 ,, 李圆圆, 陆俊杏, 张涛,*重庆师范大学生命科学学院, 重庆 401331
,, 李圆圆, 陆俊杏, 张涛,*重庆师范大学生命科学学院, 重庆 401331Morphological characteristics and cytological study of anther abortion of temperature-sensitive nuclear male sterile line 160S in Brassica napus
TANG Xin,, LI Yuan-Yuan, LU Jun-Xing, ZHANG Tao,*College of Life Sciences, Chongqing Normal University, Chongqing 401331, China通讯作者:
收稿日期:2020-09-9接受日期:2020-11-13网络出版日期:2021-05-12
| 基金资助: |
Received:2020-09-9Accepted:2020-11-13Online:2021-05-12
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (17790KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
唐鑫, 李圆圆, 陆俊杏, 张涛. 甘蓝型油菜温敏细胞核雄性不育系160S花药败育的形态学特征和细胞学研究[J]. 作物学报, 2021, 47(5): 983-990. doi:10.3724/SP.J.1006.2021.04207
TANG Xin, LI Yuan-Yuan, LU Jun-Xing, ZHANG Tao.
油菜作为世界上重要的油料作物之一, 是食用油和生物燃料的重要来源[1]。为了满足日益增长的需求, 提高油菜产量是育种工作急需解决的问题和长期目标。雄性不育植物是杂交育种所必需的育种材料, 能利用杂种优势来提高作物产量, 在农业生产上具有重要的经济价值[2,3]。
细胞质雄性不育(cytoplasmic male sterility, CMS)和细胞核雄性不育(genic male sterility, GMS)已被广泛应用于油菜杂种优势的开发与利用, 其中, GMS类型主要分为受光周期影响的光敏型雄性不育(photoperiod-sensitive genic male sterility, PGMS)和受温度控制的温敏型雄性不育(thermo-sensitive genic male sterility, TGMS)[4,5]。由于大多数P/TGMS由隐形核基因控制, 雄性不育基因容易遗传, 可以用作父本, 具有恢复源广、细胞质来源丰富等优点, 被认为是两系杂交育种的高效系统[6]。目前, 已发现的甘蓝型油菜PGMS类型主要有H90S、N196S和501-8S; TGMS类型主要有Xiangyou 91S、Xiangyou 402S、Huiyou 50S、373S、SP2S、104S、TE5A等[7]。自首次报道水稻光敏不育系58S, 并成功用于水稻两系杂交育种以来[8], 在禾本科[9,10,11]、豆科[12]和十字花科[13]等其他植物中也发现了光温敏雄性不育系材料。近年来, 甘蓝型油菜雄性不育材料花药的细胞学特征显微观察已得到广泛的关注。Sun等[7]研究发现, 373S花粉败育是由于小孢子在发育过程中出现细胞质浓缩、胞外壁不规则、单核后期绒毡层降解异常引起的。李可琪等[14]和Yan等[15]观察TE5A的花药切片发现, 在减数分裂期花粉母细胞同源染色体不能配对、凝聚形成二分体和四分体, 导致小孢子发育受阻, 最终引起败育。Yu等[16]对SP2S细胞学观察发现, 绒毡层细胞在减数分裂早期高度空泡化变大, 小孢子无法从四分体释放出来, 并伴随胼胝质降解异常, 最终导致败育。总之, 植物雄性不育是一个极其复杂的过程, 败育时期、方式多种多样, 败育特征和程度也各不相同。
前期研究表明[17], 160S育性受环境温度影响, 20℃以下可育(160S-MF), 25℃以上不育(160S-MS); 温度是影响育性的关键因素, 随着温度的升高, 花粉活力、角粒数和结角率随之降低; 研究还表明, 160S具有恢复源广泛、较强的一般配合力、杂种优势明显等优点, 可以应用培育杂交油菜新品种。前人仅仅开展了温度对不育系160S育性转化和杂种优势等方面的研究, 但其败育时期、原因和细胞学特征尚未报道。本研究通过观察甘蓝型油菜160S-MF和160S-MS形态学和细胞学差异, 明确160S-MS败育的时期和细胞学特征, 初步解析其败育原因, 旨在为深入研究其雄性不育内在分子调控机理以及在杂种优势的利用等方面奠定基础, 同时对甘蓝型油菜两系杂交育种具有指导作用。
1 材料与方法
1.1 材料
甘蓝型油菜温敏细胞核雄性不育系160S种子由重庆师范大学油用牡丹种质资源创新与利用重点实验室保存。在室温(20℃)条件下播种, 10 d后将长势良好的幼苗移栽至直径20 cm的花盆中, 一盆一株苗, 春化处理后, 分别放置在温度为15℃和28℃的冷光源植物生长箱(赛福ZRX-1100E)中培养, 光照16 h/黑暗8 h, 湿度50%。1.2 取材与固定
在初花期, 取160S-MF、160S-MS不同大小花蕾的花药压片, 用1%醋酸洋红染色, 在荧光显微镜(SOPTOP RX50)下观察小孢子发育时期。采样后, 将样品分组放入FAA固定液(50%酒精90 mL、冰醋酸5 mL、37%甲醛5 mL)中, 真空干燥箱(SKK ZK-50)抽真空, 直至花蕾完全沉入瓶底, 室温固定24 h, 70%乙醇4℃冰箱存放备用。1.3 花器形态特征观察
分别采集160S-MF、160S-MS盛花期的30朵花朵, 用游标卡尺测量花冠径、花瓣长、花瓣宽、雌蕊长、四强雄蕊长、二弱雄蕊长、四强花药长、花丝长和花柄长等花器性状。采用Microsoft Excel 2007处理数据, 用统计软件SPSS 20.0分析所测数据。体式显微镜(励扬LY-WN-OPLENIC)观察花朵形态并照相。1.4 花药石蜡切片观察
采用常规石蜡切片方法, 用70%、85%、95%和100%梯度乙醇溶液脱水, 二甲苯透明, 石蜡浸蜡和包埋, 切片厚度6~8 μm, 郝伯特配方粘片剂粘片, 苏木精-伊红染色, 中性树胶封片, 待胶完全凝固后, 在荧光显微镜(SOPTOP RX50)下观察并照相。1.5 TUNEL染色
采用Dead End Fluorometric TUNEL System (Promega)试剂盒方法检测细胞凋亡。花药石蜡切片用二甲苯脱蜡, 梯度乙醇脱水洗涤, PBS洗涤5 min, 4%多聚甲醛溶液固定15 min, 然后用20 μg mL-1蛋白酶K室温孵育10 min, 加rTdT孵育缓冲液(Equilibration Buffer, Nucleotide Mix, rTdT)对核DNA片段进行脱氧三磷酸尿苷缺口末端标记, 37℃避光孵育60 min。用碘化丙啶(PI, Sigma)复染。使用荧光显微镜(SOPTOP RX50)观察。在(520 ± 20) nm处观察荧光素的绿色荧光(TUNEL信号); 在>620 nm处观察碘化丙啶的红色荧光。2 结果与分析
2.1 160S-MF和160S-MS花朵形态学分析
对比观察160S-MF (15℃)、160S-MS (28℃)花朵的形态发现, 两者在外观形态上有明显差别, 160S-MF花朵较大(图1-A), 雄蕊长度高于雌蕊(图1-B), 花药呈黄色, 形态饱满, 能正常开裂, 释放出有活性的花粉; 160S-MS能正常开花, 花朵呈浅黄色(图1-D), 雄蕊6枚, 四强二弱, 但花朵整体偏小, 雄蕊长度不及雌蕊1/2 (图1-E), 花药褐黄色呈小三角形, 干瘪萎缩, 无花粉。160S-MS的花器形态也发生显著变化(表1)。在所测的花器特征参数中, 除雌蕊和萼片外其余花器官都达到极显著水平。与160S-MF相比, 160S-MS花瓣长度、宽度显著性变小、变窄(图1-C, F), 雄蕊(四强雄蕊和二弱雄蕊)明显退化, 花药和花丝极显著短于可育株的花丝和花药。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1160S-MF和160S-MS花形态学观察
A: 160S-MF盛花期花朵; B: 160S-MF雄蕊; C: 160S-MF花瓣和萼片; D: 160S-MS盛花期花朵; E: 160S-MS雄蕊; F: 160S-MS花瓣和萼片。
Fig. 1Flower morphology of 160S-MF and 160S-MS
A: the flower of 160S-MF; B: the flower stamens of 160S-MF; C: the petal and sepal of 160S-MF; D: the flower of 160S-MS; E: the flower stamens of 160S-MS; F: the petal and sepal of 160S-MS.
Table 1
表1
表1160S-MF和160S-MS的花器性状比较
Table 1
| 花器特征 Flower trait | 育性类型Types of male sterility | |
|---|---|---|
| 160S-MF | 160S-MS | |
| 花冠径Diameter of corolla | 18.97±1.68 | 15.58±0.78** |
| 花瓣长Length of petal | 13.98±1.16 | 12.30±0.57** |
| 花瓣宽Width of petal | 7.48±0.78 | 5.88±0.37** |
| 雌蕊长Length of pistil | 9.24±1.13 | 9.80±0.72 |
| 四强雄蕊长Length of four stronger stamens | 9.38±0.45 | 6.11±1.09** |
| 二弱雄蕊长Length of two weaker stamens | 6.99±0.81 | 4.89±1.20** |
| 四强花药长Length of four stronger stamens’ anther | 2.25±0.15 | 1.80±0.08** |
| 花丝长Length of four long filaments | 7.44±0.46 | 4.69±0.89** |
| 花柄长Length of stalk | 29.13±1.38 | 16.50±1.27** |
| 萼片长Length of sepal | 7.10±0.82 | 6.61±0.51 |
新窗口打开|下载CSV
2.2 160S-MF和160S-MS小孢子形态观察分析
通过对160S-MF、160S-MS不同发育阶段小孢子发育的细胞学观察, 研究了花药败育的特征, 如图所示, 跟大多数植物一样, 160S-MF花粉母细胞经2次减数分裂后形成四分体结构(图2-A), 周围有胼胝质; 随着胼胝质解体后, 进入单核早期(图2-B), 四分体慢慢释放出圆形的单个小孢子, 形态较小, 未明显观察到细胞核; 单核靠边期(图2-C), 明显能观察到细胞核移向细胞一侧, 此时的细胞核形态清晰, 整个细胞呈三瓣状; 之后, 随着小孢子继续发育, 细胞核经一次有丝分裂(图2-D), 形成一大一小2个细胞核, 这时进入双核期, 营养核较大位于细胞中央, 靠近细胞壁一侧较小的细胞核为生殖核; 小孢子继续发育至成熟期(图2-E), 花粉粒染色后整体颜色较深, 细胞核不能明显辨别, 形态饱满接近圆形。对160S-MS花药染色压片, 并未观察到小孢子或者花粉粒(图2-F), 表明花药很有可能在小孢子形成前就已败育。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2160S-MF和160S-MS 小孢子形态
A: 160S-MF四分体; B: 160S-MF早期小孢子; C: 160S-MF单核小孢子; D: 160S-MF双核小孢子; E: 160S-MF花粉粒; F: 160S-MS花粉粒。箭头所指为细胞核。
Fig. 2Microspores morphology of 160S-MF and 160S-MS
A: terads of 160S-MF; B: early microspores of 160S-MF; C: uninucleate microspores of 160S-MF; D: binucleate microspores of 160S-MF; E: pollen grains of 160S-MF; F: pollen grains of 160S-MS. The arrow points to the nucleus of the cells.
2.3 160S-MF和160S-MS花药细胞学观察
对160S-MF花药切片观察发现, 在造孢时期, 造孢母细胞紧密地排列在药室中央, 细胞核清晰可见, 形态呈不规则多边形(图3-A); 随着花药的发育, 造孢母细胞间稍微分开, 此时花药壁由表皮、药室内壁、中层组成, 绒毡层的发育不是很清楚(图3-B); 进入花粉母细胞时期, 花粉母细胞进一步发育, 体积变大, 形状呈椭圆形, 绒毡层细胞整齐地排列在花药壁最内层, 呈长方形, 清晰可见(图3-C); 减数分裂期(图3-D), 有的花粉母细胞有两团染色质, 表明其在进行核分裂, 此时, 绒毡层细胞处于双核状态; 四分体时期(图3-E), 小孢子呈四面体型, 绒毡层开始出现降解趋势; 小孢子细胞核靠近细胞一侧, 此时进入单核晚期, 小孢子的花粉壁开始增厚(图3-F); 双核期的小孢子细胞壁逐渐加厚且趋于圆球形, 绒毡层持续降解(图3-G); 成熟花粉期, 绒毡层不复存在, 花粉粒呈椭圆形, 形态饱满, 染色后变深(图3-H), 4个花粉囊呈蝴蝶型, 花粉囊开裂, 释放出花粉(图3-I)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3160S-MF花药不同发育阶段的显微结构
A和B: 造孢时期; C: 花粉母细胞时期; D: 减数分裂时期; E: 四分体时期; F: 单核靠边期; G: 双核期; H和I: 成熟花粉粒; Epi: 表皮层; Mid: 中层层; En: 药室内壁; T: 绒毡层; Sp: 造孢细胞; PMC: 小孢子母细胞; Te: 四分体; M: 小孢子; Po: 花粉粒。
Fig. 3Microscopical structure of anthers at different developmental stages in 160S-MF
A, B: sporogenous cell stage; C: pollen mother cell stage; D: meiotic stage; E: tetrad stage; F: late uninucleate stage; G: binucleate stage; H, I: mature pollen; Epi: epidermal; Mid: middle layer; En: endothecium; T: tapetum; Sp: sporogenous cell; PMC: pollen mother cell; Te: tetrad; M: microspore; Po: pollen.
对160S-MS花药切片观察发现, 花药也能分化形成蝴蝶型排列的4个花粉囊(图4-A)。160S-MS在造孢时期和花粉母细胞时期与160S-MF无明显差异(图4-B, C), 造孢细胞和花粉母细胞发育均正常, 此时组成花药壁的各组织结构在形态和发育也无异常。减数分裂期(图4-D, E), 160S-MS花药绒毡层发育出现异常, 尽管绒毡层为双核细胞, 但与同一时期的160S-MF花药绒毡层相比形态结构发生变化, 细胞排列不整齐, 空泡化, 出现解体, 花药开始出现败育现象。减数分裂期完成之前, 绒毡层全部解体(图4-F), 花粉母细胞也逐渐解体, 最终剩下空花粉囊。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4160S-MS花药不同发育阶段的显微结构
A和B: 造孢时期; C: 花粉母细胞时期; D和E: 减数分裂时期; F: 败育花药药室; Epi: 表皮层; Mid: 中层层; En: 药室内壁; T: 绒毡层; Sp: 造孢细胞; PMC: 小孢子母细胞。
Fig. 4Microscopical structure of anthers at different developmental stages in 160S-MS
A, B: sporogenous cell stage; C: pollen mother cell stage; D, E: meiotic stage; F: locule of aborted anther; Epi: epidermal; Mid: middle layer; En: endothecium; T: tapetum; Sp: sporogenous cell; PMC: pollen mother cell.
2.4 TUNEL染色分析
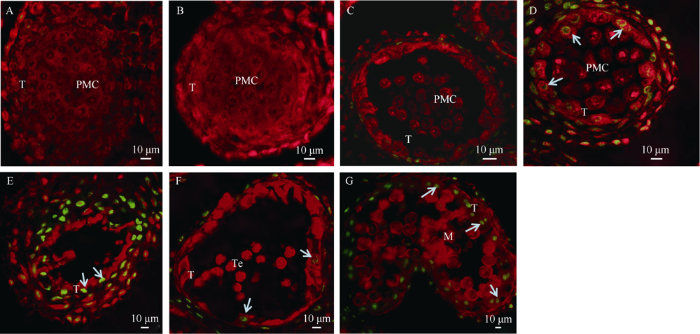
在植物花药中, 绒毡层降解被认为是细胞程序性死亡(programmed cell dead, PCD)的结果, 其特征是细胞核DNA的断裂。为进一步研究160S-MS花药绒毡层发育的异常现象, 本研究使用TUNEL法观察绒毡层细胞凋亡情况(图5)。对160S-MF、160S-MS不同发育阶段的花药切片进行TUNEL染色观察分析表明, 在花粉母细胞时期, 160S-MF、160S-MS花药绒毡层细胞的细胞核均为TUNEL阴性(图5-A, B), 表明该阶段绒毡层细胞发育正常; 减数分裂期, 160S-MF并未检测到绒毡层凋亡信号(图5-C), 相比之下, 160S-MS花药绒毡层细胞在减数分裂期检测到绿色荧光信号(图5-D), 表明其绒毡层核DNA断裂, 细胞开始凋亡; 随着花药发育, 绿色荧光信号越强烈(图5-E), 最终只剩下空的花粉囊。四分体时期(图5-F), 160S-MF花药绒毡层细胞核才出现微弱绿色荧光信号, 绒毡层开始降解, 分泌小孢子从四分体释放出来所需的物质, 并且荧光信号持续到双核期(图5-G)。表明160S-MS花药绒毡层细胞在减数分裂期提前凋亡。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5160S-MF、160S-MS花药各时期细胞凋亡TUNEL检测
A: 160S-MF花粉母细胞时期; B: 160S-MS花粉母细胞时期; C: 160S-MF减数分裂时期; D和E: 160S-MS减数分裂时期; F: 160S-MF四分体时期; G: 160S-MF双核期; T: 绒毡层; PMC: 小孢子母细胞; Te: 四分体; M: 小孢子。碘化丙染色细胞核呈红色荧光, TUNEL阳性细胞核呈绿色荧光(箭头所指)。
Fig. 5TUNEL detection of anther programmed cell dead at different developmental stages in 160S-MF and 160S-MS
A: 160S-MF at pollen mother cell stage; B: 160S-MS at pollen mother cell stage; C: 160S-MF at meiotic stage; D and E:160S-MS at meiotic stage; F: 160S-MF at tetrad stage; G:160S-MF at binucleate stage; T: tapetum; PMC: pollen mother cell; Te: tetrad; M: microspore. The fluoresce of nuclei stained by propidium iodide is red, while green fluorescence is TUNEL-positive nuclei staining (arrow).
3 讨论
在本研究中, 160S-MS雄蕊明显退化, 花丝变短, 这些形态特征与大多数温敏甘蓝型油菜雄性不育材料相同。对160S-MS醋酸洋红染色未观察到小孢子或者花粉粒, 说明160S-MS败育彻底, 由于雌蕊发育正常, 可以正常授粉。因此, 不育系160S可以用于油菜两系育种, 培育新型杂交油菜品种。在花粉发育过程中, 任何阶段的异常都会引起雄性不育, 败育发生时期和方式是多样的[18,19]。余凤群等[20]根据甘蓝型油菜雄性不育系花药败育时期和方式, 将其败育分为无花粉囊型不育系、花粉母细胞败育型不育系、单核败育型不育系3类。本研究发现, 不育系160S花药在减数分裂时期出现异常, 属于花粉母细胞败育型不育系。花粉母细胞不能完成减数分裂, 无法形成正常的四分体结构, 这一败育特征与TE5A[14,15]败育现象非常相似。160S-MS花药绒毡层在减数分裂时期出现结构和形态出现异常, 细胞空泡化, 伴随提前解体, 然而, 不育系TE5A绒毡层发育正常。从绒毡层发育层面看, 160S-MS与373S[7]、SP2S[16]和Huiyou 50S[21]都出现提前凋亡, 但这些不育系材料在发育过程中能形成正常的四分体结构, 败育发生在小孢子发育过程中, 而160S-MS在花粉母细胞减数分裂未完成时, 就形成空的花粉囊, 未观察到四分体结构和小孢子。总之160S败育时期和特点与目前报道的甘蓝型温敏核雄性不育材料存在差异。
植物花粉的生长发育需要绒毡层在适当的时期发生细胞程序性死亡[22]。 绒毡层位于花药壁最内层, 它与发育中的雄配子体直接接触, 能向小孢子提供发育需要的营养物质, 如胼胝质、孢粉素和蛋白质等[23,24,25,26]。研究表明, 绒毡层在花药和花粉的发育过程中发挥重要作用, 过早或延迟凋亡会导致雄性不育[27,28,29]。目前已在小麦[27]、拟南芥[30,31]、水稻[32,33]、油菜[34]等植物中发现大量雄性不育材料是由绒毡层发育异常引起的。李六林[35]根据前人对绒毡层发育的研究, 按照不同的植物种类, 将绒毡层解体分为2类: 一种是腺质型类型, 在花粉发育过程中, 绒毡层细胞分泌花粉母细胞和小孢子发育所需的各种营养物质, 绒毡层细胞直到花粉成熟才完全消失; 另一种是变形绒毡层, 绒毡细胞以多核原生质团的形式填充于小孢子之间的空隙中, 供小孢子利用和吸收。腺质型绒毡层多存在于被子植物中。在花药发育早期, 160S-MS能形成正常的花粉母细胞, 绒毡层细胞整齐地排列在花药壁最内层, 与160S-MF无差异。然而, 在减数分裂期, 160S-MS花药绒毡层细胞形态和结构出现异常, 细胞空泡化并伴随降解现象, 致使花粉母细胞减数分裂受阻, 慢慢解体, 无法形成四分体结构, 最终减数分裂完成前就只剩下空的花粉囊, 导致败育。可能是绒毡层细胞提前凋亡而未向腺质型转化, 不能向花粉母细胞提供发育所需的营养成分, 进而影响四分体小孢子的形成, 无成熟花粉粒产生, 导致160S败育。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:20150489 [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.tplants.2007.08.005URLPMID:17720610 [本文引用: 1]

Heterosis describes the superior performance of heterozygous hybrid plants over their homozygous parental inbred lines. Despite the rediscovery of this phenomenon a century ago and its paramount agronomic importance, the genetic and molecular basis of heterosis remains enigmatic. Recently, various pioneer studies described differences in genome organization and gene expression of hybrids and their parental inbred lines. At the genomic level, a significant loss of colinearity at many loci between different inbred lines of maize was observed. At the level of gene expression, complex transcriptional networks specific for different developmental stages and tissues were monitored in maize (Zea mays), rice (Oryza sativa) and Arabidopsis (Arabidopsis thaliana). Integration of this complex expression data might contribute to improve our understanding of the molecular basis of heterosis.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-007-0594-1URLPMID:17605126 [本文引用: 1]
S45AB, a recessive genic male sterile (RGMS) line, originated as a spontaneous mutant in Brassica napus cv. Oro. The genotypes of sterile (S45A) and fertile plants (S45B) are Bnms1ms1ms2ms2 and BnMs1ms1ms2ms2, respectively. In our previous studies, Yi et al. (Theor Appl Genet 113:643-650, 2006) mapped the BnMs1 locus to a region of 0.4 cM, candidates of which have been identified and genetic transformation is in progress. We describe the fine mapping of BnMs2 exploiting amplified fragment length polymorphism (AFLP) and amplified consensus genetic marker (ACGM) methodologies, and the identification of a collinear region probably containing BnMs2 orthologue in Arabidopsis thaliana. A near isogenic line (NIL) population S4516AB which segregated for BnMs2 locus was generated by crossing, allelism testing and repeated full-sib mating. From the survey of 1,024 AFLP primer combinations, 12 tightly linked AFLP markers were obtained and five of them were successfully converted into co-dominant or dominant sequence characterized amplified region (SCAR) markers. A population of 2,650 sterile plants was screened using these markers and a high-resolution map surrounding BnMs2 was constructed. The closest AFLP markers flanking BnMs2 were 0.038 and 0.075 cM away, respectively. Subsequently, an ACGM marker was developed to delimit the BnMs2 locus at an interval of 0.075 cM. We extended marker sequences to perform BlastN searches against the Arabidopsis genome and identified a collinear region containing 68 Arabidopsis genes, in which the orthologue of BnMs2 might be included. We further integrated BnMs2 linked AFLP or SCAR markers to two doubled-haploid (DH) populations derived from the crosses Tapidor x Ningyou7 (Qiu et al., Theor Appl Genet 114:67-80, 2006) and Quantum x No.2127-17 (available in our laboratory), and BnMs2 was mapped on N16. Molecular markers developed from these investigations will facilitate the marker-assisted selection (MAS) of RGMS lines, and the fine map and syntenic region identified will greatly hasten the process of positional cloning of BnMs2 gene.
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-006-0262-xURLPMID:16783588 [本文引用: 1]
The present study describes a novel thermo-sensitive genic male sterile (TGMS) line, Qiong68ms. To analyse the mode of fertility inheritance and tag the TGMS gene, a set of F2, BC1 and F(2:3) populations derived from a cross between Qiong68ms and K12 were evaluated for a period of 2 years. Classical genetic analyses and QTL mapping using the mean restoration percentage of the F(2:3) populations revealed that the fertility of Qiong68ms was likely to be governed by a single recessive gene, which was named tms3; the tms3 gene was mapped to a location between SSR markers umc2129 and umc1041, at a distance of 3.7 cM form umc2129 and 1.5 cM form umc1041. The molecular markers tightly linked with tms3 gene will aid in the transfer of the TGMS gene to various background inbred lines using the MAS method.
DOI:10.1007/s00122-003-1385-yURLPMID:12928780 [本文引用: 1]
A thermo-sensitive genic male-sterile (TGMS) wheat line ( Triticum aestivum L.) BNY-S was obtained from the spontaneous mutant of BNY-F. Its fertility was decided by the temperature during the differentiation stage of the spikelets. BNY-S was completely sterile when the temperature was lower than 10 degrees C during the differentiation stage of the spikelets, but fertile when the temperature was higher than 10 degrees C. Genetic analysis indicated that the sterility of BNY-S was controlled by a single recessive gene, which was named as wtms1. An F(2) population, consisting of 3,000 individuals from the cross between BNY-S and Lankao 52-24, was used for genetic analysis and statistical analysis of the TGMS and, out of them, 158 sterile and 93 fertile extremes were present for molecular tagging and mapping of the wtms1 gene. SSR (simple sequence repeat) and AFLP (amplified fragment length polymorphism) techniques combined with BSA (bulked segregant analysis) were used to screen markers linked to the target gene. As a result, wtms1 was preliminarily mapped on chromosome 2B according to SSR analysis. In AFLP analysis, 14 polymorphic AFLP loci were identified with a linkage relation to the wtms1 gene. Then linkage analysis using the F(2) population showed that three of them, E: AAG/M: CTA(163), E: AGG/M: CTC(220) and E: ACA/M: CTA(160), were linked to the wtms1 gene relatively close to a genetic distance of 6.9 cM, 6.9 cM and 13.9 cM, respectively. Finally, the wtms1 gene was mapped between the SSR marker Xgwm 374 and the AFLP marker E: AAG/M: CTA(163) with the distance of 4.8 cM and 6.9 cM, respectively. A partial linkage map was constructed according the SSR and AFLP data.
DOI:10.1093/jhered/esq100URLPMID:20864624 [本文引用: 1]
In soybean [Glycine max (L.) Merr.], manual cross-pollination to produce large quantities of hybrid seed is difficult and time consuming. Identification of an environmentally stable male-sterility system could make hybrid seed production commercially valuable. In soybean, 2 environmentally sensitive male-sterile, female-fertile mutants (ms8 and msp) have been identified. Inheritance studies showed that sterility in both mutants is inherited as a single gene. The objectives of this study were to 1) confirm that msp and ms8 are independent genes; 2) identify the soybean chromosomes that contain the msp and the ms8 genes using bulked segregant analyses (BSAs); and 3) make a genetic linkage map of the regions containing these genes. Mapping populations consisting of 176 F(2) plants for ms8 and 134 F(2) plants for msp were generated. BSA revealed that Sat_389 and Satt172 are closely associated markers with ms8 and msp, respectively. Map location of Sat_389 suggested that the ms8 gene is located on chromosome 7; molecular linkage group (MLG) M. Map location of Satt172 indicated that the msp gene is located on chromosome 2 (MLG Dlb). Genetic linkage maps developed using F(2) populations revealed that ms8 is flanked by a telomere and Sat_389 and msp is flanked by Sat_069 and GMES4176. The region between the telomere and Sat_389 is physically 160 Kb. Soybean sequence information revealed that there are 13 genes present in that region. Protein BLASTP analyses revealed that homologs of 3 of the 13 genes are known to a play role in cell division, suggesting putative candidates for ms8.
URLPMID:32231254 [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URLPMID:27670217 [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pone.0119557URLPMID:25803723 [本文引用: 1]
Chemical hybridization agent (CHA)-induced male sterility is an important tool in crop heterosis. To demonstrate that CHA-SQ-1-induced male sterility is associated with abnormal tapetal and microspore development, the cytology of CHA-SQ-1-treated plant anthers at various developmental stages was studied by light microscopy, scanning and transmission electron microscopy, in situ terminal deoxynucleotidyl transferasemediated dUTP nick end-labelling (TUNEL) assay and DAPI staining. The results indicated that the SQ-1-treated plants underwent premature tapetal programmed cell death (PCD), which was initiated at the early-uninucleate stage of microspore development and continued until the tapetal cells were completely degraded; the process of microspore development was then blocked. Microspores with low-viability (fluorescein diacetate staining) were aborted. The study suggests that premature tapetal PCD is the main cause of pollen abortion. Furthermore, it determines the starting period and a key factor in CHA-SQ-1-induced male sterility at the cell level, and provides cytological evidence to further study the mechanism between PCD and male sterility.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:15862102 [本文引用: 1]
DOI:10.1105/tpc.105.034090URLPMID:16141453 [本文引用: 1]
The tapetum, the innermost of four sporophytic layers in the anther wall, comes in direct contact with the developing male gametophyte and is thought to play a crucial role in the development and maturation of microspores. Here, we report the identification of rice (Oryza sativa) Undeveloped Tapetum1 (Udt1), which is required for the differentiation of secondary parietal cells to mature tapetal cells. T-DNA or retrotransposon Tos17 insertions in the Udt1 gene caused male sterility. The anther walls and meiocytes of the mutants were normal during the early premeiosis stage, but their tapeta failed to differentiate and became vacuolated during the meiotic stage. In addition, meiocytes did not develop to microspores, and middle layer degeneration was inhibited. Consequently, the anther locules contained no pollen. The UDT1:green fluorescent protein fusion protein was localized to the nucleus. This, together with its homology with other basic helix-loop-helix proteins, suggests that UDT1 is a transcription factor. DNA microarray analysis identified 958 downregulated and 267 upregulated genes in the udt1-1 anthers, suggesting that Udt1 plays a major role in maintaining tapetum development, starting in early meiosis.
URLPMID:12154130 [本文引用: 1]
[本文引用: 1]
DOI:10.1105/tpc.114.122986URLPMID:24781116 [本文引用: 1]
Mature pollen is covered by durable cell walls, principally composed of sporopollenin, an evolutionary conserved, highly resilient, but not fully characterized, biopolymer of aliphatic and aromatic components. Here, we report that ABORTED MICROSPORES (AMS) acts as a master regulator coordinating pollen wall development and sporopollenin biosynthesis in Arabidopsis thaliana. Genome-wide coexpression analysis revealed 98 candidate genes with specific expression in the anther and 70 that showed reduced expression in ams. Among these 70 members, we showed that AMS can directly regulate 23 genes implicated in callose dissociation, fatty acids elongation, formation of phenolic compounds, and lipidic transport putatively involved in sporopollenin precursor synthesis. Consistently, ams mutants showed defective microspore release, a lack of sporopollenin deposition, and a dramatic reduction in total phenolic compounds and cutin monomers. The functional importance of the AMS pathway was further demonstrated by the observation of impaired pollen wall architecture in plant lines with reduced expression of several AMS targets: the abundant pollen coat protein extracellular lipases (EXL5 and EXL6), and CYP98A8 and CYP98A9, which are enzymes required for the production of phenolic precursors. These findings demonstrate the central role of AMS in coordinating sporopollenin biosynthesis and the secretion of materials for pollen wall patterning.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1105/tpc.18.00907URLPMID:30886127 [本文引用: 1]
Pollen and microspore development are essential steps in the life cycle of all land plants that generate male gametes. Within flowering plants, pollen development occurs inside of the anther. Here, we report the identification of two class III peroxidase-encoding genes, PEROXIDASE9 (PRX9) and PRX40, that are genetically redundant and essential for proper anther and pollen development in Arabidopsis (Arabidopsis thaliana). Arabidopsis double mutants devoid of functional PRX9 and PRX40 are male sterile. The mutant anthers display swollen, hypertrophic tapetal cells and pollen grains, suggesting disrupted cell wall integrity. These phenotypes lead to nearly 100%-penetrant pollen degeneration upon anther maturation. Using immunochemical and biochemical approaches, we show that PRX9 and PRX40 likely cross-link extensins to contribute to tapetal cell wall integrity during anther development. This work suggests that PRX9 and PRX40 encode Arabidopsis extensin peroxidases and highlights the importance of extensin cross-linking during pollen development.
DOI:10.1093/jxb/eraa045URLPMID:31989154 [本文引用: 1]
Timely degradation of anther tapetal cells is a prerequisite for normal pollen development in flowering plants. Although several genes involved in tapetum development have been identified, the molecular basis of tapetum degeneration regulation remains poorly understood. In this study, we identified and characterized the nucleus-encoded, conserved mitochondrial aldehyde dehydrogenase OsALDH2b as a key regulator of tapetum degeneration in rice (Oryza sativa). OsALDH2b was highly expressed in anthers from meiosis to the early microspore stage. Mutation of OsALDH2b resulted in excess malonaldehyde accumulation and earlier programmed cell death in the tapetum, leading to premature tapetum degeneration and abnormal microspore development. These results demonstrate that OsALDH2b negatively regulates tapetal programmed cell death and is required for male reproductive development, providing insights into the regulation of tapetum development in plants.
URLPMID:31772077 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}