 ,1, 魏春茹1, 范润侨1, 于秀梅,1,2,*, 王逍冬2, 赵伟全2, 魏新燕2, 康振生3, 刘大群2
,1, 魏春茹1, 范润侨1, 于秀梅,1,2,*, 王逍冬2, 赵伟全2, 魏新燕2, 康振生3, 刘大群2TaPP2-A13 gene shows induced expression pattern in wheat responses to stresses and interacts with adaptor protein SKP1 from SCF complex
MENG Yu-Yu,1, WEI Chun-Ru1, FAN Run-Qiao1, YU Xiu-Mei,1,2,*, WANG Xiao-Dong2, ZHAO Wei-Quan2, WEI Xin-Yan2, KANG Zhen-Sheng3, LIU Da-Qun2通讯作者:
收稿日期:2020-05-12接受日期:2020-09-13网络出版日期:2020-09-30
| 基金资助: |
Received:2020-05-12Accepted:2020-09-13Online:2020-09-30
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (9677KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孟钰玉, 魏春茹, 范润侨, 于秀梅, 王逍冬, 赵伟全, 魏新燕, 康振生, 刘大群. 小麦TaPP2-A13基因的表达响应逆境胁迫并与SCF复合体接头蛋白TaSKP1相互作用[J]. 作物学报, 2021, 47(2): 224-236. doi:10.3724/SP.J.1006.2021.01042
MENG Yu-Yu, WEI Chun-Ru, FAN Run-Qiao, YU Xiu-Mei, WANG Xiao-Dong, ZHAO Wei-Quan, WEI Xin-Yan, KANG Zhen-Sheng, LIU Da-Qun.
F-box蛋白是SCF (SKP1-Cullin-F-box) 类E3连接酶的重要组分。第一个F-box基序于1996年在Cyclin F蛋白的N端被发现, 并因此而得名[1]。作为SCF复合体中特异性识别待降解底物的决定性蛋白, F-box蛋白广泛地存在于植物中, 负责调控植物的生长发育、激素信号传导、自交不亲和及抵御胁迫等多个代谢过程。拟南芥(Arabidopsis thaliana) F-box蛋白Hawaiian skirt和UFO (Unusual floral organs)均参与花发育的过程[2,3]; 生长素受体TIR1和茉莉酸信号受体蛋白COI1 (Coronatine-insensitive 1)均由F-box基因编码, 能与多种蛋白形成SCFTIR1和SCFCOI1, 继而在植物生长发育及激素介导的生物/非生物逆境过程中发挥作用[4,5]; 植物的自交亲和/不亲和受雌蕊专化的S-RNase基因和多个花粉专化的S-位点F-box (SLF)基因共同调控。矮牵牛(Petunia inflata) F-box基因PiSLF2 (S-locus F-box 2)编码的花粉S-决定因子可与花柱S-决定因子发生相互识别, 从而导致配子体的自交不亲和[6]。据报道, 目前鉴定了17个SLF基因, 其中至少7个参与花粉的专化性识别[7]; 在烟草(Nicotiana tabacum)中过表达小麦F-box基因TaFBA1可增强烟草的抗旱性及对热、盐和氧化胁迫的抵抗力[8,9,10,11]; F-box蛋白MAX2 (More axillary 2)不仅参与植物的发育, 而且在抵御病原细菌丁香假单胞菌(Pseudomonas syringae, P. syringae)和胡萝卜果胶杆菌(Pectobacterium carotovorum, P. carotovorum)侵染及盐和干旱胁迫过程中发挥作用[12]; F-box基因CPR30在拟南芥抵御病原细菌P. syringae的入侵过程中起着负调控作用[13]; 葡萄(Vitis vinifera) F-box基因BIG-24.1不仅受灰霉菌(Botrytis cinerea)的强烈诱导, 对UV-C、伤害、SA、ABA、MeJA、乙烯等非生物胁迫和激素也反应强烈[14]。由此可见, F-box家族在植物中发挥着广泛而重要的作用, 近年来对其在抗病和抗逆中的作用引起愈加广泛重视。
韧皮部为植物远距离信息传递和大分子物质的运输提供了一条有效的途径, 同时韧皮部也是植物防御病原物侵染的战略要地。韧皮部蛋白2 (Phloem protein 2, PP2)是存在于植物韧皮部汁液中的一种最丰富和神秘的蛋白质, 以二聚体存在, 是具有几丁质结合功能的特殊凝集素[15]。韧皮部蛋白AtPP2-B11参与了拟南芥对干旱胁迫的响应, 且发挥负调控作用[16]; 拟南芥中PP2-A1编码韧皮部凝集素类似蛋白, 研究发现该蛋白具有分子伴侣和抗真菌侵染的双重作用[17]; 南瓜(Cucurbita maxima)的韧皮部分泌蛋白PP2可以将细菌和真菌固定在交联的细丝上, 从而封闭受伤的韧皮部筛管, 使之免受病原菌的进一步侵染[18]; 黄龙病是柑橘(Citrus reticulate)上发生的一种世界级毁灭性病害。长春花(Catharanthus roseus)作为柑橘黄龙病的理想研究材料, 在受到黄龙病菌(Candidatus Liberibacter asiaticus, CLas)感染后, 长春花中的PP2类蛋白呈现强烈诱导表达, 这可能导致胼胝质沉积, 筛孔堵塞筛孔, 从而阻止病原菌的侵染[19]; 此外, PP2类蛋白CsPP2B15在高感CLas的品种锦橙(Citrus sinensis)嫩叶中受CLas诱导显著上调表达, 而在耐病品种酸柚(Citrus grandis)的嫩叶中受CLas诱导显著下调表达[20]; 苎麻(Boehmeria nivea)中鉴定了15个PP2蛋白, 这些蛋白的编码基因在响应非生物逆境、昆虫及机械伤害的过程中呈现不同的反应[21]。以上报道表明, 韧皮部蛋白2在抵御病原菌侵染, 昆虫和机械伤害及非生物胁迫中均发挥重要作用。
综上, F-box蛋白家族尤其韧皮部蛋白2亚家族(F-box/PP2, FBP)在植物抵御病原菌侵染和非生物胁迫过程中发挥了重要的调节作用。FBP成员在小麦中的功能如何, 是否参与小麦抵御生物及非生物胁迫过程, 目前尚未见报道。鉴于此, 本研究以小麦抗叶锈病近等基因系TcLr15为材料, 在获得小麦TaPP2-A13基因全长编码区的基础上, 借助生物信息学分析、qRT-PCR、亚细胞定位、酵母双杂交(Yeast 2 Hybrid, Y2H)、双分子荧光互补(Bimolecular Fluorescence Complementation, BiFC)和免疫共沉淀(Co-Immunoprecipitation, Co-IP)等技术, 明确该基因在逆境胁迫下的表达模式, 确定其亚细胞定位, 并获得与之互作的蛋白。研究结果将拓宽小麦中F-box家族基因的功能认识, 为深入解析TaPP2-A13在小麦抵御生物和非生物胁迫中的作用机制奠定了基础。
1 材料与方法
1.1 小麦和叶锈菌材料及处理方法
小麦抗叶锈病近等基因系TcLr15、小麦高感叶锈病品种郑州5389、供试叶锈菌株05-19-43②和05-5-137③均由河北省农作物病虫害生物防治技术创新中心提供。郑州5389用于扩繁叶锈菌株05-19-43②和05-5-137③的夏孢子, 方法参照Kolmer[22]进行。小麦TcLr15幼苗分别进行叶锈菌(05-19-43②和05-5-137③)侵染、激素(100 μmol L-1 ABA、200 μmol L-1 MeJA和2 mmol L-1 SA)和非生物逆境(10% PEG、7 mmol L-1 H2O2和300 mmol L-1 NaCl)处理, 取样时间及处理方法同于孟钰玉等[23]的方法。
1.2 TaPP2-A13基因全长扩增
以小麦TcLr15一叶一心期的叶片cDNA作为PCR扩增的模板。自小麦转录组数据库ExpVIP (wheat expression browser,Table 1
表1
表1本实验中涉及的引物信息
Table 1
| 引物名称 Primer name | 上游引物序列 Forward primer sequence (5'-3') | 下游引物序列 Reverse primer sequence (5'-3') | 退火温度 Annealing temperature (℃) |
|---|---|---|---|
| TaPP2-A13-F/R | AATGGCGGAATCCCTCGTG | CGGGTTTTGGACAAGAATGG | 58.0 |
| TaPP2-A13-qPCR-F/R | ATCGATGATCGGCGGTATTG | AGTGGAGCCGGAAGAGAAGG | 59.5 |
| TaGAPDH-qPCR-F/R | CTGCCTTGCTCCTCTTGCTAA | CTTGATGGAAGGACCAGCAAC | 59.5 |
| TaPP2-A13-His-F/R | gctgatatcggatccgaattcATGGGGGC GGGGGCTTCG | tgcggccgcaagcttgtcgacTTACTTGCGTATGC ACTCCTCG | 60.0 |
| TaPP2-A13-GFP-F/R | atacaccaaatcgactctagaATGGGGGC GGGGGCTTCG | catggtaccggatccactagtCTTGCGTATGCACT CCTCGG | 60.0 |
| TaPP2-A13-BD-F/R | CGgaatccATGGGGGC | ggatccTTACTTGCGTATGCACT | 60.0 |
| AD-F/R | TAATACGACTCACTATAGGGCG | AGATGGTGCACGATGCACAG | 56.0 |
| TaPP2-A13-NE-F/R | cccaggcctactagtggatccATGGGGGC GGGGGCTTCG | agcggtaccctcgaggtcgacCTTGCGTATGCACT CCTCGG | 60.0 |
| TaSKP1-FLAG-F/R | gagaacacgggggactctagaATGGCGG CCGCGGGAGAC | cgtcctaggcttaagtctagaCTCAAAGGCCCACT GGTTCTC | 60.0 |
| TaPP2-A13-HA-F/R | agacttaagcctaggacgcgtATGGGGGC GGGGGCTTCG | AtcgtatgggtacatacgcgtCTTGCGTATGCACT CCTCGG | 60.0 |
新窗口打开|下载CSV
1.3 生物信息学分析
DNAStar软件将TaPP2-A13基因的编码区翻译为蛋白质序列; ProtParam (1.4 TaPP2-A13基因表达模式分析
以1.2获得的TaPP2-A13基因全长序列为模板设计qRT-PCR引物: TaPP2-A13-qPCR-F/ TaPP2-A13-qPCR-R (表1)。RNA提取及第1链cDNA合成同于1.2。以小麦三磷酸甘油醛脱氢酶(Glyceraldehyde-3-phosphate dehydrogenase, TaGAPDH, 登录号为EU022331.1)为内参基因, 引物TaGAPDH- qPCR-F和TaGAPDH-qPCR-R序列见表1。qRT-PCR参照2×TransStart Tip Green qPCR SuperMix (TransGen, 中国)说明书进行。反应程序为94℃ 30 s; 94℃ 5 s, 59.5℃ 15 s, 72℃ 10 s, 40个循环。TaPP2-A13相对表达量采用2-ΔΔCt计算, 利用Microsoft Excel 2013进行平均值和标准误差的计算, 利用SAS 9.4软件的ANOVA进行方差分析。1.5 亚细胞定位
根据目的基因编码区和GFP融合表达载体pSuper1300-GFP序列设计引物TaPP2-A13-GFP-F和TaPP2-A13-GFP-R (表1)。以pMD19-T-TaPP2-A13为模板扩增得到带有Xba I和Spe I酶切位点的TaPP2-A13, 扩增体系与程序同于1.2。扩增产物与pSuper1300-GFP载体连接, 构建GFP融合蛋白的瞬时表达载体。测序验证正确的重组质粒转入农杆菌GV3101菌株, 注射器吸取该菌液注入6~8周龄的烟草叶片。25℃培养30 h后, 以GFP-HQ荧光滤光块(激发滤色镜470 nm/40、分光镜495 nm和吸收滤色镜525 nm/50)在尼康倒置显微镜Ti2-U (Nikon, 日本)下观察GFP绿色荧光, 以确定TaPP2-A13的亚细胞定位情况。1.6 Y2H分析
用模板pMD19-T-TaPP2-A13及引物TaPP2- A13-BD-F和TaPP2-A13-BD-R (表1)进行PCR扩增, 将酶切位点引入TaPP2-A13两侧, 扩增体系和程序同于1.2。PCR产物经EcoR I和Bam H I双酶切后, 与同样双酶切的pGBT9 (BD)载体连接, 重组载体命名为pGBT9-TaPP2-A13 (BD-TaPP2-A13)。将其转化酵母Y187后, 涂布在SD/-Trp (一缺)、SD/-Trp/-Leu (二缺)、SD/-Leu-Trp-His (三缺)和SD/-Trp/-Leu/- His/-Ade (四缺)固体培养基上, 观察酵母生长状况, 进行毒性和自激活活性验证。实验室前期已构建TcLr15接种叶锈菌株05-19-43②的抗叶锈病性酵母AH109文库[24], 用诱饵载体BD-TaPP2-A13筛选该文库, 以获取与TaPP2-A13互作的蛋白。具体操作参见Hong等[25]的方法。以四缺平板上的单克隆为模板, 利用引物TaPP2-A13-BD-F和TaPP2-A13-BD-R进行菌落PCR, 确认TaPP2-A13是否存在。再以pGADT7(AD)载体的通用引物AD-F和AD-R (表1)进行增, 鉴定捕获蛋白基因及其片段大小。挑选条带单一( >500 bp)的菌落PCR产物送北京中科希林生物科技有限责任公司测序。
通过蛋白一对一互作方法验证文库筛选结果的准确性。首先将获得的序列在NCBI数据库进行BLAST比对, 根据比对结果设计各筛选基因的全长扩增引物, 以小麦TcLr15叶片cDNA为模板, 获得各筛选基因的全长编码区。然后将各个筛选基因的编码区序列分别与AD载体连接, 构建AD-Prey载体。最后将AD-Prey和BD-TaPP2-A13共转入酵母菌AH109中, 共转化后的菌液经验证后, 再分别稀释为1∶1、1∶10和1∶100, 各取0.5 μL稀释后菌液接种至含有X-α-Gal的四缺平板上进一步筛选。
1.7 BiFC分析
根据目的基因编码区和BiFC载体pSPY-NE序列设计引物TaPP2-A13-NE-F和TaPP2-A13-NE-R (表1)。以pMD19-T-TaPP2-A13为模板扩增得到带有Bam H I和Sal I酶切位点的TaPP2-A13, 扩增体系与程序同于1.2。扩增产物与pSPY-NE载体连接, 构建BiFC载体pSPY-NE-TaPP2-A13, 随后将其转入农杆菌GV3101。同时将pSPY-CE和pSPY- CE-TaSKP1 (已构建)分别转入农杆菌GV3101。将转化pSPY-CE-TaSKP1的农杆菌与转化pSPY-NE- TaPP2-A13的农杆菌混合, 以pSPY-CE和pSPY-NE-TaPP2-A13混合菌液为阴性对照, 用注射器吸取各混合菌液注入6~8周龄的烟草叶片。25℃培养40 h后, 观察荧光信号及其分布, 仪器及设置同于1.5。1.8 Co-IP分析
以TaSKP1和TaPP2-A13的编码区序列分别设计引物TaSKP1-FLAG-F/TaSKP1-FLAG-R和TaPP2- A13-HA-F/TaPP2-A13-HA-R (表1)。以AD-TaSKP1为模板扩增得到带有Xba I的TaSKP1, 以pMD19- T-TaPP2-A13为模板扩增得到带有Mlu I酶切位点的TaPP2-A13, 扩增体系与程序同于1.2。pTF101-3FLAG和pTF101-6HA分别经单酶切(Xba I和Mlu I)后与扩增产物连接, 构建Co-IP载体pTF101-3FLAG-TaSKP1和pTF101-6HA-TaPP2-A13, 然后将重组质粒转化农杆菌GV3101。将含有重组质粒的农杆菌(OD600为0.8) 1∶1混匀后注射6~8周龄的烟草叶片。暗培养过夜, 然后转入正常光周期(16 h光照/8 h黑暗), 25℃培养40 h后, 剪取注射的叶片, 提取其总蛋白。以带有第一抗体anti-HA的beads孵育总蛋白。未经孵育的总蛋白和孵育后的免疫沉淀物分别用anti-HA和anti-FLAG杂交, 以第二抗体羊抗兔IgG-HRP结合第一抗体。杂交完成的PVDF膜漂洗干净后淋上混合液(等体积的A液和B液), 于Odyssey Fc Imaging System (LI-COR Biosciences, 美国)上以固定程序Chemi曝光2 min, 以显示目的蛋白。
2 结果与分析
2.1 获得一个小麦韧皮部蛋白2编码基因TaPP2-A13
以苗期小麦TcLr15的叶片cDNA为模板, 运用TaPP2-A13-F和TaPP2-A13-R扩增得到一特异条带, 大小与预期相符, 测序后确认该序列长度为1361 bp。在小麦转录组数据库中其ID为Traes CS2D02G397700, 说明该基因序列位于小麦2D染色体上。在NCBI网站进行BLASTx比对, 结果表明, 该序列与普通小麦D染色体组的供体种粗山羊草(Aegilops tauschii, 登录号为XP_020187178.1)的PP2-A13-like相似性最高, 为100%; 其次与二穗短柄草(Brachypodium distachyon, 登录号为XP_010240281.1)中对应序列相似性为90.66%; 与短花药野生稻(Oryza brachyantha, 登录号为XP_006652663.1, 84.87%)和小米(Setaria italic, 登录号为XP_004976538.1, 80.53%)等禾本科植物的PP2-A13/PP2-A13-like相似性均大于75%。根据以上比对结果, 将该基因命名为TaPP2-A13 (登录号为MT364339)。TaPP2-A13基因编码一个由298个氨基酸组成的蛋白质多肽链, 其相对分子量为33.18 kD, 理论等电点为6.36, 平均亲水系数为-0.322, 属于亲水性蛋白。TaPP2-A13蛋白第22~62位是F-box结构域, 第128~287位具有PP2结构域, 为F-box蛋白家族中FBP亚家族。经预测, 该蛋白二级结构中α-螺旋占22.82%, β-转角占6.04%, 无规则卷曲和β-折叠分别为46.31%和24.83%。三维结构主要由无规则卷曲、α-螺旋和β-折叠构成, 与二级结构预测结果基本一致。TaPP2-A13蛋白不含信号肽序列和跨膜结构域, 暗示其既非分泌蛋白, 亦非跨膜蛋白。亚细胞定位预测显示, TaPP2-A13定位在细胞核上。
利用Clustal X软件对小麦TaPP2-A13与其他禾本科植物PP2-A13进行氨基酸序列比对, 发现各氨基酸序列间均具有较高的相似性, 尤其是在F-box结构域和PP2结构域2个区域。F-box结构域共包含41个氨基酸, 其中31个氨基酸是完全保守的, 5个氨基酸位点仅在一个物种(1个在二穗短柄草, 1个在短花药野生稻和3个在玉米)中有差别。PP2结构域共包含160个氨基酸, 其中112个氨基酸是完全保守的, 有12个氨基酸是仅在一个物种(1个在二穗短柄草, 1个在II型少花古尔德草(Dichanthelium oligosanthes), 1个在短花药野生稻, 2个在水稻, 1个在哈氏黍(Panicum hallii), 1个在黍子(Panicum miliaceum), 2个在狗尾草和3个在玉米)中有差别。高度保守的氨基酸说明其对于维持PP2-A13/PP2-A13-like的功能至关重要(图1)。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1TaPP2-A13与其他禾本科植物PP2-A13氨基酸序列比对
黑色框区域为F-box结构域, 灰色框区域为PP2结构域; 所涉及物种为普通小麦、粗山羊草、二穗短柄草、II型少花古尔德草、短花药野生稻、粳稻、哈氏黍、糜子、谷子、狗尾草和玉米。
Fig. 1Alignment for amino acid sequence between TaPP2-A13 and other PP2-A13 from gramineous plants
Region with black box represents F-box domain, region with gray frames represents PP2 domain; plant species used in this study are Triticum aestivum, Aegilops tauschii, Brachypodium distachyon, Dichanthelium oligosanthes, Oryza brachyantha, Oryza sativa, Panicum hallii, Panicum miliaceum, Setaria italica, Setaria viridis, and Zea mays.
miliaceum)等的PP2-A13亲缘关系次之。稻类和弯叶画眉草(Eragrostis curvula)的采用MEGA7中的NJ (Neighbor-Joining)法构建系统发育树(图2)。根据亲缘关系的远近将进化树分为5个组。与小麦亲缘关系最近的为小麦近缘种属植物, 如粗山羊草、大麦(Hordeum vulgare)、二穗短柄草等, 其PP2-A13位于同一进化分支上(V组)。其中TaPP2-A13与粗山羊草AetPP2-A13的亲缘关系最近, 其氨基酸序列完全一致。TaPP2-A13与IV组的玉米、高粱(Sorghum bicolor)、谷子(Setaria italica)和糜子(Panicum PP2-A13聚为一类(III组)。乌拉尔图小麦处于单独的分支, 为II组。木本植物如胡杨树(Populus euphratica)、小垫柳(Salix brachista)和银叶玫瑰木(Rhodamnia argentea)等及双子叶植物蒺藜苜蓿(Medicago truncatula)和石斛兰(Dendrobium catenatum)等的PP2-A13聚为一类(I组)。I组、II组和III组的PP2-A13序列与第V组相应序列的亲缘关系更远些。
图2
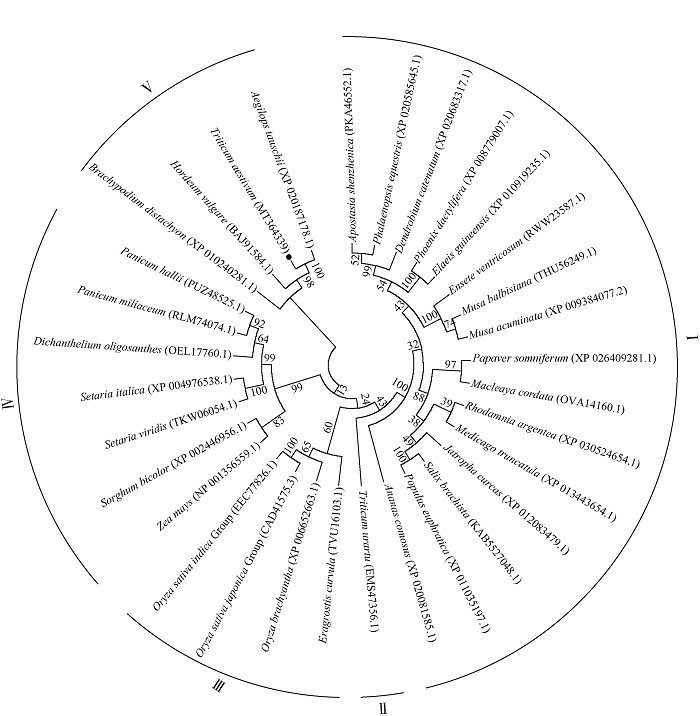 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2植物PP2-A13系统发育树图
进化树外部的实线: 植物PP2-A13蛋白的进化分支; ●: TaPP2-A13蛋白; 所涉及物种为深圳拟兰、小兰屿蝴蝶兰、石斛兰、海枣、油棕榈、食用象腿蕉、野蕉、马拉西亚野生香蕉、罂粟、博落回、银叶玫瑰木、蒺藜苜蓿、麻风树、小垫柳、胡杨树、凤梨、乌拉尔图小麦、弯叶画眉草、短花药野生稻、粳稻、籼稻、玉米、高粱、狗尾草、谷子、II型少花古尔德草、糜子、哈氏黍、二穗短柄草、大麦、普通小麦和粗山羊草。
Fig. 2Phylogenetic tree of plant PP2-A13
Solid line outside of the phylogenetic tree: the clade of plant PP2-A13 proteins; ●: TaPP2-A13 protein; Plant species used are Apostasia shenzhenica, Phalaenopsis equeatris, Dendrobium catenatum, Phoenix dactylifera, Elaeis guineensis, Ensete ventricosum, Musa balbisiana, Musa acuminate, Papaver somniferum, Macleaya cordata, Rhodamnia argentea, Medicago truncatula, Jatropha curcas, Salix brachista, Populus euphratica, Ananas comosus, Triticum urartu, Eragrostis curvula, Oryza brachyantha, Oryza sativa japonica Group, Oryza sativa indica Group, Zea mays, Sorghum bicolor, Setaria viridis, Setaria italica, Dichanthelium oligosanthes, Panicum miliaceum, Panicum hallii, Brachypodium distachyon, Hordeum vulgare, Triticum aestivum, and Aegilops tauschii.
2.2 小麦中TaPP2-A13在基因水平的表达响应激素处理及生物和非生物胁迫
为了解TaPP2-A13基因受激素诱导表达的情况, 分别对小麦TcLr15进行ABA、SA和MeJA处理。经3种外源激素处理后, 随处理时间延长, TaPP2-A13基因均呈现稳步地显著上升表达趋势。ABA处理0.5 h时, TaPP2-A13基因的表达量与0 h样品无异, 自处理2 h时开始升高, 12 h时的表达量约为0 h样品的5倍, 24 h突然降低至0 h样品的3倍左右, 然后再次升高, 在所取样时间范围内, 48 h时表达量达到最高, 为0 h样品的5.3倍; SA处理小麦叶片6 h时, TaPP2-A13基因的表达量与0 h样品差异不大, 自12 h开始表达量升高, 24 h时TaPP2-A13基因表达量达到最大, 为0 h样品的6倍, 48 h虽然有所下降但仍为对照的5.2倍; MeJA处理后, 小麦TaPP2-A13基因的表达量在6 h时开始缓慢升高, 在所取样品的时间范围内, 48 h时表达量达到最高, 为0 h样品的2.8倍(图3)。经3种抗病/抗逆相关激素处理小麦TcLr15后, SA和ABA可以诱发TaPP2-A13基因更高的表达, 其中, ABA呈现更早的诱发, 而SA诱导TaPP2-A13基因的表达更强。图3
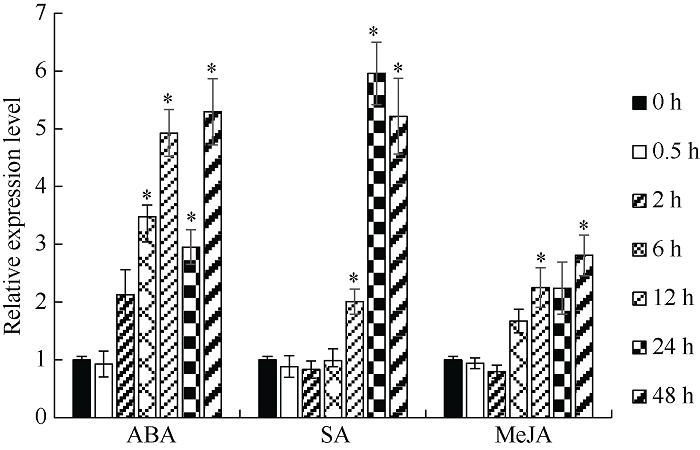 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同外源激素处理下小麦TcLr15中TaPP2-A13基因的表达模式
内参基因为TaGAPDH; *表示不同时间点样品与0 h样品相比差异显著(P < 0.05); 误差线为每组处理的标准误差(n = 3)。
Fig. 3Expression patterns of TaPP2-A13 gene under different exogenous hormones treatment in wheat
TaGAPDH served as a reference gene. * indicates signi?cant differences at P < 0.05 in samples at different treatment time points compared to the sample at 0 hour. Error bars represent the standard error of each sample (n = 3).
本研究设置3种非生物逆境处理小麦TcLr15幼苗。在H2O2胁迫下, 随着处理时间延长, TaPP2-A13基因整体呈现下调的表达趋势。0.5 h的表达量与0 h样品几乎无差异, 自2 h开始表达量持续下降, 至48 h时表达量仅为0 h的1/5; 在PEG模拟的干旱胁迫条件下, 从0.5 h开始TaPP2-A13基因表达量即出现明显下降, 在所取样的时间范围内, 24 h时表达量最低, 仅为对照的3/50; NaCl处理后, TaPP2-A13基因在前3个时间点出现明显地上调表达, 6 h时表达量为0 h的4.8倍, 而后表达量显著下调, 至24 h时达到最低值, 为0 h样品的2/5 (图4-A)。3种逆境条件下, 氧化胁迫和干旱胁迫使TaPP2-A13基因呈现显著地下调表达, 而盐胁迫则诱导TaPP2-A13基因在短时间内显著上调表达。
图4
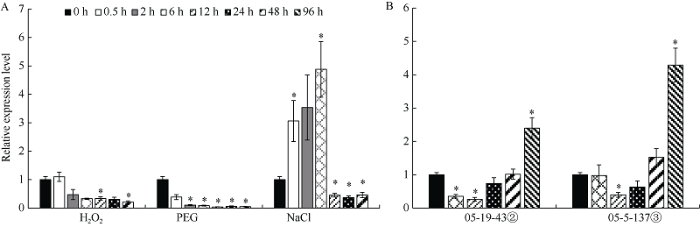 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同生物/非生物胁迫下小麦TcLr15中TaPP2-A13基因的表达模式
内参基因为TaGAPDH; *表示不同时间点样品与0 h样品相比差异显著(P < 0.05); 误差线为每组处理的标准误差(n = 3)。
Fig. 4Expression patterns of TaPP2-A13 gene under different biotic/abiotic stresses in wheat
TaGAPDH served as a reference gene. * indicates signi?cant differences at P < 0.05 in samples at different treatment time points compared to the sample at 0 hour. Error bars represent the standard error of each sample (n = 3).
小麦TcLr15与叶锈菌形成的感病(05-5-137③)及抗病(05-19-43②)组合中, TaPP2-A13基因均呈现先下调再上调的表达趋势。在抗病组合中, 接种叶锈菌株05-19-43② 6 h后, TaPP2-A13的表达量即开始下降, 12 h达到最低, 为0 h的1/4, 随后开始升高, 48 h时的表达量恢复到与0 h样品相同水平, 96 h时升高到0 h样品的2.4倍; 在感病组合中, 接菌6 h的样品表达量与0 h样品无显著变化, 12 h出现显著下调, 之后表达量开始升高, 在取样的时间范围内96 h表达量达到最高值, 为0 h样品的4.3倍(图4-B)。比较抗/感病组合间表达的趋势, 发现TaPP2-A13在感病组合各个时间点的表达量均高于抗病组合。
2.3 TaPP2-A13亚细胞定位于细胞核和细胞质中
为明确TaPP2-A13在细胞内发挥生物学功能的场所, 本试验对其亚细胞定位进行观察。结果如图5所示, 25℃培养30 h时, 阳性对照叶片中的绿色荧光遍布整个视野, 表明实验体系及烟草生长状况均较好。但此时样品中尚无绿色荧光出现。25℃继续培养至48 h时, 在细胞核和细胞质中可见较强绿色荧光。以上观察结果表明TaPP2-A13定位在细胞核和细胞质中。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5TaPP2-A13-GFP融合蛋白的亚细胞定位
绿色表示GFP蛋白在荧光显微镜下所发出的绿色荧光; 标尺为100 μm。
Fig. 5Subcellular localization of TaPP2-A13-GFP protein
The green fluorescence under fluorescence microscope represents GFP protein. Bar = 100 μm.
2.4 TaPP2-A13与TaSKP1在细胞内相互作用
2.4.1 酵母双杂交确定5种蛋白与TaPP2-A13存在物理相互作用 为解析TaPP2-A13在细胞内发挥作用的机制, 拟筛选胞内与其有相互作用的蛋白并进行验证。首先构建诱饵表达载体BD-TaPP2-A13, 将其转化酵母感受态细胞Y187后涂布于一缺、二缺、三缺和四缺平板上。经验证, 重组质粒对酵母细胞无毒性, 亦无自激活能力。以BD-TaPP2-A13为诱饵载体, 筛选酵母文库。挑选20个扩增条带单一且条带大于500 bp的克隆进行序列测定。测序结果在NCBI网站上进行BLASTx比对, 结果表明11种候选蛋白可能与TaPP2-A13存在相互作用(表2)。其中一部分与植物光合和呼吸作用相关, 如核酮糖二磷酸羧化酶小链(ribulose-bisphosphate carboxylase small chain, RbcS)、核酮糖二磷酸羧化酶活化酶B (ribulose bisphosphate carboxylase activase B, RcaB)、RUBISCO活化酶β基因(rca1_beta gene for RUBISCO activase beta, TaRca1_beta)、核酮糖活化酶A (ribulose activase A, RcaA)和Fe-S蛋白的前体蛋白(Fe-S precursor protein)等[26,27]。另有一部分与植物生长发育、抗病及激素信号转导相关, 如SCF复合体骨架蛋白SKP1[28]、UDP葡萄糖类黄酮3-O-葡糖基转移酶7 (UDP-glucose flavonoid 3-O- glucosyltransferase 7, UFGT7)[29]、蛋白磷酸酶(Protein phosphatase 2C 5, PP2C5)[30]、几丁质酶(Chitinase, CHI)[31]和酮酰CoA硫醇酶(3-ketoacyl- CoA thiolase 2, KAT2)[32]及SEC1蛋白家族的SLY1 (SEC1 family transport protein SLY1)[33]等。
Table 2
表2
表2TaPP2-A13筛选酵母文库的结果
Table 2
| 登录号 Accession No. | 蛋白名称 Protein name | 克隆数 Number of clones |
|---|---|---|
| XP_020153404.1 | Protein phosphatase 2C 5 (PP2C5) | 1 |
| KU857044.1 | S-phase kinase-associated protein 1 (SKP1) | 1 |
| XM_020332687.1 | UDP-glucose flavonoid 3-O-glucosyl transferase 7-like | 3 |
| EMS45698.1 | SEC1 family transport protein SLY1 | 1 |
| XM_020338494.1 | 3-ketoacyl-CoA thiolase 2, peroxisomal-like | 2 |
| LM992844.1 | Rca1_beta gene for RUBISCO activase beta (TaRca1_beta) | 3 |
| KC776912.1 | Ribulose activase A | 2 |
| AK330458.1 | Ribulose-bisphosphate carboxylase small chain (RbcS) | 1 |
| AF251264.1 | Ribulose bisphosphate carboxylase activase B (RcaB) | 3 |
| AY123222.1 | Putative Fe-S precursor protein | 2 |
| BAB82472.1 | Chitinase 2 | 1 |
新窗口打开|下载CSV
根据文库筛选获得序列, 结合实验室前期研究结果, 开展酵母一对一互作验证。构建的重组质粒AD-prey (筛选基因)分别与BD-TaPP2-A13共转化酵母菌株AH109, 在含有X-α-Gal的四缺固体培养基上进行回复验证。结果如图6所示, AD-TaPP2C5、AD-TaSLY1、AD-TaCHI、AD-TaRbcS和AD-TaSKP1分别与BD-TaPP2-A13形成的5个组合在含有X-α-Gal的四缺培养基上均能正常生长, 且菌落呈现蓝色, 初步证明TaPP2C5、TaSLY1、TaCHI、TaRbcS和TaSKP1与TaPP2-A13存在物理性相互作用。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6Y2H验证TaPP2-A13与筛选蛋白间的相互作用
Fig. 6Interaction of TaPP2-A13 and screening proteins identified by Y2H
2.4.2 BiFC确定TaPP2-A13与TaSKP1存在相互作用 将TaSKP1和TaPP2-A13分别与载体pSPY-CE和pSPY-NE连接, 构建pSPY-CE-TaSKP1和pSPY-NE-TaPP2-A13载体。将pSPY-CE和pSPY-CE-TaSKP1分别与pSPY-NE-TaPP2-A13混合, 注射烟草叶片, 培养40 h后进行荧光观察。pSPY-CE和pSPY-NE-TaPP2-A13形成的阴性对照中无荧光信号, 实验组合中可见绿色荧光信号, 且荧光信号主要集中在细胞核和细胞质(图7), 表明TaSKP1与TaPP2-A13存在相互作用。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7BiFC验证TaPP2-A13和TaSKP1 的相互作用
绿色表示GFP蛋白在荧光显微镜下所发出的绿色荧光; 标尺为100 μm。
Fig. 7Interaction of TaPP2-A13 and TaSKP1 identified by BiFC
The green fluorescence under fluorescence microscope represents GFP protein. Bar = 100 μm.
2.4.3 Co-IP证实TaPP2-A13与TaSKP1在胞内存在相互作用 构建重组载体pTF101-3FLAG- TaSKP1和pTF101-6HA-TaPP2-A13, 分别将其转入农杆菌GV3101, 混合2种菌液后共注射6~8周龄本氏烟草叶片。提取烟草总蛋白, 利用HA和FLAG抗体分别进行杂交实验。已知TaPP2-A13的预测分子量为33.18 kD, TaSKP1的预测分子量为19.06 kD, 载体上HA和FLAG标签蛋白的分子量分别为7.54 kD和4.06 kD。融合蛋白TaPP2-A13-6HA和TaSKP1-3FLAG的分子量大小应该为40.72 kD和23.12 kD。由图8可知, 未瞬时表达TaPP2-A13-6HA和TaSKP1-3FLAG的烟草叶片, 其全蛋白无杂交的目的条带, 而在瞬时表达TaPP2-A13-6HA和TaSKP1-3FLAG的材料中, 其全蛋白与HA抗体或FLAG抗体分别杂交获得目的蛋白。进一步利用带有anti-HA的磁珠进行总蛋白的免疫沉淀, 沉淀蛋白中富含TaPP2-A13-6HA和TaSKP1-3FLAG, 说明TaPP2-A13和TaSKP1在植物细胞内是互作的(图8)。
图8
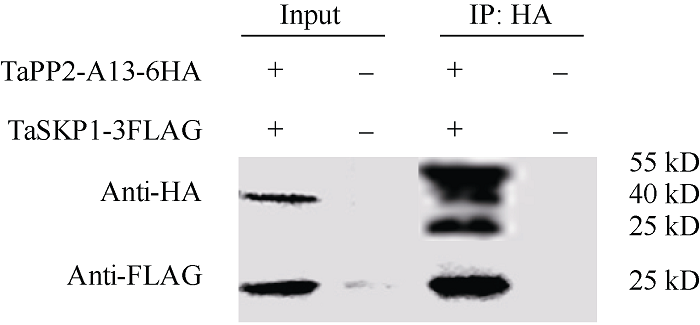 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8Co-IP分析本氏烟草叶片中TaPP2-A13与TaSKP1的互作
将烟草总蛋白提取物与带有anti-HA的beads一起孵育, 通过anti-FLAG和anti-HA进行免疫沉淀物分析。
Fig. 8Interaction of TaPP2-A13 and TaSKP1 in Nicotiana benthamiana leaves by Co-IP
Total protein extracts of N. benthamiana were incubated with anti-HA affinity beads, the immunoprecipitates were analyzed by immunoblotting with anti-FLAG and anti-HA antibodies.
3 讨论
利用同源克隆技术自小麦TcLr15中首次获得了TaPP2-A13基因, 其编码蛋白的N端为一个保守的F-box结构域, C端具有一个PP2结构域, 属于典型的FBP亚家族。本实验室前期已得到了小麦F-box基因TaFBL14和TaSKP2A[34,35], 这2个基因与TaPP2-A13虽然同属于F-box家族, 不同的是, TaFBL14和TaSKP2A的C端含有典型的LRR结构域, 属于FBL亚家族, 同时实验室也已获得了C端含有Kelch结构域的FBK亚家族成员[36]。Zhou等[8]获得的TaFBA1属于FBA亚家族。由此可见, 小麦中存在不同类型的F-box亚家族成员。各个亚家族成员是否存在功能差异尚未可知。近年来, F-box蛋白在植物抵御病原细菌和真菌侵染中的作用日益引起关注。例如, F-box基因CPR30在拟南芥抵御病原细菌P. syringae的入侵过程中起着负调控作用[13]; 棉花(Gossypium hirsutum) F-box蛋白GhACIF1通过与GhSKP1互作增强陆地棉对棉花黄萎病致病菌-大丽轮枝菌(Verticillium dahliae)的抗性[37]; BIG-24.1是分离自葡萄叶片中的F-box蛋白, 当葡萄受到灰霉菌感染时, 叶片及浆果中BIG-24.1的表达量显著上升[14]; 此外, SON1、MAX2、COI1、Nictaba等F-box家族成员在植物抵御病原菌侵染过程中均发挥着重要作用[38,39,40]。但尚未见F-box蛋白在小麦抵御叶锈菌侵染过程中发挥作用的报道。本研究对小麦叶片中TaPP2-A13基因受叶锈菌侵染后的表达情况进行分析, 发现随叶锈菌侵染进程的延伸, 该基因在抗病及感病组合中均呈现上调表达, 且在感病组合中呈现更加强烈的上调(96 h时, 感病组合的表达量是抗病组合的2倍)。长春花在遭受黄龙病菌感染后, 韧皮部蛋白基因PP2-B11和PP2-B15显著上调, 而PP2-A1和PP2-A13则下调。PP2-A13在受到不同病原菌侵染后呈现差异的表达模式。柑橘黄龙病致病菌为细菌, 而小麦叶锈病菌为专性寄生真菌, 细菌和真菌在侵染植物后引起的抗/感病路径不同, 启动抗病相关基因的表达不同, 这可能是造成相同基因在不同植物-病原物组合中表达模式不同的主要原因。
植物F-box蛋白也响应各种非生物胁迫过程。例如, FBP7、DOR、MAX2等在响应非生物胁迫方面均发挥重要作用[12,41-42]。TaPP2-A13基因受MeJA诱导呈现上调表达, 在SA、ABA和NaCl处理后呈现先上调后下调的表达趋势, 经H2O2和PEG处理后, TaPP2-A13随处理时间延长总体呈现显著下调的表达模式, 说明TaPP2-A13的表达受到包括叶锈病菌侵染、外源激素和非生物逆境在内的共同影响。相同的结果见于F-box基因BIG-24.1, 该基因不仅受灰霉菌的强烈诱导, 对UV-C、伤害、SA、ABA、MeJA、乙烯等非生物胁迫和激素也反应强烈[14]。这说明植物F-box家族的一些成员参与多种非生物胁迫的调节过程。
本研究试图找到胞内与TaPP2-A13具有相互作用的蛋白。通过筛选抗叶锈病小麦TcLr15的酵母cDNA文库, 获得了11种可能与TaPP2-A13互作的蛋白。但由于酵母文库筛选方法假阳性率较高, 进一步运用Y2H确定TaPP2C5、TaSLY1、TaCHI、TaRbcS、TaSKP1与TaPP2-A13存在相互作用。Hong等[25]发现TaKFB1和TaKFB3经过酵母文库筛选分别获得了61个和74个克隆, 进一步验证确定TaKFB1与4个蛋白存在相互作用, TaKFB3与3个蛋白存在相互作用, 且这2个TaKFB蛋白都能与TaSKP1s蛋白相互作用。这说明1个F-box蛋白可识别多个底物, 且1个F-box蛋白可能与多个SKP1蛋白相结合。
SKP1是SCF (SKP1-CUL1-F-box protein)复合体的组分, 能够与F-box蛋白中的F-box结构域直接结合, 从而参与靶蛋白的泛素化过程。经预测, TaPP2-A13和TaSKP1均无信号肽结构, 且TaSKP1的亚细胞定位预测结果为细胞核, 这为二者在空间上互作提供了可能性。据报道, AtPP2-B11能与SCF复合体骨架蛋白SKP1家族成员ASK7、ASK18和ASK19互作, 暗示其具有F-box蛋白的功能。同时过表达牡丹(Paeonia suffruticosa) PSK1基因的拟南芥耐盐性得到显著提高[28], 同样的结论在大豆GmSK1过表达的烟草中得到了印证, 此外, 过表达GmSK1的烟草还表现出对干旱胁迫有较强的耐受性。本研究通过Y2H、BiFC和Co-IP实验证实TaPP2-A13与TaSKP1存在相互作用, 因此推测TaPP2-A13可能为SCF复合体的组分, 通过与TaSKP1结合实现其响应生物/非生物逆境过程的作用。
PP2C5是可以调控植物ABA信号通路的丝氨酸/苏氨酸蛋白磷酸酶, 该酶既影响植物的生长发育, 又在植物抵御非生物胁迫和抗病过程中发挥作用[30]; SLY1属于SEC1蛋白家族, 其亚细胞定位为高尔基体。酵母中的SLY1参与了内质网到高尔基体的囊泡运输过程[43]。目前, SEC1家族蛋白的研究主要集中在酵母、线虫(Limnodrilus hoffmeisteri)和大鼠(Rattus norvegicus)中[33], 其在植物中的研究鲜见报道; CHI在植物体中普遍存在, 病原真菌攻击、真菌细胞壁的刺激和乙烯等均能诱导植物体内几丁质酶的快速积累。此外, 几丁质酶也会响应重金属、渗透压和干旱等非生物逆境胁迫[44]。在活体外, 几丁质酶能高度抑制真菌的生长[31]。根据Melchers等[45]的分类方法, 本研究中获得的小麦TaCHI为I类几丁质酶。烟草中的I类几丁质酶定位于植物液泡中[46], 本研究中获得的TaCHI亚细胞定位预测也是液泡中。RbcS基因编码的蛋白质已被证明影响Rubisco的催化效率、组装、活性及其稳定性[47]。研究发现拟南芥中RbcS的表达存在组织特异性且受光的诱导[27]。本文通过Y2H技术证实TaPP2-A13分别与TaPP2C5、TaSLY1、TaCHI和TaRbcS存在相互作用, 该结果仍需其他蛋白互作技术深入验证。
4 结论
自小麦抗叶锈病近等基因系TcLr15中分离了1个小麦FBP亚家族基因TaPP2-A13; 该基因所编码蛋白在小麦近缘种属中的进化比较保守; 定位于细胞核和细胞质上; 受叶锈菌侵染、外源激素和非生物逆境处理后, TaPP2-A13基因表达量呈现上调或下调的显著变化。文库筛选获得11种与TaPP2-A13互作的蛋白, Y2H验证表明TaPP2C5、TaSLY1、TaCHI、TaRbcS、TaSKP1与TaPP2-A13存在物理相互作用, BiFC和Co-IP深入研究确定TaSKP1与TaPP2-A13存在相互作用。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/s0092-8674(00)80098-7URLPMID:8706131 [本文引用: 1]

We have identified the yeast and human homologs of the SKP1 gene as a suppressor of cdc4 mutants and as a cyclin F-binding protein. Skp1p indirectly binds cyclin A/Cdk2 through Skp2p, and directly binds Skp2p, cyclin F, and Cdc4p through a novel structural motif called the F-box. SKP1 is required for ubiquitin-mediated proteolysis of Cin2p, Clb5p, and the Cdk inhibitor Sic1p, and provides a link between these molecules and the proteolysis machinery. A large number of proteins contain the F-box motif and are thereby implicated in the ubiquitin pathway. Different skp1 mutants arrest cells in either G1 or G2, suggesting a connection between regulation of proteolysis in different stages of the cycle.
URLPMID:28934292 [本文引用: 1]
URLPMID:7780306 [本文引用: 1]
DOI:10.1038/nature03543URLPMID:15917797 [本文引用: 1]
The plant hormone auxin regulates diverse aspects of plant growth and development. Recent studies indicate that auxin acts by promoting the degradation of the Aux/IAA transcriptional repressors through the action of the ubiquitin protein ligase SCF(TIR1). The nature of the signalling cascade that leads to this effect is not known. However, recent studies indicate that the auxin receptor and other signalling components involved in this response are soluble factors. Using an in vitro pull-down assay, we demonstrate that the interaction between transport inhibitor response 1 (TIR1) and Aux/IAA proteins does not require stable modification of either protein. Instead auxin promotes the Aux/IAA-SCF(TIR1) interaction by binding directly to SCF(TIR1). We further show that the loss of TIR1 and three related F-box proteins eliminates saturable auxin binding in plant extracts. Finally, TIR1 synthesized in insect cells binds Aux/IAA proteins in an auxin-dependent manner. Together, these results indicate that TIR1 is an auxin receptor that mediates Aux/IAA degradation and auxin-regulated transcription.
URLPMID:17637677 [本文引用: 1]
[本文引用: 1]
URLPMID:25070642 [本文引用: 1]
DOI:10.1016/j.plaphy.2014.09.017URLPMID:25299612 [本文引用: 2]
Drought is one of the most important factors limiting plant growth and development. We identified a gene in wheat (Triticum aestivum L.) under drought stress named TaFBA1. TaFBA1 encodes a putative 325-amino-acid F-box protein with a conserved N-terminal F-box domain and a C-terminal AMN1 domain. Real-time RT-PCR analysis revealed that TaFBA1 transcript accumulation was upregulated by high-salinity, water stress, and abscisic acid (ABA) treatment. To evaluate the functions of TaFBA1 in the regulation of drought stress responses, we produced transgenic tobacco lines overexpressing TaFBA1. Under water stress conditions, the transgenic tobacco plants had a higher germination rate, higher relative water content, net photosynthesis rate (Pn), less chlorophyll loss, and less growth inhibition than WT. These results demonstrate the high tolerance of the transgenic plants to drought stress compared to the WT. The enhanced oxidative stress tolerance of these plants, which may be involved in their drought tolerance, was indicated by their lower levels of reactive oxygen species (ROS) accumulation, MDA content, and cell membrane damage under drought stress compared to WT. The antioxidant enzyme activities were higher in the transgenic plants than in WT, which may be related to the upregulated expression of some antioxidant genes via overexpression of TaFBA1.
DOI:10.1371/journal.pone.0122117URLPMID:25906259 [本文引用: 1]
As one of the largest gene families, F-box domain proteins have been found to play important roles in abiotic stress responses via the ubiquitin pathway. TaFBA1 encodes a homologous F-box protein contained in E3 ubiquitin ligases. In our previous study, we found that the overexpression of TaFBA1 enhanced drought tolerance in transgenic plants. To investigate the mechanisms involved, in this study, we investigated the tolerance of the transgenic plants to oxidative stress. Methyl viologen was used to induce oxidative stress conditions. Real-time PCR and western blot analysis revealed that TaFBA1 expression was up-regulated by oxidative stress treatments. Under oxidative stress conditions, the transgenic tobacco plants showed a higher germination rate, higher root length and less growth inhibition than wild type (WT). The enhanced oxidative stress tolerance of the transgenic plants was also indicated by lower reactive oxygen species (ROS) accumulation, malondialdehyde (MDA) content and cell membrane damage under oxidative stress compared with WT. Higher activities of antioxidant enzymes, including superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX) and peroxidase (POD), were observed in the transgenic plants than those in WT, which may be related to the upregulated expression of some antioxidant genes via the overexpression of TaFBA1. In others, some stress responsive elements were found in the promoter region of TaFBA1, and TaFBA1 was located in the nucleus, cytoplasm and plasma membrane. These results suggest that TaFBA1 plays an important role in the oxidative stress tolerance of plants. This is important for understanding the functions of F-box proteins in plants' tolerance to multiple stress conditions.
DOI:10.1016/j.plantsci.2017.03.010URLPMID:28483055 [本文引用: 1]
F-box protein is a major subunit of the Skp1-Cullin-F-box (SCF) complex. We previously isolated an F-box gene from wheat, TaFBA1, and here we show that overexpression of TaFBA1 in transgenic plants under salt stress increases germination rate, root elongation, and biomass accumulation compared with WT plants. Improvements in the photosynthetic rate and its corresponding parameters were also found in the transgenic plants. These results suggest that overexpression of TaFBA1 improves salt stress tolerance in transgenic tobacco. Further, the transgenic plants displayed less membrane damage, higher antioxidant enzyme activity, and less accumulation of ROS under salt stress. The transgenic plants also had lower Na(+) content and higher K(+) content than WT plants in leaves and roots. The activity of H(+)-ATPase on the plasma membrane in the transgenic plants was higher than in WT plants, and was accompanied by a net Na(+) efflux. In the tonoplast, the activity levels of V-ATPase and PPase were also higher in the transgenic plants, thus helping to maximize intracellular Na(+) compartmentalization. The expression of some stress-related genes was upregulated by salt stress. This suggests that the enhancement of plant salt stress tolerance may be associated with an improvement in antioxidative competition and Na(+)/K(+) ion regionalization.
DOI:10.3389/fpls.2018.00521URLPMID:29740462 [本文引用: 1]
Adverse environmental conditions, including high temperature, often affect the growth and production of crops worldwide. F-box protein, a core component of the Skp1-Cullin-F-box (SCF) E3 ligase complex, plays an important role in abiotic stress responses. A previously cloned gene from wheat, TaFBA1, encodes a homologous F-box protein. A Yeast two-Hybrid (Y2H) assay showed that TaFBA1 interacted with other SCF proteins. We found that the expression of TaFBA1 could be induced by heat stress (45 degrees C). Overexpression of TaFBA1 enhanced heat stress tolerance in transgenic tobacco, because growth inhibition was reduced and photosynthesis increased as compared with those in the wild type (WT) plants. Furthermore, the accumulation of H2O2, O2(-), and carbonyl protein decreased and cell damage was alleviated in transgenic plants under heat stress, which resulted in less oxidative damage. However, the transgenic plants contained more enzymatic antioxidants after heat stress, which might be related to the regulation of some antioxidant gene expressions. The qRT-PCR analysis showed that the overexpression of TaFBA1 upregulated the expression of genes involved in reactive oxygen species (ROS) scavenging, proline biosynthesis, and abiotic stress responses. We identified the interaction of TaFBA1 with Triticum aestivum stress responsive protein 1 (TaASRP1) by Y2H assay and bimolecular fluorescence complementation (BiFC) assay. The results suggested that TaFBA1 may improve enzymatic antioxidant levels and regulate gene expression by interacting with other proteins, such as TaASRP1, which leads to the enhanced heat stress tolerance seen in the transgenic plants.
DOI:10.3389/fpls.2016.01685URLPMID:27909441 [本文引用: 2]
MAX2 (MORE AXILLARY GROWTH2) is involved in diverse physiological processes, including photomorphogenesis, the abiotic stress response, as well as karrikin and strigolactone signaling-mediated shoot branching. In this study, MdMAX2, an F-box protein that is a homolog of Arabidopsis MAX2, was identified and characterized. Overexpression of MdMAX2 in apple calli enhanced the accumulation of anthocyanin. Ectopic expression of MdMAX2 in Arabidopsis exhibited photomorphogenesis phenotypes, including increased anthocyanin content and decreased hypocotyl length. Further study indicated that MdMAX2 might promote plant photomorphogenesis by affecting the auxin signaling as well as other plant hormones. Transcripts of MdMAX2 were noticeably up-regulated in response to NaCl and Mannitol treatments. Moreover, compared with the wild-type, the MdMAX2-overexpressing apple calli and Arabidopsis exhibited increased tolerance to salt and drought stresses. Taken together, these results suggest that MdMAX2 plays a positive regulatory role in plant photomorphogenesis and stress response.
DOI:10.1111/j.1365-313X.2009.03995.xURLPMID:19682297 [本文引用: 2]
Arabidopsis gain-of-resistance mutants, which show HR-like lesion formation and SAR-like constitutive defense responses, were used well as tools to unravel the plant defense mechanisms. We have identified a novel mutant, designated constitutive expresser of PR genes 30 (cpr30), that exhibited dwarf morphology, constitutive resistance to the bacterial pathogen Pseudomonas syringae and the dramatic induction of defense-response gene expression. The cpr30-conferred growth defect morphology and defense responses are dependent on ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1), PHYTOALEXIN DEFICIENT 4 (PAD4), and NONRACE-SPECIFIC DISEASE RESISTANCE 1 (NDR1). Further studies demonstrated that salicylic acid (SA) could partially account for the cpr30-conferred constitutive PR1 gene expression, but not for the growth defect, and that the cpr30-conferred defense responses were NPR1 independent. We observed a widespread expression of CPR30 throughout the plant, and a localization of CPR30-GFP fusion protein in the cytoplasm and nucleus. As an F-box protein, CPR30 could interact with multiple Arabidopsis-SKP1-like (ASK) proteins in vivo. Co-localization of CPR30 and ASK1 or ASK2 was observed in Arabidopsis protoplasts. Based on these results, we conclude that CPR30, a novel negative regulator, regulates both SA-dependent and SA-independent defense signaling, most likely through the ubiquitin-proteasome pathway in Arabidopsis.
DOI:10.1007/s11033-010-0438-yURLPMID:21104020 [本文引用: 3]
F-box proteins are key components of the ubiquitin (Ub)/26S proteasome pathway that mediates selective degradation of regulatory proteins involved in a wide variety of cellular processes affecting eukaryotic cells. In plants, F-box genes form one of the largest multigene superfamilies and control many important biological functions. Among the F-box genes characterized to date only few have been involved in the regulation of plant defense responses. Moreover, no F-box genes have been studied and characterized in grapevine. Using a differential display approach we isolated a F-box gene (BIG-24.1), which is up-regulated during Botrytis cinerea infection of grapevine leaves. BIG-24.1 encodes a polypeptide of 386 amino acids with a conserved F-box domain in the N-terminus region and a kelch domain. By investigating expression profiles of BIG-24.1, we show that the gene expression is strongly stimulated in B. cinerea infected berries and in grapevine cells challenged by MAMP rhamnolipids, a non-host bacterium and an endophytic rhizobacterium. The gene is also strongly induced by abiotic stresses including UV-C and wounding or by salicylic acid, methyl-jasmonate, ethylene and abscisic acid that are known to be involved in defense signalling pathways. In addition, sequence analysis of the BIG-24.1 promoter revealed the presence of several regulatory elements involved in the activation of plant defense responses.
DOI:10.1104/pp.013086URLPMID:12529520 [本文引用: 1]
Phloem protein 2 (PP2) is one of the most abundant and enigmatic proteins in the phloem sap. Although thought to be associated with structural P-protein, PP2 is translocated in the assimilate stream where its lectin activity or RNA-binding properties can exert effects over long distances. Analyzing the diversity of these proteins in vascular plants led to the identification of PP2-like genes in species from 17 angiosperm and gymnosperm genera. This wide distribution of PP2 genes in the plant kingdom indicates that they are ancient and common in vascular plants. Their presence in cereals and gymnosperms, both of which lack structural P-protein, also supports a wider role for these proteins. Within this superfamily, PP2 proteins have considerable size polymorphism. This is attributable to variability in the length of the amino terminus that extends from a highly conserved domain. The conserved PP2 domain was identified in the proteins encoded by six genes from several cucurbits, celery (Apium graveolens), and Arabidopsis that are specifically expressed in the sieve element-companion cell complex. The acquisition of additional modular domains in the amino-terminal extensions of other PP2-like proteins could reflect divergence from its phloem function.
[本文引用: 1]
DOI:10.1016/j.bbrc.2013.11.034URLPMID:24269669 [本文引用: 1]
Although several phloem sap proteins have been identified from protein extracts of heat-treated Arabidopsis seedlings using FPLC gel filtration columns, many of the physiological roles played by these proteins remain to be elucidated. We functionally characterized a phloem protein 2-A1, which encodes a protein similar to phloem lectin. Using a bacterially expressed recombinant protein of AtPP2-A1, we found that it performs dual functions, showing both molecular chaperone activity and antifungal activity. mRNA expression of the AtPP2-1 gene was induced by diverse external stresses such as pathogens, and other signaling molecules, such as ethylene. These results suggest that the AtPP2-A1 molecular chaperone protein plays a critical role in the Arabidopsis defense system against diverse external stresses including fungal pathogenic attack and heat shock.
DOI:10.1111/j.1432-1033.1983.tb07603.xURLPMID:6884347 [本文引用: 1]
The two major proteins from the phloem exudate of Cucurbita maxima (pumpkin), PP1 and PP2, were stable in the absence of reducing agents after modification of their accessible cysteine residues with iodoacetamide. This permitted their purification without precautions to prevent oxidation. PP2, a lectin specific for oligomers of N-acetyl-D-glucosamine, was shown by sedimentation-equilibrium ultracentrifugation to be a dimer of Mr of 48000. Neither dithiothreitol nor tri-(N-acetyl-D-glucosamine) altered this value. The constituent polypeptides were linked by two buried disulphide bridges. PP2 behaved aberrantly on gel-filtration on both Sephadex and Bio-Gel unless tri-(N-acetyl-D-glucosamine) was added to the elution buffer; the Mr was then measured as 46000. Other proteins which bind oligomers of N-acetyl-D-glucosamine are also retarded on gel-filtration. Soluble phloem filaments were prepared by collection of exudate into deaerated buffer containing iodoacetamide but no reducing agent. Oxidative gellation of the filaments was prevented by rapid modification of their many accessible cysteine residues, and is assumed to have maintained the degree of polymerisation found in vivo. Those disulphide bridges which were present allowed the incorporation of approximately 60% of the PP1 and 80% of the PP2 into polymeric material. It is concluded that PP1 and PP2 are both structural proteins present in the filaments observable in vivo. PP2 had an elongated binding-site for oligomers of N-acetyl-D-glucosamine. It is suggested that this lectin immobilises bacteria and fungi to the cross-linked filaments which seal wounded phloem sieve-tubes, and thus maintains sterility.
DOI:10.1007/s10658-018-01607-9URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41598-017-17765-5URLPMID:29311619 [本文引用: 1]
Zika virus (ZIKV) has recently caused a pandemic disease, and many cases of ZIKV infection in pregnant women resulted in abortion, stillbirth, deaths and congenital defects including microcephaly, which now has been proposed as ZIKV congenital syndrome. This study aimed to investigate the in situ immune response profile and mechanisms of neuronal cell damage in fatal Zika microcephaly cases. Brain tissue samples were collected from 15 cases, including 10 microcephalic ZIKV-positive neonates with fatal outcome and five neonatal control flavivirus-negative neonates that died due to other causes, but with preserved central nervous system (CNS) architecture. In microcephaly cases, the histopathological features of the tissue samples were characterized in three CNS areas (meninges, perivascular space, and parenchyma). The changes found were mainly calcification, necrosis, neuronophagy, gliosis, microglial nodules, and inflammatory infiltration of mononuclear cells. The in situ immune response against ZIKV in the CNS of newborns is complex. Despite the predominant expression of Th2 cytokines, other cytokines such as Th1, Th17, Treg, Th9, and Th22 are involved to a lesser extent, but are still likely to participate in the immunopathogenic mechanisms of neural disease in fatal cases of microcephaly caused by ZIKV.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11033-019-05210-xURLPMID:31907740 [本文引用: 2]
F-box proteins play important roles in the regulation of various developmental processes in plants. Approximately 1796 F-box genes have been identified in the wheat genome, but details of their functions remain unknown. Moreover, not much was known about the roles of kelch repeat domain-containing F-box genes (TaKFBs) in wheat. In the present study, we isolated five TaKFBs to investigate the roles of KFBs at different stages of colored wheat grain development. The cDNAs encoding TaKFB1, TaKFB2, TaKFB3, TaKFB4, and TaKFB5 contained 363, 449, 353, 382, and 456 bp open reading frames, respectively. All deduced TaKFBs contained an F-box domain (IPR001810) and a kelch repeat type 1 domain (IPR006652), except TaKFB2. Expression of TaKFBs was elevated during the pigmentation stages of grain development. To clarify how TaKFB and SKP interact in wheat, we investigated whether five TaKFB proteins showed specificity for six SKP proteins using a yeast two-hybrid (Y2H) assay. An Y2H screen was performed to search for proteins capable of binding the TaKFBs and interaction was identified between TaKFB1 and aquaporin PIP1. To examine the subcellular localization of TaKFBs, we transiently expressed TaKFB-green fluorescent protein (GFP) fusions in tobacco leaves; the TaKFB-GFP fusions were detected in the nucleus and the cytoplasm. Y2H and bimolecular fluorescence complementation (BiFC) assays revealed that TaKFB1 specifically interacts with aquaporin PIP1. These results will provide useful information for further functional studies on wheat F-box proteins and their possible roles.
DOI:10.1016/j.tplants.2005.05.002URLPMID:15951221 [本文引用: 1]
Iron-sulfur (Fe-S) clusters are ubiquitous prosthetic groups required to sustain fundamental life processes. The assembly of Fe-S clusters and insertion into polypeptides in vivo has recently become an area of intense research. Many of the genes involved are conserved in bacteria, fungi, animals and plants. Plant cells can carry out both photosynthesis and respiration - two processes that require significant amounts of Fe-S proteins. Recent findings now suggest that both plastids and mitochondria are capable of assembling Fe-S proteins using assembly machineries that differ in biochemical properties, genetic make-up and evolutionary origin.
URLPMID:16453702 [本文引用: 2]
We have determined the nucleotide sequence of two members (rbcS-3A and -3C) of the pea nuclear gene family encoding the small subunit (rbcS) of ribulose-1,5-bisphosphate carboxylase. Both rbcS-3A and -3C are interrupted by two introns located at the same positions as those of the other three pea rbcS genes. Compared with the other pea rbcS genes the rbcS-3C gene has the most divergent 5'- and 3'-flanking sequences while the rbcS-3A gene has a larger and highly divergent intron 1. All five pea rbcS genes are conserved in their coding regions but show considerable sequence differences in their 3'-untranslated portion. The 3' sequence divergence of the rbcS genes has allowed us to use S1 nuclease mapping procedures to compare their expression levels in different organs and during light induction. All the rbcS genes are differentially expressed in various organs of the pea plants; moreover, specific rbcS transcripts are under-represented in seeds and petals. In leaves there is a 10-fold difference between the highest and lowest specific rbcS transcript levels. By quantitating the distribution of rbcS transcripts during light, phytochrome and blue light induction of immature (etiolated), and mature (green), pea leaves, we show that the genes are differentially activated during leaf development.
URLPMID:27787596 [本文引用: 2]
DOI:10.1105/tpc.010436URLPMID:12034899 [本文引用: 1]
Plant UDP-Glc:phenylpropanoid glucosyltransferases (UGTs) catalyze the transfer of Glc from UDP-Glc to numerous substrates and regulate the activity of compounds that play important roles in plant defense against pathogens. We previously characterized two tobacco salicylic acid- and pathogen-inducible UGTs (TOGTs) that act very efficiently on the hydroxycoumarin scopoletin and on hydroxycinnamic acids. To identify the physiological roles of these UGTs in plant defense, we generated TOGT-depleted tobacco plants by antisense expression. After inoculation with Tobacco mosaic virus (TMV), TOGT-inhibited plants exhibited a significant decrease in the glucoside form of scopoletin (scopolin) and a decrease in scopoletin UGT activity. Unexpectedly, free scopoletin levels also were reduced in TOGT antisense lines. Scopolin and scopoletin reduction in TOGT-depleted lines resulted in a strong decrease of the blue fluorescence in cells surrounding TMV lesions and was associated with weakened resistance to infection with TMV. Consistent with the proposed role of scopoletin as a reactive oxygen intermediate (ROI) scavenger, TMV also triggered a more sustained ROI accumulation in TOGT-downregulated lines. Our results demonstrate the involvement of TOGT in scopoletin glucosylation in planta and provide evidence of the crucial role of a UGT in plant defense responses. We propose that TOGT-mediated glucosylation is required for scopoletin accumulation in cells surrounding TMV lesions, where this compound could both exert a direct antiviral effect and participate in ROI buffering.
DOI:10.1104/pp.103.025569URLPMID:12970481 [本文引用: 2]
FsPP2C1 was previously isolated from beech (Fagus sylvatica) seeds as a functional protein phosphatase type-2C (PP2C) with all the conserved features of these enzymes and high homology to ABI1, ABI2, and PP2CA, PP2Cs identified as negative regulators of ABA signaling. The expression of FsPP2C1 was induced upon abscisic acid (ABA) treatment and was also up-regulated during early weeks of stratification. Furthermore, this gene was specifically expressed in ABA-treated seeds and was hardly detectable in vegetative tissues. In this report, to provide genetic evidence on FsPP2C1 function in seed dormancy and germination, we used an overexpression approach in Arabidopsis because transgenic work is not feasible in beech. Constitutive expression of FsPP2C1 under the cauliflower mosaic virus 35S promoter confers ABA insensitivity in Arabidopsis seeds and, consequently, a reduced degree of seed dormancy. Additionally, transgenic 35S:FsPP2C1 plants are able to germinate under unfavorable conditions, as inhibitory concentrations of mannitol, NaCl, or paclobutrazol. In vegetative tissues, Arabidopsis FsPP2C1 transgenic plants show ABA-resistant early root growth and diminished induction of the ABA-response genes RAB18 and KIN2, but no effect on stomatal closure regulation. Seed and vegetative phenotypes of Arabidopsis 35S:FsPP2C1 plants suggest that FsPP2C1 negatively regulates ABA signaling. The ABA inducibility of FsPP2C1 expression, together with the transcript accumulation mainly in seeds, suggest that it could play an important role modulating ABA signaling in beechnuts through a negative feedback loop. Finally, we suggest that negative regulation of ABA signaling by FsPP2C1 is a factor contributing to promote the transition from seed dormancy to germination during early weeks of stratification.
URLPMID:24263941 [本文引用: 2]
DOI:10.1093/pcp/pcr008URLPMID:21257607 [本文引用: 1]
The phytohormone ABA plays an important role in plant development and adaptation to diverse environmental stresses. Many of the components involved in ABA signaling remain to be discovered, and knowledge of these is needed to understand fully the highly complex ABA signaling network. Here, we report that an enzyme catalyzing beta-oxidation of fatty acids, 3-ketoacyl-CoA thiolase-2 (KAT2/PED1/PKT3) (EC 2.3.1.16), is involved in ABA signaling. We provide genetic evidence that KAT2 positively regulates ABA signaling in all the major ABA responses, including ABA-induced inhibition of seed germination and post-germination growth arrest, and ABA-induced stomatal closure and stomatal opening inhibition in Arabidopsis thaliana. KAT2 was shown to be important for reactive oxygen species (ROS) production in response to ABA, suggesting that KAT2 regulates ABA signaling at least partly through modulating ROS homeostasis in plant cells. Additionally, we provide data suggesting that KAT2 may function downstream of an important WRKY transcription repressor WRKY40, which may link KAT2 with the ABA receptor ABAR/CHLH-mediated signaling.
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.7668/hbnxb.2016.03.002URL [本文引用: 1]
V/V)EMS处理成熟种子12h效果最佳,筛选获得一个亚麻荠2SP1突变体。与野生型相比,该突变体种子贮藏蛋白成分中缺失了2S清蛋白,但是亚麻荠种子含油量和蛋白含量、株型和种子萌发等农艺性状未发生显著变化。这种特殊的亚麻荠2SP1突变体可作为研究种子贮藏蛋白合成机理及培育高营养品质亚麻荠新品种的优异种质资源,也可用于进行相关功能基因组学等的研究。]]>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.plantsci.2019.04.013URLPMID:31084865 [本文引用: 1]
Avr9/Cf-9-INDUCED F-BOX1 (ACIF1) was first identified during screening of Avr9/Cf-9-elicited genes in tobacco. Further analysis revealed that ACIF1 was required for hypersensitive responses triggered by various elicitors in tobacco and tomato, indicating that it may be involved in various disease resistance. Here, we cloned its cotton (Gossypium hirsutum) homolog GhACIF1, which encodes an F-box protein. We show that GhACIF1 interacts with the putative SKP1-like protein, named GhSKP1. Disease resistance assays show that GhACIF1 enhances resistance to Verticillium dahliae in Arabidopsis plants, while silencing of GhACIF1 confers sensitivity to V. dahliae in cotton. Further analysis show that PevD1 elicitor activates hypersensitive and acquired immune response mediated by GhACIF1. Collectively, these results indicate that GhACIF1 contributes to protection against V. dahliae infection.
DOI:10.1105/tpc.001867URLPMID:12119368 [本文引用: 1]
One of several induced defense responses in plants is systemic acquired resistance (SAR), which is regulated by salicylic acid and in Arabidopsis by the NIM1/NPR1 protein. To identify additional components of the SAR pathway or other genes that regulate SAR-independent resistance, we performed genetic suppressor screens of mutagenized nim1-1 seedlings, which are highly susceptible to infection by Peronospora parasitica. We isolated the son1 (suppressor of nim1-1) mutant, which shows full restoration of pathogen resistance without the induction of SAR-associated genes and expresses resistance when combined with a salicylate hydroxylase (nahG) transgene. These features indicate that son1-mediated resistance is distinct from SAR. Resistance is effective against both the virulent oomycete Peronospora and the bacterial pathogen Pseudomonas syringae pv tomato strain DC3000. We cloned SON1 and found it to encode a novel protein containing an F-box motif, an element found within the specificity determinant in the E3 ubiquitin-ligase complex. We propose the existence of a novel defense response that is independent of SAR and negatively regulated in Arabidopsis by SON1 through the ubiquitin-proteosome pathway.
DOI:10.1007/s00344-018-9863-2URL [本文引用: 1]
DOI:10.3389/fpls.2017.02218URLPMID:29375596 [本文引用: 1]
Plants are constantly exposed to a wide range of environmental stresses, but evolved complicated adaptive and defense mechanisms which allow them to survive in unfavorable conditions. These mechanisms protect and defend plants by using different immune receptors located either at the cell surface or in the cytoplasmic compartment. Lectins or carbohydrate-binding proteins are widespread in the plant kingdom and constitute an important part of these immune receptors. In the past years, lectin research has focused on the stress-inducible lectins. The Nicotiana tabacum agglutinin, abbreviated as Nictaba, served as a model for one family of stress-related lectins. Here we focus on three non-chimeric Nictaba homologs from Arabidopsis thaliana, referred to as AN3, AN4, and AN5. Confocal microscopy of ArathNictaba enhanced green fluorescent protein (EGFP) fusion constructs transiently expressed in N. benthamiana or stably expressed in A. thaliana yielded fluorescence for AN4 and AN5 in the nucleus and the cytoplasm of the plant cell, while fluorescence for AN3 was only detected in the cytoplasm. RT-qPCR analysis revealed low expression for all three ArathNictabas in different tissues throughout plant development. Stress application altered the expression levels, but all three ArathNictabas showed a different expression pattern. Pseudomonas syringae infection experiments with AN4 and AN5 overexpression lines demonstrated a significantly higher tolerance of several transgenic lines to P. syringae compared to wild type plants. Finally, AN4 was shown to interact with two enzymes involved in plant defense, namely TGG1 and BGLU23. Taken together, our data suggest that the ArathNictabas represent stress-regulated proteins with a possible role in plant stress responses. On the long term this research can contribute to the development of more stress-resistant plants.
DOI:10.1016/j.gene.2006.11.016URLPMID:17240087 [本文引用: 1]
In eukaryotes, E3 ubiquitin ligases (E3s) mediate the ubiquitylation of proteins that are destined for degradation by the ubiquitin-proteasome system. In SKP1/CDC53/F-box protein (SCF)-type E3 complexes, the interchangeable F-box protein confers specificity to the E3 ligase through direct physical interactions with the degradation substrate. The vast majority of the approximately 700 F-box proteins from the plant model organism Arabidopsis thaliana remain to be characterized. Here, we investigate the previously uncharacterized and evolutionarily conserved Arabidopsis F-box protein 7 (AtFBP7), which is encoded by a unique gene in Arabidopsis (At1g21760). Several apparent fbp7 loss-of-function alleles do not have an obvious phenotype. AtFBP7 is ubiquitously expressed and its expression is induced after cold and heat stress. When following up on a reported co-purification of the eukaryotic elongation factor-2 (eEF-2) with YLR097c, the apparent budding yeast orthologue of AtFBP7, we discovered a general defect in protein biosynthesis after cold and heat stress in fbp7 mutants. Thus, our findings suggest that AtFBP7 is required for protein synthesis during temperature stress.
DOI:10.1104/pp.108.126912URLPMID:18835996 [本文引用: 1]
Guard cells, which form stoma in leaf epidermis, sense and integrate environmental signals to modulate stomatal aperture in response to diverse conditions. Under drought stress, plants synthesize abscisic acid (ABA), which in turn induces a rapid closing of stoma, to prevent water loss by transpiration. However, many aspects of the molecular mechanism for ABA-mediated stomatal closure are still not understood. Here, we report a novel negative regulator of guard cell ABA signaling, DOR, in Arabidopsis (Arabidopsis thaliana). The DOR gene encodes a putative F-box protein, a member of the S-locus F-box-like family related to AhSLF-S(2) and specifically interacting with ASK14 and CUL1. A null mutation in DOR resulted in a hypersensitive ABA response of stomatal closing and a substantial increase of drought tolerance; in contrast, the transgenic plants overexpressing DOR were more susceptible to the drought stress. DOR is strongly expressed in guard cells and suppressed by ABA treatment, suggesting a negative feedback loop of DOR in ABA responses. Double-mutant analyses of dor with ABA-insensitive mutant abi1-1 showed that abi1-1 is epistatic to dor, but no apparent change of phospholipase Dalpha1 was detected between the wild type and dor. Affymetrix GeneChip analysis showed that DOR likely regulates ABA biosynthesis under drought stress. Taken together, our results demonstrate that DOR acts independent of phospholipase Dalpha1 in an ABA signaling pathway to inhibit the ABA-induced stomatal closure under drought stress.
DOI:10.1016/0378-1119(95)00819-5URLPMID:8647468 [本文引用: 1]
We characterized rat cDNAs that predict a protein, r-Sly1, which is similar to SLY1, a yeast protein that plays a critical role in endoplasmic reticulum to Golgi apparatus vesicle trafficking. The r-Sly1 gene is expressed in all tissues examined.
[本文引用: 1]
[本文引用: 1]
URLPMID:8012401 [本文引用: 1]
DOI:10.1104/pp.101.3.857URLPMID:12231736 [本文引用: 1]
Different isoforms of chitinases and [beta]-1,3-glucanases of tobacco (Nicotiana tabacum cv Samsun NN) were tested for their antifungal activities. The class I, vacuolar chitinase and [beta]-1,3-glucanase isoforms were the most active against Fusarium solani germlings, resulting in lysis of the hyphal tips and in growth inhibition. In additon, we observed that the class I chitinase and [beta]-1,3-glucanase acted synergistically. The class II isoforms of the two hydrolases exhibited no antifungal activity. However, the class II chitinases showed limited growth inhibitory activity in combination with higher amounts of class I [beta]-1,3-glucanase. The class II [beta]-1,3-glucanases showed no inhibitory activity in any combination. In transgenic tobacco plants producing modified forms of either a class I chitinase or a class I [beta]-1,3-glucanase, or both, these proteins were targeted extracellularly. Both modified proteins lack their C-terminal propeptide, which functions as a vacuolar targeting signal. Extracellular targeting had no effect on the specific activities of the chitinase and [beta]-1,3-glucanase enzymes. Furthermore, the extracellular washing fluid (EF) from leaves of transgenic plants expressing either of the secreted class I enzymes exhibited antifungal activity on F. solani germlings in vitro comparable to that of the purified vacuolar class I proteins. Mixing EF fractions from these plants revealed synergism in inhibitory activity against F. solani; the mixed fractions exhibited inhibitory activity similar to that of EF from plants expressing both secreted enzymes.
URLPMID:31498865 [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}