 ,**, 申树林**, 黄小虎, 马国强, 王曦彤, 杨怡玲, 李洹东, 王书贤, 朱美晨, 唐章林, 卢坤, 李加纳,*, 曲存民,*西南大学农学与生物科技学院 / 油菜工程研究中心 / 西南大学现代农业科学研究院, 重庆 400715
,**, 申树林**, 黄小虎, 马国强, 王曦彤, 杨怡玲, 李洹东, 王书贤, 朱美晨, 唐章林, 卢坤, 李加纳,*, 曲存民,*西南大学农学与生物科技学院 / 油菜工程研究中心 / 西南大学现代农业科学研究院, 重庆 400715Genome-wide association analysis reveals zinc-tolerant loci of rapeseed at germination stage
WEI Li-Juan,**, SHEN Shu-Lin**, HUANG Xiao-Hu, MA Guo-Qiang, WANG Xi-Tong, YANG Yi-Ling, LI Huan-Dong, WANG Shu-Xian, ZHU Mei-Chen, TANG Zhang-Lin, LU Kun, LI Jia-Na,*, QU Cun-Min,*College of Agronomy and Biotechnology, Southwest University / Chongqing Engineering Research Center for Rapeseed / Academy of Agricultural Sciences, Southwest University, Chongqing 400715, China通讯作者:
收稿日期:2020-02-18接受日期:2020-04-15网络出版日期:2020-05-08
| 基金资助: |
Received:2020-02-18Accepted:2020-04-15Online:2020-05-08
| Fund supported: |
作者简介 About authors
魏丽娟, E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1282KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
魏丽娟, 申树林, 黄小虎, 马国强, 王曦彤, 杨怡玲, 李洹东, 王书贤, 朱美晨, 唐章林, 卢坤, 李加纳, 曲存民. 锌胁迫下甘蓝型油菜发芽期下胚轴长的全基因组关联分析[J]. 作物学报, 2021, 47(2): 262-274. doi:10.3724/SP.J.1006.2021.04037
WEI Li-Juan, SHEN Shu-Lin, HUANG Xiao-Hu, MA Guo-Qiang, WANG Xi-Tong, YANG Yi-Ling, LI Huan-Dong, WANG Shu-Xian, ZHU Mei-Chen, TANG Zhang-Lin, LU Kun, LI Jia-Na, QU Cun-Min.
近年来, 土壤重金属污染问题愈发严重, 对人类身体健康和生态系统产生重大影响[1]。锌是植物生长发育所必需的微量元素之一, 适量锌可促进植物生长发育, 增强植物对不良环境的适应性[2,3]。然而过度使用不仅会造成严重的环境污染, 还会降低土壤微生物的活性, 减缓土壤有机质的分解, 进而影响植物根的生长和光合作用[4,5,6]。油菜被认为是修复重金属(如Cu、Cd、Zn)污染土壤的理想植物之一[7,8,9,10], 对锌具有高耐受性, 在锌胁迫下仍可获得较高的生物产量[7,11]。曹春信等[12]通过研究油菜地上部器官对锌富集的特征发现, 锌在茎秆中的含量大于籽粒。挖掘耐锌性关键位点和候选基因对耐锌性油菜品种的培育具有重要意义, 全基因组关联分析被广泛应用于作物重金属抗性位点的检测。Zhang等[13]利用全基因组关联分析方法, 在水稻中共检测到31个耐锌QTL位点。Chen等[14]利用Brassica Illumina 60K SNP芯片进行全基因组关联分析, 检测出25个甘蓝型油菜耐镉QTL位点, 并找到NRAMP6、IRT1、CAD1和PCS2四个候选基因。Zhang等[15]检测到4个甘蓝型油菜耐镉位点, 并鉴定出一些候选基因, 如HIPP27、EXPB4、EMB1793和CDSP3。曲存民等[16]检测到70个甘蓝型油菜耐砷性的显著关联位点, 并筛选出PHT3;3和PHT1;9等与甘蓝型油菜砷离子吸收转运相关的候选基因。超富集植物对重金属的积累和耐性机制研究也取得了一定进展, 许多蛋白参与锌的吸收、转运和贮存, 包括CDF (Cation Diffusion Facilitator)蛋白家族[17]、NRAMP蛋白家族[18]、ZIP蛋白家族[19]、HMA蛋白家族(P-type ATPases)[20]、YSL (yellow stripe-like)蛋白家族[21]、ABC (ATP-binding cassette)转运蛋白家族[2,22]。但关于甘蓝型油菜耐锌性QTL定位和候选基因鉴定并未见相关报道。
为挖掘甘蓝型油菜与耐锌性状相关联的关键位点及候选基因, 本研究采用GWAS方法获得与下胚轴长显著关联的SNP标记, 结合转录组数据获得锌胁迫前后差异表达基因, 进一步确定影响下胚轴长的关键候选基因。本研究不仅为甘蓝型油菜耐锌性种质资源的鉴定提供分子标记, 而且可以为耐锌性油菜资源利用和新品种培育提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为国内外收集的甘蓝型油菜自交系品种140份, 其中国内材料123份, 主要来自重庆、四川、湖北、湖南等地, 其余17份材料来自加拿大和德国等国家, 所有材料由西南大学重庆市油菜工程技术研究中心收集并提供。1.2 试验处理与性状分析
从140份研究材料中随机抽取10份材料(附表1), 分别用0、5、10、15、20、30和50 mg L-1浓度的Zn(NO3)2进行处理, 比较分析发现, 处理浓度为30 mg L-1时材料间长势差异较大, 确定为最佳处理浓度(表1)。试验分为对照组和处理组, 重复3次, 挑选大小均匀、饱满的种子各100粒, 分别播种于对照纯水和30 mg L-1的Zn(NO3)2溶液处理的培养盘 (长37.0 cm × 宽24.5 cm × 高6.5 cm, 体积为4.5 L), 覆盖保鲜膜。生长条件为昼夜温度为25℃, 光照/黑暗时间为16 h/8 h, 光照强度为100 μmol m-2 s-1, 相对湿度为60%[23]。播种7 d后, 选取长势一致的5株幼苗, 分别测量对照和锌胁迫下的下胚轴长(hypocotyl length, HL)。将处理和对照的相对值作为衡量锌胁迫对甘蓝型油菜生长的影响[24], 记为相对下胚轴长(relative hypocotyl length, RHL)。计算公式: 相对性状值(%)=处理组测定值/对照组测定值×100%[16]。利用Microsoft Excel 2013绘制正态分布图, 利用SPSS统计分析软件进行偏度和峰度分析。Table S1
附表1
附表1140份甘蓝型油菜材料信息
Table S1
| 编号 Number | 材料名称 Accessions | 生态型 Ecotype | 来源 Source | ||
|---|---|---|---|---|---|
| 1 | 中双3号Zhongshuang 3 | 半冬性Semi-winter | 中国浙江Zhejiang, China | ||
| 2 | 镇油5号Zhenyou 5 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 3 | SWU46 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 4 | WH-127 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 5 | WH-41 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 6 | WX10329 | 半冬性Semi-winter | 中国湖南Hunan, China | ||
| 7 | JY-16 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 8 | 黔油331 Qianyou 331 | 半冬性Semi-winter | 中国贵州Guizhou, China | ||
| 编号 Number | 材料名称 Accessions | 生态型 Ecotype | 来源 Source | ||
| 9 | SWU47 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 10 | 浙双72 Zheshuang 72 | 半冬性Semi-winter | 中国浙江Zhejiang, China | ||
| 11 | 沪油17 Huyou 17 | 半冬性Semi-winter | 中国上海Shanghai, China | ||
| 12 | 中双7号Zhongshuang 7 | 半冬性Semi-winter | 中国武汉Wuhan, China | ||
| 13 | WH-26 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 14 | Taisetsu | 冬性Winter | 日本Japan | ||
| 15 | M114 | 半冬性 Semi-winter | 中国China | ||
| 16 | 中油589 Zhongyou 589 | 半冬性Semi-winter | 中国武汉Wuhan, China | ||
| 17 | 农林43 Nonglin 43 | 冬性Winter | 日本Japan | ||
| 18 | 广德8104 Guangde 8104 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 19 | 至尊Zhizun | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 20 | 中双2号Zhongshuang 2 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 21 | 中双4号Zhongshuang 4 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 22 | 华油3号Huayou 3 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 23 | 宁油14 Ningyou 14 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 24 | 华油12 Huayou 12 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 25 | 湖北白花油菜Hubei white flower rape | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 26 | WH-58 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 27 | 699 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 28 | SWU63 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 29 | WH-57 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 30 | 苏油1号Shuyou 1 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 31 | WH-19 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 32 | 中双4号Zhongshuang 4 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 33 | Nakate Chousen | 春性Spring | 朝鲜DPRK | ||
| 34 | 广德761 Gaungde 761 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 35 | SWU110 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 36 | 7094 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 37 | SWU71 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 38 | 1111 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 39 | Erake | 半冬性Semi-winter | 波兰Poland | ||
| 40 | Campino | 春性Spring | 德国Germany | ||
| 41 | 宁油10号 Ningyou 10 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 42 | 中双11DH Zhongshuang 11DH | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 43 | 浙双8号 Zheshuang 8 | 半冬性Semi-winter | 中国浙江Zhejiang, China | ||
| 44 | cresor | 春性Spring | 法国France | ||
| 45 | 浙油17 Zheyou 17 | 半冬性Semi-winter | 中国浙江Zhejiang, China | ||
| 46 | 华航901 Huahang 901 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 47 | Wesreo | 春性Spring | 澳大利亚Australia | ||
| 48 | 南川长角 Nanchuansiliqua | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 49 | J-917 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 50 | 浙双6号 Zheshuang 6 | 半冬性Semi-winter | 中国浙江Zhejiang, China | ||
| 51 | 荣选 Rongxuan | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 52 | SWU106 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 编号 Number | 材料名称 Accessions | 生态型 Ecotype | 来源 Source | ||
| 53 | D2 | 春性Spring | 丹麦Denmark | ||
| 54 | 豫油1号 Yuyou 1 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 55 | WH-37 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 56 | J-951 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 57 | 全紫油菜 Quanzi rape | 春性Spring | 中国江苏Jiangsu, China | ||
| 58 | SWU69 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 59 | SWU60 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 60 | 丰油9号 Fengyou 9 | 半冬性Semi-winter | 中国河南Henan, China | ||
| 61 | 宁油1号 Ningyou 1 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 62 | SWU80 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 63 | 10-C29 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 64 | 秦油5号 Qinyou 5 | 半冬性Semi-winter | 中国陕西Shaanxi, China | ||
| 65 | Sophia | 春性Spring | 德国Germany | ||
| 66 | WaseChousen | 春性Spring | 朝鲜DPRK | ||
| 67 | Korina | 春性Spring | 德国Germany | ||
| 68 | J-915 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 69 | Bienvenu | 半冬性Semi-winter | 法国France | ||
| 70 | SWU102 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 71 | 漕泾胜利 Caojingshengli | 半冬性Semi-winter | 中国上海Shanghai, China | ||
| 72 | 563 | 半冬性Semi-winter | 中国湖南Hunan, China | ||
| 73 | 皖油15号 Wanyou 15 | 半冬性Semi-winter | 中国安徽Anhui, China | ||
| 74 | 阳光198 Yangguang 198 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 75 | 湘油13号 Xiangyou 13 | 半冬性Semi-winter | 中国湖南Hunan, China | ||
| 76 | 皖油16号 Wanyou 16 | 半冬性Semi-winter | 中国安徽Anhui, China | ||
| 77 | 华油13号 Huayou 13 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 78 | 湘油15号 Xiangyou 15 | 半冬性Semi-winter | 中国湖南Hunan, China | ||
| 79 | Tapidor | 冬性Winter | 法国France | ||
| 80 | 10-C24 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 81 | 圣光77 Shengguang 77 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 82 | 史力丰 Shilifeng | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 83 | 沪油3号 Huyou 3 | 半冬性Semi-winter | 中国上海Shanghai, China | ||
| 84 | Santana | 半冬性Semi-winter | 中国China | ||
| 85 | 10-C34 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 86 | 11-Y7-117 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 87 | SWU99 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 88 | 宁油12 Ningyou 12 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 89 | Comet | 春性Spring | 瑞典Sweden | ||
| 90 | WH-20 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 91 | 扬J6711 Yang J6711 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 92 | 7191 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 93 | 中双9号 Zhongshuang 9 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 94 | Daechosen | 半冬性Semi-winter | 朝鲜DPRK | ||
| 95 | Monty | 半冬性Semi-winter | 澳大利亚Australia | ||
| 96 | 纬隆88 Weilong 88 | 半冬性Semi-winter | 中国陕西Shanxi, China | ||
| 编号 Number | 材料名称 Accessions | 生态型 Ecotype | 来源 Source | ||
| 97 | 沪油14 Huyou 14 | 半冬性Semi-winter | 中国上海Shanghai, China | ||
| 98 | A117 | 半冬性Semi-winter | 中国陕西Shaanxi, China | ||
| 99 | WH-15 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 100 | 2011-7103 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 101 | SWU68 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 102 | 浙油18 Zheyou 18 | 半冬性Semi-winter | 中国四川Zhejiang, China | ||
| 103 | Yan 81-2 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 104 | 2012-K8053 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 105 | 淮油6号 Huaiyou 6 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 106 | 2012-3546 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 107 | wx 10296 | 半冬性Semi-winter | 中国湖南Hunan, China | ||
| 108 | 2012-8380 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 109 | SWU92 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 110 | 农林42 Nonglin 42 | 半冬性Semi-winter | 日本Japan | ||
| 111 | 大花球 Dahuaqiu | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 112 | 华油6号 Huayou 6 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 113 | CY12Q95406 | 半冬性Semi-winter | 中国四川Sichuan, China | ||
| 114 | SWU49 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 115 | SWU70 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 116 | 华双128 Huashuang 128 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 117 | 皖油早 Wanyouzao | 半冬性Semi-winter | 中国安徽Anhui, China | ||
| 118 | 宁油12 Ningyou 12 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 119 | 滁油1号 Chuyou 1 | 半冬性Semi-winter | 中国安徽Anhui, China | ||
| 120 | 10-JP3 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 121 | 2359 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 122 | 宁油8号 Ningyou 8 | 半冬性Semi-winter | 中国江苏Jiangsu, China | ||
| 123 | CY21PXW-84 | 半冬性Semi-winter | 中国四川Sichuan, China | ||
| 124 | SWU88 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 125 | Sida | 春性Spring | 加拿大Canada | ||
| 126 | 川油20 Chuanyou 20 | 半冬性Semi-winter | 中国四川Sichuan, China | ||
| 127 | 沪油19 Huyou 19 | 半冬性Semi-winter | 中国上海Shanghai, China | ||
| 128 | WH-62 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 129 | 华油14 Huayou 14 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 130 | CY12GJ-1 | 半冬性Semi-winter | 中国四川Sichuan, China | ||
| 131 | 湘油11号 Xiangyou 11 | 半冬性Semi-winter | 中国湖南Hunan, China | ||
| 132 | 浙油601 Zheyou 601 | 半冬性Semi-winter | 中国浙江Zhejiang, China | ||
| 133 | SWU108 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 134 | CY14PXW-18 | 半冬性Semi-winter | 中国四川Sichuan, China | ||
| 135 | Nca | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 136 | 甘油5号 Ganyou 5 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 137 | SWU94 | 半冬性Semi-winter | 中国重庆Chongqing, China | ||
| 138 | 中油821 Zhongyou 821 | 半冬性Semi-winter | 中国湖北Hubei, China | ||
| 139 | 川油18 Chuanyou 18 | 半冬性Semi-winter | 中国四川Sichuan, China | ||
| 140 | wx 10213 | 半冬性Semi-winter | 中国湖北Hunan, China | ||
新窗口打开|下载CSV
Table 1
表1
表1不同Zn(NO3)2溶液浓度下油菜下胚轴长度
Table 1
| 品种 Accessions | Zn(NO3)2溶液浓度 Concentration of Zn(NO3)2 solutions (mg L-1) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 20 | 30 | 50 | 70 | |
| 1 | 5.76 | 4.46 | 4.74 | 5.82 | 3.78 | 2.54 | 2.54 | 1.68 |
| 2 | 4.38 | 4.38 | 4.02 | 3.16 | 2.38 | 2.54 | 2.52 | 2.08 |
| 3 | 6.80 | 6.64 | 5.02 | 6.02 | 4.34 | 2.62 | 2.24 | 2.34 |
| 4 | 7.60 | 7.66 | 6.64 | 5.20 | 4.20 | 3.14 | 2.76 | 2.14 |
| 5 | 6.70 | 6.28 | 5.24 | 4.32 | 3.24 | 3.20 | 2.84 | 1.80 |
| 6 | 6.25 | 5.88 | 5.13 | 4.90 | 3.59 | 2.81 | 2.58 | 2.01 |
| 7 | 7.90 | 6.30 | 8.40 | 6.52 | 5.88 | 5.46 | 4.84 | 2.60 |
| 8 | 5.90 | 7.98 | 8.64 | 4.50 | 4.08 | 4.58 | 4.64 | 2.40 |
| 9 | 6.30 | 7.90 | 7.10 | 5.60 | 6.54 | 4.50 | 3.74 | 2.40 |
| 10 | 6.96 | 7.08 | 8.06 | 5.73 | 6.01 | 4.86 | 4.94 | 2.91 |
| 平均值 Mean (cm) | 6.45 | 6.46 | 6.30 | 3.78 | 3.58 | 2.48 | 3.36 | 2.24 |
| 标准差SD (cm) | 1.00 | 1.29 | 1.68 | 1.17 | 1.25 | 0.87 | 1.07 | 0.37 |
| 变异系数 CV (%) | 0.155 | 0.200 | 0.267 | 0.309 | 0.349 | 0.352 | 0.319 | 0.166 |
新窗口打开|下载CSV
1.3 基因型分析及SNP标记定位
利用芸薹属60K SNP芯片, 参照Qu等[25,26]方法, 对140份材料进行基因型分析, 去掉最小基因型频率(minor allele frequency, MAF)低于0.05和SNP得率(call frequency)小于80%的标记。为确定SNP在染色体上的位置, 利用探针序列与基因组进行BLASTN比对, E-value设定为1e-5。去掉Bit-Score相同的SNP, 最终获得30,288个SNP标记用于群体性状的关联分析。1.4 群体结构分析与亲缘关系
利用STRUCTURE软件进行甘蓝型油菜群体结构分析[27], 组群数目(K)设定为1~10, K值运行5次(number of iterations), 将MCMC (Markov Chain Monte Carlo)不作数迭代(length of burn-in period)和不作数迭代后的MCMC全部设为100,000次, 在混合模型下进行运算。最后用运算得到的后验概率值和2个连续的后验概率值的变化速率(ΔK)来确定群体中存在的亚群数目[28]。利用SPAGeDi v1.4软件进行亲缘关系(relative kinship)分析, 并计算亲缘关系的矩阵[29]。1.5 全基因组关联分析及候选基因鉴定
利用TASSEL 5.0软件[30]中的一般线性模型(General Linear Model, GLM)和混合线性模型(Mixed Linear Model, MLM), 结合SNP芯片基因型数据, 以Q值和亲缘关系K值作为协变量, 进行全基因组关联分析, 确定关联位点和标记对表型变异的解释率(R2)。本研究采用了6种模型进行关联分析, 包括GLM中的naive、Q和PCA模型, MLM中的K、Q+K和PCA+K模型。根据每个SNP位点的-log10(P)观察值和期望值, 利用R软件(1.6 甘蓝型油菜锌胁迫转录组测序
取甘蓝型油菜自交系07191锌胁迫处理前和处理7 d后的幼苗, 于液氮中保存, 参照曲存民等[16]方法提取总RNA, 利用Illumina Hiseq 2500进行转录组测序, 以甘蓝型油菜为参考基因组[32], 利用Cuffdiff筛选差异表达基因, 筛选标准为FDR < 0.01, log2|(fold change)|>1。上调和下调的差异表达基因GO富集分析采用基迪奥云平台进行分析(2 结果与分析
2.1 甘蓝型油菜发芽期锌胁迫最佳浓度确定
为了确定油菜发芽期锌胁迫的最佳处理浓度, 本研究随机挑选10个材料, 分别利用不同浓度的Zn(NO3)2溶液和纯水为对照进行处理, 发现处理浓度为30 mg L-1时, 这些材料下胚轴长度明显受到抑制, 且材料间变异较大(表1)。因此, 在发芽期筛选耐锌油菜的最佳浓度为30 mg L-1。2.2 甘蓝型油菜锌胁迫性状表型变异分析
对140份油菜正常和锌胁迫条件下的下胚轴长度进行统计发现, 在正常条件下, 甘蓝型油菜下胚轴长度为3.66~7.99 cm, 变异系数为15.8%; 锌胁迫下, 下胚轴长度为2.28~6.47 cm, 变异系数为18.8%; 相对下胚轴长的变异范围为0.38~1.08, 均值为0.73, 变异系数为17.6%, 表现出广泛的表型变异(表2)。锌胁迫下, 大部分材料的下胚轴长度都较正常生长状态的下胚轴短, 只有5个半冬性材料例外(甘油5号、SWU94、中油821、川油18和wx10213), 表明30 mg L-1锌胁迫严重抑制了甘蓝型油菜下胚轴伸长。相对下胚轴长呈连续性正态分布, 说明其受多基因控制, 表型为典型的数量性状遗传特点(图1)。Table 2
表2
表2锌胁迫下甘蓝型油菜发芽期性状统计分析
Table 2
| 性状 | 最小值 | 最大值 | 均值±标准差 | 变异系数 | 偏度 | 峰度 |
|---|---|---|---|---|---|---|
| Trait | Min.(cm) | Max.(cm) | Mean±SD | CV (%) | Skewness | Kurtosis |
| CHL | 3.66 | 7.99 | 5.77±0.92 | 15.85 | -0.13 | 0.83 |
| THL | 2.28 | 6.47 | 4.24±0.80 | 18.84 | 0.41 | 0.02 |
| RHL | 0.38 | 1.08 | 0.73±0.13 | 17.57 | 0.28 | 0.15 |
新窗口打开|下载CSV
图1
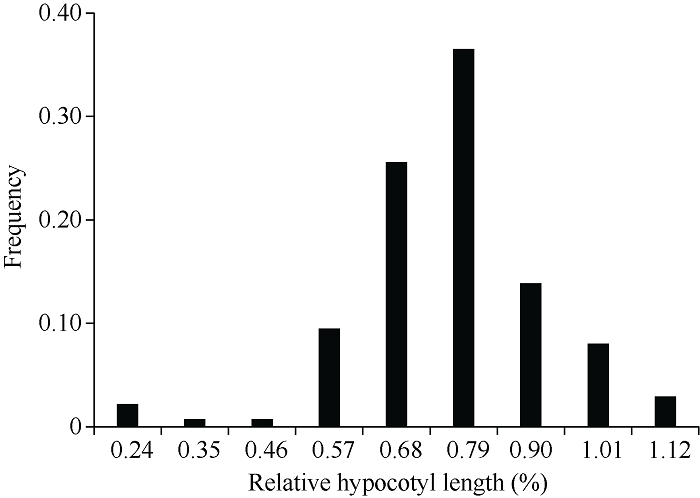 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1锌胁迫下甘蓝型油菜相对下胚轴长频率分布图
Fig. 1Frequency distribution of RHL under zinc stress in B. napus
2.3 群体结构和亲缘关系分析
利用30,288个SNP标记进行关联群体的群体结构和亲缘关系分析, 当K=2时, ΔK值最大(图2-A), 因此, 该群体的亚群数为2, 生成的最优群体结构的Q矩阵见图2-B。140份甘蓝型材料中, 89%材料间亲缘关系小于0.1 (图2-C), 表明该群体内材料间的亲缘关系较远。图2
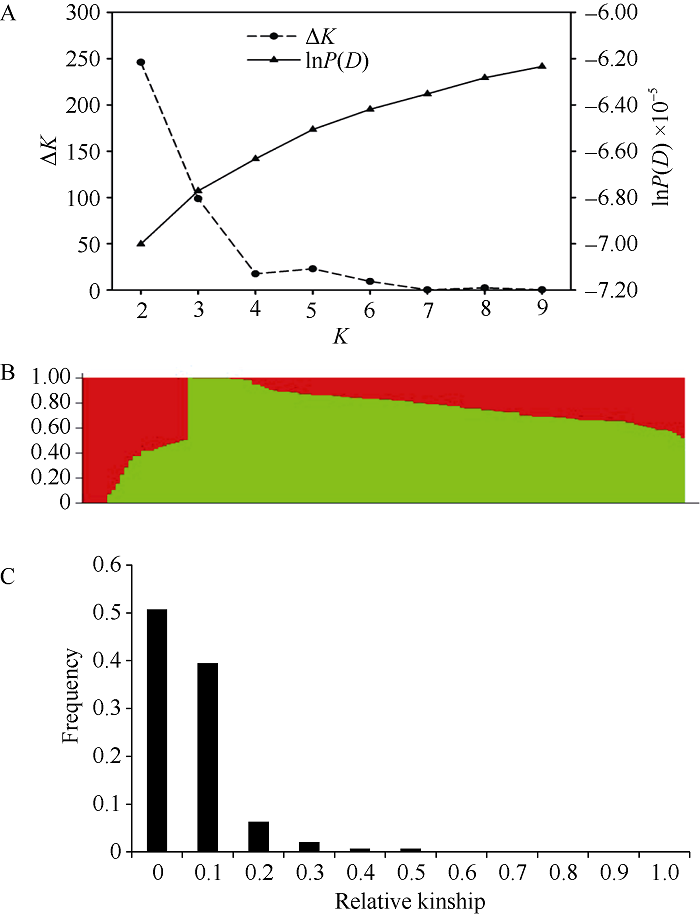 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2基于SNP标记估算群体结构关系图和亲缘关系图
A: lnP(D)和ΔK变化关系图; B: K = 2时群体结构图; C: 140份甘蓝型油菜亲缘关系图。
Fig. 2Population structure and relative kinship diagram based on SNP markers
A: plot of lnP(D) and ΔK; B: population structure profile (K = 2), the areas of the two colors (green and red) indicate the proportion of each subgroup, respectively; C: relative kinship diagram of 140 B. napus.
2.4 耐锌胁迫下相对下胚轴长度的全基因组关联分析
对相对下胚轴长度的QQ图进行分析(图3), 结果表明, K+Q模型检测到的标记的P值较其他模型更接近期望值, 其降低了假阳性, 关联分析结果更可靠。全基因组关联分析共检测到8个显著关联的SNP位点(图3和表3), 分别位于A02、A07、C04和C06染色体上, 单个SNP解释的表型变异为22.0%~33.2%。为了确定显著关联SNP位点的候选基因区间, 本研究对这些关联位点的染色体进行LD分析, 利用R2值估计LD, A02和A07染色体的衰减距离为300 kb, C04染色体衰减距离为700 kb, C06染色体的衰减距离为1 Mb (图4-A)。另外, A07染色体上22.2 Mb~23.9 Mb与C06染色体34.9 Mb~37.2 Mb位于共线性区间内(图4-B), 说明找到的显著关联的SNP位点位于这2个共线性区间内。图3
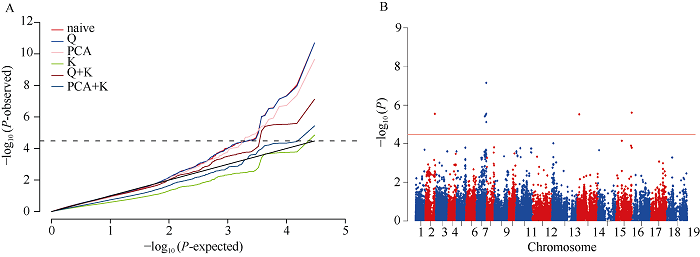 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3锌胁迫下甘蓝型油菜相对下胚轴长QQ图和曼哈顿图
Fig. 3QQ and Manhattan plots on RHL under zinc stress treatment in B. napus
Table 3
表3
表3锌胁迫下甘蓝型油菜发芽期相对下胚轴长显著关联的SNP位点
Table 3
| 染色体 Chr. | 位置 Position (bp) | P值 P-value | 显著性 -log10 (P) | 贡献率 R2 (%) |
|---|---|---|---|---|
| A02 | 22,075,111 | 2.97E-06 | 5.53 | 24.62 |
| A07 | 20,094,111 | 3.97E-06 | 5.40 | 22.11 |
| A07 | 22,001,673 | 3.49E-06 | 5.46 | 21.99 |
| A07 | 23,402,490 | 7.70E-06 | 5.11 | 19.90 |
| A07 | 23,555,825 | 2.92E-06 | 5.53 | 22.06 |
| A07 | 23,580,640 | 7.50E-08 | 7.13 | 33.18 |
| C04 | 4,456,230 | 3.12E-06 | 5.51 | 24.66 |
| C06 | 36,835,784 | 2.55E-06 | 5.59 | 24.45 |
新窗口打开|下载CSV
图4
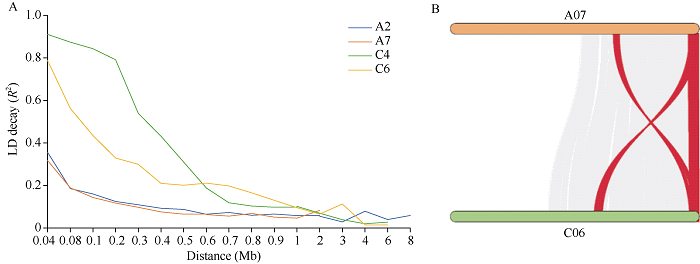 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4甘蓝型油菜染色体不同物理距离的LD分布(A)和共线性分析(B)
Fig. 4LD distribution (A) and synteny analysis (B) in different distances in B. napus
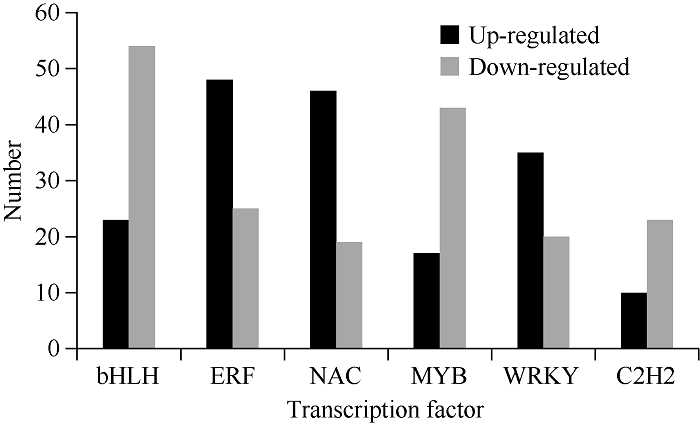
2.5 锌胁迫下油菜幼苗差异表达基因
转录组分析结果表明, 锌处理油菜幼苗7 d后, 与对照相比, 上调基因2733个, 下调基因3652个, 其中差异转录因子/基因616个, 主要包括bHLH (77)、ERF (73)、NAC (65)、MYB (60)、WRKY (55)和C2H2 (33)家族(图5)。ERF、NAC和WRKY家族基因上调数目较多, 而bHLH、MYB和C2H2家族基因下调基因数目较多。本研究对上调和下调基因分别进行GO功能富集分析, 上调基因主要富集在细胞外区域和细胞膜, 分子功能是硫酸盐跨膜转运功能, 主要参与氧化还原反应、离子转运、胁迫反应、防御反应和硫化合物转运(图6-A)。下调基因主要富集在细胞外区域、质外体和细胞膜区域, 主要的分子功能是抗氧化活性和过氧化物酶活性, 主要参与抗氧化反应、氧化还原反应和细胞壁生物合成(图6-B)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5油菜锌胁迫后上调和下调转录因子基因数目
Fig. 5Number of up-regulated and down-regulated transcription factors under zinc stress treatment in B. napus
图6
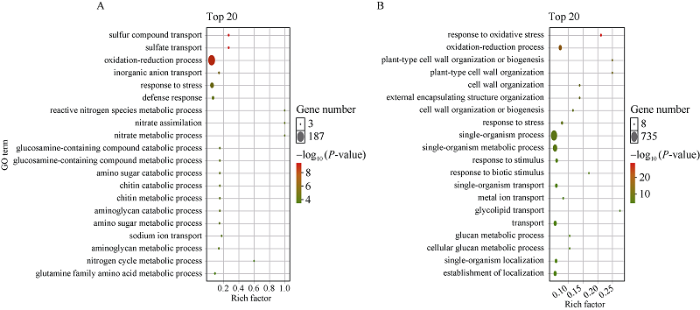 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6油菜锌胁迫后差异表达上调(A)和下调基因(B) GO富集分析
Fig. 6Differentially up-regulated (A) and down-regulated (B) genes by GO enrichment analysis under zinc stress treatment in B. napus
2.6 GWAS与转录组结合鉴定共有基因
根据各染色体的LD确定候选区间, 参照油菜基因组注释信息[32], 共找到19个与锌胁迫相关的候选基因(表4), 这些基因主要包括编码锌指蛋白家族成员(B-box型和ZFP1)、谷胱甘肽转移酶GSTU21、过氧化物酶家族蛋白、ABC和MFS转运蛋白及细胞壁相关激酶蛋白, 并找到一些重要的转录因子, 如MYB、bHLH、NAC、ERF和WRKY, 大部分基因(除GSTU21、ZFP1)。Table 4
表4
表4甘蓝型油菜锌胁迫下相关性状候选基因
Table 4
| 候选基因 Candidate genes | 物理位置 Physical position | 拟南芥同源基因 Homologs in Arabidopsis | 基因注释 Gene annotation | 差异倍数 log2 (Fold change) |
|---|---|---|---|---|
| BnaA02g30040D | Chr.A02: 21813781-21815282 | AT5G48250 | B-box型锌指蛋白 B-box type zinc finger protein with CCT domain | 2.18 |
| BnaA02g30270D | Chr.A02: 21934212-21936672 | AT5G48560 | bHLH转录因子 Basic helix-loop-helix (bHLH) DNA-binding protein | -6.62 |
| BnaA07g27330D | ChrA07: 19877781-19879164 | AT1G68670 | MYB转录因子 MYB-like transcription factor family protein | 2.15 |
| BnaA07g27340D | Chr.A07: 19884616-19887579 | AT1G68690 | 蛋白激酶家族 Protein kinase superfamily protein | 4.55 |
| BnaA07g27500D | Chr.A07: 19996402-19997781 | AT1G68850 | 过氧化物酶家族蛋白 Peroxidase superfamily protein | -2.06 |
| BnaA07g27840D | Chr.A07: 20196357-20198149 | AT1G69310 | WRKY57转录因子 WRKY DNA-binding protein 57 | 2.64 |
| BnaA07g28000D | Chr.A07: 20266703-20267687 | AT1G69490 | NAC转录因子 NAC-like, activated by AP3/PI (NAP) | 2.01 |
| BnaA07g31860D | Chr.A07: 22136736-22137327 | AT1G74930 | ERF转录因子 ORA47 | 3.71 |
| BnaA07g34210D | Chr.A07: 23311591-23312410 | AT1G78360 | 谷胱甘肽转移酶 (GSTU21) Glutathione S-transferase TAU 21 (GSTU21) | -3.13 |
| BnaA07g35030D | Chr.A07: 23695006-23698790 | AT1G79610 | Na+/H+逆向转运蛋白 Na+/H+ antiporter 6 (NHX6) | 2.29 |
| BnaA07g35090D | Chr.A07: 23714116-23714860 | AT1G79680 | 类细胞壁相关激酶WALL ASSOCIATED KINASE (WAK)-LIKE 10 (WAKL10) | 6.52 |
| BnaA07g35350D | Chr.A07: 23800896-23801571 | AT1G80730 | 锌指蛋白 Zinc-finger protein 1 (ZFP1) | -7.08 |
| BnaC04g06210D | Chr.C04: 4439046-4439940 | AT3G55090 | ABC转运蛋白 ABC-2 type transporter family protein | 2.10 |
| BnaC04g06300D | Chr.C04: 4503767-4505873 | AT2G39210 | 主要协助转运蛋白超家族 Major facilitator superfamily protein (MFS) | 3.11 |
| BnaC06g38840D | Chr.C06: 36254140-36254958 | AT1G78360 | 谷胱甘肽转移酶(GSTU21) Glutathione S-transferase TAU 21 (GSTU21) | -4.29 |
| BnaC06g38850D | Chr.C06: 36255467-36256984 | AT1G78360 | 谷胱甘肽转移酶(GSTU21) Glutathione S-transferase TAU 21 (GSTU21) | -2.35 |
| BnaC06g39970D | Chr.C06: 36849441-36853390 | AT1G79610 | Na+/H+逆向转运蛋白 Na+/H+ antiporter 6 (NHX6) | 4.01 |
| BnaC06g40020D | Chr.C06: 36867031-36869575 | AT1G69730 | 细胞壁相关激酶蛋白 Wall-associated kinase family protein | 4.43 |
| BnaC06g40250D | Chr.C06: 36958965-36959628 | AT1G80730 | 锌指蛋白 Zinc-finger protein 1 (ZFP1) | -2.49 |
新窗口打开|下载CSV
3 讨论
重金属污染可归纳为2类, 一类是一些具有显著生物毒性的重金属物, 如镉(Cd)、砷(As)等, 另一类为超过一定量后具有生物毒性的一些重金属, 如锌(Zn)、铜(Cu)等[33]。近年来, 土壤重金属污染越来越引起人们关注[34]。在土壤中过量的锌富积会抑制植物生长, 导致营养失衡、萎黄叶片, 以及光合作用受损[5,6]。研究表明, 甘蓝型油菜对重金属锌具有较强的耐受性, 在较高浓度下仍可获得较高的生物产量[2,3,4,5]。然而关于油菜响应锌胁迫的分子机制尚未见报道。在甘蓝型油菜中, 通过GWAS分析与基因组信息相结合已成为挖掘油菜数量性状基因座的常规手段, 并获得了一些与非生物胁迫相关的候选基因[14-16,35-37]。因此, 本研究通过对锌胁迫下140份甘蓝型油菜发芽期的相对下胚轴长进行了全基因组关联分析, 共检测到8个显著关联的SNP位点, A02、C04和C06染色体上显著关联的SNP位点分别位于22.0、4.4和36.8 Mb, A07染色体上显著关联的SNP位点位于22.0 Mb和23.5 Mb。曲存民等[16]将砷胁迫下甘蓝型油菜的RHL定位在A02: 19.2 Mb~22.0 Mb、A07: 19.2 Mb~22.5 Mb、A07: 23.1 Mb~23.5 Mb、C04: 4.3 Mb~4.5 Mb和C06: 31.6 Mb~36.8 Mb, 与本研究定位结果一致, 这些位点具有一因多效性, 可能在不同重金属胁迫过程中都起着重要作用。在植物中, 重金属可以引起许多代谢过程发生变化, 这些变化主要是通过氧化还原反应引起的, 同时造成抗氧化酶的活化、细胞膜通透性改变和离子的置换等反应[38]。本研究发现锌胁迫后, 上调的差异表达基因主要富集在细胞膜, 参与氧化还原反应、离子转运、防御反应和硫化合物转运。本研究结合全基因组关联分析和转录组测序, 找到19个与锌胁迫相关的候选基因, 包括转录因子/基因, 如WRKY57、ORA47和BnaA07g28000D (NAC), WRKY57可以通过清除活性氧提高植物抗旱和抗盐能力[39,40]; 植物受到伤害和水分胁迫时, ORA47可通过调控脱落酸和茉莉酸信号途径来提高抗性[41]。本研究中WRKY57和ORA47在锌胁迫后显著上调表达, 可能在锌胁迫过程中起着重要作用, 但具体的调控机制还需进一步研究。
本研究还找到了B-box型和锌指蛋白ZFP1, 锌指蛋白中的锌指结构可以结合锌离子形成稳定的指状结构, 编码锌指蛋白的众多基因已经被证明在生物和非生物胁迫过程中起着重要作用[42,43]。Sun等[44]研究表明, 菜豆MTF通过激活色氨酸生物合成调控烟草耐镉性, 锌指基序是其重要的结合金属反应的元件。拟南芥锌指蛋白ZAT6通过谷胱甘肽途径提高镉抗性[45], 而锌指蛋白ZAT11负调控拟南芥镍抗性[46]。本研究找到的B-box型锌指蛋白基因BnaA02g30040D在锌胁迫后表达量上调, 而锌指蛋白ZFP1基因表达量下调, 它们可能通过不同的机制来响应胁迫。另外, 本研究找到2个编码Na+/H+逆向转运蛋白的基因NHX6 (BnaA07g35030D和BnaC06g39970D), 锌胁迫后其表达量上调, Yang等[47]研究表明, 浮萍NHX1基因通过减少Cd2+流入和增加Cd2+外排提高重金属镉抗性。本研究中NHX6表达上调, 可能增加了Zn2+的外排, 从而提高锌抗性。
重金属处理下植物细胞产生活性氧造成细胞氧化损伤, 为了抵抗活性氧对细胞的损伤, 植物会产生抗氧化酶及非酶抗氧化剂清除自由基[48]。锌处理下小麦过氧化氢含量、过氧化物酶及过氧化氢酶活性均显著提高, 增强了小麦对锌的耐受性[49]。本研究找到一些与氧化还原反应相关的基因, 如编码过氧化物酶家族蛋白和谷胱甘肽转移酶基因GSTU21。GST可使GSH与内源电子化合物结合而产生脱毒[50], 重金属(Cu和Al)胁迫下, 拟南芥GST基因表达上调, 超表达水稻GSTU30基因提高了拟南芥重金属耐性[51,52]。但是本研究中, GSTU21基因在锌胁迫后表达量下调, 可能是由于锌处理植株后, 重金属造成的氧化胁迫反应超过了油菜自身的协调能力, 抗氧化能力受到明显抑制。
另外, 本研究还定位到一些编码ABC和MSF转运蛋白的基因, 其功能是将重金属转运到液泡内, 在重金属转运过程中起着重要作用[53,54], 这些转运蛋白基因表达量上调, 可能通过相似的途径增强植株对锌的抗性。拟南芥铝胁迫后, 可以诱导WAK1基因的表达[55]; WALK4基因启动子的突变导致水稻对钾、钠、铜和锌超敏感, WALK4突变导致水稻茎中锌含量下降[56]; WAKL10基因表达量上调, 其可能在锌胁迫过程中起着重要作用。本研究为深入研究甘蓝型油菜响应锌胁迫的分子机制奠定了基础。
4 结论
全基因组关联分析共鉴定出8个与锌胁迫显著关联的SNP位点, 结合转录组测序结果, 共找到19个候选基因, 可能在锌胁迫过程中起着重要作用。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/0048-9697(77)90043-2URL [本文引用: 1]
DOI:10.1016/j.bbamcr.2012.05.016URLPMID:22626733 [本文引用: 3]

The use of the essential element zinc (Zn) in the biochemistry of land plants is widespread, and thus comparable to that in other eukaryotes. Plants have evolved the ability to adjust to vast fluctuations in external Zn supply, and they can store considerable amounts of Zn inside cell vacuoles. Moreover, among plants there is overwhelming, but yet little explored, natural genetic diversity that phenotypically affects Zn homeostasis. This results in the ability of specific races or species to thrive in different soils ranging from extremely Zn-deficient to highly Zn-polluted. Zn homeostasis is maintained by a tightly regulated network of low-molecular-weight ligands, membrane transport and Zn-binding proteins, as well as regulators. Here we review Zn homeostasis of land plants largely based on the model plant Arabidopsis thaliana, for which our molecular understanding is most developed at present. There is some evidence for substantial conservation of Zn homeostasis networks among land pants, and this review can serve as a reference for future comparisons. Major progress has recently been made in our understanding of the regulation of transcriptional Zn deficiency responses and the role of the low-molecular-weight chelator nicotianamine in plant Zn homeostasis. Moreover, we have begun to understand how iron (Fe) and Zn homeostasis interact as a consequence of the chemical similarity between their divalent cations and the lack of specificity of the major root iron uptake transporter IRT1. The molecular analysis of Zn-hyperaccumulating plants reveals how metal homeostasis networks can be effectively modified. These insights are important for sustainable bio-fortification approaches. This article is part of a Special Issue entitled: Cell Biology of Metals.
[本文引用: 2]
DOI:10.1111/j.1469-8137.2007.01996.xURLPMID:17286818 [本文引用: 2]
Zinc (Zn) is an essential component of thousands of proteins in plants, although it is toxic in excess. In this review, the dominant fluxes of Zn in the soil-root-shoot continuum are described, including Zn inputs to soils, the plant availability of soluble Zn(2+) at the root surface, and plant uptake and accumulation of Zn. Knowledge of these fluxes can inform agronomic and genetic strategies to address the widespread problem of Zn-limited crop growth. Substantial within-species genetic variation in Zn composition is being used to alleviate human dietary Zn deficiencies through biofortification. Intriguingly, a meta-analysis of data from an extensive literature survey indicates that a small proportion of the genetic variation in shoot Zn concentration can be attributed to evolutionary processes whose effects manifest above the family level. Remarkable insights into the evolutionary potential of plants to respond to elevated soil Zn have recently been made through detailed anatomical, physiological, chemical, genetic and molecular characterizations of the brassicaceous Zn hyperaccumulators Thlaspi caerulescens and Arabidopsis halleri.
DOI:10.1016/j.chemosphere.2011.10.039URLPMID:22099539 [本文引用: 3]
The halophytic shrub Halimione portulacoides is known to be capable of growth in soils containing extremely high concentrations of Zn. This study evaluated in detail the tolerance and accumulation potential of H. portulacoides under moderate and high external Zn levels. A greenhouse experiment was conducted in order to investigate the effects of a range of Zn concentrations (0-130 mmol L(-1)) on growth and photosynthetic performance by measuring relative growth rate, total leaf area, specific leaf area, gas exchange, chlorophyll fluorescence parameters and photosynthetic pigment concentrations. We also determined the total zinc, nitrogen, phosphorus, calcium, magnesium, sodium, potassium, iron and copper concentrations in the plant tissues. H. portulacoides demonstrated hypertolerance to Zn stress, since it survived with leaf concentrations of up to 2300 mg Zn kg(-1)dry mass, when treated with 130 mmol Zn L(-1). Zinc concentrations greater than 70 mmol L(-1) in the nutrient solution negatively affected plant growth, in all probability due to the recorded decline in net photosynthesis rate. Our results indicate that the Zn-induced decline in the photosynthetic function of H. portulacoides may be attributed to the adverse effect of the high concentration of the metal on photosynthetic electron transport. Growth parameters were virtually unaffected by leaf tissue concentrations as high as 1500 mg Zn kg(-1)dry mass, demonstrating the strong capability of H. portulacoides to protect itself against toxic Zn concentrations. The results of our study indicate that this salt-marsh shrub may represent a valuable tool in the restoration of Zn-polluted areas.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s00425-006-0421-yURLPMID:17086401 [本文引用: 1]
Phytochelatins (PCs) are heavy metal binding peptides that play an important role in sequestration and detoxification of heavy metals in plants. In this study, our goal was to develop transgenic plants with increased tolerance for and accumulation of heavy metals from soil by expressing an Arabidopsis thaliana AtPCS1 gene, encoding phytochelatin synthase (PCS), in Indian mustard (Brassica juncea L.). A 35S promoter fused to a FLAG-tagged AtPCS1 cDNA was expressed in Indian mustard, and transgenic lines, designated pc lines, were evaluated for tolerance to and accumulation of Cd and Zn. Transgenic plants with moderate AtPCS1 expression levels showed significantly higher tolerance to Cd and Zn stress, but accumulated significantly less Cd and Zn than wild type plants in both shoot and root tissues. However, transgenic plants with highest expression of the transgene did not exhibit enhanced Cd and Zn tolerance. Shoots of Cd-treated pc plants had significantly higher levels of phytochelatins and thiols than wild-type plants. Significantly lower concentrations of gluthatione in Cd-treated shoot and root tissues of transgenic plants were observed. Moderate expression levels of phytochelatin synthase improved the ability of Indian mustard to tolerate certain levels of heavy metals, but at the same time did not increase the accumulation potential for Cd and Zn.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29078746 [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 6]
[本文引用: 6]
URLPMID:19495770 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:23430174 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:10835412 [本文引用: 1]
We describe a model-based clustering method for using multilocus genotype data to infer population structure and assign individuals to populations. We assume a model in which there are K populations (where K may be unknown), each of which is characterized by a set of allele frequencies at each locus. Individuals in the sample are assigned (probabilistically) to populations, or jointly to two or more populations if their genotypes indicate that they are admixed. Our model does not assume a particular mutation process, and it can be applied to most of the commonly used genetic markers, provided that they are not closely linked. Applications of our method include demonstrating the presence of population structure, assigning individuals to populations, studying hybrid zones, and identifying migrants and admixed individuals. We show that the method can produce highly accurate assignments using modest numbers of loci-e.g. , seven microsatellite loci in an example using genotype data from an endangered bird species. The software used for this article is available from http://www.stats.ox.ac.uk/ approximately pritch/home. html.
URLPMID:15969739 [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
URLPMID:28158525 [本文引用: 1]
DOI:10.1126/science.1253435URLPMID:25146293 [本文引用: 2]
Oilseed rape (Brassica napus L.) was formed ~7500 years ago by hybridization between B. rapa and B. oleracea, followed by chromosome doubling, a process known as allopolyploidy. Together with more ancient polyploidizations, this conferred an aggregate 72x genome multiplication since the origin of angiosperms and high gene content. We examined the B. napus genome and the consequences of its recent duplication. The constituent An and Cn subgenomes are engaged in subtle structural, functional, and epigenetic cross-talk, with abundant homeologous exchanges. Incipient gene loss and expression divergence have begun. Selection in B. napus oilseed types has accelerated the loss of glucosinolate genes, while preserving expansion of oil biosynthesis genes. These processes provide insights into allopolyploid evolution and its relationship with crop domestication and improvement.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.scitotenv.2014.04.127URLPMID:24875258 [本文引用: 1]
An extensive soil survey was conducted to study pollution sources and delineate contamination of heavy metals in one of the metalliferous industrial bases, in the karst areas of southwest China. A total of 597 topsoil samples were collected and the concentrations of five heavy metals, namely Cd, As (metalloid), Pb, Hg and Cr were analyzed. Stochastic models including a conditional inference tree (CIT) and a finite mixture distribution model (FMDM) were applied to identify the sources and partition the contribution from natural and anthropogenic sources for heavy metal in topsoils of the study area. Regression trees for Cd, As, Pb and Hg were proved to depend mostly on indicators of anthropogenic activities such as industrial type and distance from urban area, while the regression tree for Cr was found to be mainly influenced by the geogenic characteristics. The FMDM analysis showed that the geometric means of modeled background values for Cd, As, Pb, Hg and Cr were close to their background values previously reported in the study area, while the contamination of Cd and Hg were widespread in the study area, imposing potentially detrimental effects on organisms through the food chain. Finally, the probabilities of single and multiple heavy metals exceeding the threshold values derived from the FMDM were estimated using indicator kriging (IK) and multivariate indicator kriging (MVIK). The high probabilities exceeding the thresholds of heavy metals were associated with metalliferous production and atmospheric deposition of heavy metals transported from the urban and industrial areas. Geostatistics coupled with stochastic models provide an effective way to delineate multiple heavy metal pollution to facilitate improved environmental management. (C) 2014 Elsevier B.V.
[本文引用: 1]
DOI:10.3389/fpls.2015.01058URLPMID:26640475
Drought stress can directly inhibit seedling establishment in canola (Brassica napus), resulting in lower plant densities and reduced yields. To dissect this complex trait, 140 B. napus accessions were phenotyped under normal (0.0 MPa, S0) and water-stressed conditions simulated by polyethylene glycol (PEG) 6000 (-0.5 MPa, S5) in a hydroponic system. Phenotypic variation and heritability indicated that the root to shoot length ratio was a reliable indicator for water stress tolerance. Thereafter, 66 accessions (16 water stress tolerant, 34 moderate and 16 sensitive lines) were genotyped using 25,495 Brassica single nucleotide polymorphisms (SNPs). Genome-wide association studies (GWAS) identified 16 loci significantly associated with water stress response. Two B. napus accessions were used for RNA sequencing, with differentially-expressed genes under normal and water-stressed conditions examined. By combining differentially-expressed genes detected by RNA sequencing with significantly associated loci from GWAS, 79 candidate genes were identified, of which eight were putatively associated with drought tolerance based on gene ontology of Arabidopsis. Functional validation of these genes may confirm key drought-related genes for selection and breeding in B. napus. Our results provide insight into the genetic basis of water stress tolerance in canola.
DOI:10.1186/s12870-020-2253-5URLPMID:32028890 [本文引用: 1]
BACKGROUND: Cold stress is one of the primary environmental factors that affect plant growth and productivity, especially for crops like Brassica napus that live through cold seasons. Till recently, although a number of genes and pathways involved in B. napus cold response have been revealed by independent studies, a genome-wide identification of the key regulators and the regulatory networks is still lack. In this study, we investigated the transcriptomes of cold stressed semi-winter and winter type rapeseeds in short day condition, mainly with the purpose to systematically identify the functional conserved transcription factors (TFs) in cold response of B. napus. RESULTS: Global modulation of gene expression was observed in both the semi-winter type line (158A) and the winter type line (SGDH284) rapeseeds, in response to a seven-day chilling stress in short-day condition. Function analysis of differentially expressed genes (DEGs) revealed enhanced stresses response mechanisms and inhibited photosynthesis in both lines, as well as a more extensive inhibition of some primary biological processes in the semi-winter type line. Over 400 TFs were differentially expressed in response to cold stress, including 56 of them showed high similarity to the known cold response TFs and were consistently regulated in 158A and SGDH284, as well as 25 TFs which targets were over-represented in the total DEGs. A further investigation based on their interactions indicated the critical roles of several TFs in cold response of B. napus. CONCLUSION: In summary, our results revealed the alteration of gene expression in cold stressed semi-winter and winter ecotype B. napus lines and provided a valuable collection of candidate key regulators involved in B. napus response to cold stress, which could expand our understanding of plant stress response and benefit the future improvement of the breed of rapeseeds.
In: Sablok G, eds. Plant Metallomics and Functional Omics.
[本文引用: 1]
DOI:10.3389/fpls.2016.00145URLPMID:26904091 [本文引用: 1]
Drought stress is a severe environmental factor that greatly restricts plant distribution and crop production. Recently, we have found that overexpressing AtWRKY57 enhanced drought tolerance in Arabidopsis thaliana. In this study, we further reported that the Arabidopsis WRKY57 transcription factor was able to confer drought tolerance to transgenic rice (Oryza sativa) plants. The enhanced drought tolerance of transgenic rice was resulted from the lower water loss rates, cell death, malondialdehyde contents and relative electrolyte leakage while a higher proline content and reactive oxygen species-scavenging enzyme activities was observed during stress conditions. Moreover, further investigation revealed that the expression levels of several stress-responsive genes were up-regulated in drought-tolerant transgenic rice plants, compared with those in wild-type plants. In addition to the drought tolerance, the AtWRKY57 over-expressing plants also had enhanced salt and PEG stress tolerances. Taken together, our study indicates that over-expressing AtWRKY57 in rice improved not only drought tolerance but also salt and PEG tolerance, demonstrating its potential role in crop improvement.
URLPMID:22930734 [本文引用: 1]
[本文引用: 1]
DOI:10.3389/fpls.2016.01272URLPMID:27605931 [本文引用: 1]
Plant zinc finger proteins (ZFPs) comprise a large protein family and they are mainly involved in abiotic stress tolerance. Although Arabidopsis RING/FYVE/PHD ZFP At5g62460 (AtRZFP) is found to bind to zinc, whether it is involved in abiotic stress tolerance is still unknown. In the present study, we characterized the roles of AtRZFP in response to abiotic stresses. The expression of AtRZFP was induced significantly by salt and osmotic stress. AtRZFP positively mediates tolerance to salt and osmotic stress. Additionally, compared with wild-type Arabidopsis plants, plants overexpressing AtRZFP showed reduced reactive oxygen species (ROSs) accumulation, enhanced superoxide dismutase and peroxidase activity, increased soluble sugars and proline contents, reduced K(+) loss, decreased Na(+) accumulation, stomatal aperture and the water loss rate. Conversely, AtRZFP knockout plants displayed the opposite physiological changes when exposed to salt or osmotic stress conditions. These data suggested that AtRZFP enhances salt and osmotic tolerance through a series of physiological processes, including enhanced ROSs scavenging, maintaining Na(+) and K(+) homeostasis, controlling the stomatal aperture to reduce the water loss rate, and accumulating soluble sugars and proline to adjust the osmotic potential.
URLPMID:27041000 [本文引用: 1]
DOI:10.1104/pp.114.253096URLPMID:25624396 [本文引用: 1]
Cadmium (Cd) is highly toxic to plants. Modulation of Cd-responsive transcription is an important way for Cd detoxification in plants. Metal-responsive element (MRE) is originally described in animal metallothionein genes. Although functional MREs also exist in Cd-regulated plant genes, specific transcription factors that bind MRE to regulate Cd tolerance have not been identified. Previously, we showed that Cd-inducible bean (Phaseolus vulgaris) stress-related gene2 (PvSR2) produces a short (S) PvSR2 transcript (S-PvSR2) driven by an intronic promoter. Here, we demonstrate that S-PvSR2 encodes a bean MRE-binding transcription factor1 (PvMTF-1) that confers Cd tolerance in tobacco (Nicotiana tabacum). PvMTF-1 expression was up-regulated by Cd at the levels of RNA and protein. Importantly, expression of PvMTF-1 in tobacco enhanced Cd tolerance, indicating its role in regulating Cd resistance in planta. This was achieved through direct regulation of a feedback-insensitive Anthranilate Synthase alpha-2 chain gene (ASA2), which catalyzes the first step for tryptophan biosynthesis. In vitro and in vivo DNA-protein interaction studies further revealed that PvMTF-1 directly binds to the MRE in the ASA2 promoter, and this binding depends on the zinc finger-like motif of PvMTF-1. Through modulating ASA2 up-regulation by Cd, PvMTF-1 increased free tryptophan level and subsequently reduced Cd accumulation, thereby enhancing Cd tolerance of transgenic tobacco plants. Consistent with this observation, tobacco transiently overexpressing ASA2 also exhibited increased tolerance to Cd. We conclude that PvMTF-1 is a zinc finger-like transcription factor that links MRE to Cd resistance in transgenic tobacco through activation of tryptophan biosynthesis.
URLPMID:26983992 [本文引用: 1]
DOI:10.1007/s00299-014-1675-7URLPMID:25163803 [本文引用: 1]
KEY MESSAGE: ZAT11, a Zinc Finger of Arabidopsis Thaliana 11, is a dual-function transcriptional regulator that positively regulates primary root growth but negatively regulates Ni (2+) tolerance. Zinc Finger of Arabidopsis Thaliana 11 (ZAT11) is a C2H2-type zinc finger protein that has been reported to function as an active transcriptional repressor. However, the biological function of ZAT11 remains unknown. Here we show that GFP-tagged ZAT11 is targeted to the nucleus. Analysis of plants expressing ZAT11 promoter-GUS showed that ZAT11 is highly expressed in roots and particularly in root tips. To identify the biological function of ZAT11, we constructed three independent lines of ZAT11 overexpressing transgenic plant (ZAT11 OE). ZAT11 OE enhanced the elongation of primary root but reduced the metal tolerance against nickel ion (Ni(2+)). The reduced Ni(2+) tolerance of ZAT11 OE was correlated with decreased accumulation of Ni(2+) in plants. The decreased accumulation of Ni(2+) in ZAT11 OE was caused by the reduced transcription of a vacuolar Ni(2+) transporter gene. Taken together, our results suggest that ZAT11 is a dual function transcriptional regulator that positively regulates primary root growth but negatively regulates Ni(2+) tolerance.
DOI:10.1016/j.ecoenv.2019.109397URLPMID:31299476 [本文引用: 1]
Cadmium (Cd) is a serious threat to plants health. Though some genes have been reported to get involved in the regulation of tolerance to Cd, the mechanisms underlying this process are not fully understood. Na(+)/H(+) antiporter (NHX1) plays an important role in Na(+)/H(+) trafficking. The salt and cadmium stress tolerance were found to be enhanced by NHX1 in duckweed according to our previous study, however, its function in Cd(2+) flux under Cd stress has not been studied. Here we explored the Cd(2+) flux in wild type (WT) and NHX1 transgenic duckweed (NHX1) under Cd stress. We found that the Cd(2+) influx in NHX1 duckweed was significantly declined, followed by an increased Cd(2+) efflux after 20min treatment of Cd, which resulted a less accumulation of Cd in NHX1. Reversely, inhibition of NHX1 by amiloride treatment, enhanced Cd(2+) influx in NHX1 duckweed, subsequently delayed Cd(2+) efflux in both genotypes of duckweed under Cd(2+) shock. H(+) efflux in NHX1 duckweed was lower compare with that in WT with 20min Cd(2+) shock. NHX1 also increased the pH value with Cd(2+) stress in the transgenic rhizoid. These finding suggested a new function of NHX1 in regulation of Cd(2+) and H(+) flow during short-term Cd(2+) shock.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29930563 [本文引用: 1]
DOI:10.1104/pp.122.3.657URLPMID:10712528 [本文引用: 1]
To examine the biological role of Al-stress-induced genes, nine genes derived from Arabidopsis, tobacco (Nicotiana tabacum L.), wheat (Triticum aestivum L.), and yeast (Saccharomyces cerevisiae) were expressed in Arabidopsis ecotype Landsberg. Lines containing eight of these genes were phenotypically normal and were tested in root elongation assays for their sensitivity to Al, Cd, Cu, Na, Zn, and to oxidative stresses. An Arabidopsis blue-copper-binding protein gene (AtBCB), a tobacco glutathione S-transferase gene (parB), a tobacco peroxidase gene (NtPox), and a tobacco GDP-dissociation inhibitor gene (NtGDI1) conferred a degree of resistance to Al. Two of these genes, AtBCB and parB, and a peroxidase gene from Arabidopsis (AtPox) also showed increased resistance to oxidative stress induced by diamide, while parB conferred resistance to Cu and Na. Al content of Al-treated root tips was reduced in the four Al-resistant plant lines compared with wild-type Ler-0, as judged by morin staining. All four Al-resistant lines also showed reduced staining of roots with 2',7'-dichloro fluorescein diacetate (H(2)DCFDA), an indicator of oxidative stress. We conclude that Al-induced genes can serve to protect against Al toxicity, and also provide genetic evidence for a link between Al stress and oxidative stress in plants.
DOI:10.1039/c8mt00204eURLPMID:30516767 [本文引用: 1]
Glutathione S-transferases (GSTs) are well-known enzymes due to their role in detoxification of xenobiotic compounds. However, their biochemical action is still not so clear in imparting tolerance to several abiotic stresses in crop plants. In our previous study, we observed that rice tau class OsGSTU30 plays a significant role in the detoxification of Cr(vi). Interestingly, q-RT PCR analysis also revealed higher expression of OsGSTU30 under drought conditions. In this study, we characterize OsGSTU30 in response to drought as well as heavy metal [Cr(vi)] stresses through overexpression in Arabidopsis thaliana. Biochemical and physiological analyses revealed that OsGSTU30 overexpression lines have improved tolerance against both stresses as compared to wild-type plants. Kinetic analysis and molecular docking confirmed that OsGSTU30 enzyme possesses both GST as well as glutathione peroxidase (GPx) like activity. Differentially expressed stress-responsive genes were also identified by transcriptome analysis, involved in different biological pathways during abiotic stresses. These results suggest the signaling functions of OsGSTU30 apart from its catalytic activity during abiotic stress responses and can be further exploited for improving the stress tolerance in crops.
DOI:10.1093/pcp/pcy006URLPMID:29361141 [本文引用: 1]
Soil contamination by heavy metals and metalloids such as cadmium (Cd) and arsenic (As) poses a major threat to the environment and to human health. Vacuolar sequestration is one of the main mechanisms by which plants control toxic materials including Cd and As. Understanding the mechanisms of heavy metal tolerance and accumulation can be useful for both phytoremediation and safe crop development. In this review, we summarize recent advances in deciphering the molecular mechanisms underlying vacuolar sequestration of Cd and As, and discuss potential biotechnological applications of this knowledge and efforts towards attaining these goals.
URLPMID:17355438 [本文引用: 1]
URLPMID:19175521 [本文引用: 1]
DOI:10.1104/pp.105.066910URLPMID:16286448 [本文引用: 1]
The cell wall-associated receptor kinase (WAK) and WAK-like kinase (WAKL) gene family members are good candidates for physical linkers that signal between the cell wall and the cytoplasmic compartment. Previous studies have suggested that while some WAK/WAKL members play a role in bacterial pathogen and heavy-metal aluminum responses, others are involved in cell elongation and plant development. Here, we report a functional role for the WAKL4 gene in Arabidopsis (Arabidopsis thaliana) mineral responses. Confocal microscopic studies localized WAKL4-green fluorescent protein fusion proteins on the cell surfaces suggesting that, like other WAK/WAKL proteins, WAKL4 protein is associated with the cell wall. Histochemical analyses of the WAKL4 promoter fused with the -glucuronidase reporter gene have shown that WAKL4 expression is induced by Na+, K+, Cu2+, Ni2+, and Zn2+. A transgenic line with a T-DNA insertion at 40-bp upstream of the WAKL4 start codon was characterized. While the T-DNA insertion had little effect on the WAKL4 transcript levels under normal growth conditions, it significantly altered the expression patterns of WAKL4 under various conditions of mineral nutrients. Semiquantitative and quantitative reverse transcription-PCR analyses showed that the promoter impairment abolished WAKL4-induced expression by Na+, K+, Cu2+, and Zn2+, but not by Ni2+. Whereas the WAKL4 promoter impairment resulted in hypersensitivity to K+, Na+, Cu2+, and Zn2+, it conferred a better tolerance to toxic levels of the Ni2+ heavy metal. WAKL4 was required for the up-regulation of zinc transporter genes during zinc deficiency, and the WAKL4 T-DNA insertion resulted in a reduction of Zn2+ accumulation in shoots. A WAKL4-green fluorescent protein fusion gene driven by either the WAKL4 native promoter or the 35S constitutive promoter complemented the phenotypes. Our results suggest versatile roles for WAKL4 in Arabidopsis mineral nutrition responses.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}