Transcription characteristics of wheat glutamine synthetase isoforms and the sequence analysis of their promoters
WANG Xiao-Chun,1,2, WANG Lu-Lu1, ZHANG Zhi-Yong1, QIN Bu-Tan1, YU Mei-Qin2, WEI Yi-Hao1, MA Xin-Ming,1,*1Collaborative Innovation Center of Henan Grain Crops, Henan Agricultural University, Zhengzhou 450002, Henan, China 2College of Life Science, Henan Agricultural University, Zhengzhou 450002, Henan, China
Abstract As a key enzyme for nitrogen assimilation in wheat, glutamine synthetase is grouped into two classes: cytosolic GS and chloroplastic GS (TaGS2), and cytosolic GS includes TaGS1, TaGSr, and TaGSe. In order to study the expression characteristics and regulatory mechanisms of GS isozymes in chromosome A, B, and D of heterohexaploid wheat, transcripts of TaGS isoforms were analyzed based on the third-generation sequencing technology transcriptome analysis, and 12 promoters of TaGS isozymes of Yumai 49 were cloned based on Chinese Spring genome, and the sequence of the promoters were analyzed. The results showed that TaGS1 was mainly transcribed on chromosome 6B, TaGSe and TaGSr on chromosome 4D, and TaGS2 on chromosome 2D. Furthermore, the distance from initiation codon ATG to initiation site of transcript for each promoter of TaGS was distinct. Promoter element analysis showed that the promoter of TaGS1 in 6B had more W-box, AC-I, ABRE, as-1, and methyl jasmonic response elements, the promoter of TaGSe in 4D had more stress response elements (MYB, MBS, LTR, etc.) and auxin response element, the promoter of TaGSr in 4D had more WRE3 and transcript factor response elements, the promoter of TaGS2 in 2D had more A-box, WRE3, ARE, and an AT enrichment region. In summary, the number, type and order of cis-elements of different promoters of TaGS isozymes were distinct, which provided the foundation for further study on the regulation mechanism of TaGS isozymes. Keywords:wheat;GS isoforms;transcription;promoter;cis-element
PDF (3530KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王小纯, 王露露, 张志勇, 秦步坛, 于美琴, 韦一昊, 马新明. 小麦谷氨酰胺合成酶同工酶转录特点及其启动子序列分析[J]. 作物学报, 2021, 47(4): 761-769. doi:10.3724/SP.J.1006.2021.01046 WANG Xiao-Chun, WANG Lu-Lu, ZHANG Zhi-Yong, QIN Bu-Tan, YU Mei-Qin, WEI Yi-Hao, MA Xin-Ming. Transcription characteristics of wheat glutamine synthetase isoforms and the sequence analysis of their promoters[J]. Acta Agronomica Sinica, 2021, 47(4): 761-769. doi:10.3724/SP.J.1006.2021.01046
Miflin BJ, Habash DZ. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops J Exp Bot, 2002,53:979-987. URLPMID:11912240 [本文引用: 1]
Ortega JL, Wilson OL, Sengupta-GopalanC. The 5′ untranslated region of the soybean cytosolic glutamine synthetase β1 gene contains prokaryotic translation initiation signals and acts as a translational enhancer in plants Mol Genet Genom, 2012,287:881-893. [本文引用: 1]
Keys AJ. The re-assimilation of ammonia produced by photorespiration and the nitrogen economy of C3 higher plants Photosynth Res, 2006,87:165-175. [本文引用: 1]
KonishiN, SaitoM, ImagawaF, KannoK, YamayaT, KojimaS. Cytosolic glutamine synthetase isozymes play redundant roles in ammonium assimilation under low-ammonium conditions in roots of Arabidopsis thaliana Plant Cell Physiol, 2018,59:601-613. [本文引用: 1]
KicheyT, HeumezE, PocholleD, PageauK, VanackerH, DuboisF, Gouis JL, HirelB. Combined agronomic and physiological aspects of nitrogen management in wheat highlight a central role for glutamine synthetase New Phytol, 2006,169:265-278. URLPMID:16411930 [本文引用: 1]
Li HM, LiangH, LiZ, LiZ, Tang ZX, Fu SL, Geng YY, Yan BJ, Ren ZL. Dynamic QTL analysis of protein content and glutamine synthetase activity in re-combinant inbred wheat lines Genet Mol Res, 2015,14:8706-8715. [本文引用: 2]
KicheyT, HirelB, HeumezE, DuboisF, Le GouisJ. In winter wheat ( Triticum aestivum L.) post-anthesis nitrogen uptake and remobilization to the grain correlates with agronomic traits and nitrogen physiological markers Field Crops Res, 2007,102:22-32. [本文引用: 1]
Bernard SM, Moller AL, DionisioG, KicheyT, Jahn TP, DuboisF, BaudoM, Lopes MS, Tercé-LaforgueT, Foyer CH, Parry MA, Forde BG, Araus JL, HirelB, Schjoerring JK, Habash DZ. Gene expression, cellular localization and function of glutamine synthetase isozymes in wheat ( Triticum aestivum L.) Plant Mol Biol, 2008,67:89-105. URLPMID:18288574 [本文引用: 1]
Wang XY, Yu ZW. Study on main quality traits and related enzyme activities of different wheat varieties Sci Agric Sin, 2005,38:1980-1988 (in Chinese with English abstract). [本文引用: 1]
Jia XT, Wei YH, Gu MX, Shi AB, Wang CH. Effects of overexpression of TaGS1/TaGS2 on tobacco salt resistance and its mechanism Chin J Tob Sci, 2017,121:112-117 (in Chinese with English abstract). [本文引用: 1]
HoshidaH, TanakaY, HibinoT, HayashiY, TanakaA, TakabeT, TakabeT. Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase Plant Mol Biol, 2000,43:103-111. [本文引用: 1]
Zhang TX. Functional Analysis of Wheat Glutamine Synthase in Nitrogen Metabolism MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2012. pp 7-32 (in Chinese with English abstract). [本文引用: 3]
Céline MD, Michèle RC, KarineP, MaudL, OlivierG, JocelineK, Marie HV, MagaliF, TiphanieJ, AkiraS. Glutamine synthetase-glutamate synthase pathway and Glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco Plant Physiol, 2006,140:444-456. [本文引用: 1]
MoisonM, MarmagneA, DinantS, SoulayF, AzzopardiM, LothierJ, CiterneS, MorinH, LegayN, ChardonF, Avice JC, Reisdorf-CrenM, Masclaux-DaubresseC. Three cytosolic glutamine synthetase isoforms located in different order veins work together for N remobilization and seed filling in Arabidopsis J Exp Bot, 2018,69:4379-4393. [本文引用: 1]
TabuchiM, SugiyamaK, IshiyamaK, InoueE, SatoT, TakahashiH, YamayaT. Severe reduction in growth rate and grain filling of rice mutants lacking OsGS1:1, a cytosolic glutamine synthetase1:1 Plant J, 2005,42:641-651. [本文引用: 1]
TabuchiM, AbikoT, YamayaT. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.) . J Exp Bot, 2007,58:2319-2327. [本文引用: 1]
Gómez-MaldonadoJ, AvilaC, TorreF, Ca?asR, Cánovas FM, Campbell MM. Functional interactions between a glutamine synthetase promoter and MYB proteins Plant J, 2004,39:513-526. [本文引用: 2]
Dong JM, Xu YQ, Peng LN, FengX, Yao SK, Zhao QC, Li LF, Hu BZ. Cloning and expression characteristics analysis of some homologous genes TaEXPA7 in root of winter wheat Dongnong No.2 variety J Wheat Crops, 2017,37(11):25-33 (in Chinese with English abstract). [本文引用: 1]
HeH. Cloning of the BlOFPs Gene and Analysis of Its Interaction Proteins in Birch MS Thesis of Zhejiang A & F University, Hangzhou, Zhejiang, China, 2016. pp 15-76 (in Chinese with English abstract). [本文引用: 1]
Li ZY, Mou HZ, Gao DM, Ding GP, MaT, WangS. Cloning of cigarette type I promoter and analysis of its transcriptional starting sites Chin J Biol Eng, 2014,34(1):28-35 (in Chinese with English abstract). [本文引用: 1]
LuJ, Zhao HY, He YK, Song YR. Advances in higher plant promoters and their applications Adv Nat Sci, 2004,14:856-862 (in Chinese with English abstract). [本文引用: 1]
Wang HL, Han JJ, Li WH, LiuW. Resistant starch content of different wheat varieties (or lines) of starch branching enzyme EIIn and EII6 gene polymorphism analysis Xinjiang Agric Sci, 2015,52:981-987 (in Chinese with English abstract). [本文引用: 1]
YiH, LiA, Liu HM, Jing RL. Wheat protein phosphatase 2A gene TaPP2AbB"-α promoter cloning and expression analysis Acta Agron Sin, 2016,42:1282-1290 (in Chinese with English abstract). [本文引用: 1]
Shao YP, Yang MM, Bao GG, Sun YN, YangQ, Li WB, Wang ZK. Cloning and functional analysis of soybean GmWRI1a promoter Chin J Oil Crop Sci, 2019,41:517-523 (in Chinese with English abstract). [本文引用: 1]
Wei WJ, DengX, Yang SS. Functional analysis of TaGAPCp1 gene promoter in wheat under abiotic stress J Fujian Norm Univ(Nat Sci Edn), 2019,35(2):76-84 (in Chinese with English abstract). [本文引用: 1]
Chang JZ, Dong CL, ZhangZ, Qiao LY, YangR, JiangD, Zhang YQ, Yang LL, Wu JJ, Jing RL. Functional analysis of 5' non-translational subdomains of stress-related gene TaSAP1 in wheat Acta Agron Sin, 2019,45:1311-1318 (in Chinese with English abstract). [本文引用: 1]
Jin ZX, LiD, Li MY, Tong LG, PanD, Zhang YL, Wang HW, Han YF, Zhang ZC. Relationship between transcriptional expression of glutamine synthase isotype gene and promoter structure in rice J Northeast Agric Univ, 2017,48(10):1-10 (in Chinese with English abstract).
 ,1,2, 王露露1, 张志勇1, 秦步坛1, 于美琴2, 韦一昊1, 马新明
,1,2, 王露露1, 张志勇1, 秦步坛1, 于美琴2, 韦一昊1, 马新明
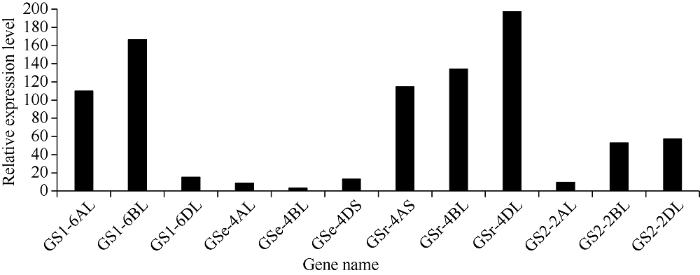 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT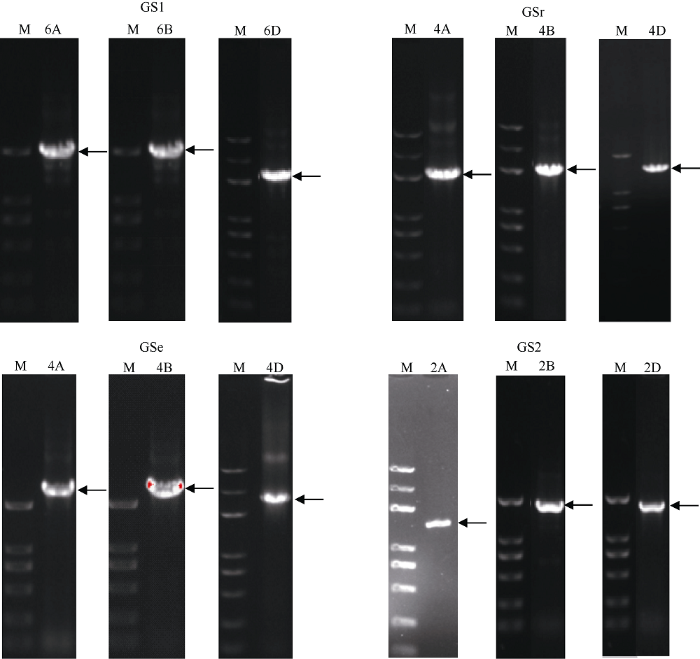 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT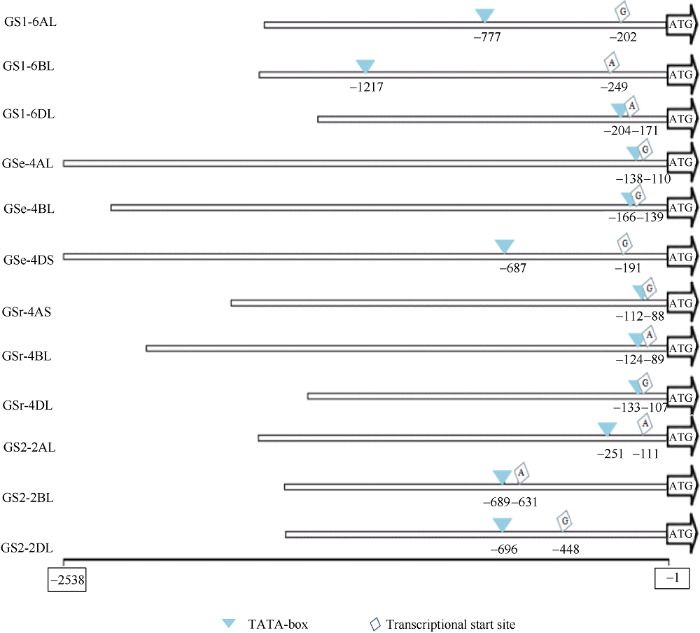 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}