毛蕊异黄酮、β-硅酸盐、柚配基查尔酮、紫铆花素、甘草素、异甘草素、3,4’,7-三羟基黄烷酮、吡啶甲酸、紫苜蓿素等等, 但是本研究发现上调的代谢物中绝大部分(14/17)属于氨基酸、脂质代谢、有机酸和碳水化合物, 主要包括L-谷氨酸、D-葡萄糖6-磷酸、生物素、L-精氨酸、L-天冬酰胺、尿苷酸、4-鸟苷丁酸酯等, 表明外源褪黑素处理能够一定程度的提高大豆叶片碳氮代谢水平和抗旱能力。为验证这个结果的可靠性, 利用KEGG Mapper对检测到正常供水(WW)和干旱胁迫下同时进行褪黑素处理(DM)的大豆叶片共同上调和下调代谢物所参与的代谢通路进行分析(图7-B)发现, 这些上调代谢物直接和间接参与的通路集中在碳氮代谢方面, 进一步证明了外源褪黑素能够通过改变大豆叶片中代谢物的表达而提高其抗旱能力。
图7
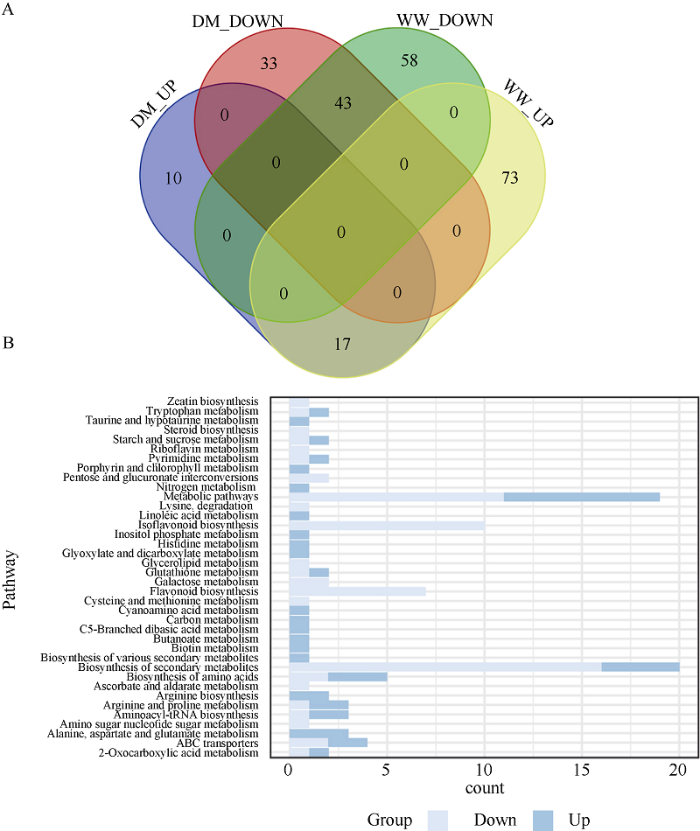 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7干旱胁迫和外源褪黑素处理对代谢组的影响
A: 韦恩图显示了WW/D和DM/D比较中共同的差异累积代谢物。B: WW/D和DM/D共同上调和下调基因的KEGG富集图。横坐标是差异表达基因数。处理同
Fig. 7Effects of drought stress and exogenous melatonin treatment on the metabolome in soybean
A: the venn diagram shows the overlapped differentially-accumulated metabolites between the WW/D and DM/D comparisons. B: the histogram of KEGG pathway enrichment for differentially-accumulated metabolites both in WW/D and DM/D comparison. X-axis is the number of differentially expressed genes. Treatments are the same as those given in
2.5 关键代谢通路的转录组和代谢组联合分析
为了更清楚了解外源褪黑素对干旱条件下大豆叶片抗旱能力的生理变化, 本研究针对干旱(D)处理, 正常供水(WW)处理和干旱胁迫下外源褪黑素(DM)处理大豆叶片中上调的转录组和代谢组进行联合分析。如图4-A和图8所示, 上调的转录组和代谢组所参与的通路均包括淀粉和蔗糖代谢、氨基酸的生物合成等。为了进一步具体了解其参与的代谢通路, 通过整合上述筛选出上调的基因和代谢产物发现, 它们同时参与了淀粉、蔗糖代谢和氨基酸代谢, 包括酪氨酸代谢、缬氨酸代谢、亮氨酸代谢、异亮氨酸代谢和苯丙氨酸代谢。其中, 主要是通过β-葡萄糖苷酶基因表达的增加, 最终提高了L-天冬酰胺和6-磷酸葡萄糖代谢物的含量。转录组和代谢组联合分析表明, 外源褪黑素通过增强β-葡萄糖苷酶基因的表达, 促进了干旱胁迫下大豆碳氮代谢进程, 增加了L-天冬酰胺和6-磷酸葡萄糖等碳氮代谢化合物的积累, 进而提高大豆的抗旱性。图8
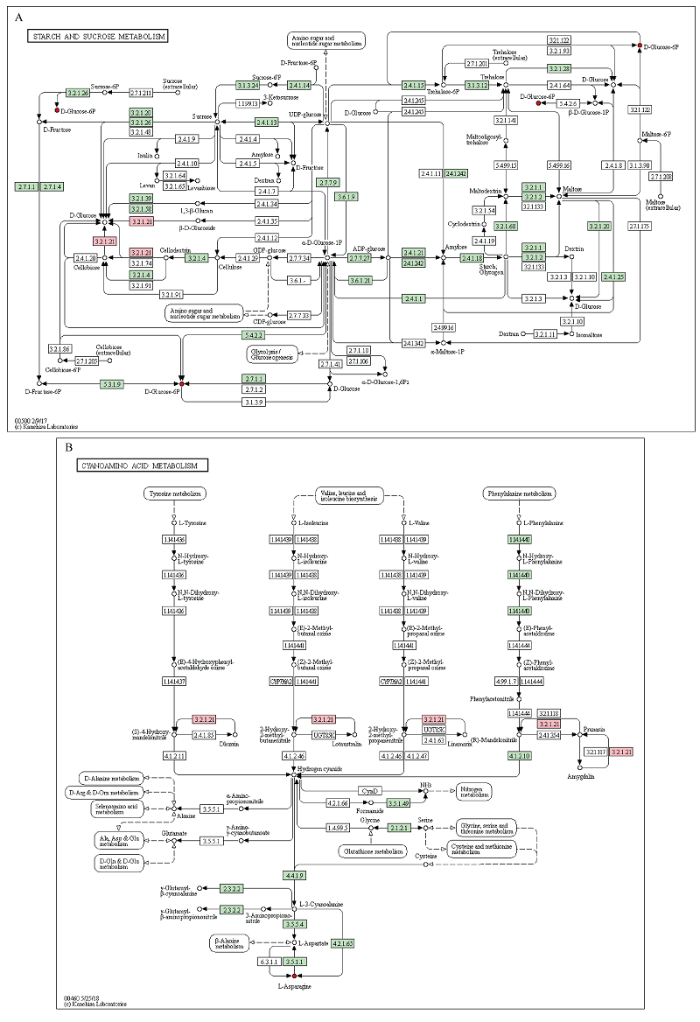 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8淀粉蔗糖代谢和L-天冬酰胺代谢的KEGG途径
A: 淀粉和蔗糖代谢。B: L-天冬酰胺代谢。红色和粉色分别表示与干旱胁迫处理相比, 正常供水处理和干旱胁迫下喷施外源褪黑素大豆叶片中共同上调的代谢产物和基因。
Fig. 8KEGG pathway of starch sucrose metabolism and cyanoamino acid metabolism
A: starch and sucrose metabolism. B: cyanoamino acid metabolism. Red and pink denote up-regulated metabolites and genes both in WW/D and DM/D.
3 讨论
课题组之前的研究已经证明, 外源褪黑素处理可以提高干旱胁迫下大豆的碳氮代谢能力[16], 然而, 褪黑素缓解干旱胁迫的具体机制尚不清楚。因此, 本研究利用转录组和代谢组的方法研究了褪黑素在大豆碳氮代谢及抗旱能力中的作用, 从而确定了与大豆碳氮代谢能力相关的候选基因和关键代谢途径。在转录组分析中, 用WW/D和DM/D 2种方法对差异表达基因进行了鉴定, 这一试验设计使我们能够鉴定出在干旱胁迫下褪黑素改变的差异表达基因的表达水平。在WW/D比较中, 共有3986个差异表达基因, 其中1604个基因上调, 2382个基因下调, 而干旱胁迫下同时进行褪黑素处理(DM)的大豆叶片相比于干旱条件下(D)有68个基因上调, 600个基因下调。进一步整合WW/D和DM/D两组差异表达的结果发现, 共同上调和下调分别有37个和493个基因。倪知游[17]研究发现, 褪黑素处理上调了干旱胁迫下猕猴桃幼苗光合作用、碳代谢、抗坏血酸代谢等相关基因。李超[18]研究发现, 褪黑素处理干旱胁迫下苹果幼苗氮代谢和光合作用等相关基因显著上调。本研究中GO富集结果表明, 一些差异表达的基因能够直接和间接的参与碳氮代谢的过程, 比如半胱氨酸合成基因、光合相关基因和β-葡萄糖苷酶等, 这与前人的研究结果基本一致, 表明褪黑素的确能够从碳氮代谢方面改善大豆抗旱能力。魏志为[19]研究表明, 褪黑素可以通过调节相关基因的表达, 促进逆境胁迫下苹果叶片中葡萄糖、淀粉、氨基酸代谢物的积累, 在本研究中, 通过代谢组分析表明, 与干旱胁迫(D)相比, 共同被正常供水处理(WW)和干旱胁迫条件下外源褪黑素处理(DM)大豆叶片的17个上调和43个下调代谢产物, 17个上调的代谢物中包括氨基酸、脂质、有机酸和碳水化合物, 进一步验证了外源褪黑素能够提高大豆的碳氮代谢能力。
为探索褪黑素通过哪些基因和代谢途径来提高大豆叶片在干旱胁迫条件下的抗旱能力, 本研究整合转录组和代谢组上调的37个基因和17个代谢化合物进行KEGG分析发现, 同时存在差异表达基因和代谢化合物的有2个代谢途径, 即淀粉蔗糖代谢和氰基氨基酸代谢。糖是调节逆境诱导基因表达的关键信号传递物质, 糖除了行使自身代谢功能外, 还在分子水平上调控多种生理和发育过程, 包括光合作用、同化物的储存和淀粉的动员等[20]。植物发生非生物胁迫时会导致糖的含量发生显著的变化, 可以影响许多基因的表达量, 例如在拟南芥中发现了干旱胁迫下调了参与碳水化合物代谢的基因表达, 包括蔗糖磷酸合成酶、蔗糖合成酶、甘油醛-3-磷酸脱氢酶、磷酸烯醇丙酮酸羧化酶、β-葡萄糖苷酶等相关基因表达, 因此碳水化合物在植物生长发育过程中起着至关重要的作用[21]。本研究结果表明, 干旱胁迫导致大豆叶片碳水化合物、糖代谢和光合作用相关基因下调, 干旱胁迫条件下喷施外源褪黑素可促进上调相关基因表达, 其中包括β-葡萄糖苷酶表达的增加, 参与光合作用的(Glyma.19G261400)上调, 以及6-磷酸葡萄糖代谢物的含量的增加, 进而增强大豆的抗旱能力。这与Wei等[22]研究褪黑素可以促进非生物胁迫条件大豆下光合作用过程中的光反应和暗反应相关基因上调, 改善逆境胁迫下大豆的光合相关过程, 可以促进蔗糖和葡萄糖相关合成基因表达上调的研究结果一致。氨基酸可以作为活性氧清除剂和潜在的调节分子, 对提高植物的抗逆能力至关重要, 特别是亮氨酸和异亮氨酸可通过植物呼吸系统来调节植物的抗逆性[23,24]。色氨酸、苯丙氨酸和酪氨酸(芳香族氨基酸)作为重要的细胞成分, 参与植物的发育和环境适应[25]。半胱氨酸是必需氨基酸和生糖氨基酸, 植物生长过程中可由植物体内的蛋氨酸转化而来, 可与胱氨酸互相转化, 可以延缓植物衰老, 减轻非生物胁迫对作物的氧化损伤[26]。本研究发现, 干旱胁迫导致大豆叶片氨基酸相关合成基因下调, 氨基酸代谢产物下降, 此外, 尽管检测到受褪黑素影响的上调代谢化合物只有17个, 但是发现它们还参与其他的氨基酸代谢过程, 比如丙氨酸、天冬氨酸和谷氨酸代谢、半胱氨酸和甲硫氨酸代谢等, 提高了大豆的抗旱能力及氮代谢能力, 减少了干旱胁迫对大豆植株造成的氧化损伤。这与Verslues等[27]和Antoniou等[28]研究发现氨基酸可能是一种自由基清除剂, 褪黑素可以通过调节氨基酸代谢系统地改善苜蓿植株干旱胁迫诱导的损伤相一致。
4 结论
转录组和代谢组联合分析发现, 褪黑素通过调节“氨基酸代谢”和“淀粉蔗糖代谢”途径, 促进干旱胁迫下β-葡萄糖苷酶基因表达增加, 提高了L-天冬酰胺和6-磷酸葡萄糖代谢物的含量, 最终提高了大豆的抗旱性。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIPMID [本文引用: 1]

Global climate change is predicted to increase temperatures, alter geographical patterns of rainfall and increase the frequency of extreme climatic events. Such changes are likely to alter the timing and magnitude of drought stresses experienced by crops. This study used new developments in the classification of crop water stress to first characterize the typology and frequency of drought-stress patterns experienced by European maize crops and their associated distributions of grain yield, and second determine the influence of the breeding traits anthesis-silking synchrony, maturity and kernel number on yield in different drought-stress scenarios, under current and future climates. Under historical conditions, a low-stress scenario occurred most frequently (ca. 40%), and three other stress types exposing crops to late-season stresses each occurred in ca. 20% of cases. A key revelation shown was that the four patterns will also be the most dominant stress patterns under 2050 conditions. Future frequencies of low drought stress were reduced by ca. 15%, and those of severe water deficit during grain filling increased from 18% to 25%. Despite this, effects of elevated CO2 on crop growth moderated detrimental effects of climate change on yield. Increasing anthesis-silking synchrony had the greatest effect on yield in low drought-stress seasonal patterns, whereas earlier maturity had the greatest effect in crops exposed to severe early-terminal drought stress. Segregating drought-stress patterns into key groups allowed greater insight into the effects of trait perturbation on crop yield under different weather conditions. We demonstrate that for crops exposed to the same drought-stress pattern, trait perturbation under current climates will have a similar impact on yield as that expected in future, even though the frequencies of severe drought stress will increase in future. These results have important ramifications for breeding of maize and have implications for studies examining genetic and physiological crop responses to environmental stresses. © 2013 John Wiley & Sons Ltd.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIPMID [本文引用: 1]
In recent years melatonin has emerged as a research highlight in plant studies. Melatonin has different functions in many aspects of plant growth and development. The most frequently mentioned functions of melatonin are related to abiotic stresses such as drought, radiation, extreme temperature, and chemical stresses. This review mainly focuses on the regulatory effects of melatonin when plants face harsh environmental conditions. Evidence indicates that environmental stress can increase the level of endogenous melatonin in plants. Overexpression of the melatonin biosynthetic genes elevates melatonin levels in transgenic plants. The transgenic plants show enhanced tolerance to abiotic stresses. Exogenously applied melatonin can also improve the ability of plants to tolerate abiotic stresses. The mechanisms by which melatonin alleviates abiotic stresses are discussed. © The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI [本文引用: 1]
We studied the effects of melatonin on soybean photosynthesis and seed growth at different developmental stages under drought stress induced by polyethylene glycol in the growth medium. Experimental groups were supplied with a normal nutrient solution after seven days of continuous treatment; overall, five treatments were completed when the early maturity stage was reached. Under normal water supply, melatonin supplementation had no significant effect on the photosynthetic characteristics or soybean seed growth. However, drought stress reduced leaf relative water content, chlorophyll content, and relative photosynthetic gas-exchange parameters, while it increased a leaf content of osmoregulatory substances, antioxidant-related enzyme activities, and seed growth-related indicators. Compared to the drought-stressed group, the melatonin treatment under drought stress improved the above indicators; specifically, melatonin effectively increased the photosynthetic rate, 100-seed mass, and total seed mass per plant. All the effects were most significant during the first seven days after the early seed-filling stage.
DOIURL [本文引用: 1]
DOI [本文引用: 1]
In order to investigate the role of melatonin in the drought tolerance, we examined pigments, gas exchange, osmolytes, and reactive oxygen radical metabolism in soybean plants. Drought declined photosynthetic pigments and caused irreversible reduction in net photosynthesis, which was followed by stomatal limitation for 5 and 10 d and nonstomatal limitation for 15 d. Soluble sugar, soluble proteins, and proline concentrations were higher in drought-stressed seedlings compared with the control. The contents of superoxide anion, hydrogen peroxide, and malondialdehyde increased during drought stress indicating oxidative stress. Drought stress also increased superoxide dismutase, peroxidase, and catalase activities. Melatonin treatment improved the tolerance of drought-treated plants, which was possibly due to the enhanced content of osmolytes and higher antioxidant enzyme activities that retard dehydration and lipid peroxidation.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIPMID [本文引用: 1]
Melatonin is a well-known agent that plays multiple roles in animals. Its possible function in plants is less clear. In the present study, we tested the effect of melatonin (N-acetyl-5-methoxytryptamine) on soybean growth and development. Coating seeds with melatonin significantly promoted soybean growth as judged from leaf size and plant height. This enhancement was also observed in soybean production and their fatty acid content. Melatonin increased pod number and seed number, but not 100-seed weight. Melatonin also improved soybean tolerance to salt and drought stresses. Transcriptome analysis revealed that salt stress inhibited expressions of genes related to binding, oxidoreductase activity/process, and secondary metabolic processes. Melatonin up-regulated expressions of the genes inhibited by salt stress, and hence alleviated the inhibitory effects of salt stress on gene expressions. Further detailed analysis of the affected pathways documents that melatonin probably achieved its promotional roles in soybean through enhancement of genes involved in cell division, photosynthesis, carbohydrate metabolism, fatty acid biosynthesis, and ascorbate metabolism. Our results demonstrate that melatonin has significant potential for improvement of soybean growth and seed production. Further study should uncover more about the molecular mechanisms of melatonin's function in soybeans and other crops. © The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
DOIPMID [本文引用: 1]
Amino acids have various prominent functions in plants. Besides their usage during protein biosynthesis, they also represent building blocks for several other biosynthesis pathways and play pivotal roles during signaling processes as well as in plant stress response. In general, pool sizes of the 20 amino acids differ strongly and change dynamically depending on the developmental and physiological state of the plant cell. Besides amino acid biosynthesis, which has already been investigated in great detail, the catabolism of amino acids is of central importance for adjusting their pool sizes but so far has drawn much less attention. The degradation of amino acids can also contribute substantially to the energy state of plant cells under certain physiological conditions, e.g. carbon starvation. In this review, we discuss the biological role of amino acid catabolism and summarize current knowledge on amino acid degradation pathways and their regulation in the context of plant cell physiology. Copyright © 2015 The Author. Published by Elsevier Inc. All rights reserved.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}