Phenotype analysis and gene mapping of small kernel 7 (smk7) mutant in maize
JIANG Cheng-Gong1,2, SHI Hui-Min2, WANG Hong-Wu2, LI Kun2, HUANG Chang-Ling2, LIU Zhi-Fang2, WU Yu-Jin2, LI Shu-Qiang2, HU Xiao-Jiao2,*, MA Qing1,*通讯作者:
收稿日期:2020-03-11接受日期:2020-08-19网络出版日期:2020-09-10
| 基金资助: |
Corresponding authors:
Received:2020-03-11Accepted:2020-08-19Online:2020-09-10
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (4699KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
蒋成功, 石慧敏, 王红武, 李坤, 黄长玲, 刘志芳, 吴宇锦, 李树强, 胡小娇, 马庆. 玉米籽粒突变体smk7的表型分析和基因定位[J]. 作物学报, 2021, 47(2): 285-293. doi:10.3724/SP.J.1006.2021.03015
JIANG Cheng-Gong, SHI Hui-Min, WANG Hong-Wu, LI Kun, HUANG Chang-Ling, LIU Zhi-Fang, WU Yu-Jin, LI Shu-Qiang, HU Xiao-Jiao, MA Qing.
玉米是世界第一大粮食作物, 也是重要的饲料和工业原料[1], 玉米产量对全球粮食安全具有举足轻重的作用。玉米籽粒发育状况决定着玉米的产量和品质, 因此籽粒遗传发育机制一直是玉米研究的热点[2,3]。玉米的籽粒由胚、胚乳和表皮3个部分组成。胚是籽粒的关键部分, 是精子与卵细胞融合形成的二倍体。胚乳占籽粒的绝大部分, 为籽粒的早期发育提供营养物质, 是精子与中央细胞极核融合形成的三倍体[4]。发掘调控胚和胚乳发育的关键基因并解析其分子机制, 对指导玉米高产优质育种具有重要意义[5]。
籽粒突变体是研究胚和胚乳发育机制的良好材料。早在1980年, Neuffer和Sheridan[6]利用EMS化学诱变玉米材料, 发现大约300个基因上的相关位点与玉米籽粒发育有关。随后, ****们根据突变籽粒的表型将玉米籽粒突变体大致分为, 小籽粒(small kernel, smk)、籽粒严重缺陷(defective kernel, dek), 胚特异缺陷(embryo defective, emb), 粉质胚乳(opaque或floury)和空果皮(empty pericarp, emp)等类型[7]。
胚乳是玉米籽粒营养的主要储存部位, 包括淀粉胚乳、糊粉层、转移层及胚周层4个区域。粉质胚乳突变体具有不透明胚乳, 通常赖氨酸含量较高。赖氨酸具有促进生长发育及增强免疫的作用, 因此这类突变体被广泛研究。目前已克隆opaque或floury突变体有14个, 如隐性突变o2、o5-7、o10、o11[8,9,10], 半显性突变fl1[11]等。其中研究最早的o2突变体赖氨酸含量较野生型增加约69% [12]。O2基因编码Bzip转录因子调控醇溶蛋白基因表达, 突变后醇溶蛋白含量显著降低, 富含赖氨酸的非醇溶蛋白含量增加, 从而极大提高籽粒中赖氨酸含量[13,14,15]。而其余opaque或floury基因也均与醇溶蛋白的结构、分布和积累密切相关。近期克隆的fl3在淀粉胚乳细胞中特异表达, 通过与RNA聚合酶III互作调控5S rRNA和tRNA的转录[16]。胚乳基部转移层(basal endosperm transfer layer, BETL)位于胚和胚乳之间, 是籽粒从母体获取营养的重要部位, 并且还具有细胞分裂素合成、能量代谢、防御反应以及母本和籽粒之间信号传导等多种功能。一旦受损会严重影响籽粒发育, 如mn1、dek37、dek41、dek44等突变体。Mn1编码细胞壁转化酶, mn1突变体胚乳基部转移层缺乏己糖的合成, 导致BETL细胞发育受到影响, 引起籽粒缺陷表型[17]。DEK37编码P亚家族的三角状五肽重复蛋白(PPR), 该蛋白功能的丧失导致线粒体复合体I亚基nad2的第一个内含子剪接效率降低[15]。DEK41编码1个P型的PPR蛋白, 与玉米nad4内含子3的顺式剪切有关, 突变导致转移层细胞中线粒体结构和功能遭到破坏, 引起BETL层细胞发育受损[18]。糊粉层包裹着胚和胚乳细胞, 种子萌发时, 糊粉层产生水解酶, 分解胚乳中的营养物质。目前已知调控糊粉层发育的基因有DEK1、CR4、SAL1、THK1和NKD1。DEK1基因编码胚乳中糊粉层细胞的1个蛋白酶, CR4编码受体激酶, SAL1编码1个E类液泡蛋白, 在质膜的囊泡运输上起作用, CR4和DEK1参与糊粉层细胞特化的信号接收与传导, SAL1降解或者循环DEK1, CR4来维持质膜上二者的浓度, 起负调控作用[19]。
胚是玉米籽粒具有生命活性的部分。胚特异突变体通常表现为籽粒大小正常, 但是没有可见胚或胚发育畸形。已克隆的胚发育特异基因有EMB8156、EMB8522、LEM1、EMB12、EMB14、EMB16、EMB-7L[20]等。EMB8516、LEM1、EMB14和EMB16的功能均与质体核糖体形成相关, Emb12编码质体起始因子3 (IF3), 质体基因的正确表达对玉米胚发生具有重要意义。PPR8522与EMB-7L均编码叶绿体靶向的PPR蛋白, 突变后叶绿体受损, 代谢产物合成受抑制[21]。除了上述突变体外, 还有一类胚和胚乳都有缺陷的突变体, 如small kernel突变体的胚和胚乳发育都比较滞后。SMK6基因编码1个PPR-E+型蛋白, 突变导致线粒体nad1-740、nad4L-110、nad7-739和mttB-138、139位置上C-U的编辑受到影响, 损害了线粒体活性[22]。SMK4基因碱基的缺失突变导致线粒体复合物IV细胞色素C氧化酶1 (cox1)转录本的1489位C被U编辑[23]。总而言之, 在玉米籽粒突变中, 大部分的籽粒突变是由PPR蛋白的突变导致的。而PPR蛋白分为PLS、E、E+、DYW几种类型, 直接或间接参与RNA稳定、翻译、切割、剪切、编辑等[24]。
本研究以玉米自交系B73经EMS诱变产生的籽粒突变体smk7为研究材料, 对突变籽粒的表型和显微结构进行观察, 同时构建不同遗传背景的F2分离群体, 开展性状的遗传解析和基因定位。结合对定位区间的基因注释, 为候选基因的克隆及功能研究奠定了基础。
1 材料与方法
1.1 实验材料
利用EMS诱变玉米B73自交系花粉构建突变体库, 经过筛选获得1个表型可稳定遗传的小粒突变体smk7 (small kernel 7)。由于smk7发芽率低, 且不能生长成正常植株, 我们将杂合植株(+/smk7)自交4代(M4)获得的分离果穗用于突变表型分析。2017年和2018年在北京昌平基地, 分别以昌7-2、Mo17和郑58为母本, 以M4代杂合植株(+/smk7)为父本, 杂交得到F1群体, 再自交获得不同遗传背景的F2分离群体。F2群体用于后续的遗传分析和基因定位。1.2 实验方法
1.2.1 扫描电镜观察淀粉粒结构 取M4代分离果穗上的野生型和突变体籽粒, 在65℃烘箱内连续烘48 h确保籽粒完全脱水干燥, 把胚乳部分敲碎, 固定在贴有导电条的圆形金属样品台上, 置于离子溅射仪中镀金膜, 用HITACHI SU8020扫描电镜对突变体胚乳的淀粉大小和结构变化进行观察。1.2.2 石蜡切片 取授粉后12 d的M4代分离果穗上的突变体和野生型籽粒, 用FAA固定液保存, 固定后的组织材料进行脱水、透明、浸蜡与包埋, 将包埋好的蜡块切片黏附于载玻片上, 然后盖上盖玻片在45℃烘箱里干燥。再用OLYMPLUS BX53正置显微镜(奥林巴斯, 日本)观察并照相。
1.2.3 籽粒蛋白、油分和淀粉含量的测定 淀粉含量的测定: 取野生型籽粒和突变体籽粒在65℃烘箱中烘6 h以上, 用磨样机磨样, 称取2.5 g粉末, 加10 mL CaCl2-乙酸溶液润湿, 再加50 mL CaCl2溶液充分混匀, 置于120℃油浴锅中加热30 min, 放入冷水槽冷却至室温, 将水解液全部加入带漏斗100 mL容量瓶中, 并加1 mL硫酸锌溶液摇匀, 在加1 mL硫酸亚铁溶液充分沉淀蛋白质, 蒸馏水定容至100 mL, 摇匀过滤。用空白液调整旋光仪零点, 再将滤液装满旋光仪室温下测定。结果计算:
粗淀粉(%) = α×105/L × W × (100 - H) × 203。
式中, α为旋光仪上读出的旋转角度; L为炫光管长度(dm); W为样品重(g); 203为淀粉比旋度; H为样品含水量(%)。
籽粒蛋白和油分测定: 用德国BRUKER MPA近红外光谱仪测定油分和蛋白含量, 分别选取大小均一的突变体和野生型籽粒置于样品杯中, 用透射方式扫描获得籽粒近红外样品的吸收光谱图, 每个样品重复装样扫描3次, 实验设置3个生物学重复。根据本实验室已构建的蛋白和油分模型, 利用BRUKER公司的OPUS软件进行数据分析。
1.2.4 基因的初定位和精细定位 以突变体与Mo17杂交构建的F2群体为材料开展基因定位。提取F2果穗上的45粒突变籽粒及双亲的基因组DNA。委托博瑞迪生物技术有限公司进行基于20K GBTS (genotyping by target sequencing) 靶向测序基因型分型[3]。计算每个多态性SNP标记的基因型频率(SNP index), 即亲本型基因型占所有F2样本基因型的频率, SNP index越接近1, 表明标记与目标基因连锁越紧密。然后用MaizeGDB和Gramene等网站比对和下载定位区间内的核苷酸序列, 用Primer 5等软件开发InDel和SNP标记。进一步扩大定位群体, 筛选标记处的交换单株, 完成基因精细定位。
利用试剂盒DP360 (天根生化科技(北京)有限公司)提取正常籽粒和突变籽粒的DNA, 操作步骤如下: 样品充分研磨, 加入裂解液, 涡旋离心, 转移上清过柱, 将滤液和无水乙醇等体积混合, 将混合液过柱, 倒掉滤液, 加漂洗液漂洗2次, 离心, 加ddH2O溶解2 min, 得到的DNA溶液置于-20℃保存。选用25 μL扩增体系, 上下游引物各1.25 μL, PCR mix 12.5 μL, DNA 2 μL, ddH2O 8 μL。扩增程序为: 98℃预变性3 min; 98℃变性10 s、60℃退火20 s、72℃延伸30 s、共35个循环; 72℃终延伸5 min。利用1%琼脂糖凝胶电泳检测PCR产物。
2 结果与分析
2.1 小粒突变体smk7的表型分析
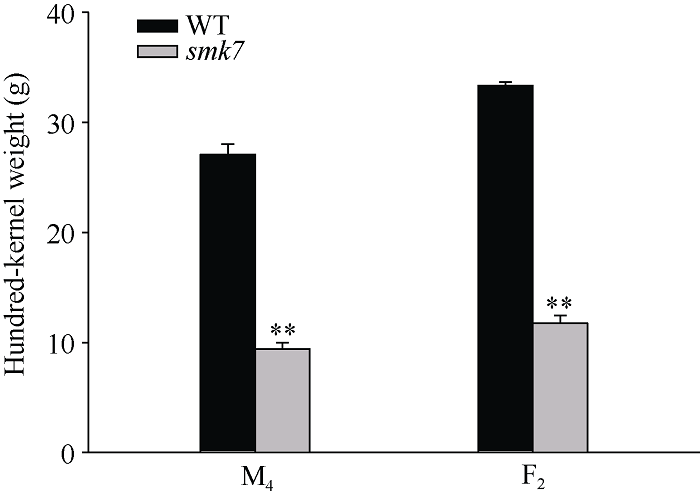
smk7是玉米自交系B73经EMS诱变产生的小籽粒突变体。对M4和F2代成熟分离果穗进行观察发现, smk7突变体与野生型相比, 种皮皱缩, 籽粒扁小, 胚与胚乳发育缺陷(图1-A, B)。仅有10%的突变籽粒可以发育成苗, 但幼苗植株生长发育缓慢, 叶色浅黄, 最终不能发育为正常植株(图1-C)。百粒重测量结果表明, 突变体籽粒百粒重显著降低, 仅约野生型籽粒的35% (图2)。对smk7和野生型籽粒胚乳结构进行扫描电镜观察发现, 野生型淀粉粒呈球形, 形状规则, 而突变体淀粉粒变小, 结构不规则。突变体与野生型蛋白体结构无明显差异(图3)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1smk7突变体表型
A: M4代果穗; B: smk7与野生型籽粒表型比较; C: smk7和野生型幼苗表型比较。
Fig. 1Morphological phenotypes of the smk7
A: maize ear of M4 generation, B: comparison of the phenotype between smk7 and WT kernels; C: comparison of the phenotype between smk7 and WT seedlings.
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2突变体籽粒和野生型籽粒的百粒重
**表示野生型与突变体在0.01水平差异显著。
Fig. 2Hundred-kernel weight of wild type and smk7 kernels
** represents significant differences between the smk7 mutant and wild type at the 0.01 probability level.
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3 smk7及野生型成熟籽粒扫描电镜照片
A: 野生型胚乳; B: smk7胚乳; SG为淀粉粒; PB为蛋白体; 标尺为20 μm。
Fig. 3Scanning electron microscopy (SEM) of mature smk7 and WT kernels
A: wild type endosperm; B: smk7 endosperm; SG: starch granule; PB: protein body; Bar = 20 μm.
Table 1
表1
表1基因定位引物序列
Table 1
| 引物 Primer | 正向序列 Forward sequence (5°-3′) | 反向序列 Reverse sequence (5′-3′) |
|---|---|---|
| In4.3 | AACGCATCATCCTATGTCCAAC | GGGTGAAGCCAGCCATTATTT |
| In1.9 | ATGGTACGATCAACATAAAGGGAA | GGCGTCACCGAAGAAATACAC |
| In0.83 | GCAAGAAGCACCAGCCCT | CGAGCGAAAGAAAGGAATGT |
| In1.16 | ACATGACCCACGATCCAGACA | TGCAGCCACTCTCCTTATGGT |
| SNP1 | TTAGGGTGGAGTTGCTTCGC | TTATGAATGAAGCACGGAAATGA |
| SNP2 | TTAGGGTGGAGTTGCTTCGC | TTATGAATGAAGCACGGAAATGA |
| SNP3 | AAGGAATGGAGGCTTGGGTT | ACAGCCGCCTTCGGATTC |
| SNP4 | AGCGGTCCTTGACTTTATTTGA | CATTACACGACCAATACAGCCA |
| SNP5 | TGGCTTTTCATACCCTCCTCC | CCTTCGCTGTGACTTGGATGT |
新窗口打开|下载CSV
2.2 小粒突变体smk7的籽粒成分测定
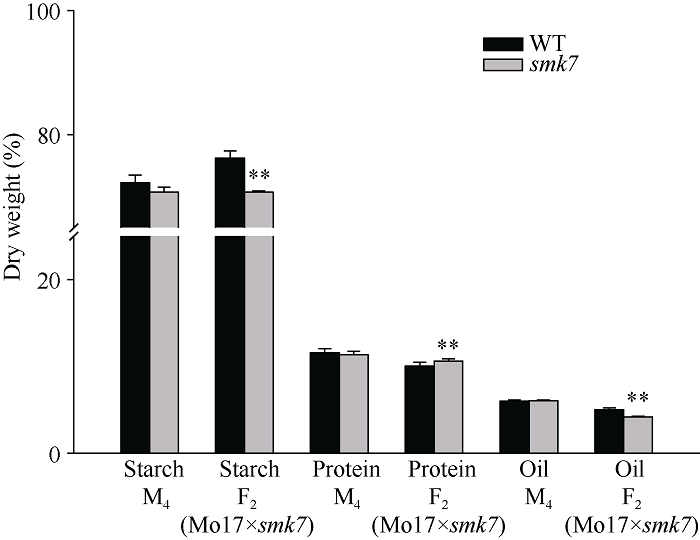
smk7突变体籽粒成分测定结果表明, M4代分离果穗的smk7突变籽粒淀粉含量和蛋白含量低于野生型籽粒, 油分含量突变籽粒略高于野生型籽粒, 均未达到显著水平。F2代分离果穗的smk7突变籽粒淀粉和油分含量显著低于野生型, 而蛋白含量显著高于野生型, 可能是由于不同背景的影响导致F2代籽粒淀粉、油分和蛋白的含量发生变化(图4)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4野生型籽粒和smk7籽粒的成分测定
**表示野生型与突变体在0.01水平差异显著。
Fig. 4Determination and analysis of WT and smk7 kernel components
** represents significant differences between smk7 mutant and wild type at the 0.01 probability level.
2.3 小粒突变体smk7授粉后不同时期胚和胚乳的观察分析
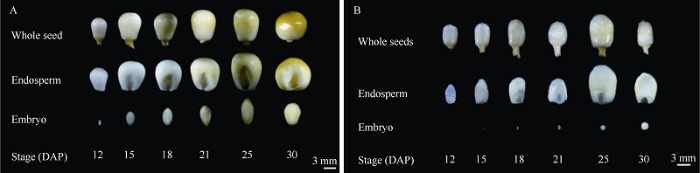
对授粉后12、15、18、21、25和30 d的M4代分离果穗进行观察发现, 授粉后12 d即可明显区分出smk7突变籽粒。与野生型籽粒相比, smk7籽粒明显体积较小, 胚乳不饱满, 胚发育迟滞, 几乎不可见。随着发育进程的增加, 突变表型更加显著, 授粉后21~25 d, 胚发育明显滞后, 大小不足野生型胚的1/10, 胚乳灌浆不饱满。授粉后30 d, 突变籽粒果皮和胚乳间出现较大空隙, 籽粒皱缩体积约为野生型的1/3, 胚和胚乳略有增大, 但结构畸形明显, 以上结果表明smk7突变产生与籽粒发育的早期(图5-A, B)。进一步对授粉后12 d的野生型和smk7籽粒制作石蜡切片, 显微观察发现, 结果发现与野生型籽粒相比, 突变体发育严重滞后, 突变体的胚乳转移层细胞(BETL)存在明显发育异常。授粉后12 d, 野生型籽粒BETL区细胞发育完善, 成带状和基部胚乳细胞接触更为紧密, 有明显的细胞壁内突, 而smk7突变体BETL区细胞与基部分离, 细胞壁内突不明显(图6)。由此推测SMK7基因突变可能导致BETL细胞发育异常, 影响母体营养物质向胚和胚乳传递, 造成了籽粒的发育缺陷表型。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5授粉后不同时期胚和胚乳观察分析
A: 野生型籽粒在12、15、18、21、25和30 DAP胚和胚乳及完整种子的观察分析; 标尺为3 mm。B: 突变型籽粒(smk7)在12、15、18、21、25和30 DAP胚和胚乳及完整种子的观察分析; DAP: 授粉后天数; 标尺为3 mm。
Fig. 5Observation of embryo and endosperm of WT and smk7 at different development stages
A: the observation of embryo, endosperm and whole seed of wild type at 12, 15, 18, 21, 25, and 30 DAP; Bar =3 mm. B: the observation of embryo, endosperm and whole seed of smk7 at 12, 15, 18, 21, 25, and 30 DAP; DAP: days after pollination; Bar = 3 mm.
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6授粉后不同天数野生型和突变体籽粒石蜡切片观察
A: 野生型授粉后12 d籽粒; 标尺为 1000 μm。B: smk7授粉后12 d籽粒; 标尺为1000 μm。C: WT授粉后12 d籽粒胚乳基部转移; 标尺为150 μm; D: smk7授粉后12 d籽粒胚乳基部转移层; 标尺为150 μm; BETL: 胚乳基部转移层细胞。
Fig. 6Observation of paraffin sections of wild type and smk7 kernels on different days after pollination
A: wild type kernels at 12 DAP; Bar = 1000 μm. B: smk7 kernels at 12 DAP; Bar = 1000 μm. C: the basal endosperm transfer layer of WT endosperm at 12 DAP; Bar = 150 μm. D: the basal endosperm transfer layer of smk7 at 12 DAP; Bar = 150 μm; BETL: basal endosperm transfer layers.
2.3 smk7受单隐性核基因控制
利用杂合植株(+/smk7)分别与自交系Mo17、昌7-2、郑58杂交得到F1群体, F1自交获得不同遗传背景的F2群体。挑选杂合体F1植株进行自交, 对M2、M3和F2分离果穗中的小粒突变体和正常籽粒进行统计, 计算分离比, 卡方测验结果表明, 正常籽粒与突变籽粒之比均符合3∶1分离规律(χ2<3.84) (表2)。表明smk7小籽粒性状受1对隐性单基因控制, 是细胞核遗传。Table 2
表2
表2不同群体的遗传分离比检验
Table 2
| 世代 Populations | 总粒数 Total kernels | 正常籽粒数 Normal kernels | 突变籽粒数 Mutant kernels | 实际比例 Actual ratio | 理论比例 Theoretical ratio | 卡方值 χ2 |
|---|---|---|---|---|---|---|
| M2 | 6372 | 4742 | 1630 | 2.91:1 | 3:1 | 1.1151 |
| M3 | 9821 | 7412 | 2409 | 3.08:1 | 3:1 | 1.1367 |
| F2 (Mo17×smk7) | 5123 | 3870 | 1253 | 3.09:1 | 3:1 | 0.7730 |
| F2 (Z58×smk7) | 2016 | 1512 | 504 | 3:1 | 3:1 | 0.1120 |
新窗口打开|下载CSV
2.4 SMK7基因的定位
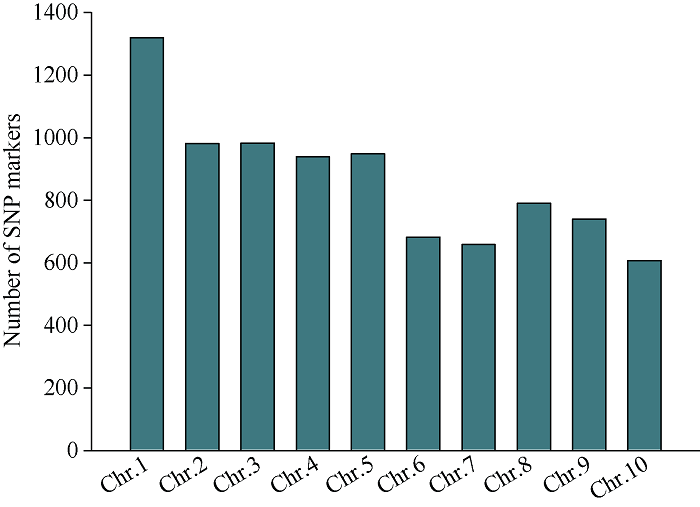
以Mo17为遗传背景的F2分离果穗为材料, 利用CTAB法提取45个粒突变籽粒及其亲本DNA, 利用20K GBTS靶向测序技术分析样本基因型。去除杂合及双亲之间无多态的位点后, 共得到7903个SNP多态性标记, 占总位点数的39.5%, 其中1号染色体最多为1314个, 10号染色体最少为586个(图7)。通过计算各条染色体, 每个多态性SNP标记的基因型频率(SNP index), 发现2号染色体上0.04 Mb~9.54 Mb区间可能与目标性状连锁(图8)。扩大定位群体, 提取274个突变籽粒DNA, 同时开发In0.83、In1.16、In1.9和In4.3若干对InDel标记, 将SMK7定位在0.83 Mb~1.9 Mb区间。继续扩大定位群体到399, 开发SNP1、SNP2、SNP3、SNP4和SNP5等若干对SNP标记, 最终将SMK7基因定位在2号染色体的SNP2和SNP3两个SNP标记之间, 左侧标记处交换单株数为4, 右侧标记处交换单株数为9, 物理距离为120 kb (图9)。利用Gramene (图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7多态性SNP标记的数目
Fig. 7Number of SNP markers
图8
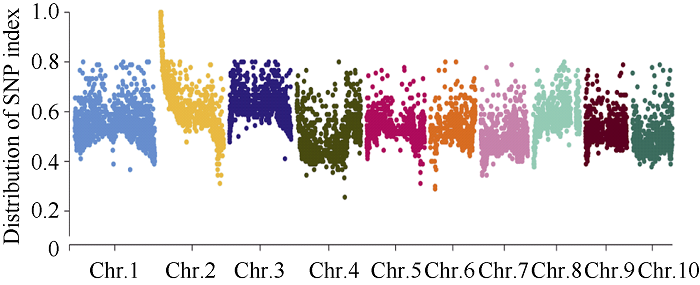 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8SNP index全基因组频率分布图
Fig. 8Distribution profile of SNP index on whole genome
图9
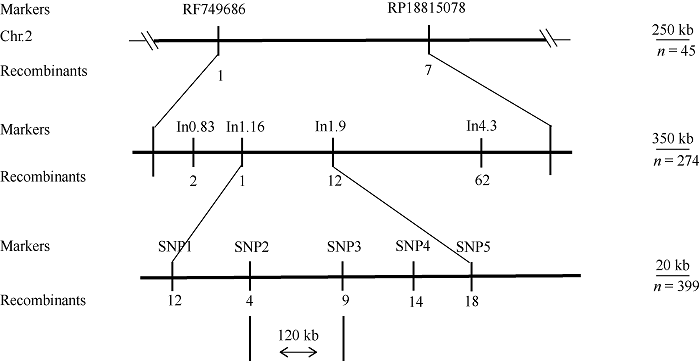 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9SMK7基因的精细定位
群体大小; Recombinants: 重组单株数。
Fig. 9Fine mapping of SMK7
the number of individuals; Recombinants: the number of recombinants.
Table 3
表3
表3候选区间内基因信息
Table 3
| 基因名称 Locus name | 基因注释 Gene annotation |
|---|---|
| ZM00001D001818 | Probable polyol transporter 4 |
| ZM00001D001819 | N-acetylglucosaminyl-phosphatidylinositol de-N-acetylase family protein |
| ZM00001D001820 | Protochlorophyllide reductase1 |
| ZM00001D001821 | Unknown |
| ZM00001D001822 | Unknown |
| ZM00001D001823 | Anthranilate synthase beta subunit 1 chloroplastic |
| ZM00001D001824 | Dof zinc finger protein DOF1.6 |
| ZM00001D001825 | Transducin/WD40 repeat-like superfamily protein |
新窗口打开|下载CSV
3 讨论
籽粒是玉米主要营养储存器官, 也是研究禾本科种子发育的模式器官。开展籽粒突变体研究对于克隆籽粒发育关键基因及解析籽粒发育调控分子机制具有重要意义。本研究中smk7突变体属于小粒(small kernel)类型突变体, 表现为胚和胚乳发育迟滞的表型。成熟的smk7突变体种子体积较小, 胚和胚乳发育缺陷, 尚有少数可以成苗, 幼苗具有黄化现象。这种表型不同于emp、emb等类型的突变体, emp是胚和胚乳严重缺陷的致死突变, 而emb突变体仅胚发育存在异常。小部分dek突变体表型与smk7类似, 如dek36、dek37、dek40、dek42[25]等发育成幼苗。对smk7授粉后12 d 的籽粒石蜡切片观察发现, 突变体BETL层细胞壁向内生长受阻, 发育迟缓。胚乳基部转移层是一类2~3层高度特异的细胞, 含有大量线粒体, 参与细胞的信号转导, 激素合成等重要功能, 种子灌浆发育需要通过BETL从母体中获取营养。表明SMK7基因可能调控胚乳基部转移层细胞形成, 进而影响胚和胚乳发育。
目前已克隆的smk突变体有smk1、smk2、Zmsmk3、smk4、smk6和Zmsmk9[26,27,28,29]。smk1突变体胚和胚乳发育滞后, 种皮和胚乳之间有很大空腔, 约10%的突变籽粒可萌发。SMK1基因编码1个E型PPR蛋白, 参与了线粒体nad7-836的编辑。smk2突变体籽粒变小, 胚败育, 胚乳细胞少, BETL细胞发育受阻。SMK2位于4号染色体, 编码谷氨酰胺酶参与vitamin B6的合成。Zmsmk3突变体表现为胚与胚乳发育滞后, BETL细胞发育严重受损, 约30%突变籽粒可萌发成苗。ZmSMK3位于3号染色体, 编码1个线粒体转录终止因子(mTERF)蛋白, 参与nad1的第4内含子和nad4的第1内含子剪切过程; smk4小粒突变体95%可以成苗, 但是植株生长发育缓慢且花期延后。基因克隆结构表明SMK4位于4号染色体编码E亚类PPR蛋白, 参与线粒体cox1转录本C-U的编辑; smk6是1个小粒胚致死突变体, SMK6基因位于10号染色体, 该基因编码E型PPR蛋白, 影响线粒体基因的编辑功能; Zmsmk9突变体胚与胚乳发育滞后, 但是能正常萌发并发育成株。ZmSMK9位于1号染色体, 编码1个P型PPR蛋白, 参与nad5的剪切。由此可知大部分smk突变均与线粒体功能缺陷有关, 线粒体作为细胞的能量工厂和进行有氧呼吸的主要场所, 在胚乳BETL细胞功能及籽粒发育中具有重要的作用。
本研究中smk7被定位在RM1433917~RM15 35316两个SNP标记之间, 物理距离约为120 kb。通过MaizeGDB和Gramene生物信息学网站分析发现该区段包含了8个新的蛋白编码基因。对其中Zm00001d001818和Zm00001d001820进行PCR扩增并和诱变亲本B73进行序列比对发现无突变位点; Zm00001d001819编码氮-乙酰氨基葡萄糖基磷脂酰肌醇-氮-乙酰化酶家族蛋白, 目前今发现在哺乳动物, 酵母和原生动物中有活性[30]; Zm00001d001823的编码叶绿体β亚基邻氨基苯甲酸合成酶1, 可能参与IAA合成的色氨酸途径[31]。在核分化过程中, IAA控制着糖和蛋白质的代谢, 是胚乳BETL形成的必要条件, IAA通过CK或ABA直接或间接地调控玉米醇溶蛋白编码基因的转录。Zm00001d001824编码Dof锌指蛋白, 广泛参与碳氮代谢、花和花粉发育、种子发育和萌发、次生代谢、维管发育和叶片等植物生长发育过程。Zm00001d001825编码WD40重复超家族转导蛋白, 可能与AP2形成复合物协同作用于网格蛋白介导的内吞作用[32]。玉米edh1突变体中与网格蛋白内吞作用的基因发生突变, 导致突变体籽粒变小, 因此WD40也可能参与籽粒发育调控。所以后续还需通过候选基因测序、基因表达分析等分子实验进一步确定候选基因。
4 结论
本研究以B73为背景材料通过EMS诱变得到1个可以稳定遗传的籽粒突变体, 该突变体籽粒变小, 胚和胚乳发育严重滞后, 胚乳BETL层细胞发育迟缓, 细胞壁发育向内生长受阻。利用图位克隆将SMK7定位在2号染色体短臂RM1433917~ RM1535316这2个SNP标记内, 物理距离约为120 kb, 在该区间内没有已克隆的籽粒发育相关基因, 因此SMK7可能是1个调控籽粒发育的新基因。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3945/an.110.000182URLPMID:22332054 [本文引用: 1]

Africa shares a unique relationship with maize (Zea mays). After its introduction from New World explorers, maize was quickly adopted as the cornerstone of local cuisine, especially in sub-Saharan countries. Although maize provides macro- and micronutrients required for humans, it lacks adequate amounts of the essential amino acids lysine and tryptophan. For those consuming >50% of their daily energy from maize, pandemic protein malnutrition may exist. Severe protein and energy malnutrition increases susceptibility to life-threatening diseases such as tuberculosis and gastroenteritis. A nutritionally superior maize cultivar named quality protein maize (QPM) represents nearly one-half century of research dedicated to malnutrition eradication. Compared with traditional maize types, QPM has twice the amount of lysine and tryptophan, as well as protein bioavailability that rivals milk casein. Animal and human studies suggest that substituting QPM for common maize results in improved health. However, QPM's practical contribution to maize-subsisting populations remains unresolved. Herein, total protein and essential amino acid requirements recommended by the WHO and the Institute of Medicine were applied to estimate QPM target intake levels for young children and adults, and these were compared with mean daily maize intakes by African country. The comparisons revealed that ~100 g QPM is required for children to maintain adequacy of lysine, the most limiting amino acid, and nearly 500 g is required for adults. This represents a 40% reduction in maize intake relative to common maize to meet protein requirements. The importance of maize in Africa underlines the potential for QPM to assist in closing the protein inadequacy gap.
DOI:10.1016/s1369-5266(99)80006-5URLPMID:10047564 [本文引用: 1]
There is a renewal of interest in endosperm development. Recent studies are leading the way to a better understanding of fundamental processes such as cell cycle control and the mechanisms of imprinting. A more global view of interactions between the endosperm and the embryo is emerging and will initiate an integrated approach to the study of seed development.
[本文引用: 2]
DOI:10.1016/j.pbi.2010.08.009URLPMID:20875768 [本文引用: 1]
In angiosperms, a double-fertilization event leads to the formation of a diploid embryo and a triploid endosperm. In Arabidopsis and many dicots, seed development undergoes an initial phase of active endosperm proliferation followed by a second phase in which embryo grows to full size and replaces most of the endosperm volume at its maturity. Since the seed coat and endosperm growth in Arabidopsis precedes embryo growth, the major volume of the mature seed is largely attained before the enlargement of the embryo. Therefore, the seed size is coordinately regulated by the growth of the triploid endosperm, the diploid maternal ovule, and the diploid embryo. Recent studies have identified many new pathway components and revealed possible mechanisms that underlie seed development and size regulation in Arabidopsis. In this review, we shall discuss the regulation of endosperm proliferation by a few newly identified pathways involving transcriptional, epigenetic, and imprinting regulators, the regulation of integument or seed coat development by a few transcription factors, and the regulation of embryo proliferation by AP2-like and bHLH proteins and phytohormones.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:17249053 [本文引用: 1]
A planting of 3,919 M(1) kernels from normal ears crossed by EMS-treated pollen produced 3,461 M(1) plants and 3,172 selfed ears. These plants yielded 2,477 (72%) total heritable changes; the selfed ears yielded 2,457 (78%) recessive mutants, including 855 (27%) recessive kernel mutants and 8 (0.23%) viable dominant mutants. The ratio of recessive to dominant mutants was 201:1. The average mutation frequency for four known loci was three per 3,172 genomes analyzed. The estimated total number of loci mutated was 535 and the estimated number of kernel mutant loci mutated was 285. Among the 855 kernel mutants, 432 had a nonviable embryo, and 59 germinated but had a lethal seedling. A sample of 194 of the latter two types was tested for heritability, lethality, chromosome arm location and endosperm-embryo interaction between mutant and nonmutant tissues in special hyper-hypoploid combinations produced by manipulation of B-A translocations. The selected 194 mutants were characterized and catalogued according to endosperm phenotype and investigated to determine their effects on the morphology and development of the associated embryo. The possibility of rescuing some of the lethal mutants by covering the mutant embryo with a normal endosperm was investigated. Ninety of these 194 mutants were located on 17 of the 18 chromosome arms tested. Nineteen of the located mutants were examined to determine the effect of having a normal embryo in the same kernel with a mutant endosperm, and vice versa, as compared to the expression observed in kernels with both embryo and endosperm in a mutant condition. In the first situation, for three of the 19 mutants, the mutant endosperm was less extreme (the embryo helped); for seven cases, the mutant endosperm was more extreme (the embryo hindered); and for nine cases, there was no change. In the reverse situation, for four cases the normal endosperm helped the mutant embryo; for 14 cases there was no change and one case was inconclusive.
DOI:10.1534/genetics.111.133967URL [本文引用: 1]
In maize, a series of seed mutants with starchy endosperm could increase the lysine content by decreased amount of zeins, the main storage proteins in endosperm. Cloning and characterization of these mutants could reveal regulatory mechanisms for zeins accumulation in maize endosperm. Opaque7 (o7) is a classic maize starchy endosperm mutant with large effects on zeins accumulation and high lysine content. In this study, the O7 gene was cloned by map-based cloning and confirmed by transgenic functional complementation and RNAi. The o7-ref allele has a 12-bp in-frame deletion. The four-amino-acid deletion caused low accumulation of o7 protein in vivo. The O7 gene encodes an acyl-activating enzyme with high similarity to AAE3. The opaque phenotype of the o7 mutant was produced by the reduction of protein body size and number caused by a decrease in the a-zeins concentrations. Analysis of amino acids and metabolites suggested that the O7 gene might affect amino acid biosynthesis by affecting a-ketoglutaric acid and oxaloacetic acid. Transgenic rice seeds containing RNAi constructs targeting the rice ortholog of maize O7 also produced lower amounts of seed proteins and displayed an opaque endosperm phenotype, indicating a conserved biological function of O7 in cereal crops. The cloning of O7 revealed a novel regulatory mechanism for storage protein synthesis and highlighted an effective target for the genetic manipulation of storage protein contents in cereal seeds.
[本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pgen.1006270URLPMID:27541862 [本文引用: 1]
Cereal storage proteins are major nitrogen sources for humans and livestock. Prolamins are the most abundant storage protein in most cereals. They are deposited into protein bodies (PBs) in seed endosperm. The inner structure and the storage mechanism for prolamin PBs is poorly understood. Maize opaque10 (o10) is a classic opaque endosperm mutant with misshapen PBs. Through positional cloning, we found that O10 encodes a novel cereal-specific PB protein. Its middle domain contains a seven-repeat sequence that is responsible for its dimerization. Its C terminus contains a transmembrane motif that is required for its ER localization and PB deposition. A cellular fractionation assay indicated that O10 is initially synthesized in the cytoplasm and then anchored to the ER and eventually deposited in the PB. O10 can interact with 19-kD and 22-kD alpha-zeins and 16-kD and 50-kD gamma-zeins through its N-terminal domain. An immunolocalization assay indicated that O10 co-localizes with 16-kD gamma-zein and 22-kD alpha-zein in PBs, forming a ring-shaped structure at the interface between the alpha-zein-rich core and the gamma-zein-rich peripheral region. The loss of O10 function disrupts this ring-shaped distribution of 22-kD and 16-kD zeins, resulting in misshapen PBs. These results showed that O10, as a newly evolved PB protein, is essential for the ring-shaped distribution of 22-kD and 16-kD zeins and controls PB morphology in maize endosperm.
DOI:10.1105/tpc.107.053538URLPMID:17693529 [本文引用: 1]
The maize (Zea mays) floury1 (fl1) mutant was first reported almost 100 years ago, but its molecular identity has remained unknown. We report the cloning of Fl1, which encodes a novel zein protein body membrane protein with three predicted transmembrane domains and a C-terminal plant-specific domain of unknown function (DUF593). In wild-type endosperm, the FL1 protein accumulates at a high level during the period of zein synthesis and protein body development and declines to a low level at kernel maturity. Immunogold labeling showed that FL1 resides in the endoplasmic reticulum surrounding the protein body. Zein protein bodies in fl1 mutants are of normal size, shape, and abundance. However, mutant protein bodies ectopically accumulate 22-kD alpha-zeins in the gamma-zein-rich periphery and center of the core, rather than their normal discrete location in a ring at outer edge of the core. The 19-kD alpha-zein is uniformly distributed throughout the core in wild-type protein bodies, and this distribution is unaffected in fl1 mutants. Pairwise yeast two-hybrid experiments showed that FL1 DUF593 interacts with the 22-kD alpha-zein. Results of these studies suggest that FL1 participates in protein body formation by facilitating the localization of 22-kD alpha-zein and that this is essential for the formation of vitreous endosperm.
URLPMID:14171571 [本文引用: 1]
DOI:10.1073/pnas.87.1.46URLPMID:2296602 [本文引用: 1]
The opaque-2 locus (o2) in maize regulates the expression of many members of the zein multigene family of storage proteins. cDNA clones for a wild-type allele of the (o2) locus (O2) were isolated from a maize endosperm cDNA library and sequenced. We found a 258-nucleotide 5' leader sequence containing three short open reading frames followed by a sequence specifying a protein of 437 amino acids. The presumptive amino acid sequence of the protein (O2) specified by the O2 cDNA contains a
DOI:10.1105/tpc.4.6.689URLPMID:1392590 [本文引用: 1]
opaque-2 (o2) is a regulatory locus in maize that plays an essential role in controlling the expression of genes encoding the 22-kD zein proteins. Through DNase I footprinting and DNA binding analyses, we have identified the binding site for the O2 protein (O2) in the promoter of 22-kD zein genes. The sequence in the 22-kD zein gene promoter that is recognized by O2 is similar to the target site recognized by other
DOI:10.1007/BF00037028URLPMID:7766871 [本文引用: 2]
The maize opaque 2 (o2) mutation is known to have numerous pleiotropic effects. Some polypeptides have their expression depressed while others are enhanced. The best characterized effects of the o2 mutation are those exerted on endosperm genes encoding the storage protein class of the 22 kDa alpha-zeins and the ribosome inactivating protein b-32. The Opaque 2 (O2) locus encodes a basic domain-leucine zipper DNA-binding factor, O2, which transcriptionally regulates these genes. In the maize-related grass Coix lacryma-jobi, an O2-homologous protein regulates the 25 kDa alpha-coixin family. We show in this paper that O2 transcriptionally regulates the structurally and developmentally different class of the beta-prolamins. A new O2-binding box was identified in beta-prolamin genes from maize and Coix that, together with the boxes previously identified in other endosperm expressed genes, forms a curious collection of O2 cis elements. This may have regulatory implications on the role of O2 in the mechanism that controls coordinated gene expression in the developing endosperm. Considering that the O2 locus controls at least three distinct classes of genes in maize endosperm, we propose that the O2 protein may play a more general role in maize endosperm development than previously conceived.
DOI:10.1105/tpc.17.00576URLPMID:28874509 [本文引用: 1]
Maize (Zea mays) floury3 (fl3) is a classic semidominant negative mutant that exhibits severe defects in the endosperm but fl3 plants otherwise appear normal. We cloned the fl3 gene and determined that it encodes a PLATZ (plant AT-rich sequence and zinc binding) protein. The mutation in fl3 resulted in an Asn-to-His replacement in the conserved PLATZ domain, creating a dominant allele. Fl3 is specifically expressed in starchy endosperm cells and regulated by genomic imprinting, which leads to the suppressed expression of fl3 when transmitted through the male, perhaps as a consequence the semidominant behavior. Yeast two-hybrid screening and bimolecular luciferase complementation experiments revealed that FL3 interacts with the RNA polymerase III subunit 53 (RPC53) and transcription factor class C 1 (TFC1), two critical factors of the RNA polymerase III (RNAPIII) transcription complex. In the fl3 endosperm, the levels of many tRNAs and 5S rRNA that are transcribed by RNAPIII are significantly reduced, suggesting that the incorrectly folded fl3 protein may impair the function of RNAPIII. The transcriptome is dramatically altered in fl3 mutants, in which the downregulated genes are primarily enriched in pathways related to translation, ribosome, misfolded protein responses, and nutrient reservoir activity. Collectively, these changes may lead to defects in endosperm development and storage reserve filling in fl3 seeds.
DOI:10.3389/fpls.2014.00063URLPMID:24616729 [本文引用: 1]
In maize developing seeds, transfer cells are prominently located at the basal endosperm transfer layer (BETL). As the first filial cell layer, BETL is a gateway to sugars, nutrients and water from mother plant; and anchor of numerous functions such as sucrose turnover, auxin and cytokinin biosynthesis/accumulation, energy metabolism, defense response, and signaling between maternal and filial generations. Previous studies showed that basal developing endosperms of miniature1 (mn1) mutant seeds lacking the Mn1-encoded cell wall invertase II, are also deficient for hexose. Given the role of glucose as one of the key sugars in protein glycosylation and proper protein folding; we performed a comparative large scale glycoproteome profiling of total proteins of these two genotypes (mn1 mutant vs. Mn1 wild type) using 2D gel electrophoresis and glycosylation/total protein staining, followed by image analysis. Protein identification was done by LC-MS/MS. A total of 413 spots were detected; from which, 113 spots matched between the two genotypes. Of these, 45 showed >20% decrease/increase in glycosylation level and were selected for protein identification. A large number of identified proteins showed decreased glycosylation levels in mn1 developing endosperms as compared to the Mn1. Functional classification of proteins, showed mainly of post-translational modification, protein turnover, chaperone activities, carbohydrate and amino acid biosynthesis/transport, and cell wall biosynthesis. These proteins and activities were related to endoplasmic reticulum (ER) stress and unfolded protein response (UPR) as a result of the low glycolsylation levels of the mutant proteins. Overall, these results provide for the first time a global glycoproteome profile of maize BETL-enriched basal endosperm to better understand their role in seed development in maize.
DOI:10.1093/jxb/erz193URLPMID:31020318 [本文引用: 1]
The splicing of organelle-encoded mRNA in plants requires proteins encoded in the nucleus. The mechanism of splicing and the factors involved are not well understood. Pentatricopeptide repeat (PPR) proteins are known to participate in such RNA-protein interactions. Maize defective kernel 41 (dek41) is a seedling-lethal mutant that causes developmental defects. In this study, the Dek41 gene was cloned by Mutator tag isolation and allelic confirmation, and was found to encode a P-type PPR protein that targets mitochondria. Analysis of the mitochondrial RNA transcript profile revealed that dek41 mutations cause reduced splicing efficiency of mitochondrial nad4 intron 3. Immature dek41 kernels exhibited severe reductions in complex I assembly and NADH dehydrogenase activity. Up-regulated expression of alternative oxidase genes and deformed inner cristae of mitochondria in dek41, as revealed by TEM, indicated that proper splicing of nad4 is essential for correct mitochondrial functioning and morphology. Consistent with this finding, differentially expressed genes in the dek41 endosperm included those related to mitochondrial function and activity. Our results indicate that DEK41 is a PPR protein that affects cis-splicing of mitochondrial nad4 intron 3 and is required for correct mitochondrial functioning and maize kernel development.
URLPMID:17933905 [本文引用: 1]
DOI:10.1016/j.plantsci.2019.110203URLPMID:31481208 [本文引用: 1]
Embryo and endosperm originate from the double fertilization, but they have different developmental fates and biological functions. We identified a previously undescribed maize seed mutant, wherein the embryo appears to be more severely affected than the endosperm (embryo-specific, emb). In the W22 background, the emb embryo arrests at the transition stage whereas its endosperm appears nearly normal in size. At maturity, the embryo in W22-emb is apparently small or even invisible. In contrast, the emb endosperm develops into a relative normal size. We cloned the mutant gene on the Chromosome 7L and designated it emb-7L. This gene is generally expressed, but it has a relatively higher expression level in leaves. Emb-7L encodes a chloroplast-localized P-type pentatricopeptide repeat (PPR) protein, consistent with the severe chloroplast deficiency in emb-7L albino seedling leaves. Full transcriptome analysis of the leaves of WT and emb-7L seedlings reveals that transcription of chloroplast protein-encoding genes are dramatically variable with pre-mRNA intron splicing apparently affected in a tissue-dependent pattern and the chloroplast structure and activity were dramatically affected including chloroplast membrane and photosynthesis machinery component and synthesis of metabolic products (e.g., fatty acids, amino acids, starch).
DOI:10.1111/tpj.12161URLPMID:23451851 [本文引用: 1]
Embryo-specific mutants in maize define a unique class of genetic loci that affect embryogenesis without a significant deleterious impact on endosperm development. Here we report the characterization of an embryo specific12 (emb12) mutant in maize. Embryogenesis in the emb12 mutants is arrested at or before transition stage. The mutant embryo at an early stage exhibits abnormal cell structure with increased vacuoles and dramatically reduced internal membrane organelles. In contrast, the mutant endosperm appears normal in morphology, cell structure, starch, lipid and protein accumulation. The Emb12 locus was cloned by transposon tagging and predicts a protein with a high similarity to prokaryotic translation initiation factor 3 (IF3). EMB12-GFP fusion analysis indicates that EMB12 is localized in plastids. The RNA in situ hybridization and protein immunohistochemical analyses indicate that a high level of Emb12 expression localizes in the embryo proper at early developmental stages and in the embryo axis at later stages. Western analysis indicates that plastid protein synthesis is impaired. These results indicate that Emb12 encodes the plastid IF3 which is essential for embryogenesis but not for endosperm development in maize.
URLPMID:31234031 [本文引用: 1]
URLPMID:33215867 [本文引用: 1]
DOI:10.1105/tpc.104.022236URLPMID:15269332 [本文引用: 1]
The complete sequence of the Arabidopsis thaliana genome revealed thousands of previously unsuspected genes, many of which cannot be ascribed even putative functions. One of the largest and most enigmatic gene families discovered in this way is characterized by tandem arrays of pentatricopeptide repeats (PPRs). We describe a detailed bioinformatic analysis of 441 members of the Arabidopsis PPR family plus genomic and genetic data on the expression (microarray data), localization (green fluorescent protein and red fluorescent protein fusions), and general function (insertion mutants and RNA binding assays) of many family members. The basic picture that arises from these studies is that PPR proteins play constitutive, often essential roles in mitochondria and chloroplasts, probably via binding to organellar transcripts. These results confirm, but massively extend, the very sparse observations previously obtained from detailed characterization of individual mutants in other organisms.
DOI:10.1104/pp.19.01336URLPMID:31857426 [本文引用: 1]
The roles of C-terminal Eps15 homology domain (EHD) proteins in clathrin-mediated endocytosis in plants are poorly understood. Here, we isolated a maize (Zea mays) mutant, designated ehd1, which showed defects in kernel development and vegetative growth. Positional cloning and transgenic analysis revealed that ehd1 encodes an EHD protein. Internalization of the endocytic tracer FM4-64 was substantially reduced in the ehd1 mutant and ZmEHD1 knockout mutants. We further demonstrated that ZmEHD1 and the ZmAP2 sigma subunit physically interact at the plasma membrane. Auxin distribution and ZmPIN1a-YFP localization were altered in the ehd1 mutant. Kernel indole-3-acetic acid levels were substantially lower in the ehd1 mutant than in wild-type maize. Exogenous application of 1-naphthaleneacetic acid, but not GA3 or 2-naphthaleneacetic acid, rescued the seed germination and seedling emergency phenotypic defects of ehd1 mutants. Taken together, these results indicate that ZmEHD1 regulates auxin homeostasis by mediating clathrin-mediated endocytosis through its interaction with the ZmAP2 sigma subunit, which is crucial for kernel development and vegetative growth of maize.
DOI:10.1111/tpj.12584URLPMID:24923534 [本文引用: 1]
RNA editing modifies cytidines (C) to uridines (U) at specific sites in the transcripts of mitochondria and plastids, altering the amino acid specified by the DNA sequence. Here we report the identification of a critical editing factor of mitochondrial nad7 transcript via molecular characterization of a small kernel 1 (smk1) mutant in Zea mays (maize). Mutations in Smk1 arrest both the embryo and endosperm development. Cloning of Smk1 indicates that it encodes an E-subclass pentatricopeptide repeat (PPR) protein that is targeted to mitochondria. Loss of SMK1 function abolishes the C --> U editing at the nad7-836 site, leading to the retention of a proline codon that is edited to encode leucine in the wild type. The smk1 mutant showed dramatically reduced complex-I assembly and NADH dehydrogenase activity, and abnormal biogenesis of the mitochondria. Analysis of the ortholog in Oryza sativa (rice) reveals that rice SMK1 has a conserved function in C --> U editing of the mitochondrial nad7-836 site. T-DNA knock-out mutants showed abnormal embryo and endosperm development, resulting in embryo or seedling lethality. The leucine at NAD7-279 is highly conserved from bacteria to flowering plants, and analysis of genome sequences from many plants revealed a molecular coevolution between the requirement for C --> U editing at this site and the existence of an SMK1 homolog. These results demonstrate that Smk1 encodes a PPR-E protein that is required for nad7-836 editing, and this editing is critical to NAD7 function in complex-I assembly in mitochondria, and hence to embryo and endosperm development in maize and rice.
URLPMID:28408540 [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.plantsci.2019.110205URLPMID:31521217 [本文引用: 1]
Maize kernel size and weight are essential contributors to its yield. So the identification of the genes controlling kernel size and weight can give us a chance to gain the yield. Here, we identified a small kernel mutant, Zea mays small kernel 9 (Zmsmk9), in maize. Cytological observation showed that the development of the endosperm and embryo was delayed in Zmsmk9 mutants at the early stages, resulting in a small kernel phenotype. Interestingly, despite substantial variation in kernel size, the germination of Zmsmk9 seeds was comparable to that of WT, and could develop into normal plants with upright leaf architecture. We cloned Zmsmk9 via map-based cloning. ZmSMK9 encodes a P-type pentatricopeptide repeat protein that targets to mitochondria, and is involved in RNA splicing in mitochondrial NADH dehydrogenase5 (nad5) intron-1 and intron-4. Consistent with the delayed development phenotype, transcriptome analysis of 12-DAP endosperm showed that starch and zeins biosynthesis related genes were dramatically down regulated in Zmsmk9, while cell cycle and cell growth related genes were dramatically increased. As a result, ZmSMK9 is a novel gene required for the splicing of nad5 intron-1 and intron-4, kernel development, and plant architecture in maize.
DOI:10.1110/gad.03104003URLPMID:12876312 [本文引用: 1]
The TT1542 protein from Thermus thermophilus HB8 is annotated as a conserved hypothetical protein, and belongs to the DUF158 family in the Pfam database. A BLAST search revealed that homologs of TT1542 are present in a wide range of organisms. The TT1542 homologs in eukaryotes, PIG-L in mammals, and GPI12 in yeast and protozoa, have N-acetylglucosaminylphosphatidylinositol (GlcNAc-PI) de-N-acetylase activity. Although most of the homologs in prokaryotes are hypothetical and have no known function, Rv1082 and Rv1170 from Mycobacterium tuberculosis are enzymes involved in the mycothiol detoxification pathway. Here we report the crystal structure of the TT1542 protein at 2.0 A resolution, which represents the first structure for this superfamily of proteins. The structure of the TT1542 monomer consists of a twisted beta-sheet composed of six parallel beta-strands and one antiparallel beta-strand (with the strand order 3-2-1-4-5-7-6) sandwiched between six alpha-helices. The N-terminal five beta-strands and four alpha-helices form an incomplete Rossmann fold-like structure. The structure shares some similarity to the sugar-processing enzymes with Rossmann fold-like domains, especially those of the GPGTF (glycogen phosphorylase/glycosyl transferase) superfamily, and also to the NAD(P)-binding Rossmann fold domains. TT1542 is a homohexamer in the crystal and in solution, the six monomers forming a cylindrical structure. Putative active sites are suggested by the structure and conserved amino acid residues.
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1509292112URLPMID:26417106 [本文引用: 1]
Cellulose biosynthesis is performed exclusively by plasma membrane-localized cellulose synthases (CESAs). Therefore, the trafficking of CESAs to and from the plasma membrane is an important mechanism for regulating cellulose biosynthesis. CESAs were recently identified as cargo proteins of the classic adaptor protein 2 (AP2) complex of the clathrin-mediated endocytosis (CME) pathway. The AP2 complex of the CME pathway is conserved in yeast, animals, and plants, and has been well-characterized in many systems. In contrast, the recently discovered TPLATE complex (TPC), which is proposed to function as a CME adaptor complex, is only conserved in plants and a few other eukaryotes. In this study, we discovered that the TWD40-2 protein, a putative member of the TPC, is also important for the endocytosis of CESAs. Genetic analysis between TWD40-2 and AP2M of the AP2 complex revealed that the roles of TWD40-2 in CME are both distinct from and cooperative with the AP2 complex. Loss of efficient CME in twd40-2-3 resulted in the unregulated overaccumulation of CESAs at the plasma membrane. In seedlings of twd40-2-3 and other CME-deficient mutants, a direct correlation was revealed between endocytic deficiency and cellulose content deficiency, highlighting the importance of controlled CESA endocytosis in regulating cellulose biosynthesis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}