 ,1,2,3,**, 白建芳,2,3,**, 张风廷2,3, 郭丽萍2,3, 苑少华2,3, 李艳梅2,3, 张胜全2,3, 赵昌平,2,3,*, 张立平,1,2,3,*
,1,2,3,**, 白建芳,2,3,**, 张风廷2,3, 郭丽萍2,3, 苑少华2,3, 李艳梅2,3, 张胜全2,3, 赵昌平,2,3,*, 张立平,1,2,3,*Cloning and potential function analysis of ascorbic peroxidase gene of hybrid wheat in seed aging
YUE Jie-Ru,1,2,3,**, BAI Jian-Fang,2,3,**, ZHANG Feng-Ting2,3, GUO Li-Ping2,3, YUAN Shao-Hua2,3, LI Yan-Mei2,3, ZHANG Sheng-Quan2,3, ZHAO Chang-Ping,2,3,*, ZHANG Li-Ping,1,2,3,*通讯作者:
收稿日期:2020-06-12接受日期:2020-10-14网络出版日期:2021-03-12
| 基金资助: |
Received:2020-06-12Accepted:2020-10-14Online:2021-03-12
| Fund supported: |
作者简介 About authors
岳洁茹, E-mail:
白建芳, E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (11431KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
岳洁茹, 白建芳, 张风廷, 郭丽萍, 苑少华, 李艳梅, 张胜全, 赵昌平, 张立平. 杂交小麦抗坏血酸过氧化物酶基因克隆及其在种子老化中的潜在功能分析[J]. 作物学报, 2021, 47(3): 405-415. doi:10.3724/SP.J.1006.2021.01049
YUE Jie-Ru, BAI Jian-Fang, ZHANG Feng-Ting, GUO Li-Ping, YUAN Shao-Hua, LI Yan-Mei, ZHANG Sheng-Quan, ZHAO Chang-Ping, ZHANG Li-Ping.
抗坏血酸过氧化物酶(ascorbate peroxidase, APX)是降解过氧化氢的关键酶, 能清除活性氧从而降低氧自由基的浓度, 降低细胞的氧化损伤。近年来关于APX基因功能的研究主要集中在植物抗逆方面。例如, 在非生物胁迫条件下, 水稻、白桦中APX基因表达上调[1,2]。在低温条件下, 甘蔗体内的ScAPX1基因表达量上升[3], 且抗寒能力高的甘蔗内ScAPX1酶活性高于抗寒能力低的甘蔗; 在高温条件下, 转大麦APX基因的拟南芥叶片正常, 而非转基因拟南芥的叶片出现大量枯黄[4]; 在盐胁迫下, 胞质超氧化物歧化酶(superoxide dismutase, SOD)和胞质抗坏血酸过氧化物酶(APX)双基因过表达的拟南芥植株与野生型及单基因型相比表现更好, 生物量积累和纤维素含量更高[5]; 在高盐、PEG、低温处理条件下, 葡萄体内7个APX基因均呈现不同程度的上调表达来提高葡萄的抗逆性[6]。水稻类囊体APX酶在干旱胁迫下能够使叶绿体内的H2O2和参与几个重要代谢途径的基本蛋白的积累保持稳态, 从而可以更好地进行光合作用, 使植株正常生长[7]。种子活力是指种子迅速整齐萌发的发芽潜力、生长潜势和生产潜力, 是种子质量的重要指标和种用价值的重要组成部分, 其高低由遗传因素、种子发育期间的环境条件及种子贮藏条件3个方面决定[8]。二系杂交小麦是目前小麦杂种优势利用的主要途径之一, BS型不育系是光温敏二系杂交小麦的代表类型之一, 具有不育度高、易恢复、制种性状好等特性, 是北京杂交小麦品种京麦6号、京麦7号、京麦8号、京麦9号的不育系来源[9]。目前人工老化处理是研究种子活力最常用的一种方法, 并且针对二系小麦杂交种在种子活力方面已经有一定的研究, 如BNS型小麦杂交种在不同老化处理下种子发芽率、发芽势等发芽指标以及POD酶活性均随着老化时间的延长而降低, 但相同处理条件下各项指标高于常规小麦, 表明杂交种的抗老化能力明显高于常规小麦品种[10]。BS型杂交小麦是由BS型不育系和恢复系杂交产生的京麦系列杂交种, 是不同于BNS型杂交小麦的一种二系杂交小麦, 它们具有不同的遗传背景。目前尚未有关于TaAPX基因对BS型杂交小麦种子活力调控方面的研究。
因此, 本研究拟利用BS型杂交小麦及其父母本种子为研究材料, 通过对其克隆TaAPX基因, 并通过生物信息学手段分析该基因的核酸及蛋白序列特征, 预测与TaAPX基因互作的miRNAs及与其互作的相关蛋白, 分析该基因的分子结构及功能, 并运用实时荧光定量PCR (fluorescent quantitative real-time PCR, qPCR) 技术, 验证该基因对BS型杂交小麦种子活力调控方面的功能, 结合种子不同老化时间APX酶活性变化, 初步探讨TaAPX基因对BS型杂交小麦和常规种在老化过程中的调控方式及它们之间的区别, 结合预测的miRNAs的功能及APX蛋白与其他蛋白互作的预测, 为更深入研究TaAPX基因在BS型杂交小麦种子活力中的调控方式奠定基础。
1 材料与方法
1.1 试验材料与试剂
本研究以一个杂交小麦组合BS1453 (母本)、11GF5135 (父本)、BS1453/11GF5135 (杂交种)的种子为材料, 由北京市农林科学院/北京杂交小麦工程技术研究中心提供。APX酶活测定试剂盒购于北京索莱宝公司; RNA提取试剂购于北京化工厂; TRIzol试剂购自(Invitrogen, USA); 培养基所用试剂和氨苄青霉素均购自中国区西格玛公司(北京); 酶切试剂Smart buffer、BamH I均购于BioLabs; 10×EasyTaq buffer购于全式金; 凝胶回收试剂盒、质粒提取试剂盒均购自北京天根生化科技有限公司; 反转录试剂盒和荧光定量试剂盒均购于TaKaRa (Japan)。1.2 方法
1.2.1 人工老化处理 参照宋松泉等[11]的方法人工加速老化小麦, 将BS1453 (母本)、11GF5135 (父本)、BS1453/11GF5135 (杂交种)种子置于高温(41℃)、高湿(相对湿度为90%)的老化箱中老化处理, 时间分别为0、3、5、7和9 d。处理完毕后, 取出种子在室温下通风晾至初始重量备用。1.2.2 种子形态观察及老化对种子发芽的影响
对杂交种及亲本进行种子外表饱满度形态观察, 通过室内发芽纸发芽试验对老化0、5 d的BS1453/ 11GF5135及BS1453、11GF5135种子发芽情况进行观察, 发芽第4天统计发芽率、根长、芽长。发芽试验设置3次生物学重复, 每个重复50粒种子。
1.2.3 酶活性的测定 用索莱宝APX酶活性测定试剂盒测定不同老化时间下BS1453/11GF5135及BS1453、11GF5135种子中的APX酶活性, 分别进行3次生物学重复。
1.3 APX基因克隆与序列分析
以拟南芥APX3蛋白序列(GenBank登录号为AT4G350001)为探针, 对小麦蛋白数据库进行本地BLAST, 获得小麦APX基因(ID: 100125717)。提取小麦APX的CDS序列, 利用Primer 5.0软件设计CDS全长克隆引物, 由金维智(北京)合成(表1)。以BS1453/11GF5135种子的cDNA为模板进行PCR扩增, PCR扩增体系20 μL: 2×Easy Taq SuperMix10 μL, 上下游引物(10 μmol L-1)各1 μL, cDNA模板2 μL, ddH2O 6 μL, 扩增程序: 94℃ 5 min; 94℃ 30 s, 71℃ 45 s, 72℃ 1 min, 42个循环; 72℃ 10 min。对扩增产物进行琼脂糖凝胶电泳检测, 回收目的条带, 连接到16318hGFP载体上, 连接产物转入DH5α感受态细胞, 经菌落PCR验证后, 各挑选7个阳性克隆子进行测序, 测序和引物合成均由金维智(北京)公司完成。Table 1
表1
表1TaAPX基因克隆及qPCR引物序列
Table 1
| 引物名称 Primer name | 引物序列 Primer sequence (5°-3°) | 退火温度 Annealing temperature (℃) | 引物用途 Primer purpose |
|---|---|---|---|
| APX-F | TATCTCTAGAGGATCCATGGCGGCTCCGGT | 71.37 | 扩增基因引物序列 Amplified gene primer sequences |
| APX-R | TGCTCACCATGGATCCTTACTTGCTCCTCTTGGAAGCCTCG | 72.27 | |
| APX-qF | CAACCGTTGAGTTCATCCCTG | 57.8 | 荧光定量PCR引物序列 RT-qPCR primer sequences |
| APX-qR | TGGTGCGCCTTTCTTAGCAT | 60 | |
| 18S rRNA-qF | CGCGCTACGGCTTTGACCTA | 56 | 内参引物序列 Internal reference primer sequences |
| 18S rRNA-qR | CGGCAGATTCCCACGCGTTACG | 58 | |
| miR396-qF | CGGTCTTCCACAGCTTTCTT | 75 | 荧光定量PCR引物序列 RT-qPCR primer sequences |
| miR396-qR | GTGCAGGGTCCGAGGT | 76 | |
| U6snRNA-qF | CAAGGATGACACGCAAATTCG | 80 | 内参引物序列 Internal reference primer sequences |
| U6snRNA-qR | GTGCAGGGTCCGAGGT | 82.5 |
新窗口打开|下载CSV
1.4 生物信息学分析
利用在线分析工具ExPASy ProtParam分析TaAPX基因编码蛋白的理化性质; 利用ProtScale分析TaAPX基因编码蛋白的亲疏水性; 利用SignalP 4.1 Server分析TaAPX基因编码的蛋白是否为信号肽蛋白; 利用TargetP 1.1 Server对TaAPX基因编码蛋白的亚细胞定位进行分析。TaAPX蛋白的二级结构和三级结构分别利用SOPMA和SWISS-MODEL进行预测; 利用NCBI蛋白比对工具预测TaAPX蛋白的保守结构域; 利用DNAMAN (ver.6.0)软件将TaAPX基因编码的蛋白与其他物种的APX基因编码的蛋白进行多序列比对; 采用MEGA 6.0软件, 依照邻位归并法(NJ)构建系统进化树; 利用PsRNATarget在线网站(http://plantgrn.noble.org/psRNATarget/analysis)预测与TaAPX基因互作的miRNAs; 利用在线蛋白互作预测网站STRING预测可能与TaAPX蛋白互作的蛋白。1.5 TaAPX基因表达模式的分析
设计TaAPX基因特异性引物APX-F和APX-R并进行qPCR扩增(表1), 反应程序: 95℃ 30 s, 95℃ 5 s, 60℃ 30 s, 45个循环; 95℃ 15 s, 60℃ 1 min, 95℃ 15 s; 60℃时收集荧光信号, 设置3次生物学重复。基因表达量采用相对定量法2-ΔΔCt法进行分析, 选用18S rRNA作为qPCR内参基因[12], 计算杂交小麦及其父母本在不同老化时间下TaAPX基因的相对表达量。1.6 miRNA与靶基因qPCR验证
利用miRNA的提取试剂盒(康为世纪, CW0627S)提取miRNA, 并进行cDNA合成。miRNA qPCR分析使用miRNA 荧光定量PCR检测试剂盒(康为世纪, CW2142)进行。应用程序: 95℃ 30 s, 95℃ 5 s, 60℃ 30 s, 45个循环。3次生物学重复, U6snRNA作为miRNA qPCR分析的内参, 利用2-ΔΔCt法进行表达定量分析。2 结果与分析
2.1 种子形态观察及老化对种子发芽的影响
本研究中以光温敏雄性不育系BS1453 (母本), 常规种11GF5135 (父本)及杂交种BS1453/11GF5135为研究对象, 通过对种子外观进行观察, 发现母本BS1453种皮皱瘪, 不饱满, 种皮颜色偏红; 父本种皮颜色为白色, 种子比较饱满; 杂交种BS1453/11GF5135种子饱满, 种皮颜色较父本深(图1)。发芽率统计发现, 杂交种老化前后的发芽率分别为90%和35%, 父本老化前后的发芽率分别为85%和25%, 母本老化前后的发芽率分别为82%和28%, 表明杂交种的发芽率高于其亲本。用发芽纸进行室内发芽实验, 在发芽第四天统计根长和芽长(图2-A), 发现老化前后杂交种和父母本的发芽率差异极显著(P<0.01), 根长差异显著(P<0.05) (图2-B)。图1
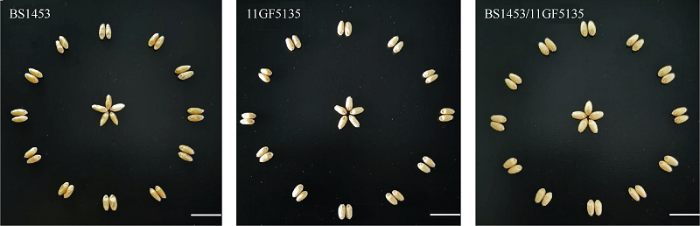 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1亲本及杂交种籽粒形态
Fig. 1Grain morphology of parents and hybrids
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2人工老化前后种子萌发表型和发芽统计
A: 老化处理和未老化处理的杂交种及父母本种子的萌发表型。B: 人工老化处理对杂交种及父母本的发芽率、根长、芽长的影响。发芽率在(P<0.01)时差异极显著, 用**表示; 根长在(P < 0.05)时差异显著, 用*表示; 芽长差异不显著。0~5 d: 老化处理的天数。
Fig. 2Seed germination phenotype and germination statistics before and after artificial aging
A: germination phenotypes of artificially aged and untreated seeds of hybrid and parents. B: effects of artificial aging on germination rate, root length and shoot length of hybrids and parents. ** and * indicate significant differences at the 0.01 and 0.05 probability levels, respectively. 0-5 d: Day(s) of aging treatment.
2.2 小麦APX基因的鉴定和序列分析
为了进一步研究杂交种高活力的机理, 以种子活力相关的小麦APX基因为目的基因, 以BS1453/11GF5135种子的cDNA为模板, 进行PCR扩增, 得到一条约850 bp左右的明亮条带(图3), 与预期长度一致。切胶回收并测序, 测序结果显示, TaAPX基因包含一个长度为832 bp的开放阅读框, 编码277个氨基酸。通过氨基酸序列多重比对分析后进一步发现, 杂交小麦APX基因所编码蛋白的氨基酸序列与小麦A、B和D基因组上的序列相同, 与普通小麦、大麦、粗山羊草的相似性达到90%以上(图4), 并且杂交种和父母本之间不存在序列变异(图5)。图3
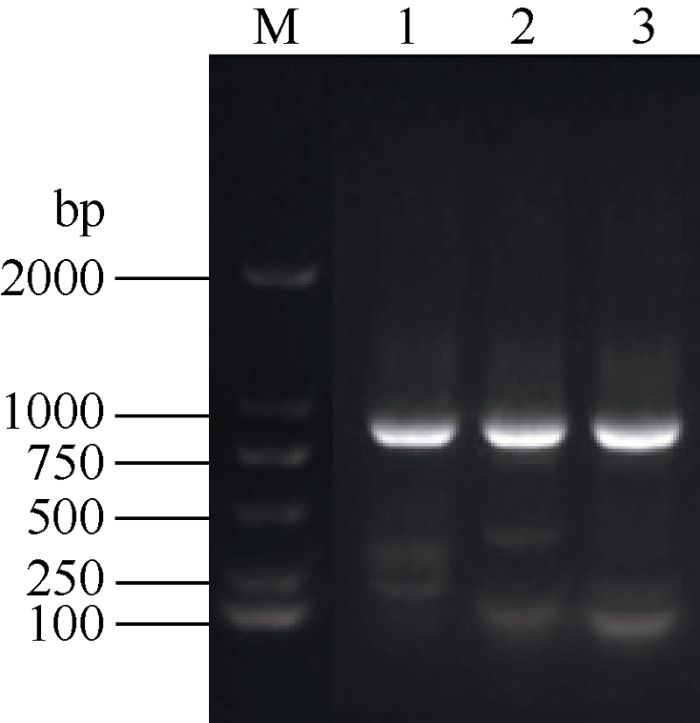 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3TaAPX基因的扩增产物
M: DL2000; 1: BS1453扩增产物; 2: 11GF5135扩增产物; 3: BS1453/11GF5135扩增产物。
Fig. 3Amplification products of TaAPX gene
M: DL2000 DNA marker; 1: amplified product of BS1453; 2: amplified product of 11GF5135; 3: amplified product of BS1453/ 11GF5135.
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4TaAPX氨基酸多序列比对分析
Fig. 4Multiple alignment analysis of TaAPX amino acid sequences
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5父母本和杂交种CDS序列比对分析
Fig. 5CDS alignment comparative analysis of parents and hybrid
2.3 TaAPX基因编码蛋白的特征分析
运用ExPASy ProtParam对TaAPX蛋白进行理化性质分析, 结果表明TaAPX蛋白的分子式为C1342H2127N375O402S5, 原子总数为4251, 相对分子质量为30.1 kD, 理论等电点(pI)为6.67, 由20种氨基酸组成, 其中Ala和Leu含量最高分别为12.6%和10.5%, 酸性氨基酸(Asp+Glu)为13.7%, 碱性氨基酸(Arg+Lys)为13.4%, 不稳定系数为29.86, 脂肪指数为88.48, 总平均疏水性为-0.241, 表明该蛋白是一个稳定亲水的中性蛋白。TaAPX蛋白的亚细胞定位及信号肽预测结果表明, TaAPX定位于细胞质且为非信号肽蛋白, 该蛋白由α-螺旋(41.58%), β-折叠(7.22%), 无规则卷曲(40.89%), 延伸链(10.31%)组成。其三级结构主要由螺旋和卷曲组成(图6-A)。利用NCBI进一步对TaAPX蛋白进行保守结构域分析发现, TaAPX蛋白具有典型的APX蛋白保守结构域, 隶属于PLN02608超家族。利用NetPhos对TaAPX蛋白进行磷酸化位点分析, 结果表明该蛋白的氨基酸序列上存在18个潜在的磷酸化位点, 包括9个丝氨酸(Ser)、4个苏氨酸(Thr)和5个酪氨酸(Tyr)位点(图6-B)。PROSITE scan数据库检测到的TaAPX蛋白序列中存在3个位点, 其中功能位点位于第40个氨基酸, 其他位点分别位于第36、162个氨基酸, TaAPX蛋白过氧化物酶区域在31~258氨基酸之间。图6
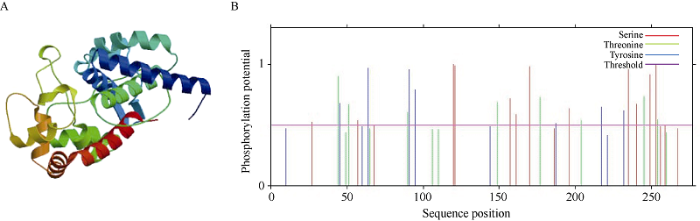 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6TaAPX蛋白的三级结构及磷酸化位点预测
A: TaAPX蛋白的三级结构; B: TaAPX蛋白磷酸化位点的预测。
Fig. 6Prediction of tertiary structure and phosphorylation site of TaAPX protein
A: tertiary structure of TaAPX protein; B: phosphorylation site prediction of TaAPX protein.
2.4 TaAPX基因编码蛋白的进化树分析
为进一步了解TaAPX基因的生物学功能, 采用MEGA6.0软件构建了植物APX蛋白氨基酸序列的系统进化树(图7)。将二系小麦杂交种APX蛋白序列与小麦(Triticum aestivum L.)、粗山羊草(Aegilops tauschii)、玉米(Zea mays L.)、水稻(Oryza sativa L.)、大麦(Hordeum vulgare L.)、高粱(Sorghum bicolor L.)和拟南芥(Arabidopsis thaliana) 7个物种的APX蛋白序列进行进化分析, 结果发现该蛋白序列与粗山羊草(Aegilops tauschii)的蛋白序列相似性为95.19%, 与印度小麦Raj3765 (Indian wheat cv. Raj 3765) (TaAPX-AGG23553.1)的蛋白序列相似性为94.5%, 与硬粒圆锥小麦(TaAPX-VAI74770.1)的蛋白序列相似性为94.16%, 与普通小麦(TaAPX-ABQ53157.1)的蛋白序列相似性为92.78%, 与大麦(HvAPX-AXF50251.1)的蛋白序列相似性为92.44%, 与拟南芥(AtAPX- OAO98886.1)的蛋白序列相似性为73.54%, 与玉米(ZmAPX-ACO90192.1)的蛋白序列相似性为55%, 与高粱(SbAPX-002463451.1)的蛋白序列相似性为57.41%, 与水稻(OsAPX-015630498.1)的蛋白序列相似性为56.79%, 表明, 二系小麦杂交种与小麦野生亲缘种粗山羊草(Aegilops tauschii)的亲缘关系最近, 都属于PLN02608超家族, 具有保守的结构域。图7
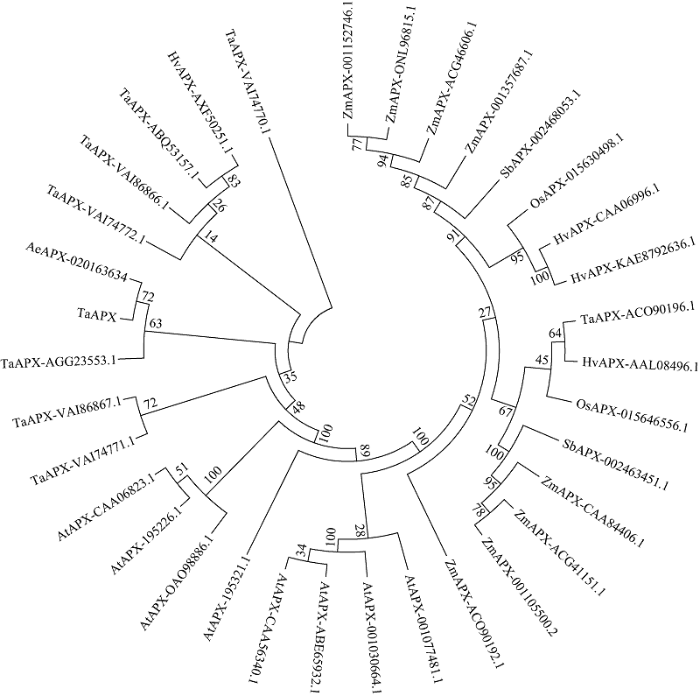 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7TaAPX蛋白系统进化分析
Fig. 7Phylogenetic analysis of TaAPX
2.5 TaAPX基因与miRNA互作的预测分析及与TaAPX蛋白互作蛋白的预测分析
前期研究发现miRNA对植物种子活力有一定的调控功能, 本研究中通过预测与TaAPX基因互作的miRNAs (图8-A), 结果发现与TaAPX互作的已知的miRNAs主要包括拟南芥(aly-miR396b-3p)、(ath-miR396b-3p)、芜菁(bra-miR396-3p)、大豆(gma-miR396b-3p)、(gma-miR396k-3p)、蒺藜状苜蓿(mtr-miR396a-3p)、刺菜蓟(cca-miR396a-3p)、杨树(ptc-miR396e-3p), 其非配对能UPE (unpairedenergy非配对能值越低, 蛋白和miRNA互作性越强)值为15.82; 水稻(osa-miR444a-3p.2、osa-miR444d.2、osa-miR444e、osa-miR444f)、大麦(hvu-miR444b)、蔗糖(ssp-miR444a、ssp-miR444b.2)二穗短柄草(bdi-miR444b) UPE值为22.175; 杨树(ptc-miR169aa)、木薯(mes-miR169m、mes-miR169l)、大豆(gma-miR169k、gma-miR169l-5p)、番木瓜(cpa-miR169), UPE值为19.968; 水稻(osa-miR167a-3p)、玉米(zma-miRa-3p), UPE值为19.365; 二穗短柄草(bdi-miR166h-5p)、水稻(osa-miR166i-5p), UPE值为17.618, 其中miR396与TaAPX的互作性最高, TaAPX基因与miR396的互作位置在TaAPX基因序列552~571碱基的位置(图8-D), 表明TaAPX基因可能主要受miR396的调控。miRNA具有发卡结构(图8-B), 在转录调控中作为负调控因子, 通过降解靶基因达到调控基因表达的目的。图8
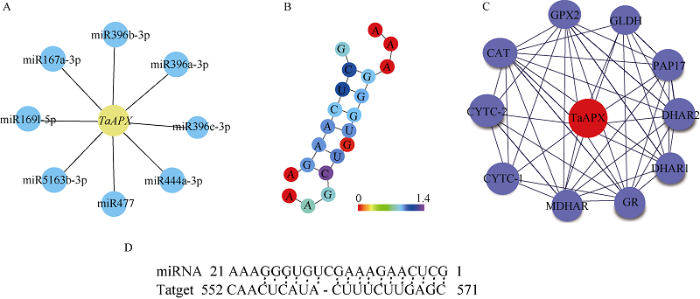 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8TaAPX基因与miRNA和蛋白的互作模型及互作位点
A: TaAPX基因与miRNA的互作图; B: miRNA的发卡结构图; C: 与TaAPX蛋白互作的蛋白的网络模型; D: TaAPX基因与miR396的互作位点。
Fig. 8Interaction model and site of TaAPX gene with miRNA and protein
A: interaction mapping between TaAPX gene and miRNA; B: the hairpin structure diagram of miRNA; C: network model of proteins interacting with TaAPX proteins; D: the interaction site between TaAPX gene and miR396.
为进一步了解TaAPX基因的功能, 本研究利用STRING对TaAPX蛋白潜在的互作关系进行预测分析, 结果表明APX3可能与谷胱甘肽过氧化酶(Glutathione Peroxidase 2, GPX2)、谷氨酸脱氢酶(Glutamate dehydrogenase, GLDH)、编码过氧化物酶体的过氧化氢酶(Catalase, CAT)、传递电子到细胞色素氧化酶复合物的电子载体蛋白(Cytc oxidasec, CYTC-1、CYTC-2)催化抗坏血酸合成的还原酶(MDHAR)、谷胱甘肽还原酶(glutathione reductase, GR)等蛋白互作(图8-C)。
2.6 不同老化时间下亲本和子代种子内TaAPX基因和miR396表达模式分析及酶活性分析
为了进一步研究不同老化时间对种子的影响, 设置0、3、5、7和9 d老化梯度, 利用RT-qPCR对5个不同时间老化处理后的杂交种和亲本种子的胚进行TaAPX基因表达模式分析, 结果发现2个亲本BS1453、11GF5135和杂交种BS1453/11GF5135种子内TaAPX基因的表达均呈先升后降的趋势。在亲本种子中, TaAPX基因的表达量在老化第3天上升并达到峰值, 随后开始下降, 老化第5天TaAPX基因的表达量低于第3天, 老化第7天TaAPX基因的表达量低于第5天, 老化第9天的表达量达到最低(图9-A, B); 而杂交种BS1453/11GF5135种子内TaAPX基因的表达量在老化7 d内均呈显著上升趋势, 老化第5天TaAPX基因的表达量达到峰值, 老化第7天TaAPX基因的表达量低于第5天, 但是仍然高于对照, 老化第9天降到最低(图9-C)。图9
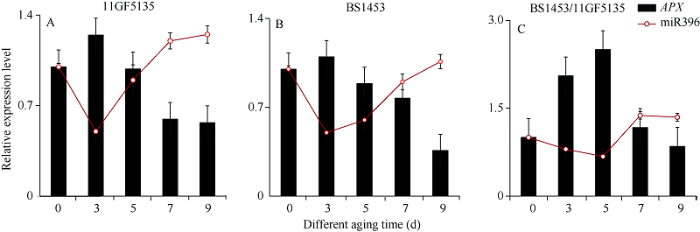 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9不同老化时间下TaAPX基因和miR396在杂交种和亲本种子胚中的表达模式分析
A: 不同老化条件下, TaAPX基因和miR396在父本种子胚中的表达; B: 不同老化条件下, TaAPX基因和miR396在母本种子胚中的表达; C: 不同老化条件下, TaAPX基因和miR396在杂交种种子胚中的表达。0~9 d: 老化处理的天数。
Fig. 9Expression pattern analysis of TaAPX gene and miR396 in hybrid and parental seed embryos at different aging time
A: the expression of TaAPX gene and miR396 in embryo of male seeds, under different aging conditions; B: the expression of TaAPX gene and miR396 in embryo of female seeds, under different aging conditions; C: the expression of TaAPX gene and miR396 in embryo of hybrid seeds, under different aging conditions. 0-9 d: day(s) of aging treatment.
为了进一步研究TaAPX基因在种子活力中的调控作用, 本研究在相同的老化条件下测定杂交种和父母本种子内的APX酶活性变化, 结果发现, 两个亲本BS1453、11GF5135种子内的APX酶活性呈下降趋势, 但是老化第5天的APX酶活性高于老化第3天, 随后开始下降, 老化第9天APX酶活性最低(图10-A, B); 而杂交种种子内酶活性变化基本与TaAPX基因的表达一致。随着老化时间的延长杂交种BS1453/11GF5135种子内的APX酶活性在老化第3天出现短暂的下降, 第5天开始上升, 老化第7天APX酶活性高于第5天达到峰值, 老化第9天开始下降(图10-C)。
图10
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10不同老化时间下杂交种和亲本内APX酶活性分析
A: 不同老化条件下, APX酶活在父本种子胚中的变化; B: 不同老化条件下, APX酶活在母本种子胚中的变化; C: 不同老化条件下, APX酶活在杂交种种子胚中的变化。0~9 d: 老化处理的天数。
Fig. 10Analysis of APX enzyme activity in hybrids and parents at different aging times
A: changes of APX enzyme activity in embryo of male seeds, under different aging conditions; B: changes of APX enzyme activity in embryo of female seeds, under different aging conditions; C: changes of APX enzyme activity in embryo of hybrid seeds, under different aging conditions. 0-9 d: day(s) of aging treatment.
为了解miRNA对种子活力的调控方式, 本研究对预测到的互作关系比较强烈的miR396, 在不同老化程度的父母本及杂交种中进行表达模式分析, 同时分析miRNA对靶基因TaAPX的调控关系。结果发现miR396与TaAPX基因表达趋势相反, 两者呈拮抗作用(图9)。
3 讨论
人工老化前后, BS型杂交小麦BS1453/ 11GF5135均显示出优于亲本的种子活力(图2), 为研究种子活力相关基因TaAPX在杂交小麦中的调控功能, 本研究以BS型二系小麦杂交种为研究对象, 克隆TaAPX基因, 通过生物信息学分析预测与TaAPX基因互作的miRNAs, 发现TaAPX可能与miR396、miR444、miR169、miR167、miR166互作, 其中互作效应最强的miRNA为miR396。有报道发现, 这些miRNA与植物抗逆相关, 如在大豆中miR396家族调控GmGRF家族基因, 在高温胁迫下大豆中Gma-miR396在叶片中上调表达, 而在根中下调表达, 有利于抵抗高温胁迫[13]; 在高温胁迫下, 将大豆gma-miR169c基因转入拟南芥发现拟南芥叶片加快萎蔫, 根生长减慢等症状, 表明gma-miR169c在高温胁迫下起负调控作用[14]; 在盐胁迫条件下, 将黄瓜miR166基因转入拟南芥使拟南芥发芽率、根长等各项指标都比野生型高, 过表达miR166可以使拟南芥更好地抵抗高温胁迫[15]。miRNA主要是通过对靶基因的负调控来进一步调控生理生化反应[16]。有研究表明miRNA通过与靶基因互作, 对种子发育及活力有一定的调控作用[17]。在水稻ZR02种子内osa-miR164c的下调和osamiR168a的上调有利于维持种子活力, 且osa-miR164c和osa-miR168a通过调节细胞膜的完整性来影响种子活力; miR167A作为一个母体效应基因参与胚胎及种子发育, 当种皮的基因型为mir167a+/+和mir167a+/-时, 胚胎正常发育, 不会影响种子的发育, 当种皮的基因型为mir167a-/-时, 胚胎发育出现缺陷, 从而影响种子的发育[18]。本研究中发现, miR396在3个品种中, 表达量与APX基因随着老化时间的变化而呈相反的趋势(图9), 表明miR396可能是通过负调控APX发挥功能。因此, 这些预测的miRNA可能通过调控TaAPX基因来提高植物的抗逆能力以及杂交种的种子活力, 其具体功能有待进一步的研究验证。进一步的试验发现父母本和杂交种种子胚内TaAPX基因的表达量随着老化天数的增加是先上升后下降的趋势(图9-A, B), 但是在杂交种子中TaAPX基因的表达量在3、5和7 d均高于对照, 而其父母本种子中TaAPX基因的表达量只在3 d高于对照, 说明杂交种子的抗老化能力要高于其亲本(图9-C)。可能是在老化初期, 由于外界环境的变化激发了种子的抗逆应激反应, 从而激发APX基因的上调表达, 以保护细胞免受氧化损伤。有研究表明在42℃热应激条件下, 印度小麦Raj 3765种子内TaAPX基因的表达量上调了203倍来抵抗高温胁迫[19], 因此推测TaAPX基因在抵抗高温胁迫维持种子活力方面具有重要作用。
研究表明正常的种子内的活性氧的产生与清除之间是动态平衡的[20], 经过老化处理的种子会破坏这种平衡, 造成细胞内H2O2的过量累积, 从而影响种子活力。但是燕麦种子随老化时间的延长, APX活性呈先上升, 随后下降的趋势; 高阳茅种子中APX活性随着老化程度的加深而升高[21]。本实验酶活性分析结果表明亲本种子随着老化天数的增加, 其APX酶活性总体是呈下降趋势的, 但是在杂交种中APX的酶活变化与这一结论正好相反, 从图中可以看出在老化3 d时APX酶活性出现短暂地下降, 但是在老化5 d和7 d呈上升趋势, 在老化9 d开始下降(图10-C)。说明长时间老化处理对BS型小麦杂交种子内APX酶活的影响大于对其亲本的影响, 即老化处理会增强APX酶清除体内H2O2的能力, 以保护种子免受损伤。另外通过蛋白互作预测研究表明, TaAPX基因对种子活力的调控作用是一个复杂的酶活反应过程, 是多种酶共同作用的结果, 具体的APX酶反应程序为: 2APX (抗坏血酸过氧化物酶)+2AsA (抗坏血酸)+H2O2 (过氧化氢)→2MDHA (单脱氢抗坏血酸)+H2O; GR (谷胱甘肽还原酶)也是一种抗氧化酶, 通过NADPH可将GSSG (氧化型谷胱甘肽)还原为GSH (还原型谷胱甘肽)[22]; CAT和APX一样作为一种抗氧化酶在维持植物细胞稳态方面具有重要作用[23]。
4 结论
BS型二系小麦杂交种BS1453/11GF5135老化前后发芽率及根长、芽长都高于亲本。研究发现APX酶在维持BS型小麦杂交种较高种子活力方面具有重要作用, 短时间的老化处理会提高BS型小麦的抗老化能力, 表明BS型小麦杂交种的抗老化能力要高于其亲本。研究通过对与TaAPX互作的miRNA的表达量进行分析, 发现该基因可能通过miRNA负调控来发挥功能。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/s00299-007-0395-7URLPMID:17571267 [本文引用: 1]

In order to determine the different roles of rice (Oryza sativa L.) cytosolic ascorbate peroxidases (OsAPXa and OsAPXb, GenBank accession nos. D45423 and AB053297, respectively) under salt stress, transgenic Arabidopsis plants over-expressing OsAPXa or OsAPXb were generated, and they all exhibited increased tolerance to salt stress compared to wild-type plants. Moreover, transgenic lines over-expressing OsAPXb showed higher salt tolerance than OsAPXa transgenic lines as indicated by root length and total chlorophyll content. In addition to ascorbate peroxidase (APX) activity, antioxidant enzyme activities of catalase (CAT), superoxide dismutase (SOD) and glutathione reductase (GR), which are also involved in the salt tolerance process, and the content of H2O2 were also assayed in both transgenic and wild-type plants. The results showed that the overproduction of OsAPXb enhanced and maintained APX activity to a much higher degree than OsAPXa in transgenic Arabidopsis during treatment with different concentrations of NaCl, enhanced the active oxygen scavenging system, and protected plants from salt stress by equilibrating H2O2 metabolism. Our findings suggest that the rice cytosolic OsAPXb gene has a more functional role than OsAPXa in the improvement of salt tolerance in transgenic plants.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11033-019-04648-3URLPMID:30706357 [本文引用: 1]
Hydrogen peroxide (H2O2) is known to accumulate in plants during abiotic stress conditions and also acts as a signalling molecule. In this study, Arabidopsis thaliana transgenics overexpressing cytosolic CuZn-superoxide dismutase (PaSOD) from poly-extremophile high-altitude Himalayan plant Potentilla atrosanguinea, cytosolic ascorbate peroxidase (RaAPX) from Rheum australe and dual transgenics overexpressing both the genes were developed and analyzed under salt stress. In comparison to wild-type (WT) or single transgenics, the performance of dual transgenics under salt stress was better with higher biomass accumulation and cellulose content. We identified genes involved in cell wall biosynthesis, including nine cellulose synthases (CesA), seven cellulose synthase-like proteins together with other wall-related genes. RNA-seq analysis and qPCR revealed differential regulation of genes (CesA 4, 7 and 8) and transcription factors (MYB46 and 83) involved in secondary cell wall cellulose biosynthesis, amongst which most of the cellulose biosynthesis gene showed upregulation in single (PaSOD line) and dual transgenics at 100 mM salt stress. A positive correlation between cellulose content and H2O2 accumulation was observed in these transgenic lines. Further, cellulose content was 1.6-2 folds significantly higher in PaSOD and dual transgenic lines, 1.4 fold higher in RaAPX lines as compared to WT plants under stress conditions. Additionally, transgenics overexpressing PaSOD and RaAPX also displayed higher amounts of phenolics as compared to WT. The novelty of present study is that H2O2 apart from its role in signalling, it also provides mechanical strength to plants and aid in plant biomass production during salt stress by transcriptional activation of cellulose biosynthesis pathway. This modulation of the cellulose biosynthetic machinery in plants has the potential to provide insight into plant growth, morphogenesis and to create plants with enhanced cellulose content for biofuel use.
[本文引用: 1]
[本文引用: 1]
URLPMID:30170113 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:31203895 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}