 ,1,2, 叶亚峰1, 何丹1, 任艳1, 杨阳1, 谢建1, 程维民1, 陶亮之1, 周利斌3, 吴跃进1, 刘斌美,1,*
,1,2, 叶亚峰1, 何丹1, 任艳1, 杨阳1, 谢建1, 程维民1, 陶亮之1, 周利斌3, 吴跃进1, 刘斌美,1,*Identification and gene localization of a novel rice brittle culm mutant bc17
JIANG Hong-Rui,1,2, YE Ya-Feng1, HE Dan1, REN Yan1, YANG Yang1, XIE Jian1, CHENG Wei-Min1, TAO Liang-Zhi1, ZHOU Li-Bin3, WU Yue-Jin1, LIU Bin-Mei,1,*通讯作者:
收稿日期:2020-04-3接受日期:2020-09-13网络出版日期:2021-01-12
| 基金资助: |
Received:2020-04-3Accepted:2020-09-13Online:2021-01-12
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (3527KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
姜鸿瑞, 叶亚峰, 何丹, 任艳, 杨阳, 谢建, 程维民, 陶亮之, 周利斌, 吴跃进, 刘斌美. 一个新的水稻脆秆突变体bc17的鉴定及基因定位[J]. 作物学报, 2021, 47(1): 71-79. doi:10.3724/SP.J.1006.2021.02025
JIANG Hong-Rui, YE Ya-Feng, HE Dan, REN Yan, YANG Yang, XIE Jian, CHENG Wei-Min, TAO Liang-Zhi, ZHOU Li-Bin, WU Yue-Jin, LIU Bin-Mei.
水稻茎秆的机械强度是重要的农艺性状之一, 直接影响着植株的抗倒伏能力。水稻脆性突变体是一类比较常见的突变体[1], 是研究细胞壁合成机制的重要材料。脆性突变一般是指植株茎秆机械强度下降, 通常表现为纤维素和半纤维素含量下降, 木质素含量升高[2], 在结构上表现为厚壁组织次生细胞壁变薄[3]。由于茎秆机械强度下降, 存在倒伏发生的隐患, 生产上一般认为是不利的性状。然而脆性突变体秸秆脆嫩、纤维素含量下降, 有利于反刍动物采食, 消化更加容易, 具备成为粮饲兼用型水稻品种的应用潜力[4]。另外, 脆秆突变体的茎秆易粉碎降解, 对于秸秆生态还田有着很好的应用前景, 是一种重要的种质资源[5]。
从第一个水稻脆秆基因bc1[6]被发现以来, 对水稻脆性突变体的研究已有50多年的历史了。截止目前, 报道的水稻脆性突变体仅有20多个, 大多数基因都是涉及细胞壁纤维素合成酶OsCesA (cellulose synthase)的变异, 其中以bc (brittle culm)命名的水稻脆性突变体有16个(bc1~bc16), 相对研究的比较系统和深入, 并且对细胞壁合成调控的机制做了详细解析[7,8]。细胞壁化学成分的变化、合成和转运的阻滞以及厚壁组织结构发育异常等都可能引起水稻茎秆机械强度的改变[9,10,11,12,13,14,15,16,17,18,19]。近来发现水稻转录因子也参与细胞壁生物合成的过程[20,21,22]。水稻纤维素合酶基因是引起脆秆特性的关键基因, 其中最重要的是OsCesA4、OsCesA7、OsCesA9, 可能形成一个纤维素合成复合体参与次生细胞壁的合成[12]。水稻脆秆基因bc7和bc11基因[13,14]是OsCesA4的等位变异; bc6和bc13基因[15,16]是OsCesA9的等位变异。bc10、bc14、bc15可能参与细胞壁的修饰, 其中bc10基因编码一个定位在高尔基体的Ⅱ型内整合膜蛋白, 调节细胞壁纤维素合成和阿拉伯半乳聚糖蛋白含量[17], bc14基因编码核苷酸糖转运蛋白OsNST1, 定位于高尔基体, 转运UDPG, 可能为多糖生物合成提供底物[18]。bc15基因编码一个膜相关的类几丁质酶蛋白[19]。转录因子MYB家族和NAC家族参与到细胞壁的合成途径, 比如NAC29/31-MYB61-CESA通路调控细胞壁生物合成和组装来影响次生细胞壁纤维素合成[20,21,22]。bc12基因编码一个双靶向的驱动蛋白4, 控制水稻细胞周期进程, 揭示细胞生长和细胞壁修饰存在潜在的联系[23]。细胞壁的合成主要由不同类型的糖基转移酶来完成, 而且目前被揭示功能的酶还很少, 明确具体生化特征的更少[8]。由于水稻细胞壁的生物合成过程非常复杂, 涉及的调控途径类型多样, 新基因的发掘和研究有助于完善细胞壁合成机制, 为水稻细胞壁定向改良育种奠定理论基础。
本实验以重离子辐照诱变粳稻品种武运粳7号(Wuyunjing 7, wyj7)获得的一个脆秆突变体为研究对象, 根据水稻脆秆突变基因命名规则和顺序, 将其称为bc17突变体(brittle culm 17), 分析bc17突变体的脆性性状对农艺特征及机械强度的影响, 通过茎秆组织结构的解剖学及细胞壁成分的测定来分析脆性形成的生物学机制, 利用遗传学及图位克隆技术将bc17突变位点进行染色体定位, 通过生物信息学分析鉴定可能的候选基因, 为进一步分离目标基因奠定基础, 同时为育种应用提供高效的筛选标记。
1 材料与方法
1.1 材料
脆秆突变体bc17、野生型武运粳7号以及用于遗传分析和基因定位的群体材料, 均种植于中国科学院合肥物质科学研究院水稻实验基地(安徽合肥), 种植方式为人工栽插, 常规田间管理。1.2 农艺性状分析
在田间随机取10株成熟期供试水稻材料, 考察其主要农艺特征和经济性状, 包括株高、穗长、分蘖数、每穗粒数、千粒重、结实率等。1.3 细胞壁组分含量分析
根据Van Soest等[24]方法测定细胞壁纤维素、半纤维素和木质素等主要组成组分含量。取成熟期的水稻材料, 将茎秆和叶片分开, 剪成适当的长度放入65℃烘箱中, 待完全烘干后取出, 用旋风磨分别将茎秆和叶片粉碎, 粉末装入干净的封口袋中以备后续实验使用。重复3次。1.4 bc17茎秆机械强度测定
1.4.1 茎秆拉伸力的测定 在成熟期从田间选取新鲜水稻材料, 将茎鞘分离后截取第2节间茎秆夹持在万能力学试验机(LD23.502, 力试(上海)科学仪器有限公司)上, 在茎秆断裂失去载荷后自动停止拉伸, 记录数据。1.4.2 茎秆抗折力的测定 在成熟期从田间选取新鲜水稻材料, 将茎鞘分离后截取第2节间茎秆, 放置在等间距的支架上, 用数显式拉压力计(HP-50, 乐清市艾德堡仪器有限公司)从茎秆中部向下施压使水稻茎秆折断, 记录施加力的峰值, 即为茎秆抗折力。
1.4.3 倒伏指数的计算 根据章忠贵等[25]的方法略有修改, 在成熟期采集新鲜样品, 测量相应位置的长度和重量, 计算突变体和野生型第2节间的弯曲力矩和倒伏指数。
弯曲力矩 = 节间基部至穗顶的长度(cm) × 该节间基部至穗顶的鲜重(g) × 0.001 × 9.8
倒伏指数 = (弯曲力矩/抗折力) × 100
倒伏指数越大, 则茎秆越易倒伏, 以倒伏指数200为抗倒伏临界值。
1.5 茎秆结构的电镜观察
取成熟期相同部位的脆秆突变体及其野生型茎秆切成2~4 mm的小段, 投入含有2.5%戊二醛的PBS缓冲液(4 mmol L-1 sodium phosphate, pH 7.2; 200 mmol L-1 NaCl)中, 4℃固定48 h后用1%锇酸PBS (phosphate buffer saline)溶液固定。固定后的样品经过PBS缓冲液充分漂洗后, 顺序用10%、20%、40%、60%和80%的乙醇梯度各处理30 min脱水, 再用100%乙醇处理2次各30 min, 100%丙酮处理2次各5 min。脱水后的材料依次用London Resin White (Sigma, USA)树脂与乙醇顺序以1∶3、1∶1和3∶1的比例混合进行置换和浸透, 最后换入纯树脂继续浸透48 h, 并于模块中包埋, 60℃聚合24 h。样品修块后以80 nm超薄切片, 放置于铜网上, 观察前用醋酸铀和柠檬酸铅复染。样品放置在透射电镜上观察[26]。1.6 脆秆突变体bc17的遗传学分析
利用bc17突变体分别与粳稻品种(武运粳7号和当粳8号)和籼稻品种(特青、黄花占和9311)进行人工杂交, F1代经自交获得F2代种子。在成熟期对F2代植株进行茎秆脆性鉴定, 统计野生型和突变表型个体的数目, 并用统计学方法计算分离比例和卡方检验。1.7 bc17基因的染色体定位
利用对bc17/9311杂交构建的F2分离群体对bc17基因进行初定位, 采用BSA (bulked segregant analysis)法进行基因定位, 即混合分组分析, 也称分离群体分组分析, 是一种通过在群体中挑选极端或代表性性状的个体组成混池进行分析的方法。通过研究混池之间等位基因/分子标记频率的差异, 将与性状相关的位点在基因组上进行定位。选择在水稻12条染色体上均匀分布的408个SSR (simple sequence repeat)标记进行亲本多态性检测, 利用筛选出的差异SSR标记对亲本、F1植株以及混合池(20个脆秆单株等量DNA混合)进行PCR扩增, 对电泳产物的统计分析。在精细定位阶段, 根据网上公布的Nipponbare和9311序列, 并选择定位区间中存在差异的序列进行引物设计与合成。用这些引物分别对bc17和9311基因组DNA进行PCR扩增。PCR程序按常规方法进行。如果PCR产物在bc17突变体和9311间产生多态性, 就直接用作新的SSR或者InDel (insertion and deletion)分子标记。1.8 数据分析
采用Origin 2018对本文实验数据进行差异性分析和作图。2 结果与分析
2.1 bc17突变体的表型特征
通过对突变体bc17和野生型wyj7在整个生育期中农艺特征的观察发现: 该突变体从抽穗期开始, 株高显著低于野生型, 分蘖数明显减少(图1-A), 在拔节期时通过手工折断实验, 突变体bc17茎秆表现出韧性丧失, 能够明显折成两截折断且断口清晰, 而野生型茎秆则表现出韧性, 没有断口出现难以折断(图1-B)。突变体的叶片和野生型的类似, 均只能折出痕迹, 无明显断裂(图1-C), 突变体的穗子比野生型变短, 穗粒数减少, 结实率显著降低(图1-D)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1突变体bc17和野生型wyj7植株及茎秆折断表现型
A: 成熟期植株; B: 成熟期茎秆折断表型; C: 成熟期叶片; D: 穗部表型。
Fig. 1Plants and broken culm phenotype of mutant type bc17 and wild type wyj7 in mature stage
A: Plants of mature stage; B: An easily broken bc17 culm indicated by arrows; C: Leaves of mature stage; D: Panicle phenotype. wyj7: Wuyunjing 7; bc17: brittle culm 17.
2.2 bc17突变体的农艺性状分析
与野生型相比, 突变体bc17的株高、穗长以及结实率极显著降低, 降低程度分别为26.61%、27.51%以及18.54%, 每穗粒数降低了13.18%, 达到显著水平, 而分蘖数和千粒重虽然降低, 但是差异不显著(表1)。农艺性状的变化表明, bc17脆秆性状的突变对于植株的生长和发育也产生了较大的影响, 可能存在一因多效。Table 1
表1
表1突变体bc17与野生型wyj7农艺性状分析
Table 1
| 性状 Trait | 突变体 Mutant (bc17) | 野生型 Wild type (wyj7) | |
|---|---|---|---|
| 株高 Plant height (cm) | 69.43±1.95** | 94.6±0.80 | |
| 分蘖数Tiller number per plant | 9.67±0.58* | 11.33±0.58 | |
| 结实率 Seed fertility (%) | 63.45±2.39** | 87.53±4.38 | |
| 千粒重 1000-grain weight (g) | 23.6±0.67** | 26.62±0.84 | |
| 穗长 Panicle length (cm) | 12.73±0.55** | 15.56±0.21 | |
| 每穗粒数Number of grain per panicle | 111.94±12.06 | 128.94±9.34 | |
新窗口打开|下载CSV
2.3 细胞壁组分含量的分析
木质素(lignin)、纤维素(cellulose)、半纤维素(hemi-cellulose)以及灰分(主要成分是二氧化硅)是构成水稻细胞壁的主要成分。通过测定成熟期突变体bc17和野生型wyj7茎秆、叶片的细胞壁组分含量, 结果表明, 突变体bc17叶片和茎秆的纤维素含量比野生型分别降低18.67%和22.70%, 半纤维素含量分别升高31.36%和45.76%; 野生型叶片中木质素含量显著低于突变体, 而灰分含量显著高于野生型, 但是在茎秆中差异不明显(表2)。细胞壁成分在不同组织中比例的差异, 可能是引起脆性特征特异表达的主要因素。Table 2
表2
表2野生型和突变体bc17茎秆和叶片细胞壁成分分析
Table 2
| 成分 Component | 茎秆Culm | 叶片Leaf | ||
|---|---|---|---|---|
| 突变体 Mutant (bc17) | 野生型 Wild type (wyj7) | 突变体 Mutant (bc17) | 野生型 Wild type (wyj7) | |
| 纤维素 Cellulose | 19.91±1.77* | 25.77±1.36 | 29.19±2.16* | 35.89±1.74 |
| 半纤维素 Hemicellulose | 22.68±1.78* | 15.56±0.63 | 30.03±1.17 | 22.86±2.40 |
| 木质素 Lignin | 4.39±0.40 | 3.99±0.66 | 6.52±0.30* | 4.61±0.24 |
| 灰分 Ash | 4.21±0.13 | 4.19±0.59 | 6.65±0.12* | 8.07±0.29 |
新窗口打开|下载CSV
2.4 脆秆突变体bc17茎秆的物理特征
机械强度是水稻茎秆重要的物理特征之一, 而脆秆突变是引起机械强度改变的。在成熟期时测定水稻茎秆的第2节间和第3节间的折断力、拉伸力。野生型wyj7与突变体bc17茎秆的第2节间茎秆抗折力分别为6.38和3.33, 第3节间茎秆抗折力分别为7.30和4.73。bc17茎秆第2节间抗折力比野生型降低了47.80%, 第3节间茎秆抗折力降低了35.20% (图2), 这与田间观察的茎秆脆性特征相符, 突变体所需要的茎秆折断力明显减少, 拉伸力的测定也验证了这一点, 第2节间突变体相对于野生型分别降低了63.65%。第3节间突变体相对于野生型分别降低了60.34%。脆秆突变体茎秆抗折力比野生型显著下降, 可能会引起倒伏的发生, 因此计算了第2节间的倒伏指数。结果表明bc17突变体第2节间弯曲力矩比野生型显著下降, 引起倒伏指数比野生型升高, 但是二者差异不显著, 说明bc17突变体抗倒伏能力较强, 可能是由于突变体株高下降, 植株重心下移产生的后果。图2
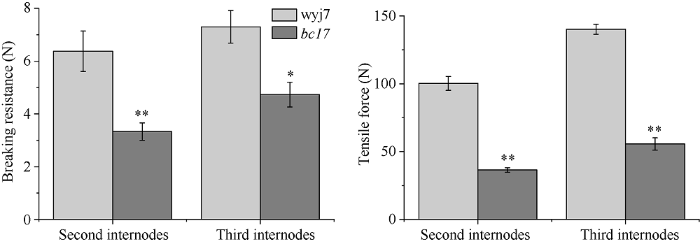 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2野生型和突变体第2节间、第3节间茎秆折断力和茎秆拉伸力分析
* 表示显著差异(P < 0.05), ** 表示极显著差异(P < 0.01)。
Fig. 2Breaking force and tension force of wild type and mutant in the 2nd and 3rd internodes
* and ** represent significant differences between the bc17 mutant and wild type at the 0.05 and 0.01 probability levels, respectively. wyj7: Wuyunjing 7; bc17: brittle culm 17.
Table 3
表3
表3野生型和突变体bc17倒伏指数计算
Table 3
| 参数 Parameters | 野生型 Wild type (wyj7) | 突变体 Mutant (bc17) |
|---|---|---|
| 抗折力Breaking resistance (N) | 6.38±0.59 | 3.33±0.11** |
| 弯曲力矩Bending moment (N m) | 6.31±0.34 | 3.75±0.34** |
| 倒伏指数Lodging index | 99.38±76.40 | 112.15±57.40 |
新窗口打开|下载CSV
2.5 组织解剖学观察
为了观察茎秆性状对细胞壁结构的影响, 成熟期分别对野生型wyj7和突变体bc17茎秆第2节间横切样品进行电镜观察, 结果发现, 突变体茎秆的厚壁组织明显变薄, 细胞之间空隙变大, 相应地细胞密度降低, 细胞数目减少, 从而造成支撑力下降, 对外力响应变得敏感(图3)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3野生型(左)和突变体(右)茎秆横截面
Fig. 3Cross section of wild type (left) and mutant (right) in stalks
2.6 遗传分析
将突变体bc17分别与野生型wyj7正反交得到F1, 杂交后F1代表型正常。统计杂交组合F2代中的群体单株总数以及具有脆秆表型的单株数, 计算分离比例并进行卡方检验, 结果表明, 突变体bc17与不同品种杂交的F2群体脆秆植株分离比例均符合3∶1 (P>0.05)的理论比值(表4), 这表明bc17的脆秆特征是由一对单隐性核基因控制的。Table 4
表4
表4突变体bc17的遗传分析
Table 4
| 杂交组合 Cross | 正常植株 Normal plants | 脆秆株数 Number of brittle culm plants | 群体总数 Total number of F2 | χ2 (χ20.05 = 3.84) |
|---|---|---|---|---|
| bc17/wyj7 | 223 | 78 | 301 | 0.13 |
| bc17/9311 | 329 | 97 | 426 | 1.13 |
新窗口打开|下载CSV
2.7 基因定位
用bc17/9311杂交组合F2分离群体中的97株脆秆表型植株作为定位群体。选取均匀分布于12条染色体上的408对SSR分子标记对bc17和9311两亲本进行多态性筛选, 将具有多态性的110对引物检测亲本。F1基因组DNA以及突变混池DNA (选取20个脆性单株等量DNA混合)进行连锁分析, 其中7号染色体上的SSR引物RM500和RM3743与突变体具有良好的连锁性。利用bc17/9311杂交组合的682株F2脆秆表型植株扩大群体进行精细定位, 在RM500~RM3743之间设计多个InDel分子标记, 最终将bc17定位在UP-81至DN-25之间, 物理距离约162 kb的区间内(表5和图4)。在水稻基因组注释网站(Table 5
表5
表5bc17基因定位部分引物
Table 5
| 标记 Marker | 正向引物 Forward primer (5′-3′) | 反向引物 Reverse primer (5′-3′) |
|---|---|---|
| RM500 | GAGCTTGCCAGAGTGGAAAG | GTTACACCGAGAGCCAGCTC |
| RM3743 | TAGCCTTGTTCCATCCATCC | CTTCTCCCTCTCCTCCTTCC |
| UP-81 | TGCATCTCATCTCCCCTCTT | TGGAGTATAACGCCGACCTC |
| DN-25 | AGGGAAAATGCGCTGAACTA | ATTCATCCATGCCCTATCCA |
新窗口打开|下载CSV
图4
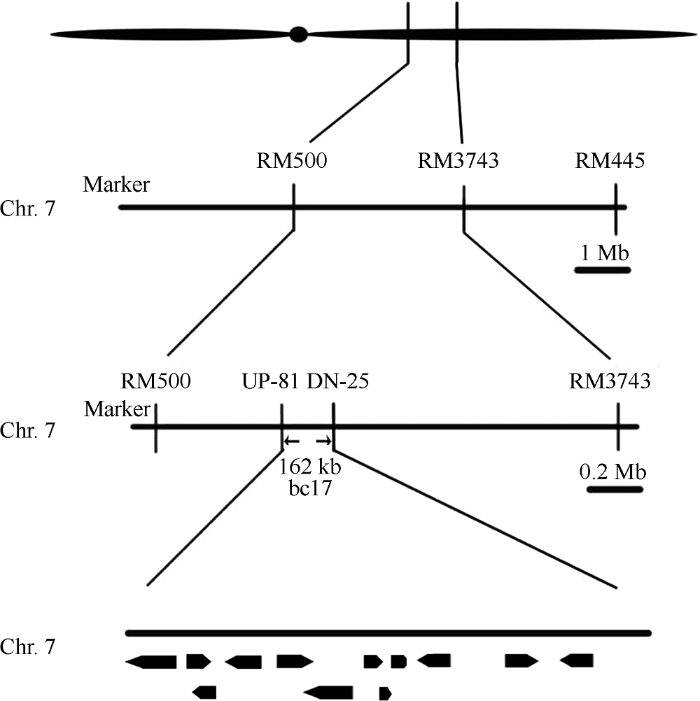 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4利用分子标记对bc17基因定位
Fig. 4Positional mapping of bc17 by molecular makers
3 讨论
目前报道的许多水稻脆性突变体都伴随着明显的表型变化, 如植株矮小, 分蘖数减少, 结实率降低以及产量下降等, 且脆性全生育期、全组织表达, 类似的有bc3和bc12等[23,26]。bc3突变体出现轻微矮化, 与野生型相比bc3突变体的茎、叶、根中纤维素含量降低了28%~36%, 而其他细胞壁组分含量没有发生变化[26]。bc12突变体由于细胞数目的显著减少导致植株矮化, 纤维素微纤维的方向和细胞壁组分改变致使出现脆秆表型[23]。bc17突变体虽然植株变矮, 但是脆性特征比较独特, 具有组织特异性和时间表达特异性, 与其他矮秆脆性突变体具有明显的差异。茎秆和叶片细胞壁成分测定显示, 与野生型相比, bc17茎秆和叶片都出现纤维素含量降低, 半纤维素含量升高。值得一提的是, 与野生型相比, bc17叶片木质素含量降低, 灰分升高, 也可能是导致叶片没有表现出脆性的原因。水稻脆秆性状发生的原因, 除了涉及经典的纤维素合成途径(纤维素合酶)、多糖生物合成转运途径外, 转录因子调控途径以及细胞壁多糖乙酰化修饰等[27]对细胞壁的功能也起到重要的调控作用。叶亚峰等[20]发现OsMYB103L是MYB家族转录因子, C端具有较高的转录活性, 可直接与OsCesA4、OsCesA7、OsCesA9以及bc1启动子区域结合调控他们的表达, 此外, OsMYB103L还参与GA介导的纤维素合成途径的调控。周奕华等发现水稻BS1蛋白为木聚糖乙酰酯酶[28], 可从木糖吡喃糖基残基的O-2和O-3位置的木聚糖骨架上切割乙酰基部分。在维持木聚糖主链上适当的乙酰化水平中起重要作用, 参与次生细胞壁的形态建成, 这表明水稻次生细胞壁的生物合成过程非常复杂。本研究将bc17定位在7号染色体分子标记UP-81和DN-25之间, 没有发现相关报道的其他脆秆基因。在bc17定位区间预测有功能的基因涉及到氨基转移酶、O-甲基转移酶、硫酯酶等, 都有可能影响到细胞壁组分的改变, 而这些基因目前还没有相关研究报道, 可能会为次生细胞壁合成的机制解析发现一条新的调控途径。
农作物传统秸秆处理如弃置农田或者直接焚烧, 由于自然状态下降解速率缓慢, 容易造成环境污染和资源浪费[29]。水稻脆秆突变体, 由于其独特的细胞壁组成和结构的变化, 收获时秸秆易破碎降解, 有利于水稻秸秆的生态还田[30]。同时, 还可以将脆性材料直接作为家畜饲料, 较低的纤维素含量更有利于反刍动物消化, 具有良好的饲料应用前景[31]。水稻秸秆是可再生的生物质原料, 可以经过水解发酵产生生物乙醇[32,33]。黄峰等利用水稻脆性秸秆为原料, 利用酸预处理且酶促糖化发酵72 h后, 脆性秸秆发酵的乙醇产量可达为普通秸秆的1.1倍[34]。bc17突变体细胞壁纤维素含量降低, 半纤维素含量升高, 有望通过优异等位基因的发掘, 应用到生产中, 从秸秆饲料化和能源化角度解决秸秆利用的难题。
4 结论
利用重离子辐照武运粳7号(wyj7)获得一个脆秆突变体bc17, 在抽穗后茎秆出现脆性特征, 且随着生育期进程脆秆脆性程度增加, 叶片基本表现正常。较野生型, bc17叶片和茎秆纤维素含量显著降低, 半纤维素含量显著升高, 厚壁组织变薄, 细胞之间空隙变大, 相应地细胞密度降低, 细胞数目减少。该性状受一对隐性核基因控制, 被定为在7号染色体长臂上162 kb区间内, 生物信息学分析表明bc17可能是一个新的脆秆基因, 为进一步揭示水稻细胞壁合成机制奠定材料基础。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1744-7909.2007.00629.xURLPMID:18713364 [本文引用: 1]

Plant mechanical strength is an important agronomic trait of rice. An ethyl methane sulfonate (EMS)-induced rice mutant, fragile plant 2 (fp2), showed morphological changes and reduced mechanical strength. Genetic analysis indicated that the brittle of fp2 was controlled by a recessive gene. The fp2 gene was mapped on chromosome 10. Anatomical analyses showed that the fp2 mutation caused the reduction of cell length and cell wall thickness, increasing of cell width, and the alteration of cell wall structure as well as the vessel elements. The consequence was a global alteration in plant morphology. Chemical analyses indicated that the contents of cellulose and lignin decreased, and hemicelluloses and silicon increased in fp2. These results were different from the other mutants reported in rice. Thus, fp2 might affect the deposition and patterning of microfibrils, the biosynthesis and deposition of cell wall components, which influences the formation of primary and secondary cell walls, the thickness of cell walls, cell elongation and expansion, plant morphology and plant strength in rice.
DOI:10.3724/SP.J.1006.2008.01417URL [本文引用: 1]
bc7(t)]叶进行了细胞学观察及叶细胞化学组成分析。光镜和电镜观察都发现突变体厚壁细胞的细胞壁变薄; 对细胞壁成分的化学分析显示突变体纤维素含量明显低于对照, 硅含量明显升高, 而木质素变化不明显; 木质素的组化反应也显示了木质素在突变体和对照之间差异不大; X-射线微区分析表明, 硅元素在突变体叶表面明显提高。上述结果表明, 突变体叶纤维素含量的降低影响了厚壁细胞次生壁的形成, 导致细胞壁变薄, 机械强度降低, 硅含量的升高有助于突变体增强机械强度。]]>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:21205179 [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1111/j.1365-313X.2010.04308.xURLPMID:20663087 [本文引用: 1]
Membrane trafficking between the plasma membrane (PM) and intracellular compartments is an important process that regulates the deposition and metabolism of cell wall polysaccharides. Dynamin-related proteins (DRPs), which function in membrane tubulation and vesiculation are closely associated with cell wall biogenesis. However, the molecular mechanisms by which DRPs participate in cell wall formation are poorly understood. Here, we report the functional characterization of Brittle Culm3 (BC3), a gene encoding OsDRP2B. Consistent with the expression of BC3 in mechanical tissues, the bc3 mutation reduces mechanical strength, which results from decreased cellulose content and altered secondary wall structure. OsDRP2B, one of three members of the DRP2 subfamily in rice (Oryza sativa L.), was identified as an authentic membrane-associated dynamin via in vitro biochemical analyses. Subcellular localization of fluorescence-tagged OsDRP2B and several compartment markers in protoplast cells showed that this protein not only lies at the PM and the clathrin-mediated vesicles, but also is targeted to the trans-Golgi network (TGN). An FM4-64 uptake assay in transgenic plants that express green fluorescent protein-tagged OsDRP2B verified its involvement in an endocytic pathway. BC3 mutation and overexpression altered the abundance of cellulose synthase catalytic subunit 4 (OsCESA4) in the PM and in the endomembrane systems. All of these findings lead us to conclude that OsDRP2B participates in the endocytic pathway, probably as well as in post-Golgi membrane trafficking. Mutation of OsDRP2B disturbs the membrane trafficking that is essential for normal cellulose biosynthesis of the secondary cell wall, thereby leading to inferior mechanical properties in rice plants.
DOI:10.1093/jxb/erq395URLPMID:21209026 [本文引用: 1]
The brittle culm (bc) mutants of Gramineae plants having brittle skeletal structures are valuable materials for studying secondary cell walls. In contrast to other recessive bc mutants, rice Bc6 is a semi-dominant bc mutant with easily breakable plant bodies. In this study, the Bc6 gene was cloned by positional cloning. Bc6 encodes a cellulose synthase catalytic subunit, OsCesA9, and has a missense mutation in its highly conserved region. In culms of the Bc6 mutant, the proportion of cellulose was reduced by 38%, while that of hemicellulose was increased by 34%. Introduction of the semi-dominant Bc6 mutant gene into wild-type rice significantly reduced the percentage of cellulose, causing brittle phenotypes. Transmission electron microscopy analysis revealed that Bc6 mutation reduced the cell wall thickness of sclerenchymal cells in culms. In rice expressing a reporter construct, BC6 promoter activity was detected in the culms, nodes, and flowers, and was localized primarily in xylem tissues. This expression pattern was highly similar to that of BC1, which encodes a COBRA-like protein involved in cellulose synthesis in secondary cell walls in rice. These results indicate that BC6 is a secondary cell wall-specific CesA that plays an important role in proper deposition of cellulose in the secondary cell walls.
[本文引用: 1]
[本文引用: 1]
URLPMID:12970476 [本文引用: 2]
DOI:10.1016/S1673-8527(07)60115-5URL [本文引用: 2]
AbstractSeveral brittle culm mutants of rice were identified and characterized. In this study, we characterized a brittle mutant (bc7(t)) identified from japonica variety Zhonghua 11 by means of 60Co-γ radiation. This mutant displays normal phenotype similar to its wild type plants except for the fragility of all plant body, with ∼10% decrease in the cellulose content. The genetic analysis and gene fine mapping showed that the bc7(t) mutant was controlled by a recessive gene, residing on an 8.4 kb region of the long arm of chromosome 1. The gene annotation indicated that there was only one putative gene encoding cellulose synthase catalytic subunit (CesA) in this region, which was allelic to OsCesA4. Furthermore, the sequence analysis was carried out and 7 bases deletion in the junction of exon 10 and intron 10 was done in bc7(t) mutant, resulting in the change of reading frame and the consequent failure to generate functional protein. In addition, the result of RNA interference experiment showed that when the Bc7(t) was knocked down, the transplants exhibited fragility, similar to bc7(t) mutant. The finding of novel allele of OsCesA4 locus will facilitate the understanding of the mechanism of cell wall biosynthesis. The potential utilization of the bc7(t) mutant in animal food was discussed as well.]]>
DOI:10.1007/s11103-009-9536-4URL [本文引用: 2]
Cellulose synthase (CESA) is a critical catalytic subunit of the cellulose synthase complex responsible for glucan chain elongation. Our knowledge about how CESA functions is still very limited. Here, we report the functional characterization of a rice mutant, brittle culm11, that shows growth retardation and dramatically reduced plant strength. Map-based cloning revealed that all the mutant phenotypes result from a missense mutation in OsCESA4 (G858R), a highly conserved residue at the end of the fifth transmembrane domain. The aberrant secondary cell wall of the mutant plants is attributed to significantly reduced cellulose content, abnormal secondary wall structure of sclerenchyma cells, and overall altered wall composition, as detected by chemical analyses and immunochemical staining. Importantly, we have found that this point mutation decreases the abundance of OsCESA4 in the plasma membrane, probably due to a defect in the process of CESA complex secretion. The data from our biochemical, genetic, and pharmacological analyses indicate that this residue is critical for maintaining the normal level of CESA proteins in the plasma membrane.]]>
DOI:10.1093/jxb/erq395URL [本文引用: 2]
The brittle culm (bc) mutants of Gramineae plants having brittle skeletal structures are valuable materials for studying secondary cell walls. In contrast to other recessive bc mutants, rice Bc6 is a semi-dominant bc mutant with easily breakable plant bodies. In this study, the Bc6 gene was cloned by positional cloning. Bc6 encodes a cellulose synthase catalytic subunit, OsCesA9, and has a missense mutation in its highly conserved region. In culms of the Bc6 mutant, the proportion of cellulose was reduced by 38%, while that of hemicellulose was increased by 34%. Introduction of the semi-dominant Bc6 mutant gene into wild-type rice significantly reduced the percentage of cellulose, causing brittle phenotypes. Transmission electron microscopy analysis revealed that Bc6 mutation reduced the cell wall thickness of sclerenchymal cells in culms. In rice expressing a reporter construct, BC6 promoter activity was detected in the culms, nodes, and flowers, and was localized primarily in xylem tissues. This expression pattern was highly similar to that of BC1, which encodes a COBRA-like protein involved in cellulose synthesis in secondary cell walls in rice. These results indicate that BC6 is a secondary cell wall-specific CesA that plays an important role in proper deposition of cellulose in the secondary cell walls.
DOI:10.1093/mp/sst025URL [本文引用: 2]
Tricheary elements (TEs), wrapped by secondary cell wall, play essential roles in water, mineral, and nutrient transduction. Cadmium (Cd) is a toxic heavy metal that is absorbed by roots and transported to shoot, leaves, and grains through vascular systems in plants. As rice is a major source of Cd intake, many efforts have been made to establish 'low-Cd rice'. However, no links have been found between cellulose biosynthesis and cadmium accumulation. We report here a rice brittle culm13 mutant, resulting from a novel missense mutation (G101K) in the N-terminus of cellulose synthase subunit 9 (CESA9). Except for the abnormal mechanical strength, the mutant plants are morphologically indistinguishable from the wild-type plants. Transmission electron microscopy (TEM) and chemical analyses showed a slight reduction in secondary wall thickness and 22% decrease in cellulose content in bc13 plants. Moreover, this mutation unexpectedly confers the mutant plants Cd tolerance due to less Cd accumulation in leaves. Expression analysis of the genes required for Cd uptake and transport revealed complicated alterations after applying Cd to wild-type and bc13. The mutants were further found to have altered vascular structure. More importantly, Cd concentration in the xylem saps from the bc13 plants was significantly lower than that from the wild-type. Combining the analyses of CESA9 gene expression and Cd content retention in the cell-wall residues, we conclude that CESA9(G101K) mutation alters cell-wall properties in the conducting tissues, which consequently affects Cd translocation efficiency that largely contributes to the low Cd accumulation in the mutant plants.
[本文引用: 2]
URLPMID:21383162 [本文引用: 2]
URLPMID:22665444 [本文引用: 2]
URLPMID:26350403 [本文引用: 3]
URLPMID:26002868 [本文引用: 2]
DOI:10.1186/s12284-017-0196-8URLPMID:29305728 [本文引用: 2]
BACKGROUND: Rice zebra mutants are leaf variegation mutants that exhibit transverse sectors of green/yellow or green/white in developing or mature leaves. In most cases, leaf variegation is caused by defects in chloroplast biogenesis pathways, leading to an accumulation of reactive oxygen species in a transverse pattern in the leaves. Here, we examine a new type of leaf variegation mutant in rice, zebra3 (z3), which exhibits transverse dark-green/green sectors in mature leaves and lacks the typical yellow or white sectors. RESULTS: Map-based cloning revealed that the Z3 locus encodes a putative citrate transporter that belongs to the citrate-metal hydrogen symport (CitMHS) family. CitMHS family members have been extensively studied in bacteria and function as secondary transporters that can transport metal-citrate complexes, but whether CitMHS family transporters exist in eukaryotes remains unknown. To investigate whether Z3 acts as a citrate transporter in rice, we measured citrate levels in wild-type leaves and in the dark-green and green sectors of the leaves of z3 mutants. The results showed that citrates accumulated to high levels in the dark-green sectors of z3 mutant leaves, but not in the green sectors as compared with the wild-type leaves. CONCLUSIONS: These results suggest that leaf variegation in the z3 mutant is caused by an unbalanced accumulation of citrate in a transverse pattern in the leaves. Taking these results together, we propose that Z3 plays an important role in citrate transport and distribution during leaf development and is a possible candidate for a CitMHS family member in plants.
DOI:10.1111/j.1365-313X.2010.04238.xURLPMID:20444225 [本文引用: 3]
Kinesins are encoded by a large gene family involved in many basic processes of plant development. However, the number of functionally identified kinesins in rice is very limited. Here, we report the functional characterization of Brittle Culm12 (BC12), a gene encoding a kinesin-4 protein. bc12 mutants display dwarfism resulting from a significant reduction in cell number and brittleness due to an alteration in cellulose microfibril orientation and wall composition. BC12 is expressed mainly in tissues undergoing cell division and secondary wall thickening. In vitro biochemical analyses verified BC12 as an authentic motor protein. This protein was present in both the nucleus and cytoplasm and associated with microtubule arrays during cell division. Mitotic microtubule array comparison, flow cytometric analysis and expression assays of cyclin-dependent kinase (CDK) complexes in root-tip cells showed that cell-cycle progression is affected in bc12 mutants. BC12 is very probably regulated by CDKA;3 based on yeast two-hybrid and microarray data. Therefore, BC12 functions as a dual-targeting kinesin protein and is implicated in cell-cycle progression, cellulose microfibril deposition and wall composition in the monocot plant rice.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00425-010-1145-6URLPMID:20369251 [本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}