 ,1,2,*, ����1, ������1, ���1, ������1, ���Ƿ�2, ��ʾ��2
,1,2,*, ����1, ������1, ���1, ������1, ���Ƿ�2, ��ʾ��2Analysis on the structures of polygalacturonase-inhibiting proteins and the expression profile of its encoding genes in rice
CHEN Xi-Jun,1,2,*, TANG Tao1, LI Li-Li1, CHEN Chen1, CHEN Yu-Wen1, ZHANG Ya-Fang2, ZUO Shi-Min2ͨѶ����:
�ո�����:2020-02-16��������:2020-07-2�����������:2020-07-17
| ��������: |
Received:2020-02-16Accepted:2020-07-2Online:2020-07-17
| Fund supported: |

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (3268KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
��Ϧ��, ����, ������, ���, ������, ���Ƿ�, ��ʾ��. ˮ����۰�����ȩ��ø���Ƶ�����OsPGIP�ṹ�������������
���� [J]. ����ѧ��, 2020, 46(12): 1884-1893. doi:10.3724/SP.J.1006.2020.02011
CHEN Xi-Jun, TANG Tao, LI Li-Li, CHEN Chen, CHEN Yu-Wen, ZHANG Ya-Fang, ZUO Shi-Min.
��������ά���ǹ���ֲ��ϸ���ڹǼܵĻ�������, Ҳ�Dz�ԭ����������������ϡ��ڲ�ԭ���������������, PG��������ĵ�һ�����ڽ���ø[1]��PGIP��ֲ��ϸ���ڵ���Ҫ��ɲ���, ���˿�������ڹ����γɸ�����ӹ�ϸ������, ���������Խ�ϲ���PG, ����PG�Լ���ϸ�����ƻ�����, ��������ά��ϸ����������, �Ӷ��谭��������, ���Ͳ������ض�[2,3]��ͬʱ, ���ڲ�ԭ��PG���Ա�����, ���һ���ֽ�����ѽ����Ѿ۰�����ȩ����(oligalacutronides, OG)�������½�, �Ӷ�����OG�ڼ������ڸ���, ��Щ���м����ӻ��Ե�OG�ֿɼ�������ķ�����Ӧ, ��һ�������������[4,5,6,7]��
Ҫ��ȷij��PGIP�Բ���PG���Ե���������, ��ֱ�ӵķ������ô�����PGIP�����вⶨ, �������о���Ϊ��ֲ����֯����ȡPGIP, ��Щ�����ܰ����Ŷ���߶����Ƶ�PGIP�칹��, ���ý��Ҳ����ЩPGIP��ͬ������, �����ǵ�������[8]�����, Ҫ�о�ijһ��PGIP������, ������Դ����ķ�����ô�PGIP�����ǽϺõ�;��������ĿǰΪֹ, �ѿ�¡��170��PGIP�����н������������о�, ��Ϊ���������ϵͳ��PGIP�����ܱ���, ���γɰ�����, ���������Ҫ�����, �����������û�л���[9,10,11,12]�����, ���о���ЩPGIP֮ǰ, ����������ʽ���������ϢѧԤ��, Ϊ��һ���о��ṩ����֧�ַdz���Ҫ��
��ֲ������, PGIP������Լ�����ʽ����, ��Ȼ��Щ�������Ŵ��Ͼ��и߶ȵ�������, ����Ӧ�Բ�ͬ��ԭ��������澳����ʱ, ����ˮƽ����������; ��ʹ����ͬ�Ļ���, ��ֲ�������IJ�ͬ������, ����������в�ͬ[13,14,15]����˶�����4��PvPGIP, ������������������, ��PvPGIP2��������֯�б���, ��PvPGIP3��PvPGIP4ֻ�ڸ���������, PvPGIP1������Ҷ�������ᡢ���Ͷ����о�������[16]�����ܵ�������Ⱦ�ͻ�е���˵��澳��OG��ˮ�����ֲ�D�صȾ����յ����õ����Ӵ�����, ���PvPGIP������в�ͬ��Ӧ��, ֻ��PvPGIP4�����д��������±������������Ա仯[17,18]����Ŀǰ���б���, ˮ���й�����7��OsPGIP����[19,20,21]��Lu��[19]�о�����, �ھ���Ʒ���л�11��, 7��OsPGIP������������˳ʽ����Ԫ����ɶ���, ��Ը���ֲ�D�ش����ķ�ӦҲ������ͬ��Ϊ�˸�����ȷ��OsPGIP����ı�������, ���о�ѡ�ÿ������ƿݲ�ˮ��Ʒ�ֲ�ͨ�����¡��ڹ�ͽ��ֲ�ԭ�����澳����, ���о���ͬ�����¸�OsPGIP����ı���ˮƽ, Ϊ��һ������Щ����Ӧ����ˮ�����������ṩ�������ݡ�
1 �����뷽��
1.1 ���Բ���
���Ծ���: ˮ���ƿݲ���YN-7, �ɱ�ʵ���ҷ����Խ���ˮ���ƿݲ����ꡣ����ˮ��: YSBR1 (��)���쵾3��(�и�)��Lemont (�߸�)�������ݴ�ѧũѧԺˮ�����������Ŵ��������о����ṩ��1.2 OsPGIP����Ŀ�¡�����
ˮ��������DNA��ȡ����Khanuja��[22]�ķ�����RNA��ȡ������Ĭ�������Ƽ���˾(�й�)ֲ��RNA��ȡ�Լ���, ��ת¼cDNAʹ����������Ƽ�(����)����˾FastKing RT�Լ��С�����NCBI�Ͳο������и������������, ʹ��Primer V5.0������Ʊ��о���������(��1)�����������ɱ����﹤������˾(����TaKaRa��˾)�ϳɡ��ֱ���ˮ��������DNA��cDNAΪ�������PCR����, ����������������¡����pMD19-T, ���Կ�¡�����Ϻ��������﹤������˾����Table 1
��1
��1������OsPGIP������������
Table 1
| ���� Gene | �������� Forward primer (5��-3��) | �������� Reverse primer (5��-3��) |
|---|---|---|
| OsPGIP1 | TGACTCGCTATTGCATGCG | TGGGAGCTTAATTGCAGGGA |
| OsPGIP2 | ATACACGGCATTGCATGCAC | CTTACACTCGTTCTCCGTAC |
| OsPGIP3 | TAGAAGAGAGGAAGCACGCA | TTGGTGGCCTGAGATAGGT |
| OsPGIP4 | TGTCGTGCACTTGTGTTCAA | GCATTAGCTGGTTGCTTC |
| OsFOR1 | TTCAGGTAGATACAATGGCG | ATGGATGGATGGATGCTC |
| OsPGIP6 | GAGCCGAGACGAGACGA | ATATGTACCCAAGCCCAAA |
| OsPGIP7 | TCCTGCACGGATTTGAGC | TAACAACAGCCAGTCAGCAAT |
�´��ڴ�|����CSV
1.3 ϵͳ�������Ĺ���
�ԡ�Polygalacturonase inhibiting protein��Ϊ�ؼ��ʴ�NCBI��������ص�������, ȥ����PGIP��δ��ȷ����ΪPGIP������, ����Щ�����뱾ʵ���п�¡����ķ���������ϵͳ��������, ����MEGA 5.1��������ϵͳ������������1.4 OsPGIP�ṹԤ��
������һ���ṹ�������ź���Ԥ������ֱ�ʹ����վ(1.5 OsPGIP�����빦��Ԥ��
�����ʵķ��������ȵ�㡢�ȶ��ԡ�֬���ԡ���ˮ�ԡ���Ĥ�ṹ����ϸ����λ��N-�ǻ���λ���Լ��˾��б��������ܽ��Ե������빦�ֱܷ�ͨ����վ(1.6 ˮ���紦������֯ȡ��
��ͬ������ȡ��: ѡ����������Ŀ������ƿݲ�ˮ��Ʒ��YSBR1���쵾3�ź�Lemont��ͬ������ (������Ҷ�ڡ������ں�����)��ֲ��, ȡ�в�Ҷ����֯���ڹ��: ȡ��Ʒ��ˮ�����Ӿ����֡���ѿ��, ѡȡѿ����һ�µ���������ƽ����������(150 mm)�ײ���ʪɴ����, 26������������йⰵ����(14 h/10 h)����14 d��, �ڹ�ڰ�������������ˮ���嵽��Ҷ��, ȡֲ���в�Ҷ����֯, ���������������µ���Ϊ���ա�
���´���: ����������14 d��ˮ����4���������������4 d, ȡֲ���в�Ҷ����֯, �������ⰵ������������Ϊ���ա�
��ԭ�����ִ���: �������ǩ���ֵķ���[40], �����ƿݲ�����ˮ��Ҷ����, �Բ��Ӿ�������, 48 h��ȡֲ���в�Ҷ����֯��
������֯ȡ������������Һ����, -80�汣��, ����ֲ����RNA��ȡ��
1.7 qRT-PCR����
��ȡ��ˮ����֯��Ʒ��RNA, ��ת¼��cDNA��, ����CFX96 Real-time���ϵͳ(Bio-Rad)���м�⡣20 ��L��Ӧ��ϵ: AceQqPCR SYBR Green Master Mix 10 ��L��Forward Primer 0.4 ��L��Reverse Primer 0.4 ��L��ROX Reference Dye 1 0.4 ��L��ģ��cDNA 2 ��L��ddH2O 6.8 ��L����Ӧ��������: 95�� 5 min; 95�� 15 s, 60�� 20 s, 40��ѭ��; 95�� 15 s����OsACTIN1 (NCBI���к�ΪX16280)Ϊ�ڲλ���, ��������ΪOsACTIN- F/OsACTIN-R (5��-CAGCATGAAGATCAAGGTGG-3�� / 5��-TTCCTGTGCACAATGGATGG-3��), ������OsPGIP������������вμ�����[19]��ÿ����Ʒ��3������ѧ�ظ�, ÿʵ���ظ�3�Ρ�1.8 ����ͳ�������
�������ݲ���SPSS 12.05�������з������, ��Tukey������ж��رȽ�, ��P<0.05ʱ, ���ʾ����������2 ��������
2.1 OsPGIPϵͳ��������
��ˮ��cDNA��������7��OsPGIP�������в����Ϻ��������﹤������˾����, ��NCBI�ȶ�ȷ����������������ȷ���ԡ�Polygalacturonase inhibiting protein��Ϊ�ؼ��ʴ�NCBI��վ�й���������������333��, ������ȷע��������Դ������ΪPGIP�Ĺ�258��������Щ�����������OsPGIPһ�����бȶ���ϵͳ�����������������, �������ͬ��������ֵ�PGIP�����Խϸ�, �ۼ���1��2����֧(Ⱥ)��, ��ʮ�ֻ��ơ����ơ��̱��Ƶ�ֲ���PGIP (ͼ1-A); ������Щ�ۼ�Ⱥ��Ҳ������, ���Ϊ���������������PGIP (NCBI���к�ΪEPS70789)��ˮ���ļ���PGIP���и߶�����, ��������PGIP (NCBI���к�ΪAEX34755)�붹��PGIP����һ��; ͬ��, ��Դ��ˮ����7��OsPGIP��OsPGIP2��OsPGIP4��OsPGIP6��OsPGIP7���������Խϸ�, ������ͬһ��֦, ��������OsPGIP�ֱ�λ�ڽ�������ͬ�ķ�֦(ͼ1-B)����Щ�����������, ����ͬһ�����е�PGIP������Դ��1�����������������������š���ͻ�䡢СƬ�β����ȱʧ�Ļ������IJ���, ����ͬ�����е�PGIPҲ�п���Դ��ͬһ���ȡ�ͼ1
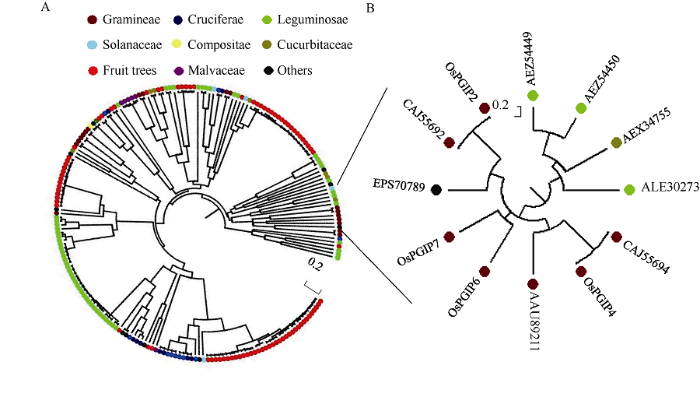 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1OsPGIP������������Դ��PGIPϵͳ������
ͼ�г�OsPGIPΪ������д��, ������Ϊ��ӦPGIP��NCBI���кš�A: �ܽ�����; B: ������
Fig. 1Phylogenetic relationship among OsPGIPs and PGIPs from other plant species
OsPGIPs is the abbreviations of polygalacturonase-inhibiting protein from Oryza sativa, the others are the NCBI accession numbers of the corresponding sequences. A: phylogenetic tree; B: subtree.
2.2 OsPGIP�ռ�ṹԤ��
һ���ṹ��������, 7��OsPGIP����ɷֱ���뺬��309~380���������OsPGIP����, ÿ����������һ���ź���, ������17~31��������; ���ź�����, ��OsPGP������N-�ˡ�9~11��LRR (Leucine-rich repeat)Ƭ�κ�C-�˵ȵ��͵�PGIP�ṹ��; ����ЩLRRƬ����, �����ڱ��ص�xxLxLxx����, LΪ�������֬���就����л�, xΪ���ⰱ����(ͼ2)��OsPGIP�Ķ����ṹ��Ҫ�ɦ�-��������-�۵�������������, �Ҷ����������Ϊ��; OsPGIP��N-�˺�C-�˷ֱ���8~13�����װ���, ���γ�3~5�������, ��Щ��������ȶ�OsPGIP���ض��ռ�ṹ����Ҫ����(��2)��Table 2
��2
��2OsPGIP�����ṹԤ����
Table 2
| ���� Protein | ��-���� ��-helix | ������ Extended strand | ������� Random coil | ���װ����� Number of cysteine | �����λ�� Position of disulfide bond |
|---|---|---|---|---|---|
| OsPGIP1 | 36.25 | 10.68 | 53.07 | 8 | 56-63, 278-298, 300-308 |
| OsPGIP2 | 46.20 | 10.23 | 43.57 | 9 | 34-64, 65-72, 310-323, 331-339 |
| OsPGIP3 | 32.74 | 13.86 | 53.39 | 10 | 17-27, 56-64, 322-328, 330-337 |
| OsPGIP4 | 44.70 | 9.74 | 45.56 | 9 | 33-63, 64-71, 333-339, 341-348 |
| OsFOR1 | 30.12 | 10.24 | 59.64 | 10 | 27-58, 59-66, 312-320 |
| OsPGIP6 | 49.21 | 4.47 | 46.32 | 10 | 64-73, 114-137, 348-370, 372-379 |
| OsPGIP7 | 27.19 | 10.82 | 61.99 | 13 | 16-65, 25-34, 66-73, 322-328, 330-337 |
�´��ڴ�|����CSV
ͼ2
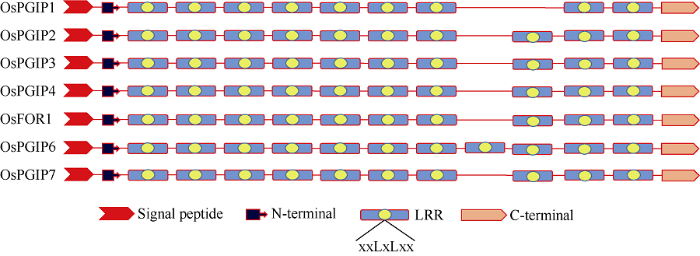 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ27��OsPGIPһ���ṹ
Fig. 2Primary structure of OsPGIPs
����ͬԴ��ģ�ķ���Ԥ��OsPGIP�������ṹ, �������, ����SWISS-MODEL��CPHmodelsԤ�������OsPGIP����ά�ռ�ṹ���Ƚ�����, �����ɶ���������-��-����-�������-��-�۵�����ɵ���Ȧ״�ṹ���������������γ�һ���ض��İ���(ͼ3-A)������ͬԤ�������õ���ͬһOsPGIP��ά�ռ�ṹ������ȫ��ͬ, �е���������ϴ�, �ر���SCRATCH��ģ�������������ַ������ý��������ȫ��ͬ���ֱ���SWISS-MODEL��CPHmodelsģ��OsPGIP6�Ŀռ�ṹ, �����ʾ, �������Dz���Ԥ��İ�������(34~369��39~354)�������Ḳ����(88.4%��83.2%)����ģ���������(30.1%��33.6%)���������, ����Ԥ�������в���, �����ַ���Ԥ���OsPGIP6�ռ�ṹ��, ��LRR���ֱ����13����10����-�۵�, ����SCRATCHģ��������͵�PGIP�ռ�ṹ����, LRR������ڦ�-�۵�; ͬ���ķ���Ӧ����Ԥ��OsPGIP7�Ŀռ�ṹ�õ��Ľ������(ͼ3-B)��������3�ַ�ʽģ���OsPGIP7�ṹ��ˮ���ƿݲ�����RsPG2���е����ʶԽӷ���, ���ܻ�õĶԽ�ʾ��ͼ������ȫһ��, ������ʾΪOsPGIP7�İ�����RsPG2����϶����λ���н��ܽ��, ˵���ò�λ�����ߵĻ�������, ��������ǰ���о�OsPGIP1��OsPGIP2��RsPG1�ԽӵĽ����һ��[12], ˵����ͬPGIP�벻ͬPG�Ļ�����ʽ�Ƕ��ֶ�����(ͼ3-C)��
ͼ3
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3OsPGIP������3D�ṹģ��ͼ
��ͨͼ�к�ɫ����ɫ����ɫ�ֱ��ʾ��-��������-�۵������������A: ���е���PGIP�ռ�ṹ�ĸ�OsPGIP3Dģ��; B: �ֱ����SWISS-MODEL��CPHmodels��SCRATCH����ģ���OsPGIP6��OsPGIP7�ռ�ṹģ��; C: OsPGIP7��RsPG2�ĵ����ӶԽ�, ����ͼΪOsPGIP7, ��ͨͼΪRsPG2��
Fig. 33D structure of OsPGIPs
Red, yellow, and green in the cartoon figures mean ��-helix, ��-sheet, and random coil, respectively. A: 3D structure of OsPGIPs with typical spatial structure of PGIP. B: 3D structures of OsPGIP6 and OsPGIP7 constructed with homologous modeling methods of SWISS-MODEL, CPH model and SCRATCH. C: Protein docking of OsPGIP7 and RsPG2. Mesh and cartoon figures mean OsPGIP7 and RsPG2, respectively.
2.3 OsPGIP�����빦��Ԥ��
����ǰ�˱���, 7��OsPGIP�ı������ֱ�λ��3��Ⱦɫ����, ����OsPGIP1~OsPGIP4λ��ˮ��5��Ⱦɫ��, OsFOR1��OsPGIP6λ��8��Ⱦɫ��, OsPGIP7λ��9��Ⱦɫ��[19,20,21]����Ԥ��, 7��OsPGIP�ķ���������35 kD����(32.75~38.79 kD), �ȵ��Ϊ4.73~8.37������, �ѱ��������Ʋ���PG���Ի�����������ֲ��Ե�OsPGIP1��OsPGIP4��OsFOR1[20,23-24]��ȵ��������Ի�ƫ���ԡ�Ԥ��ı�������OsPGIP2���ȶ���, �������ȶ�; ��������Ԥ�⵰��Ϊ��ˮ���ס�֬���Ժá����п�Ĥ�ṹ����λ��ϸ����, �Ҿ�����1�����N-�ǻ���λ�㡣����, ��Щ�����ڴ˾���ԭ�˱������ܽ�ȼ��ͻ���ȫ����(��3)��Table 3
��3
��3��������Ԥ���OsPGIP���������빦��
Table 3
| ���� Protein | Ⱦɫ�� λ��Location | ������ Molecular weight (kD) | �ȵ�� pI | ���ȶ� ָ�� Instability index | ֬���� ָ�� Aliphatic index | ��ƽ�� ��ˮ�� Grand average of hydropathicity | ��Ĥ�� Trans- membrane | ��ϸ����λ Subcellular localization | N-�ǻ��� λ���� Number of N-glycosylation sites | �˾��б�����ܽ��� Chance of solubility expressed in E. coli (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| OsPGIP1 | Chr. 5 | 32.75 | 6.98 | 31.82 | 98.58 | 0.183 | Yes | Extracellular | 4 | 0 |
| OsPGIP2 | Chr. 5 | 36.97 | 4.73 | 40.16 | 103.33 | 0.080 | Yes | Extracellular | 8 | 0 |
| OsPGIP3 | Chr. 5 | 36.14 | 5.94 | 34.36 | 95.63 | 0.181 | Yes | Extracellular | 5 | 0 |
| OsPGIP4 | Chr. 5 | 37.22 | 8.37 | 33.98 | 101.58 | 0.203 | Yes | Extracellular | 4 | 0 |
| OsFOR1 | Chr. 8 | 35.46 | 7.09 | 36.08 | 99.94 | 0.041 | Yes | Extracellular | 1 | 0 |
| OsPGIP6 | Chr. 8 | 38.79 | 5.86 | 35.69 | 106.42 | 0.286 | Yes | Extracellular | 1 | 0 |
| OsPGIP7 | Chr. 9 | 35.88 | 5.90 | 33.26 | 101.87 | 0.265 | Yes | Extracellular | 3 | 0.7 |
�´��ڴ�|����CSV
2.4 OsPGIP����ı�������
�ֱ�ȡ��ͬ�����ڿ����к߸��ƿݲ�ˮ��Ʒ��YSBR1���쵾3�ź�Lemont��Ҷ����֯, ��ȡ��RNA, qRT-PCR����OsPGIP����ı����������ݷ����������, ���ڸ�OsPGIP��������ڿ���Ʒ��YSBR1�б��������, �������ں�����������������ɲ����ԡ����������������, OsFOR1��OsPGIP7�ڳ�����ˮ��Ҷ���б������ϸ�, ��OsPGIP2��OsPGIP4��OsFOR1��������Ҷ���б������ϸ�; ��OsPGIP������ܱ��������Ƚ�, ���Ը߸�Ʒ��Lemont���(ͼ4)���˽��������ǰ���о�OsPGIP1�����������ʱ�Ľ���һ��[25]��ͼ4
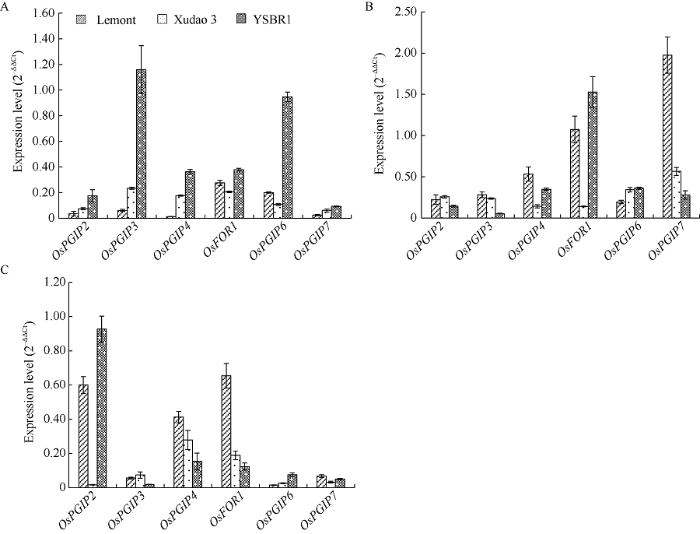 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4��ͬ�����ڿ������ƿݲ�ˮ��Ʒ��Ҷ���и�OsPGIP����ı�����
A: ����; B: ������; C: ���ڡ�
Fig. 4Expression levels of OsPGIP genes in rice cultivars with different resistance to sheath blight at different growth stages
A: seedling stage; B: adult-plant stage; C: spike stage.
�ֱ��Խ���ˮ���ƿݲ��������º��ڹ������������澳�ͷ������澳, ����OsPGIP�����ڲ�ͬ�澳�������ڲ�ͬˮ��Ʒ��ֲ��Ҷ���еı���ˮƽ���������, �ڵ���������, �߸�Ʒ��Lemont�е�OsPGIP2��OsPGIP3����Ϳ���Ʒ��YSBR1�е�OsPGIP6����ı������������µ�, ������Ʒ���е����л�����ϵ�����, ��YSBR1�е�OsPGIP4��OsFOR1������������(��Ա������ֱ�Ϊ8.55��14.53), �Ҹ����������ƽ���ϵ������Կ���Ʒ��YSBR1�е�Ϊ���; �ڹ����, ��OsPGIP2������Lemont���쵾3���б����������Ա仯��(��Ա������ֱ�Ϊ0.97��1.10), ������OsPGIP�����ڸ�Ʒ���о������ϵ�����, ���Կ���Ʒ��YSBR1�еı������ϵ��������, ƽ��Ϊ9.09, OsPGIP2�ı������Ƕ��յ�18.11��; �����ƿݲ�����, ��ͬ����ı���ˮƽ�����ϵ����µ�, ��������ȶ�����˵, �������ϵ�����, �ر���OsFOR1�����OsPGIP6����, ����ϵ�����Ϊ9.53��6.66����Щ���������ǰ���о�OsPGIP1������������Ľ��һ��, �����澳�����´����OsPGIP�����ͨ����������ı���ˮƽ������ֲ�����ӿ����ԡ�
ͼ5
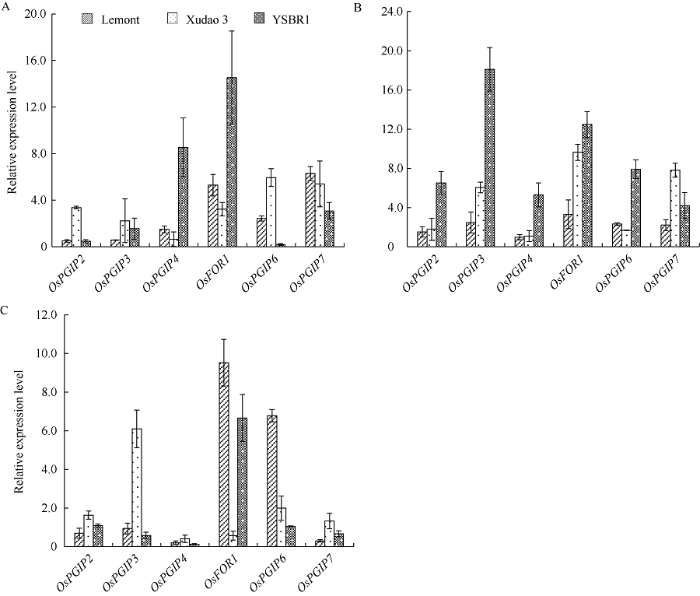 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5��ͬ�澳�����¿������ƿݲ�ˮ��Ʒ��Ҷ���и�OsPGIP����ı�����
A: ����; B: �ڹ�; C: ���֡�
Fig. 5Expression levels of OsPGIP genes in rice cultivars with different resistance to sheath blight under different stress condition.
A: low temperature; B: dark; C: inoculation.
3 ����
PGIP��Ϊֲ��ϸ���ڵ���Ҫ��ɲ���, �����������ڴ��ģ������, ֻ������������ʮ���������Լ������ʽ����, һ��Ϊ2~16��[26,27,28]������Щ������, �����Ͻ�2��AtPGIP������, ������δ�����ں��ӵı���, �����ǿ���ת���ӵIJ�������»���ʧ��[26,28]��ˮ����Ŀǰ������7��PGIP����, ����DNA��cDNA�������, 7��OsPGIP��������ں���, Ҳ������ת���Ӳ�������������7��OsPGIP��, ��Щ���ƶȽϸ��������̱���ֲ��PGIP��ϵͳ������������ʱ����ͬһȺ��, ����ЩOsPGIP���Ŵ�����ϴ�, ���������ڲ�ͬ�����Ȼ���Liu��[10]����ˮ��PGIP������ֲ����Դ��PGIP����ϵͳ��������, ��Ϊ���к̱��ơ����ơ�ʮ�ֻ��ƺ���ľ�����ɸ��Ա�������Ϊһ��, ˮ���е�7��OsPGIP��С�����ס������ȵ�PGIP�߶����ơ��������о���ѡ����������������������Դ��51��PGIP, ��ʮ�ֻ��ƽ�ѡ�����Ͳ˺����Ͻ�, ����Ҳ��ѡ���˴�ͬ����ͬ�ֵ����ֲ˶��������������Ŵ�, ��NCBI��������ȷ����ΪPGIP�ĵ������н��ж����бȶ���ϵͳ������������ʱ, ���Ƿ���, ������ͬ�����������Դ��PGIP�й�Ϊһ�������, ����������PGIP���������ֵ�PGIP�������Ÿ��ߵ������ԡ���Щ���˵��, ��ʹ����ͬ��������ֵ�PGIPҲ�п��������Բ�ͬ������, ����ͬ�����е�PGIP�������Թ�ͬ���ȵĿ�����, ������һ������Խ�С��ֲ��ϸ�����Ǹ߶ȶ�̬�İ���ṹ, ��ֲ�������������ܵ��澳в��ʱ, ��ɲ��ϱ��ع�, ��Ϊϸ���ڵ�һ����, PGIP���������ڵ���������N-�˺�LRR���й�[29]�������LRRƬ�ι��ɵİ���, ���д����ĸ����, �������벡��PG�Ļ���[30,31,32]������ͬ����PGIP������ͬһ���ֵIJ�ͬPGIP�Բ�ͬ��ԭ����ͬһ��ԭ����ͬPG��������������ϴ�, ��������ʽ����(�羺�������ơ��Ǿ��������ƺͻ���͵�) ��, �����ʱ������������С��۵���ʽ�������ɺͿռ�ṹ�ȶ��Ի�������Ӱ��[11,33-35]��ֻ����ĿǰΪֹ, ���ڶ�PGIP����ṹ���о����dz���, ����PGIP-PG�����ľ���ṹģ���ǻ�δ������, ��˶���PGIP���ܼ�����PG�������о��ͣ���ڽ���������Ϣѧ�����Ľ���ϡ�Ŀǰ���е��ṹԤ������������ܶ�, ��SWISS- MODEL��CHPmodels��SCRATCH��Modeller��3D-JIGSAW��EsyPred3D��RaptorX��Hhpred�ȡ����ܲ�ͬ����Ԥ��Ľ��������ȫ��ͬ, ��������������������С�Ƕ�X���䷨Ԥ��PvPGIP2��FpPG�Ļ���ʱ, ���ַ����õ��Ľ���ֱ���PvPGIP2�İ�����FvPG��N-�����Ħ�-�۵�����C-�˼�������϶������, ����Щ�������ͨ����������Ķ���ͻ�������֤[31,36-37]������, ���е����ʿռ�ṹ�����ܡ�����, �Լ������������ĶԽ�Ԥ����������о������ܼ��������ƵĻ���, ֻ����Ԥ�����ʱҪ���ж��ַ����ıȽ�, ����Ԥ��ģ�ͼ������κ����ۡ��籾�о��зֱ����SWISS-MODEL��CHPmodels��SCRATCH��������Ԥ����OsPGIP�Ŀռ�ṹ, SCRATCHԤ���ģ����Ȼ�������о��߶�ֲ��PGIP�ռ�ṹ����֪, ���Ҳ������֮����һ���о�OsPGIP�Ľṹ�����ʡ����ܼ���PG�Ļ����ȡ�
PGIP�������������ͱ�������, �������ܵ�������Դ���ص��յ�����, ��ֲ�D�ء���е���ˡ�����ͷ������澳�ȡ��˶��е�4��PvPGIP����ֲ�����B2����Ⱥһ��50 kb��������, ��Щ�����ں�����ˮƽ�ϵ������Դ���80%, ˵������Դ�ڹ�ͬ�����Ȼ���, �����ǵı���ȴ���켫��, ������Ҷ�������ᡢ���ͼ���PvPGIP1�����������, ��PvPGIP2ȴ�����������б���, PvPGIP3��PvPGIP4��ֻ�ڸ���������[16,18]���ܲ�ԭ����Ⱦ��, ��������������Ի����ͷ����Ի������ֱ���PGIP�����ת¼����������ӳ��ڼ����ض���λ�ֲ�����[16,38-39]����ù��������Ⱦ���������Ͻ�������AtPGIP�����ϵ�����, ��ȴ��ͨ����ͬ���źŴ���;��[26]����Щ���������, ֲ����PGIP����ı���ˮƽ�������������ڡ���λ�������Դ������Ӱ�졣ˮ���е�7��OsPGIP�����ڲ�ͬ��֯�����еı���ˮƽ�ͬ; �ڳ�ù�ء������غ������ᴦ����, �̵�Ʒ������63�г�OsPGIP2�����ڼ����ش�����������µ���, ��������ı������������ϵ�; ���ھ���Ʒ���л�11��, ʹ�������ᡢܿ����������ù�ء������ء������ء��������ˮ���ᴦ����, ����ʱ�������, �������OsPGIP����ı����������ϵ�[19]������ͬ����ˮ����, ���о�����, ���ƿݲ��߲�ͬ���Ե�ˮ��Ʒ��������������������澳ʱ, ������OsPGIP����ı�����������������ġ������������Щ����ı�����ؼ��벡ԭ��PG�����Ļ���, ��Ϊ����ˮ�����������е�Ӧ�õ춨������
4 ����
��¡��ˮ��OsPGIP��������OsPGIP1���ʣ��6������, �����ǽ�����ϵͳ��������, Ԥ�������������һ�����������ṹ������ѧ����, �����˸�������ˮ����ͬ�����ں��澳�µı�������, Ϊ��ʾOsPGIP������岻ͬ��Ա�ڲ�ͬ�澳�����µ�DZ�ڹ��ܺ���ˮ�����������е�Ӧ�õ춨�˻������ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
DOI:10.1111/ppa.1956.5.issue-4URL [��������: 1]
DOI:10.1134/s0006297908100015URLPMID:18991551 [��������: 1]

It is generally believed that plants
DOI:10.1111/j.1438-8677.2008.00175.xURLPMID:19689781 [��������: 1]
Polygalacturonase-inhibiting proteins (PGIPs) are plant cell wall proteins that specifically inhibit the activity of endopolygalacturonases (PGs) produced by fungi during the infection process. The interaction with PGIPs limits the destructive potential of PGs and may trigger plant defence responses through the release of elicitor active oligogalacturonides. In order to pinpoint the residues of PvPGIP2 from Phaseolus vulgaris involved in the interaction with PGs, we used site-directed mutagenesis to mutate the residues D131, D157 and D203, and tested for the inhibitory activity of the mutant proteins expressed in Pichia pastoris against Fusarium phyllophilum and Aspergillus niger PGs. Here, we report that mutation of these residues affects the inhibition capacity of PvPGIP2 against F. phyllophilum PG.
DOI:10.1111/j.1365-313X.2007.03398.xURLPMID:18088306 [��������: 1]
In addition to the role of the cell wall as a physical barrier against pathogens, some of its constituents, such as pectin-derived oligogalacturonides (OGA), are essential components for elicitation of defence responses. To investigate how modifications of pectin alter defence responses, we expressed the fruit-specific Fragaria x ananassa pectin methyl esterase FaPE1 in the wild strawberry Fragaria vesca. Pectin from transgenic ripe fruits differed from the wild-type with regard to the degree and pattern of methyl esterification, as well as the average size of pectin polymers. Purified oligogalacturonides from the transgenic fruits showed a reduced degree of esterification compared to oligogalacturonides from wild-type fruits. This reduced esterification is necessary to elicit defence responses in strawberry. The transgenic F. vesca lines had constitutively activated pathogen defence responses, resulting in higher resistance to the necrotropic fungus Botrytis cinerea. Further studies in F. vesca and Nicotiana benthamiana leaves showed that the elicitation capacity of the oligogalacturonides is more specific than previously envisaged.
DOI:10.3389/fpls.2013.00030URLPMID:23440336 [��������: 1]
Systemic acquired resistance (SAR) is an inducible defense mechanism in plants that confers enhanced resistance against a variety of pathogens. SAR is activated in the uninfected systemic (distal) organs in response to a prior (primary) infection elsewhere in the plant. SAR is associated with the activation of salicylic acid (SA) signaling and the priming of defense responses for robust activation in response to subsequent infections. The activation of SAR requires communication by the primary infected tissues with the distal organs. The vasculature functions as a conduit for the translocation of factors that facilitate long-distance intra-plant communication. In recent years, several metabolites putatively involved in long-distance signaling have been identified. These include the methyl ester of SA (MeSA), the abietane diterpenoid dehydroabietinal (DA), the dicarboxylic acid azelaic acid (AzA), and a glycerol-3-phosphate (G3P)-dependent factor. Long-distance signaling by some of these metabolites also requires the lipid-transfer protein DIR1 (DEFECTIVE IN INDUCED RESISTANCE 1). The relative contribution of these factors in long-distance signaling is likely influenced by environmental conditions, for example light. In the systemic leaves, the AGD2-LIKE DEFENSE RESPONSE PROTEIN1 (ALD1)-dependent production of the lysine catabolite pipecolic acid (Pip), FLAVIN-DEPENDENT MONOOXYGENASE1 (FMO1) signaling, as well as SA synthesis and downstream signaling are required for the activation of SAR. This review summarizes the involvement and interaction between long-distance SAR signals and details the recently discovered role of Pip in defense amplification and priming that allows plants to acquire immunity at the systemic level. Recent advances in SA signaling and perception are also highlighted.
DOI:10.1186/s12870-016-0959-1URLPMID:28103793 [��������: 1]
BACKGROUND: Oligogalacturonides (OGs) are important components of damage-associated molecular pattern (DAMP) signaling and influence growth regulation in plants. Recent studies have focused on the impact of long OGs (degree of polymerization (DP) from 10-15), demonstrating the induction of plant defense signaling resulting in enhanced defenses to necrotrophic pathogens. To clarify the role of trimers (trimeric OGs, DP3) in DAMP signaling and their impact on plant growth regulation, we performed a transcriptomic analysis through the RNA sequencing of Arabidopsis thaliana exposed to trimers. RESULTS: The transcriptomic data from trimer-treated Arabidopsis seedlings indicate a clear activation of genes involved in defense signaling, phytohormone signaling and a down-regulation of genes involved in processes related to growth regulation and development. This is further accompanied with improved defenses against necrotrophic pathogens triggered by the trimer treatment, indicating that short OGs have a clear impact on plant responses, similar to those described for long OGs. CONCLUSIONS: Our results demonstrate that trimers are indeed active elicitors of plant defenses. This is clearly indicated by the up-regulation of genes associated with plant defense signaling, accompanied with improved defenses against necrotrophic pathogens. Moreover, trimers simultaneously trigger a clear down-regulation of genes and gene sets associated with growth and development, leading to stunted seedling growth in Arabidopsis.
DOI:10.1073/pnas.92.10.4095URLPMID:11607534 [��������: 1]
Jasmonic acid, synthesized from linolenic acid (the octadecanoid pathway), has been proposed to be part of a signal transduction pathway that mediates the induction of defensive genes in plants in response to oligouronide and polypeptide signals generated by insect and pathogen attacks. We report here that the induction of proteinase inhibitor accumulation in tomato leaves by plant-derived oligogalacturonides and fungal-derived chitosan oligosaccharides is severely reduced by two inhibitors (salicylic acid and diethyldi-thiocarbamic acid) of the octadecanoid pathway, supporting a role for the pathway in signaling by oligosaccharides. Jasmonic acid levels in leaves of tomato plants increased several fold within 2 hr after supplying the polypeptide systemin, oligogalacturonides, or chitosan to the plants through their cut stems, as expected if they utilize the octadecanoid pathway. The time course of jasmonic acid accumulation in tomato leaves in response to wounding was consistent with its proposed role in signaling proteinase inhibitor mRNA and protein synthesis. The cumulative evidence supports a model for the activation of defensive genes in plants in response to insect and pathogen attacks in which various elicitors generated at the attack sites activate the octadecanoid pathway via different recognition events to induce the expression of defensive genes in local and distal tissues of the plants.
DOI:10.1146/annurev.phyto.39.1.313URLPMID:11701868 [��������: 1]
Polygalacturonase-inhibiting proteins (PGIPs) are extracellular plant proteins capable of inhibiting fungal endopolygalacturonases (PGs). Plants have evolved different PGIPs with specific recognition abilities against the many PGs produced by fungi. The genes encoding PGIPs are organized into families, and different members of each family may encode proteins with nearly identical characteristics but different specificities and regulation. PGIPs are typically induced by pathogen infection and stress-related signals. The recognition ability of PGIPs resides in their LRR (leucine-rich repeat) structure, where solvent-exposed residues in the beta-strand/beta-turn motifs of the LRRs are determinants of specificity. Manipulation of the primary structure of PGIPs is expected to generate more efficient PGIPs with novel recognition specificities to protect crop plants against pathogens.
DOI:10.1016/S0981-9428(00)01228-6URL [��������: 1]
DOI:10.1038/srep39840URLPMID:28079053 [��������: 2]
Polygalacturonase-inhibiting protein (PGIP), belonging to a group of plant defence proteins, specifically inhibits endopolygalacturonases secreted by pathogens. Herein, we showed that purified GhPGIP1 is a functional inhibitor of Verticillium dahliae and Fusarium oxysporum f. sp. vasinfectum, the two fungal pathogens causing cotton wilt. Transcription of GhPGIP1 was increased in cotton upon infection, wounding, and treatment with defence hormone and H2O2. Resistance by GhPGIP1 was examined by its virus-induced gene silencing in cotton and overexpression in Arabidopsis. GhPGIP1-silenced cotton was highly susceptible to the infections. GhPGIP1 overexpression in transgenic Arabidopsis conferred resistance to the infection, accompanied by enhanced expression of pathogenesis-related proteins (PRs), isochorismate synthase 1 (ICS1), enhanced disease susceptibility 1 (EDS1), and phytoalexin-deficient 4 (PAD4) genes. Transmission electron microscopy revealed cell wall alteration and cell disintegration in plants inoculated with polygalacturonase (PGs), implying its role in damaging the cell wall. Docking studies showed that GhPGIP1 interacted strongly with C-terminal of V. dahliae PG1 (VdPG1) beyond the active site but weakly interacted with C-terminal of F. oxysporum f. sp. vasinfectum (FovPG1). These findings will contribute towards the understanding of the roles of PGIPs and in screening potential combat proteins with novel recognition specificities against evolving pathogenic factors for countering pathogen invasion.
DOI:10.3389/fpls.2015.00146URLPMID:25852708 [��������: 2]
Polygalacturonase inhibiting proteins (PGIPs) are cell wall proteins that inhibit the pectin-depolymerizing activity of polygalacturonases secreted by microbial pathogens and insects. These ubiquitous inhibitors have a leucine-rich repeat structure that is strongly conserved in monocot and dicot plants. Previous reviews have summarized the importance of PGIP in plant defense and the structural basis of PG-PGIP interaction; here we update the current knowledge about PGIPs with the recent findings on the composition and evolution of pgip gene families, with a special emphasis on legume and cereal crops. We also update the information about the inhibition properties of single pgip gene products against microbial PGs and the results, including field tests, showing the capacity of PGIP to protect crop plants against fungal, oomycetes and bacterial pathogens.
DOI:10.1186/s12284-019-0318-6URLPMID:31359264 [��������: 2]
BACKGROUND: An economic strategy to control plant disease is to improve plant defense to pathogens by deploying resistance genes. Plant polygalacturonase inhibiting proteins (PGIPs) have a vital role in plant defense against phytopathogenic fungi by inhibiting fungal polygalacturonase (PG) activity. We previously reported that rice PGIP1 (OsPGIP1) inhibits PG activity in Rhizoctonia solani, the causal agent of rice sheath blight (SB), and is involved in regulating resistance to SB. RESULT: Here, we report that OsPGIP2, the protein ortholog of OsPGIP1, does not possess PGIP activity; however, a few amino acid substitutions in a derivative of OsPGIP2, of which we provide support for L233F being the causative mutation, appear to impart OsPGIP2 with PG inhibition capability. Furthermore, the overexpression of mutated OsPGIP2(L233F) in rice significantly increased the resistance of transgenic lines and decreased SB disease rating scores. OsPGIP2(L233F) transgenic lines displayed an increased ability to reduce the tissue degradation caused by R. solani PGs as compared to control plants. Rice plants overexpressing OsPGIP2(L233F) showed no difference in agronomic traits and grain yield as compared to controls, thus demonstrating its potential use in rice breeding programs. CONCLUSIONS: In summary, our results provide a new target gene for breeding SB resistance through genome-editing or natural allele mining.
DOI:10.1007/s10265-012-0515-5URL [��������: 1]
Regulation of defense in plants is a complex process mediated by various signaling pathways. Promoter analysis of defense-related genes is useful to understand these signaling pathways involved in regulation. To this end, the regulation of the polygalacturonase-inhibiting protein encoding gene from Vitis vinifera L. (Vvpgip1) was analyzed with regard to expression pattern and induction profile as well as the promoter in terms of putative regulatory elements present, core promoter size and the start of transcription. Expression of Vvpgip1 is tissue-specific and developmentally regulated. Vvpgip1 expression was induced in response to auxin, salicylic acid and sugar treatment, wounding and pathogen infection. The start of transcription was mapped to 17?bp upstream of the ATG and the core promoter was mapped to the 137?bp upstream of the ATG. Fructose- and Botrytis responsiveness were identified in the region between positions �C3.1 and �C1.5?kb. The analyses showed induction in water when the leaves were submersed and this response and the response to wounding mapped to the region between positions �C1.1 and �C0.1?kb. In silico analyses revealed putative cis-acting elements in these areas that correspond well to the induction stimuli tested.]]>
DOI:10.1186/s12870-014-0189-3URLPMID:25034494 [��������: 1]
BACKGROUND: Polygalacturonase-inhibiting proteins (PGIPs) are leucine-rich repeat (LRR) plant cell wall glycoproteins involved in plant immunity. They are typically encoded by gene families with a small number of gene copies whose evolutionary origin has been poorly investigated. Here we report the complete characterization of the full complement of the pgip family in soybean (Glycine max [L.] Merr.) and the characterization of the genomic region surrounding the pgip family in four legume species. RESULTS: BAC clone and genome sequence analyses showed that the soybean genome contains two pgip loci. Each locus is composed of three clustered genes that are induced following infection with the fungal pathogen Sclerotinia sclerotiorum (Lib.) de Bary, and remnant sequences of pgip genes. The analyzed homeologous soybean genomic regions (about 126 Kb) that include the pgip loci are strongly conserved and this conservation extends also to the genomes of the legume species Phaseolus vulgaris L., Medicago truncatula Gaertn. and Cicer arietinum L., each containing a single pgip locus. Maximum likelihood-based gene trees suggest that the genes within the pgip clusters have independently undergone tandem duplication in each species. CONCLUSIONS: The paleopolyploid soybean genome contains two pgip loci comprised in large and highly conserved duplicated regions, which are also conserved in bean, M. truncatula and C. arietinum. The genomic features of these legume pgip families suggest that the forces driving the evolution of pgip genes follow the birth-and-death model, similar to that proposed for the evolution of resistance (R) genes of NBS-LRR-type.
[��������: 1]
URL [��������: 3]
The expression analysis of the four polygalacturonase-inhibiting protein (Pgip) genes composing the bean (Phaseolus vulgaris L.) Pgip family showed a pattern of transcriptional variation in young leaves, hypocotyls, roots and pods with Pvpgip1 not expressed, Pvpgip2 expressed in all organs, Pvpgip3 and Pvpgip4 poorly expressed in roots. We compared also transcript accumulation of the four Pvpgip genes during infection of bean plants with the fungal pathogens Botrytis cinerea, Sclerotinia sclerotiorum and Colletotrichum demuthianum. qRT-PCR analyses showed that the transcript level of Pvpgip1, Pvpgip2 and Pvpgip3 increases significantly following fungal infection, whereas Pvpgip4 remains unchanged. The level of induction was different between the three genes, Pvpgip2 exhibiting the strongest transcript accumulation. The induction pattern was similar in the pathosystems bean-S. sclerotiorum, bean-B. cinerea, and in the compatible interaction bean-C. lindemuthianum, with a maximum of transcript accumulation in the late stage of infection. Instead, in the incompatible interaction bean-C. lindemuthianum, Pvpgip1, Pvpgip2 and Pvpgip3 showed an early and transient transcript accumulation, with Pvpgip2 exhibiting an earlier and higher induction. These results extend previous analyses of the whole Pup gip transcript and provide additional evidences of the relevant role of PvPGIP2 in plant defence.
DOI:10.1007/s004250050308URLPMID:9637069 [��������: 1]
Polygalacturonase-inhibiting proteins (PGIPs), leucine-rich repeat (LRR) proteins evolutionarily related to several plant resistance genes, bind to and regulate the action of fungal endopolygalacturonases. In Phaseolus vulgaris L., PGIPs are encoded by a gene family comprising at least five members. As a start for a systematic analysis of the regulation of the pgip family, we have analysed the ability of the promoter of the bean gene pgip-1 to direct expression of beta-glucuronidase (GUS) in transfected tobacco protoplasts, microbombarded bean and tobacco leaves, and transgenic tobacco plants. In protoplasts, the pgip-1 gene region from nucleotide (nt) -2004 to nt +27 directed a level of expression that was as high as that directed by the cauliflower mosaic virus (CaMV) 35S promoter and could not be further induced by elicitor treatment; alteration of the region immediately following the TATAA sequence at nt -29 abolished expression. Upon stable integration into tobacco plants of the pgip-1 promoter-GUS construct, as well as of a -394 deletion, expression was detected for both constructs mainly in the stigma and, to a lesser extent, in the anthers and in the conductive vascular tissue. The promoter responded to wounding but not to oligogalacturonides, fungal glucan, salicylic acid, cryptogein, or pathogen infection. This expression pattern does not mirror that of the whole pgip gene family.
DOI:10.1104/pp.104.044644URLPMID:15299124 [��������: 2]
Polygalacturonase-inhibiting proteins (PGIPs) are extracellular plant inhibitors of fungal endopolygalacturonases (PGs) that belong to the superfamily of Leu-rich repeat proteins. We have characterized the full complement of pgip genes in the bean (Phaseolus vulgaris) genotype BAT93. This comprises four clustered members that span a 50-kb region and, based on their similarity, form two pairs (Pvpgip1/Pvpgip2 and Pvpgip3/Pvpgip4). Characterization of the encoded products revealed both partial redundancy and subfunctionalization against fungal-derived PGs. Notably, the pair PvPGIP3/PvPGIP4 also inhibited PGs of two mirid bugs (Lygus rugulipennis and Adelphocoris lineolatus). Characterization of Pvpgip genes of Pinto bean showed variations limited to single synonymous substitutions or small deletions. A three-amino acid deletion encompassing a residue previously identified as crucial for recognition of PG of Fusarium moniliforme was responsible for the inability of BAT93 PvPGIP2 to inhibit this enzyme. Consistent with the large variations observed in the promoter sequences, reverse transcription-PCR expression analysis revealed that the different family members differentially respond to elicitors, wounding, and salicylic acid. We conclude that both biochemical and regulatory redundancy and subfunctionalization of pgip genes are important for the adaptation of plants to pathogenic fungi and phytophagous insects.
DOI:10.1007/s00299-012-1239-7URL [��������: 5]
All the seven Ospgip genes showed variable expression patterns in Minghui 63 and their expressions were regulated by different phytohormone treatments or fungal infection in Minghui 63 and Zhonghua 11.]]>
DOI:10.1023/B:PLAN.0000006940.89955.f1URL [��������: 3]
We have isolated a cDNA clone, OsFOR1, from the immature panicles of rice. The OsFOR1 (Oryza sativa floral organ regulator 1) gene encodes a protein that contains a leucine-rich repeat (LRR) domain. This domain comprises 10 tandem repeats of a canonical 24-amino acid LRR sequence. The structure and the number of LRRs for OsFOR1 are similar to those of polygalacturonase-inhibiting proteins (PGIPs) from various other plant species. Moreover, the OsFOR1 recombinant protein, when fused to maltose-binding protein (MBP), shows PGIP activity against the Aspergillus niger polygalacturonase. OsFOR1 is highly expressed in the calli and immature and mature panicles, while detectable at only low levels in seedling roots and mature stems. Insitu hybridization experiments showed the transcripts of OsFOR1 are present in young spikelet primordia and in almost all of the young floral organs. Transgenic approaches were used to study invivo functioning. Antisense expression of OsFOR1 resulted in an increase in the numbers of floral organs, including the stamen, carpel, palea/lemma, stigma, and lodicule. OsFOR1 transcript was not detected in the frizzy panicle mutant, which is defective in its spikelet formation but normal in inflorescence-meristem initiation and maintenance. Therefore, we suggest that OsFOR1 plays a role in the formation and/or maintenance of floral organ primordia.]]>
DOI:10.1007/s00122-006-0378-zURL [��������: 2]
Polygalacturonase-inhibiting proteins (PGIPs) are leucine-rich repeat (LRR) proteins involved in plant defence. A number of PGIPs have been characterized from dicot species, whereas only a few data are available from monocots. Database searches and genome-specific cloning strategies allowed the identification of four rice (Oryza sativa L.) and two wheat (Triticum aestivum L.) Pgip genes. The rice Pgip genes (Ospgip1, Ospgip2, Ospgip3 and Ospgip4) are distributed over a 30kbp region of the short arm of chromosome 5, whereas the wheat Pgip genes, Tapgip1 and Tapgip2, are localized on the short arm of chromosome 7B and 7D, respectively. Deduced amino acid sequences show the typical LRR modular organization and a conserved distribution of the eight cysteines at the N- and C-terminal regions. Sequence comparison suggests that monocot and dicot PGIPs form two separate clusters sharing about 40% identity and shows that this value is close to the extent of variability observed within each cluster. Gene-specific RT-PCR and biochemical analyses demonstrate that both Ospgips and Tapgips are expressed in the whole plant or in a tissue-specific manner, and that OsPGIP1, lacking an entire LRR repeat, is an active inhibitor of fungal polygalacturonases. This last finding can contribute to define the molecular features of PG–PGIP interactions and highlights that the genetic events that can generate variability at the Pgip locus are not only limited to substitutions or small insertions/deletions, as so far reported, but can also involve variation in the number of LRRs.]]>
DOI:10.1023/A:1017213630972URL [��������: 1]
DOI:10.1094/PDIS-03-15-0305-REURLPMID:30694142 [��������: 1]
Rice sheath blight (SB), caused by necrotrophic pathogen Rhizoctonia solani, is one of the most destructive rice diseases, and no major resistance genes are available. Polygalacturonase-inhibiting proteins (PGIP) are extracellular leucine-rich repeat proteins and play important roles in plant defense against different pathogenic fungi by counteracting secreted fungal polygalacturonases (PG). However, the role of PGIP in conferring resistance to rice SB remains to be thoroughly investigated. Here, we showed that OsPGIP1 is capable of inhibiting PG derived from R. solani. Our real-time reverse-transcription polymerase chain reaction results indicated that resistant rice 'YSBR1' and 'Jasmine 85' express significantly higher levels of OsPGIP1 than susceptible 'Lemont'. Our results also show that OsPGIP1 is most highly expressed at the late tillering stage in the sheath of YSBR1, coinciding with the critical stage of SB development in field. More importantly, the OsPGIP1 level is highly elevated by inoculation with R. solani in resistant cultivars but not in susceptible Lemont. Overexpression of OsPGIP1 significantly increased rice resistance to SB and inhibited tissue degradation caused by R. solani-secreted PG. Furthermore, OsPGIP1 overexpression did not affect rice agronomic traits or yield components. Together, our results not only demonstrate the important role of OsPGIP1 in combatting the rice SB disease but also provide a new avenue to the improvement of rice SB resistance by manipulating an endogenous gene.
DOI:10.1007/s00425-016-2480-zURLPMID:26945855 [��������: 1]
MAIN CONCLUSION: OsPGIP4 overexpression enhances resistance to bacterial leaf streak in rice. Polygalacturonase-inhibiting proteins are thought to play important roles in the innate immunity of rice against fungi. Here, we show that the chromosomal location of OsPGIP4 coincides with the major bacterial leaf streak resistance quantitative trait locus qBlsr5a on the short arm of chromosome 5. OsPGIP4 expression was up-regulated upon inoculation with the pathogen Xanthomonas oryzae pv. oryzicola strain RS105. OsPGIP4 overexpression enhanced the resistance of the susceptible rice variety Zhonghua 11 to RS105. In contrast, repressing OsPGIP4 expression resulted in an increase in disease lesions caused by RS105 in Zhonghua 11 and in Acc8558, a qBlsr5a resistance donor. More interestingly, upon inoculation, the activated expression of pathogenesis-related genes was attenuated for those genes involved in the salicylic acid pathway, while the activated expression of jasmonic acid pathway markers was increased in the overexpression lines. Our results not only provide the first report that rice PGIP could enhance resistant against a bacterial pathogen but also indicate that OsPGIP4 is a potential component of the qBlsr5a locus for bacterial leaf streak in rice.
DOI:10.3969/j.issn.1001��7216.2012.05.018URL [��������: 1]
��ˮ��������DNA��������1 184 bp��Ospgip1����Ƭ�Ρ���Ƭ�ΰ���930 bp�����������Ķ�����ֹ������ΪTAA�����ں��ӡ�RT��PCR���������Ospgip1������ˮ���������ƿݲ�Ʒ���о��ܱ������ͬ�����ڡ���ͬ��λ�������в��졣ʵʱPCR�����ʾ������Ʒ��YSBR1���и�Ʒ��Jasmine 85��Ospgip1�ı�����Ҫ���Ը��ڸв�Ʒ��Lemont������ƻ������������ǿ���Ʒ�ֻ��Ǹв�Ʒ�ֵ�Ospgip1��������������ߣ��ƿݲ�����Ⱦʹ�ÿ���Ʒ��Ospgip1������������ӣ����Ըв�Ʒ��Ӱ�첻��
[��������: 1]
DOI:10.1105/tpc.005165URLPMID:12509524 [��������: 3]
Polygalacturonase-inhibiting proteins (PGIPs) are plant proteins that counteract fungal polygalacturonases, which are important virulence factors. Like many other plant defense proteins, PGIPs are encoded by gene families, but the roles of individual genes in these families are poorly understood. Here, we show that in Arabidopsis, two tandemly duplicated PGIP genes are upregulated coordinately in response to Botrytis cinerea infection, but through separate signal transduction pathways. AtPGIP2 expression is mediated by jasmonate and requires COI1 and JAR1, whereas AtPGIP1 expression is upregulated strongly by oligogalacturonides but is unaffected by salicylic acid, jasmonate, or ethylene. Both AtPGIP1 and AtPGIP2 encode functional inhibitors of polygalacturonase from Botrytis, and their overexpression in Arabidopsis significantly reduces Botrytis disease symptoms. Therefore, gene duplication followed by the divergence of promoter regions may result in different modes of regulation of similar defensive proteins, thereby enhancing the likelihood of defense gene activation during pathogen infection.
DOI:10.1007/s00425-008-0733-1URL [��������: 1]
Most plants encode a limited set of polygalacturonase inhibitor (PGIP) genes that may be involved in aspects of plant development, but more importantly in the inactivation of polygalacturonases (PG) secreted by pathogens. Previously, we characterized two Brassica napus PGIP genes, BnPgip1 and BnPgip2, which were differentially expressed in response to pathogen infection and wounding. Here we report that the B. napus genome encodes a set of at least 16 PGIP genes that are similar to BnPgip1 or BnPgip2. This is the largest Pgip gene family reported to date. Comparison of the BnPGIPs revealed several sites within the xxLxLxx region of leucine rich repeats that form β-sheets along the interacting face of the PGIP that are hypervariable and represent good candidates for generating PGIP diversity. Characterization of the regulatory regions and RT-PCR studies with gene-specific primers revealed that individual genes were differentially responsive to pathogen infection, mechanical wounding and signaling molecules. Many of the BnPgip genes responded to infection by the necrotic pathogen, Sclerotinia sclerotiorum; however, these genes were also induced either by jasmonic acid, wounding and salicylic acid or some combination thereof. The large number of PGIPs and the differential manner in which they are regulated likely ensures that B. napus can respond to attack from a broad spectrum of pathogens and pests.]]>
DOI:10.1007/s00122-008-0719-1URL [��������: 2]
DOI:10.3389/fpls.2015.00660URLPMID:26379688 [��������: 1]
The plant endomembrane system is massively involved in the synthesis, transport and secretion of cell wall polysaccharides and proteins; however, the molecular mechanisms underlying trafficking toward the apoplast are largely unknown. Besides constitutive, the existence of a regulated secretory pathway has been proposed. A polygalacturonase inhibitor protein (PGIP2), known to move as soluble cargo and reach the cell wall through a mechanism distinguishable from default, was dissected in its main functional domains (A, B, C, D), and C sub-fragments (C1-10), to identify signals essential for its regulated targeting. The secretion patterns of the fluorescent chimeras obtained by fusing different PGIP2 domains to the green fluorescent protein (GFP) were analyzed. PGIP2 N-terminal and leucine-rich repeat domains (B and C, respectively) seem to operate as holding/releasing signals, respectively, during PGIP2 transit through the Golgi. The B domain slows down PGIP2 secretion by transiently interacting with Golgi membranes. Its depletion leads, in fact, to the secretion via default (Sp2-susceptible) of the ACD-GFP chimera faster than PGIP2. Depending on its length (at least the first 5 leucine-rich repeats are required), the C domain modulates B interaction with Golgi membranes allowing the release of chimeras and their extracellular secretion through a Sp2 independent pathway. The addition of the vacuolar sorting determinant Chi to PGIP2 diverts the path of the protein from cell wall to vacuole, suggesting that C domain is a releasing rather than a cell wall sorting signal.
DOI:10.1021/bi0017632URLPMID:11148052 [��������: 1]
A detailed analysis of the secondary structure has been carried out on the polygalacturonase-inhibiting protein (PGIP) from Phaseolus vulgaris, a leucine-rich repeat (LRR) protein present in the cell wall of many plants. Far-UV CD and infrared spectroscopies coupled to constrained secondary structure prediction methods indicated the presence of 12 alpha- and 12 beta-segments, thus allowing a schematic representation of three domains of the protein, namely, the central LRR region and the two cysteine-rich flanking domains. Peptides from endoproteinase-degraded PGIP were analyzed by mass spectrometry, and four disulfide bonds were identified. Mass spectrometric analysis in combination with glycosidase treatments revealed two N-linked oligosaccharides located on Asn 64 and Asn 141. The main structure resembled the typical complex plant N-glycan consisting of a core pentasaccharide beta1,2-xylosylated, carrying an alpha1,3-fucose linked to the innermost N-acetylglucosamine and one outer arm N-acetylglucosamine residue. The schematic representation of PGIP structural domains is discussed in the framework of the structure and function of LRR proteins.
DOI:10.1371/journal.pone.0080610URLPMID:24260434 [��������: 2]
Polygalacturonases (PGs) are secreted by phytopathogenic fungi to degrade the plant cell wall homogalacturonan during plant infection. To counteract Pgs, plants have evolved polygalacturonase-inhibiting proteins (PGIPs) that slow down fungal infection and defend cell wall integrity. PGIPs favour the accumulation of oligogalacturonides, which are homogalacturonan fragments that act as endogenous elicitors of plant defence responses. We have previously shown that PGIP2 from Phaseolus vulgaris (PvPGIP2) forms a complex with PG from Fusarium phyllophilum (FpPG), hindering the enzyme active site cleft from substrate. Here we analyse by small angle X-ray scattering (SAXS) the interaction between PvPGIP2 and a PG from Colletotrichum lupini (CluPG1). We show a different shape of the PG-PGIP complex, which allows substrate entry and provides a structural explanation for the different inhibition kinetics exhibited by PvPGIP2 towards the two isoenzymes. The analysis of SAXS structures allowed us to investigate the basis of the inability of PG from Fusarium verticilloides (FvPG) to be inhibited by PvPGIP2 or by any other known PGIP. FvPG is 92.5% identical to FpPG, and we show here, by both loss- and gain-of-function mutations, that a single amino acid site acts as a switch for FvPG recognition by PvPGIP2.
URL [��������: 1]
Polygalacturonase-inhibiting proteins (PGIPs), extracellular proteins that specifically inhibit fungal endopolygalacturonases (PGs), play a critical role in plant protection by favouring the accumulation of oligogalacturonides (OGs), which are elicitors of plant defence responses. The genes encoding PGIP2 of P. vulgaris and the variant PGIP2.Q224K were subjected to error prone PCR (epPCR) to generate mutated inhibitors with novel and improved recognition capabilities. Using a Pichia pastoris expression library and a high-throughput screening method, two mutated PvPGIP2.Q224K-derived inhibitors active against the PG produced by the phytopathogenic fungus F. phyllophilum (FpPG) were isolated. Both variants were better inhibitors than PGIP2.Q224K and were characterized by the replacement of the lysine in position 224, supporting the view that the absence of this positively charged amino acid at position 224 is a primary requirement for gaining the inhibition capability against FpPG.
DOI:10.1016/j.bbapap.2003.08.012URL [��������: 1]
DOI:10.1104/pp.105.067546URLPMID:16244152
Botrytis cinerea is a phytopathogenic fungus that causes gray mold in >1,000 plant species. During infection, it secretes several endopolygalacturonases (PGs) to degrade cell wall pectin, and among them, BcPG1 is constitutively expressed and is an important virulence factor. To counteract the action of PGs, plants express polygalacturonase-inhibiting proteins (PGIPs) that have been shown to inhibit a variety of PGs with different inhibition kinetics, both competitive and noncompetitive. The PG-PGIP interaction promotes the accumulation of oligogalacturonides, fragments of the plant cell wall that are general elicitors of plant defense responses. Here, we characterize the enzymatic activity of BcPG1 and investigate its interaction with PGIP isoform 2 from Phaseolus vulgaris (PvPGIP2) by means of inhibition assays, homology modeling, and molecular docking simulations. Our results indicate a mixed mode of inhibition. This is compatible with a model for the interaction where PvPGIP2 binds the N-terminal portion of BcPG1, partially covering its active site and decreasing the enzyme affinity for the substrate. The structural framework provided by the docking model is confirmed by site-directed mutagenesis of the residues that distinguish PvPGIP2 from the isoform PvPGIP1. The finding that PvPGIP2 inhibits BcPG1 with a mixed-type kinetics further indicates the versatility of PGIPs to evolve different recognition specificities.
DOI:10.1111/j.1438-8677.2008.00175.xURLPMID:19689781 [��������: 1]
Polygalacturonase-inhibiting proteins (PGIPs) are plant cell wall proteins that specifically inhibit the activity of endopolygalacturonases (PGs) produced by fungi during the infection process. The interaction with PGIPs limits the destructive potential of PGs and may trigger plant defence responses through the release of elicitor active oligogalacturonides. In order to pinpoint the residues of PvPGIP2 from Phaseolus vulgaris involved in the interaction with PGs, we used site-directed mutagenesis to mutate the residues D131, D157 and D203, and tested for the inhibitory activity of the mutant proteins expressed in Pichia pastoris against Fusarium phyllophilum and Aspergillus niger PGs. Here, we report that mutation of these residues affects the inhibition capacity of PvPGIP2 against F. phyllophilum PG.
DOI:10.1016/j.funbio.2012.04.010URLPMID:22749160 [��������: 1]
By using surface plasmon resonance (SPR) technology, the kinetics of the interaction of various fungal endopolygalacturonases (EPGs) (13 EPGs) with Phaseolus vulgaris (bean) PGIP2 was carried out to determine whether or not there is any interaction between polygalacturonases-inhibiting protein (PGIP) and EPG. The effect of polygalacturonic acid (PGA) on these interactions was also evaluated. The results show that all EPGs evaluated bind to PGIP2, except for AnPGb and the strength of the interaction depends on the EPG/PGIP2 pairing. Further, the presence of PGA has a moderate to strong effect on the EPG/PGIP2 interaction and the strength of the effect is dependent on the exact EPG/PGIP2 pairing. The differences in affinity in the absence and presence of PGA, suggest a certain level of cooperativity. These results indicate a three-component complex similar to that observed for the heparin-ATIII-thrombin, the FGF-FGFR-heparin, or the hedgehog-interference hedgehog-heparan complexes. This data points to an architecture in which the inhibitor binds at a location distant from the substrate binding site. Furthermore, we applied differential proteolysis mass spectrometry (DPMS) to study the location of the binding site between EPG and PGIP2. DPMS studies indicate that PGIP2 does not bind AnPGII, AnPGa, and AnPGc directly over the active site but instead binds on the face opposite to the active site, creating an allosteric interaction.
DOI:10.1104/pp.111.181057URL [��������: 1]
We report here the low-resolution structure of the complex formed by the endo-polygalacturonase from Fusarium phyllophilum and one of the polygalacturonase-inhibiting protein from Phaseolus vulgaris after chemical cross-linking as determined by small-angle x-ray scattering analysis. The inhibitor engages its concave surface of the leucine-rich repeat domain with the enzyme. Both sides of the enzyme active site cleft interact with the inhibitor, accounting for the competitive mechanism of inhibition observed. The structure is in agreement with previous site-directed mutagenesis data and has been further validated with structure-guided mutations and subsequent assay of the inhibitory activity. The structure of the complex may help the design of inhibitors with improved or new recognition capabilities to be used for crop protection.
DOI:10.1007/s004250050130URL [��������: 1]
DOI:10.1006/pmpp.1996.0008URL [��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}