 ,1,*, ����־1, ������1, �2, ĸΰ��3, ������3, ��Τ��3, ���ҽ�1, ������1, ������1, ��ϲ��1, ����2, ����1, ����1, ���Ļ�,2,*
,1,*, ����־1, ������1, �2, ĸΰ��3, ������3, ��Τ��3, ���ҽ�1, ������1, ������1, ��ϲ��1, ����2, ����1, ����1, ���Ļ�,2,*Detection of QTLs controlling cold tolerance at bud bursting stage by using a high-density SNP linkage map in japonica rice
JIANG Shu-Kun,1,*, WANG Li-Zhi1, YANG Xian-Li1, LI Bo2, MU Wei-Jie3, DONG Shi-Chen3, CHE Wei-Cai3, LI Zhong-Jie1, CHI Li-Yong1, LI Ming-Xian1, ZHANG Xi-Juan1, JIANG Hui2, LI Rui1, ZHAO Qian1, LI Wen-Hua,2,*ͨѶ����:
�ո�����:2019-12-08��������:2020-03-24�����������:2020-08-12
| ��������: |
Received:2019-12-08Accepted:2020-03-24Online:2020-08-12
| Fund supported: |
����� About authors
E-mail:sk_jiang@126.com, E-mail��sk_jiang@126.com

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (3422KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
������, ����־, ������, �, ĸΰ��, ������, ��Τ��, ���ҽ�, ������, ������, ��ϲ��, ����, ����, ����, ���Ļ�. ���ڸ��ܶ�SNP�Ŵ�ͼ�ľ���ѿ���͵���QTL����[J]. ����ѧ��, 2020, 46(8): 1174-1184. doi:10.3724/SP.J.1006.2020.92066
JIANG Shu-Kun, WANG Li-Zhi, YANG Xian-Li, LI Bo, MU Wei-Jie, DONG Shi-Chen, CHE Wei-Cai, LI Zhong-Jie, CHI Li-Yong, LI Ming-Xian, ZHANG Xi-Juan, JIANG Hui, LI Rui, ZHAO Qian, LI Wen-Hua.
��Ϊ����������Ҫ����ʳ����֮һ, ˮ����Դ���ȴ������ȴ�, ���С���������, �Ե��¸�������[1]���亦��ȫ������Ȼ�ֺ�, ������������Ҿ��з���, ���Ĵ������ϲ����ձ�������������������Ϊ��, ���ģ�亦�ķ����������ʮ�ϰ��ڹ�����ʳ����ʧ[2,3,4,5,6]���ҹ��������������е����亦����, �ڻ��Ϻͳ��������ε�˫������, �����ġ������������^�ġ���¶�硱������������亦[7]����������ƽ��3~4��ͻ�����һ�δ�Χ���亦, ���ˮ����������, Σ����ʳ��ȫ[8]��ˮ���ȷ��ڡ�ѿ�ڡ����ڡ������ڡ������ںཬ�����ܵ����亦Ӱ��[9,10], �������ֱ�ӻ��ӹ�ϵ��20����90�������, �ҹ���ˮ�����������˽ϴ�ı仯��һ����, ˮ������Ŀ���������ͳɱ�����߾���Ч��ת��; ��һ����, �����ҹ����õĿ��ٷ�չ, ũ����Ҫ�Ͷ��������ת��, ũ���Ͷ�����ȱ�ѳ�Ϊˮ��������չ����Ҫ�������ء��������ı�����, ˮ��ֱ������ʡʱ��ʡ���ͽ�Լ�ɱ�, ���ũ��Ļ�ӭ, ��Ѹ���ƹ㡣�ݲ���ȫͳ��, ˮ��ֱ������ڰ���ʡ�Ѵ�56.7����, ����ʡΪ49����, ����ʡΪ43.3����, ����ʡΪ69.3����, �㽭ʡΪ37.3����, �㶫ʡΪ10����, ������ʡ��ֱ�������Ϊ40����[11,12]������˫�������絾ֱ���Ͷ����ߺ������ĵ�����ֱ������������������������, ���ȱ���¢, �������ؼ���[9,10,11,12]�����Ҫ������ֱ����ˮ�������н�ǿ��ѿ���͵���������������������һֱ����������Ϊ��, ѿ���͵����������ڲ���Ϊ����Ŀ��, ��ʹ����߲������ʵ�ˮ��Ʒ������ѿ���͵�������������������ֱ����, ������ֱ���������������, ˮ��ѿ�������������������������в��ɺ��ӵ���Ҫ��״��
�Դ�20����30���, ������Ա��ʼ�о�ˮ���亦����, �����亦����������ѧԭ��ˮ��������������ơ��Ŵ������ͻ����¡�ȷ�������˴������о��ɹ�[1,13-19]�������ȷ��ڡ����ں��������亦���, ������ˮ��ѿ�������Է�����о����Բ���, Ŀǰ��Ҫ��չ����ѿ����������QTL��λ�о����ϳ��ܵ�[20]�����Ͼ�11�Ͱ�������DHȺ��, ��7��Ⱦɫ���ϼ�����1��ѿ���亦QTL��Zhang��[21]����Lemont������������Խ�ϵȺ��, ��3�š�7�ź�11��Ⱦɫ���ϼ�3������ˮ��ѿ�������Ե�QTL����������[22]��������23�ͼ���1�ŵ�F2:3Ⱥ��, ��2�š�4�ź�7��Ⱦɫ���ϼ�3��ѿ������QTL�������[23]����Lemont������������Խ�ϵȺ��, ��1�š�3�š�7�ź�11��Ⱦɫ���ϼ�4������ˮ��ѿ�������Ե�QTL����¶ϼ��[24]����Asominori��IR24�������Խ�ϵȺ�����Ӧ��Ⱦɫ��Ƭ���û�ϵ, ��5�ź�12��Ⱦɫ���ϼ�3��ѿ������QTL����ӭ����[25]����Խ���Kasalath�Ļؽ������Խ�ϵȺ��, ��4�š�6�ź�11��Ⱦɫ���ϼ�4��ѿ������QTL���־���[26]�������̵�9311Ϊ���塢�����ձ���Ϊ�����Ⱦɫ��Ƭ���û�ϵ, ��5�ź�7��Ⱦɫ���ϼ�����4��ѿ������QTL��Baruah��[27]�������ൾA58��Ұ����W107�������Խ�ϵȺ��, ��1�š�11�ź�12��Ⱦɫ���ϼ���3��ѿ������QTL��Ji��[28]����TN1�ʹ���06��DHȺ��, ��2�š�4�ź�11��Ⱦɫ���ϼ�����3��ѿ������QTL�����µ�[29]�����̵�9311Ϊ���塢�����ձ���Ϊ�����Ⱦɫ�嵥Ƭ���û�ϵ, �ڳ�1�ź�11��Ⱦɫ�������10��Ⱦɫ���ϼ���18��ѿ������QTL����������[30]���ÿ���131�Ͷ�ũ422�������Խ�ϵȺ��, ��4��Ⱦɫ���ϼ�1��ѿ������QTL��������[31]���ù�½��4��Ϊ����, �ձ���Ϊ�����Ⱦɫ��Ƭ���û�ϵ, ��1�š�6�š�8�š�9�ź�10��Ⱦɫ���ϼ�8��ѿ������QTL��Yang��[32]���û�����74Ϊ����, ����ռΪ�����Ⱦɫ��Ƭ���û�ϵ, ��5�ź�6��Ⱦɫ���ϼ�����2��ѿ������QTL��Zhang��[33]����249���̵�������Դ���Ͻ��5Kˮ��оƬ����ѿ�������GWAS����, ��1�š�3�š�4�š�5�š�6�š�10�ź�12��Ⱦɫ���Ϲ���13��ѿ�������������
��Ȼ���Ƕ�ѿ�������Խ����˽϶��QTL��������, ������ˮ����12��Ⱦɫ���Ͼ�����QTL�Ĵ���, ����������QTL��ΪЧQTL, ���о�ϸ��λ�ͺ����о��ѶȽϴ�ͬʱ, �����о����õ��Ŵ�Ⱥ������黷���ȷ���IJ�ͬ, �о����������ͬ, ����б�Ҫ������������ͬ���Ŵ�Ⱥ������ܶȵ��Ŵ�ͼ�������ѱ�����ѿ������QTL������֤, ������µ�ѿ������λ�㡣���о������������źڹȺ���ũ265�����������Խ�ϵȺ��, ����ز����������ĸ��ܶ�bin�Ŵ�ͼ��, �ھ����ѿ�������Ե�QTL, ����Ϊֱ������������������ṩ���õġ�������Դ��, ͬʱΪ����ˮ��ѿ��������Ŵ��ͷ��ӻ����ṩ�ο���
1 �����뷽��
1.1 �������
2004��, ��ѿ��������ǿ�����ϵط�����Ʒ���������źڹ�(LTH)Ϊĸ��, ѿ�����������ı���������Ʒ����ũ265 (SN265)Ϊ���������ӽ����, 2005��2013��ֱ�����������(2005��2006, 2008��2009)����������(2008)�ͺ�����������(2011��2015)����, ���õ�������(single seed descent, SSD)�״��Խ�11����ð���144����ϵ���ȶ��Ŵ������Խ�ϵȺ��(recombinant inbred lines, RILs)������Huang��[34]�������ز����ֶ�, ���á��������ڡ����������������źڹȺ���ũ265�������Խ�ϵȺ��bin�Ŵ�ͼ�ס���bin����ͼ������58,738�������ȵ�, ƽ��ÿ����ϵ405�������ȵ㡣���ղ�����2818������bin���(ͼ1-A), ��Щ��Ǹ����˴ֵ������ȵ�(ͼ1-B), ƽ��ÿ��Ⱦɫ���ϸ���235��bin���, �������ƽ��Ϊ128.8 kb[35]��
ͼ1
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1�������źڹ�-��ũ265�����Խ�ϵȺ�������ͼ��(A)�ͻ�����(B)
����ͼ��λ�û����ձ���ο�������(MSU Rice Genome Annotation Project Release 7), ��ɫ��ʾ��ũ265; ��ɫ��ʾ�������źڹȡ�
Fig. 1Physical map (A) and genotype (B) of Lijiangxintuanheigu-shennong 265 (LTH-SN265) RIL population
Physical position is based on MSU Rice Genome Annotation Project Release 7 sequence. Yellow: SN265 genotype; Blue: LTH genotype.
1.2 ѿ�����������۷���
�ױ��������źڹ�(LTH)����ũ265 (SN265)��144�������Խ�ϵȺ����ϴ�������Ӵ��ع��д���, ��������Ϊ�¶�5��C, ���ʪ��Ϊ10%������ѿ�������Բ��պ���ֲ��[36]�ķ���������, ÿ����ϵ3���ظ�, ����3�ε�ƽ��ֵ���к������������Ƚ����ӴӴ��ع���ȡ��, ��50��C���¸������ڸ��´���48 h, ʹ���ָ��ﲢ�����������ߡ�ÿ���ظ���ѡ������100��, ��0.1%���Ȼ�����Һ����10 min, ����ˮ��ϴ3~4��, ȥ����ˮϴ��3��, �ڵ�˫����ֽ��ֱ��9 cm��������, ����10 mLȥ����ˮ�������������, ��30��C����������(�Ϻ�һ��MGC-250)�ڴ�ѿ2~3 d��Ȼ��Ӻ�������ȡ��, ��ȥ����ˮ��ϴ1~2�Ρ��ӷ�ѿ�������о�����ѡѿ��Լ1~2 cm������50��, ��ֲ�ڱ߳�2 cm��Ӫ��������, ���������ȷ������ƺõ�Ӫ����������28��C����������(�Ϻ�һ��MGC-450HP)�ڷ���1 d��, ת������˹�������(�ձ�����ƽԭ��ĿԮ��˪����ʵ����)�ڴ���48 h, �¶�����Ϊ2��C��������, ���������ת�Ƶ��¶�Ϊ20~30��C, ��������ĵط�, ʹ��ָ�����, ÿ��������ˮ���ָ�10 d��, ������������״̬������ͼ2�ı��������������ָ�״̬��ֵ, ����Ϊ5��: �ָ�����ѿ�����ĸ�ֵΪ0; �ָ�����ѿֹͣ����, ���ֻ��εĸ�ֵΪ1; �ָ�����һ����������, Ҷɫ�ϵ�, �������dz������ĸ�ֵΪ2; �ָ�������������ǿ, Ҷɫ��, �����ϻ����ĸ�ֵΪ3; �ָ���Ҷɫ����, �ܹ����������ĸ�ֵΪ4�������ղ�ͬ������м�Ȩ�Ʒ֡�ѿ�����伶��=(0��0��������+1��1��������+2��2��������+3��3��������+4��4��������)/�����IJ�����������ͼ2
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2ˮ��ѿ���亦���۵ȼ���
Fig. 2Evaluation criteria for chilling injury at rice bud stage SN265: Shennong 265; LTH: Lijiangxintuanheigu.
1.3 ѿ�������Ե�QTL������QTLЧӦ����
����R/qtl��CIM��������QTL��λ, ����mqmpermutation��������������1000�ε�LOD��ֵ(��=0.05)ȷ��, ��ʵ����õ�LODֵ����LOD��ֵʱ, ����Ϊ�����δ���1��QTL, ����������ΪLOD��ֵ����1��LODֵ��λ������[37]��Ϊ�˽�һ����֤�����鶨λ�����ȷ��, ͬʱ����ɸѡ����QTL�ۼ�ЧӦ, ���á�ѡ����ͼ�����Խ���QTL��֤���ۼ�ЧӦ����[38]�����Ƚ�����ѿ�����������QTLλ����ݷ��ӱ�����ݽ��з���Ч����Ĺ̶�, �Դ�����������QTL�Ժ���������Ӱ��, Ȼ����RILsȺ����ɸѡĿ��QTL��������齻������, ������QTL������֤���ۼ�ЧӦ������
2 ��������
2.1 �ױ���ѿ�������ԱȽϼ�Ⱥ���ѿ�������Էֲ�
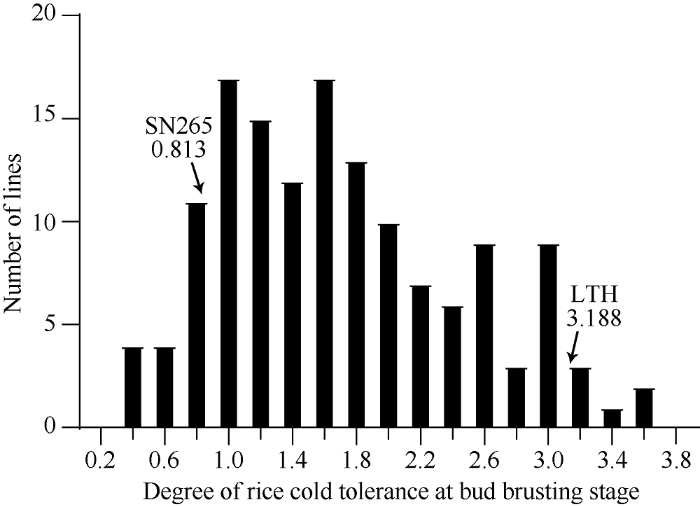
������ǰ�ڵ��о�����, �������źڹ�����ũ265�ڶ����ͬʱ�ڵ����������״�ϴ�����������, �������źڹȾ��и�������ۺ��������ԡ����о���, 2���ױ���2��C���´���2 d���ָ�10 d��, ��ũ265��ƽ��ѿ������ȼ���Ϊ0.813, ���������źڹȸߴ�3.188 (ͼ3��ͼ4), ����ﵽ������ˮƽ�������Խ�ϵȺ���ѿ�������Եȼ��ֲ���ΧΪ0.313~3.688, ƽ��ֵΪ1.699, ����Ϊ0.767�������ϳ��ֽ�����̬�ֲ�, �����ֳ���ǿ�ij�����(ͼ4)��ͼ3
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3�ױ��������źڹȺ���ũ265ѿ�ڲ����䴦��(A)���䴦����(B)�ıȽ�
Fig. 3Comparison of no cold treatment (A) and cold treatment (B) at bud bursting stage between LTH and SN265
ͼ4
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4�����Խ�ϵȺ���ѿ�������Էֲ�
Fig. 4Distribution of cold tolerance at bud bursting stage in LTH-SN265 RILs
2.2 Ⱥ��ѿ�������Ե�QTL�ֲ���ЧӦ
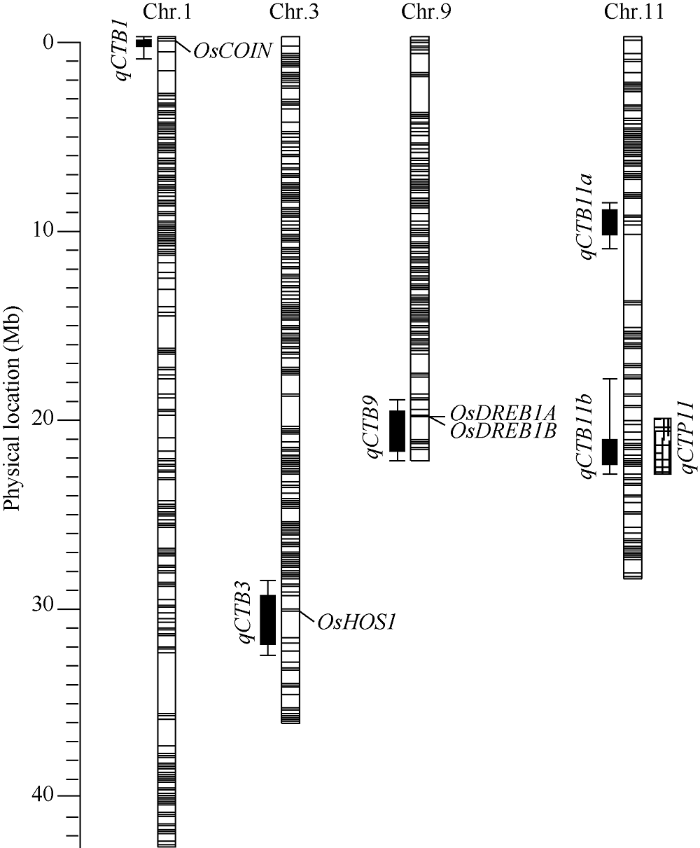
����R/qtl��������5������ˮ��ѿ�������Ե�QTL, �ֲ���ˮ����1�š�3�š�9�ź�11��Ⱦɫ����, ��ЩQTL����Ч��λ������������������źڹ�(��1��ͼ5)����������������ЧQTL��qCTB11b (LODֵΪ24.01), λ��11��Ⱦɫ�峤�۶˵�21.24 Mb~22.03 Mb֮��, �����С0.79 Mb���������������ʴ���15%����ЧQTL��qCTB9 (LODֵΪ5.36)��qCTB11a (LODֵΪ8.58), �ֱ�λ��ˮ����9Ⱦɫ����19.90 Mb~22.30 Mb֮���11��Ⱦɫ����8.53 Mb~9.93 Mb֮��, �����С�ֱ�Ϊ2.40 Mb��1.40 Mb������ЧQTL��qCTB1 (LODֵΪ3.26)��qCTB3 (LODֵΪ3.05), �ֱ�λ��ˮ��1��Ⱦɫ����0.43 Mb~0.93 Mb֮���3��Ⱦɫ����29.40 Mb~32.87 Mb֮��, �����С�ֱ�Ϊ0.50 Mb��0.61 Mb��û�ܴ���ũ265�м�ѿ�������Ե���Ч��λQTL��Table 1
��1
��1�����������źڹ�-��ũ265�������Խ�ϵȺ�����ѿ������QTL��Ϣ
Table 1
| ������״ ������ QTL | Ⱦɫ�� Chr. | ��ֵλ��a Peak pos.a | ��ֵ��� Peak marker | QTL����QTL interval | LODֵ LOD value | �������� Var. (%) | ���� ЧӦ Add. | ��Ч��λ ������Դ Positive allele | |
|---|---|---|---|---|---|---|---|---|---|
| ����ͼ�� Physical (Mb) | ��λ���� Location interval (Mb) | ||||||||
| qCTB1 | 1 | 0.73 | Bin01-002 | 0.43-0.93 | 0.50 | 3.26 | 8.20 | 0.22 | LTH |
| qCTB3 | 3 | 29.40 | Bin03-281 | 29.40-32.87 | 3.47 | 3.05 | 8.00 | 0.21 | LTH |
| qCTB9 | 9 | 21.45 | Bin09-185 | 19.90-22.30 | 2.40 | 5.36 | 16.60 | 0.34 | LTH |
| qCTB11a | 11 | 9.71 | Bin11-087 | 8.53-9.93 | 1.40 | 8.58 | 24.10 | 0.39 | LTH |
| qCTB11b | 11 | 21.80 | Bin11-141 | 21.24-22.03 | 0.79 | 24.01 | 53.50 | 0.64 | LTH |
�´��ڴ�|����CSV
ͼ5
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5����ѿ�������Ե�QTL��Ⱦɫ���ϵ�λ�÷ֲ�
OsCOIN: �����յ���пָ����[46]; OsHOS1: ������в�ȵ�E3��������ø[47]; OsDREB1A, OsDREB1B: �����յ���AP2/ EREBPת¼���ӻ���[48]; qCTP11: ѿ������QTL[27]��
Fig. 5Chromosome location of putative QTL for cold tolerance at bud bursting stage in LTH-SN265 RILs
OsCOIN: cold inducible zinc finger protein[46]; OsHOS1: a rice E3-Ubiquitin Ligase in the modulation of cold stress response[47]; OsDREB1A, OsDREB1B: cold inducible AP2/EREBP transcription factor gene[48]; qCTP11: a QTL for cold tolerance at bud stage[27].
2.3 ѿ��������QTL����֤���ۼ�ЧӦ����
���ǶԼ���ѿ������QTL��������֤����������ЩQTL�Ļ������ۼ�ЧӦ������, ������ѿ�������Ե�����QTL���÷��ӱ�Ǹ������з���Ч����Ĺ̶�, Ȼ���������Խ�ϵȺ����ɸѡĿ������(������QTL����)Ϊ��Ч��λ����ĵ���, ��������5��QTL������֤�����, �������ƵIJ���, ͨ����Ǹ���ѡ�����ͬQTL��ϵĵ���, ����QTL�Ļ����������ۼ�ЧӦ���ۡ��������κ�ѿ������QTL��ϵ��ѿ�ڵ��¿��Եȼ�Ϊ0.80 (ͼ6��ͼ7-A, X), ����1��QTL��ϵ��ѿ�ڵ��¿��Եȼ���1.03��2.33 (ͼ6��ͼ7-C~F), ����qCTB11b����ϵ������ߵ�ѿ�ڵ��¿��Եȼ�(ͼ6��ͼ7-G)������2��QTL��ϵ��ѿ�ڵ��¿��Եȼ���1.67��2.82 (ͼ6��ͼ7-H~M), ����qCTB11b��ϵ��ѿ�ڵ��¿��Եȼ��ձ�ϸ�(2.44~2.82), 2��QTL֮������������ϱ���Ϊ�ļ���ЧӦ, ����ЧӦ������������Ⱥ���ģ��Ӱ��, ��û�и��ǵ�ȫ����3��QTL�ۺ�����, ɸѡ����6�����ͺ���3��QTL��ϵ��ѿ�ڵ��¿��Եȼ���1.33��2.98 (ͼ6��ͼ7-N~S), ������QTL�ۺϵĹ�������, ����qCTB11b��ϵ��ѿ�ڵ��¿��Եȼ��ձ�ϸ�(2.52~2.98), 3��QTL֮�������������Ҳ����Ϊ�ļ���ЧӦ��4��QTL�ۺϵ���ϵ��3������, ������qCTB11a��qCTB11b, ���ǵ�ѿ�ڵ��¿��Եȼ���3.00��3.06 (ͼ6��ͼ7-T~V)������ͬʱ�ۺ�5��QTL����ϵ8��, ���ǵ�ѿ�ڵ��¿��Եȼ���2.94��3.50 (ͼ6��ͼ7-W)��ͨ����������, ����ȷ����ѿ���͵���QTL�������Խ�ϵȺ���оۺ�ʱ��Ҫ����Ϊ����ЧӦ��ͼ6
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6ѿ������QTL����֤�;ۺ�ЧӦ����ʹ�õ����鵥�������
��-��: ȱʧ��
Fig. 6Genotype of recombinant plant for validating cold tolerance QTL and analyzing pyramiding effect at bud busting stage
��-��: missing.
ͼ7
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ7����ѿ�������Ե�QTLЧӦ����LTH-SN265�����Խ�ϵȺ���еľۺ�Ч������
A: �ױ���ũ265 (SN265); B: �ױ��������źڹ�(LTH); C~G: qCTB1��qCTB3��qCTB9��qCTB11a��qCTB11b��ѿ������ЧӦ; H: qCTB1��qCTB3��ѿ������ۺ�ЧӦ; I: qCTB3��qCTB9��ѿ������ۺ�ЧӦ; J: qCTB9��qCTB11a��ѿ������ۺ�ЧӦ; K: qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; L: qCTB1��qCTB11b��ѿ������ۺ�ЧӦ; M: qCTB9��qCTB11b��ѿ������ۺ�ЧӦ; N: qCTB1��qCTB3��qCTB9��ѿ������ۺ�ЧӦ; O: qCTB3��qCTB9��qCTB11a��ѿ������ۺ�ЧӦ; P: qCTB1��qCTB3��qCTB11a��ѿ������ۺ�ЧӦ; Q: qCTB1��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; R: qCTB3��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; S: qCTB9��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; T: qCTB1��qCTB3��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; U: qCTB1��qCTB9��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; V: qCTB3��qCTB9��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; W: qCTB1��qCTB3��qCTB9��qCTB11a��qCTB11b��ѿ������ۺ�ЧӦ; X: ��QTL����ϵ���͡�
Fig. 7Effect of cold tolerance QTL and pyramiding effect at bud stage in LTH-SN265 RILs
A: Shennong 265 (SN265); B: Lijiangxintuanheigu (LTH); C-G: Cold tolerance effect of qCTB1, qCTB3, qCTB9, qCTB11a, and qCTB11b at bud stage; H: Cold tolerance cluster effect of qCTB1 and qCTB3 at bud stage; I: Cold tolerance cluster effect of qCTB3 and qCTB9 at bud stage; J: Cold tolerance cluster effect of qCTB9 and qCTB11a at bud stage; K: Cold tolerance cluster effect of qCTB11a and qCTB11b at bud stage; L: Cold tolerance cluster effect of qCTB11a and qCTB11b at bud stage; M: Cold tolerance cluster effect of qCTB9 and qCTB11b at bud stage; N: Cold tolerance cluster effect of qCTB1, qCTB3, and qCTB9 at bud stage; O: Cold tolerance cluster effect of qCTB3, qCTB9, and qCTB11a at bud stage; P: Cold tolerance cluster effect of qCTB1, qCTB3, and qCTB11a at bud stage; Q: Cold tolerance cluster effect of qCTB1, qCTB11a, and qCTB11b at bud stage; R: Cold tolerance cluster effect of qCTB3, qCTB11a, and qCTB11b at bud stage; S: Cold tolerance cluster effect of qCTB9, qCTB11a, and qCTB11b at bud stage; T: Cold tolerance cluster effect of qCTB1, qCTB3, qCTB11a, and qCTB11b at bud stage; U: Cold tolerance cluster effect of qCTB1, qCTB9, qCTB11a and qCTB11b at bud stage; V: Cold tolerance cluster effect of qCTB3, qCTB9, qCTB11a, and qCTB11b at bud stage; W: Cold tolerance cluster effect of qCTB1, qCTB3, qCTB9, qCTB11a, and qCTB11b at bud stage; X: none QTL line.
3 ����
3.1 ˮ��ѿ�����������۷����ıȽ�
ˮ��ѿ�����������Ǿ���ˮ��ֱ�������ʺ�����ȵ���Ҫũ����״, �������ڱ����������Ϸ�˫�����絾����ֱ���������Ҫ������״��������, ����ֱ�������ĸ���, Խ��Խ��ĵ�����ˮ��ֱ��ʱѡ��ѿ��ֱ������ä�Ȳ��֡�, ʹ��ѿ���������Գ�Ϊ����Ʒ���Ƿ�����ֱ������Ĺؼ����ء����, ȷ����������ϵ�ѿ���������Ծͱ��Խ��Խ��Ҫ��ѿ��������������бȽϳ��õ������õ��������䴦����ѿ�������, ����������ɻ�����Ϊ����ָ��, ���ַ�����ֱ�ۡ�����ɿ�[22,23,24,25,26,27,28,29,30,31,32,33]�����о������ַ��������˽�һ���ĸĽ�, ��Ҫ�������¼�������: һ�ǵ��´������¶Ⱥ�ʱ��, ǰ�˱�����, �����5��C 5 d [22,23,24,25,26,27,28,29,30,31,32,33], ���о�������2��C 2 d, �������˵��´����ĵ�������, �������˴���ʱ�䡣���ǵ��´����������������ʽ, ǰ�˶�ʹ��ȫ��ˮ��[22,23,24,25], ���о�ѡ������, �������ӽ�ˮ��ֱ������ʵ�ʡ�����ѿ���亦�ȼ������۷�ʽ, ǰ�˶��Ǽؽ���������������ɻ�����Ϊѿ�������Լ�����ָ��[22,23,24,25], ���о�Ϊ�˸�ȫ��ط�ӳÿһ����ϵ���ۺϿ���, �����˲�ͬ�����Ȩ�ϼƵķ�ʽ����ijһ��ϵ���������ˮƽ��ͨ�������Ľ�, ���������˴���ʱ��, ���һ�����˼����ľ��ȡ�3.2 �ز��������ܶ�bin����Ŵ�ͼ����ˮ��QTL�����е�Ӧ��
�����ز���ɱ��Ĵ���½��Ͳ����������ü������������, �����ز�������ϡ��������ڡ�������bin�Ŵ�ͼ���ѳ�Ϊˮ��[39,40,41]������[42,43]������[44]������ũ����״�Ŵ��о�����Ҫ�����ֶΡ��봫ͳ�ķ��ӱ�����, �ز����õ�SNP��Ǿ��и����鸲�ǶȺ��ܶȵ�����, ���������������QTL�����Ͷ�λ��ȷ�Ժ;�ȷ��, �����ܹ�Ϊ�����Ļ������о��ṩ��Խ�С��Ⱦɫ������ο������о����ð���2818��bin��ǵ��Ŵ�ͼ�ֱ���ˮ��ѿ�������Ե�QTL��λ����0.50 Mb~2.40 Mb��������, ��С����������ֱ���500 kb (qCTB1)��790 kb (qCTB11b), ��Խϴ����������Ҳ���ֱ���1.40 Mb (qCTB11a)��2.40 Mb (qCTB9)��3.47 Mb (qCTB3), ��ԶԶС�ڴ�ͳ���ӱ�Ǹߴ�5~10 cM�Ķ�λ����(Լ4 Mb~10 Mb)��500 kb���ҵ�QTL��λ������Ȼ��ֱ����ɻ����¡����һ���IJ��[45], ����ҲΪ�����ľ�ϸ��λ�ṩ���㹻С���������䡣����, ��ͳ��QTL������ͼȺ����Ҫ����-�����ּ��ӽ��õ���, ����Ϊ��-�̽�Ⱥ��, ����ʹ������������Ϊ�ױ����ϵ��Ŵ�Ⱥ��, ��Ҫ����Ϊ���������ڵĶ�̬��ԶԶС���̵�������, ����-�����ּ�Ķ�̬�����, ���Dz�༫��[46]���ز�������Ϊ�����һ�������Ч�ֶ�, ʹ�����þ�-����Ⱥ��Ҳ���Թ������ܶȵ��Ŵ�ͼ��, Ϊ�о�����ʳζƷ�ʡ��������ϰ����亦��Ҫ��˫��Ϊ������ʵ���ṩ�˼���ı�����3.3 ����QTLȫ������LTH�Ŀ���ԭ������RILȺ�����QTL�ۺ������Ŀ�����
���о��з���SN265��LTH�����������Խ�ϵȺ�����ѿ�������Եij�����, ����������QTLȫ������LTH, ��ԭ����Ҫ��QTL���ʱ, Ϊ�˾����ܽ��ͼ�����, ʹ������Խϸߵ�LOD��ֵ(3.0)�������LOD��ֵ���͵�2.4, �ܼ�һ�����ܵ�QTL-qCTB7, ��LODֵ��Ϊ2.45, �������ʺ�С, ��Ϊ5.8%, ����ЧӦֵΪ-0.13����QTL��������Ч��λ����������ũ265, �����ڱ�������̫С, ����Ҫ������QTL�ĸ������ؽ�Ⱥ�������α��֤������, �Լ���QTL����ЧӦ����, �����Ƕ�QTL���оۺ�Ч��������QTL����ˮ������ʵ������Ҫǰ�ᡣ����, һ�����, ���������Խ�ϵ�е���Ƭ����Խ϶�, �ų��Ŵ�����������Ӱ���ѶȺܴ�, �ܿ���Ӱ��QTL�ۼ�ЧӦ����������, �����Խ�ϵ��������QTL���ۼ�ЧӦ���������о������ˡ�ѡ����ͼ���ķ�������QTL���ۼ�ЧӦ����, �÷����Ĺؼ����dz�ַ����ѹ����ĸ��ܶ�����ͼ����ϸ��Ⱥ����������ݽ�Ŀ��QTL���������QTL���л����̶�, �Ծ������ų��Ŵ�������Ӱ�졣����֮ǰҲ�ɹ�������һ����������ˮ����ϵ��QTL�ۺ�Ч��[38]����Ȼ, ��ѡ����ͼ����QTL�ۺ�Ч���������϶�������Ȼ���ϵ��Ⱦɫ��Ƭ�δ���ϵ, ����û�и������Ŵ�Ⱥ��ʱ, �Բ�ʧΪһ����Ч�ֶΡ�3.4 ���о�������ˮ��ѿ���͵���QTL�������о�����ıȽϺ�����Ӧ��DZ��
����ũ���Ͷ����Ĵ��ģת�ƺ�ũҵ�����ɱ��IJ�������, ˮ��ֱ������Խ��Խ�ܵ�������ʡ�ع���ϡ��ũҵ������ũ�������ӡ�����, ���ڶ��صĺ�����̬����, ������ˮ��ֱ�����ò���Դ����ĵ��»�����ˮ��ѿ�����������ڵ�����ˮ����ѿϸ��ά�����������, ������ϱ���Ϊ��ѿ�շ���ά�����������, ��ˮ��ֱ���ؾ߱�����Ҫũ����״[22]�����Ĵ�ѿ��������ǿ�����ϵط�����Ʒ���������źڹ���, ������5������ѿ���͵���������QTL, ͬQTL���ݿ�(Q-TARO: http://qtaro.abr.affrc. go.jp/)�Ѽ��������������QTL�ȶԷ���, qCTB1������һ����֪�������յ���пָ������OsCOIN, �û���������������ǿˮ��������, ͬʱ���Ӹ����Ậ��[47]��qCTB3�����ڰ���һ����֪��������ػ���OsHOS1, ��ͨ����ICE1����������ˮ������в�ȷ�Ӧ[48]��qCTB9�����ڰ���2���ɴش��ڵ�������ػ���, �ֱ���OsDREB1A��OsDREB1B, ���߾������յ�, ����ˮ������в�ȷ�Ӧ[49]��qCTB11a����û�з����Ѿ��������亦���QTL�ͻ���, ��һ���·��ֵ�λ�㡣��ЧQTL- qCTB11b��Baruah��[27]������ѿ������QTL- qCTP11λ����ͬ������, qCTP11��Դ�ڱ������ط�Ʒ��A58�����Ͽ��Կ���, ����ˮ��ѿ�������Ե�QTL�ڲ�ͬ�Ŵ����������¾���һ����������, ���ҵ�11Ⱦɫ���ϼ�������Чλ��qCTB11b���нϴ�ĺ����о���ֵ������Ҳ���á�ѡ����ͼ���ķ�����������ЩQTL������DZ���;ۺ�Ч��, ����ѿ���͵���QTL��Ҫ����Ϊ����ЧӦ, �dz�����ˮ���ķ��Ӹ������֡�������ЧQTL-qCTB11b�����������кܴ���Ŵ�ЧӦ, ������������QTL�ۺ�ʱ, �������ֳ�����ͻ���ĸ���Ч��, ��������Ӧ�ü�ֵ��4 ����
�������������źڹȺ���ũ265�����Խ�ϵȺ�����2818��bin��ǵ��Ŵ�ͼ�ס����ô�Ⱥ���bin���ͼ��, ����5��ѿ������QTL, �ֲ���ˮ����1�š�3�š�9�ź�11��Ⱦɫ����, �������ʷֱ�Ϊ8.0%��8.2%��16.6%��24.4%��53.5%, ��Ч��λ��������������ױ��������źڹȡ���ЧQTL-qCTB11bλ��11��Ⱦɫ�峤�۶˵�17.97 Mb~23.03 Mb֮�䡣���á�ѡ����ͼ�����Խ�����QTL��֤���ۼ�ЧӦ����, ��ȷ�˿���ͨ��QTL���ۼӾۺ�ʵ��ѿ�������������Ŵ�����, �ۺϵ���ЧQTLԽ��, ���������ȼ�Խ�ߡ��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.5923/j.food.20120205.05URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
DOI:10.1073/pnas.0805303105URLPMID:18719107

Tolerance to abiotic stress is an important agronomic trait in crops and is controlled by many genes, which are called quantitative trait loci (QTLs). Identification of these QTLs will contribute not only to the understanding of plant biology but also for plant breeding, to achieve stable crop production around the world. Previously, we mapped three QTLs controlling low-temperature tolerance at the germination stage (called low-temperature germinability). To understand the molecular basis of one of these QTLs, qLTG3-1 (quantitative trait locus for low-temperature germinability on chromosome 3), map-based cloning was performed, and this QTL was shown to be encoded by a protein of unknown function. The QTL qLTG3-1 is strongly expressed in the embryo during seed germination. Before and during seed germination, specific localization of beta-glucuronidase staining in the tissues covering the embryo, which involved the epiblast covering the coleoptile and the aleurone layer of the seed coat, was observed. Expression of qLTG3-1 was tightly associated with vacuolation of the tissues covering the embryo. This may cause tissue weakening, resulting in reduction of the mechanical resistance to the growth potential of the coleoptile. These phenomena are quite similar to the model system of seed germination presented by dicot plants, suggesting that this model may be conserved in both dicot and monocot plants.
URLPMID:30120236
DOI:10.1016/j.plantsci.2010.04.004URL
DOI:10.1038/ncomms14788URLPMID:28332574
Low temperature is a major factor limiting rice productivity and geographical distribution. Improved cold tolerance and expanded cultivation to high-altitude or high-latitude regions would help meet growing rice demand. Here we explored a QTL for cold tolerance and cloned the gene, CTB4a (cold tolerance at booting stage), encoding a conserved leucine-rich repeat receptor-like kinase. We show that different CTB4a alleles confer distinct levels of cold tolerance and selection for variation in the CTB4a promoter region has occurred on the basis of environmental temperature. The newly generated cold-tolerant haplotype Tej-Hap-KMXBG was retained by artificial selection during temperate japonica evolution in cold habitats for low-temperature acclimation. Moreover, CTB4a interacts with AtpB, a beta subunit of ATP synthase. Upregulation of CTB4a correlates with increased ATP synthase activity, ATP content, enhanced seed setting and improved yield under cold stress conditions. These findings suggest strategies to improve cold tolerance in crop plants.
DOI:10.1016/j.cell.2015.01.046URLPMID:25728666
Rice is sensitive to cold and can be grown only in certain climate zones. Human selection of japonica rice has extended its growth zone to regions with lower temperature, while the molecular basis of this adaptation remains unknown. Here, we identify the quantitative trait locus COLD1 that confers chilling tolerance in japonica rice. Overexpression of COLD1(jap) significantly enhances chilling tolerance, whereas rice lines with deficiency or downregulation of COLD1(jap) are sensitive to cold. COLD1 encodes a regulator of G-protein signaling that localizes on plasma membrane and endoplasmic reticulum (ER). It interacts with the G-protein alpha subunit to activate the Ca(2+) channel for sensing low temperature and to accelerate G-protein GTPase activity. We further identify that a SNP in COLD1, SNP2, originated from Chinese Oryza rufipogon, is responsible for the ability of COLD(jap/ind) to confer chilling tolerance, supporting the importance of COLD1 in plant adaptation.
DOI:10.1073/pnas.1819769116URLPMID:30808744 [��������: 1]
Rice (Oryza sativa L.) is a chilling-sensitive staple crop that originated in subtropical regions of Asia. Introduction of the chilling tolerance trait enables the expansion of rice cultivation to temperate regions. Here we report the cloning and characterization of HAN1, a quantitative trait locus (QTL) that confers chilling tolerance on temperate japonica rice. HAN1 encodes an oxidase that catalyzes the conversion of biologically active jasmonoyl-L-isoleucine (JA-Ile) to the inactive form 12-hydroxy-JA-Ile (12OH-JA-Ile) and fine-tunes the JA-mediated chilling response. Natural variants in HAN1 diverged between indica and japonica rice during domestication. A specific allele from temperate japonica rice, which gained a putative MYB cis-element in the promoter of HAN1 during the divergence of the two japonica ecotypes, enhances the chilling tolerance of temperate japonica rice and allows it to adapt to a temperate climate. The results of this study extend our understanding of the northward expansion of rice cultivation and provide a target gene for the improvement of chilling tolerance in rice.
URL [��������: 1]
�����̵�Ʒ���Ͼ�11�;���Ʒ�ְ�������F1��ҩ������֯���������67���ӱ�������ֲ��(Doubled haploid, DH)�� �Ը�Ⱥ�幹����һ�Ű���131��RFLP��ǵ�ˮ������ͼ�ס� ��ѿ�ڵ�������Ϊָ�꣬ �����ױ����� DHϵ�ڵ���(4��5��)�����µ������ԡ������������������DHȺ���г�˫�������ֲ��� ����ѿ��������������������Ƶ�������״������������Ϊ������״����QTL��������ͼ������ �����ڵ�7Ⱦɫ����G379b-RG4������������������йصĻ���(Cts7) ��
URL [��������: 1]
�����̵�Ʒ���Ͼ�11�;���Ʒ�ְ�������F1��ҩ������֯���������67���ӱ�������ֲ��(Doubled haploid, DH)�� �Ը�Ⱥ�幹����һ�Ű���131��RFLP��ǵ�ˮ������ͼ�ס� ��ѿ�ڵ�������Ϊָ�꣬ �����ױ����� DHϵ�ڵ���(4��5��)�����µ������ԡ������������������DHȺ���г�˫�������ֲ��� ����ѿ��������������������Ƶ�������״������������Ϊ������״����QTL��������ͼ������ �����ڵ�7Ⱦɫ����G379b-RG4������������������йصĻ���(Cts7) ��
DOI:10.1016/j.plantsci.2004.09.021URL [��������: 1]
URL [��������: 6]
���̾�������23��/����1�ŵ�F2��3 ��200����ϵ��Ϊ��ͼȺ�壬������һ�ź���97������ ��SSR����ǵķ�������ͼ�ס���5����������£���F3��ϵ����ѿ�������Լ�����������SSR��ǽ�����ѿ��������������״λ�㣨QTL���������о����������ѿ����������F3��ϵȺ�гʵ��������ֲ�������Ϊ�ɶ������Ƶ�������״��������ѿ���������йص�QTL 3�����ֱ�λ�ڵ�2��4 ��7Ⱦɫ���ϣ��Ա��ͱ���Ĺ����ʷ�ΧΪ11.5%��20.5%�����У�λ�ڵ�4Ⱦɫ��RM273��RM303��qCTBP4�Ա��ͱ���Ĺ��������
URL [��������: 6]
���̾�������23��/����1�ŵ�F2��3 ��200����ϵ��Ϊ��ͼȺ�壬������һ�ź���97������ ��SSR����ǵķ�������ͼ�ס���5����������£���F3��ϵ����ѿ�������Լ�����������SSR��ǽ�����ѿ��������������״λ�㣨QTL���������о����������ѿ����������F3��ϵȺ�гʵ��������ֲ�������Ϊ�ɶ������Ƶ�������״��������ѿ���������йص�QTL 3�����ֱ�λ�ڵ�2��4 ��7Ⱦɫ���ϣ��Ա��ͱ���Ĺ����ʷ�ΧΪ11.5%��20.5%�����У�λ�ڵ�4Ⱦɫ��RM273��RM303��qCTBP4�Ա��ͱ���Ĺ��������
[��������: 5]
[��������: 5]
[��������: 5]
[��������: 5]
[��������: 5]
[��������: 5]
DOI:10.3969/j.issn.1001-7216.2010.03.004URL [��������: 3]
���̵�Ʒ��9311Ϊ���塢����Ʒ���ձ���Ϊ���幹����95��Ⱦɫ��Ƭ���û�ϵΪ���ϣ� ��5����������½���ѿ�������Լ��������������6���û�ϵ���´�����ij������������ױ�9311��һ�����죬����������ǿ��9311�����ô�����ͼ����������4����ѿ����������ص�QTL���ֱ�λ��ˮ����5�͵�7Ⱦɫ���ϡ�����qCTB��5��1��qCTB��5��2��qCTB��5��3�ֱ𱻶�λ�ڵ�5Ⱦɫ��RM267��RM1237��RM2422��RM6054��RM3321��RM1054֮���Ŵ�����ֱ�Ϊ21.3 cM��27.4 cM��12.7 cM���û�Ƭ����;qCTB��7����λ�ڵ�7Ⱦɫ��RM11-RM2752�����Ŵ�����Ϊ6.8 cM���û�Ƭ���ϡ�
DOI:10.3969/j.issn.1001-7216.2010.03.004URL [��������: 3]
���̵�Ʒ��9311Ϊ���塢����Ʒ���ձ���Ϊ���幹����95��Ⱦɫ��Ƭ���û�ϵΪ���ϣ� ��5����������½���ѿ�������Լ��������������6���û�ϵ���´�����ij������������ױ�9311��һ�����죬����������ǿ��9311�����ô�����ͼ����������4����ѿ����������ص�QTL���ֱ�λ��ˮ����5�͵�7Ⱦɫ���ϡ�����qCTB��5��1��qCTB��5��2��qCTB��5��3�ֱ𱻶�λ�ڵ�5Ⱦɫ��RM267��RM1237��RM2422��RM6054��RM3321��RM1054֮���Ŵ�����ֱ�Ϊ21.3 cM��27.4 cM��12.7 cM���û�Ƭ����;qCTB��7����λ�ڵ�7Ⱦɫ��RM11-RM2752�����Ŵ�����Ϊ6.8 cM���û�Ƭ���ϡ�
DOI:10.1007/s10681-008-9753-yURL [��������: 6]
The present study was conducted to understand the pattern of variation and the genetic bases for cold tolerance at the early growth stage in Asian rice. The genetic variation was investigated at the germination, plumule and seedling stages among 57 strains including cultivated rice (Oryza sativa ssp. indica and ssp. japonica) and its wild progenitor (Oryza rufipogon). The significant differentiation of cold tolerance was observed among the taxonomically divided groups. At the germination stage, both indica and japonica subspecies tended to be more tolerant than O. rufipogon, whereas at the plumule and seedling stages, ssp. japonica tended to be more tolerant than ssp. indica and O. rufipogon. Furthermore, in cold tolerance at the plumule stage, the clinal variation across the latitude of origins was observed within O. rufipogon and ssp. japonica, suggesting that the current pattern of variation seems to have been shaped by both their phylogenetic histories and on-going adaptation to the local environments. QTL analysis between O. sativa ssp. japonica (tolerant) and O. rufipogon (susceptible) revealed five putative QTLs for cold tolerance at the plumule and seedling stages but not at the germination stage. Substitution mapping was also carried out to precisely locate the two major QTLs for cold tolerance at the plumule stage, which could be used for improvement of tolerance to cold stress in ssp. indica.
DOI:10.1016/S1672-6308(09)60028-7URL [��������: 3]
A doubled haploid (DH) population consisted of 120 lines, derived from a cross between an indica variety, TN1, and a japonica variety, Chunjiang 06, was used to identify QTLs controlling rice cold tolerance at the plumule and 3-leaf-seedling stages by using the QTLNetwork software. The percentages of normal plumules after 4°C treatments for 7 d, 9 d, 11 d, and 14 d, as well as the cold stress tolerance index (CSTI) and the withering index (WI) of rice seedling were investigated. A total of five single-effect QTLs, each for percentages of normal plumules after 4°C treatments for 9 d, 11 d and 14 d, and CSTI and WI, respectively were identified. The QTLs for the percentages of normal plumules after low temperature treatments for 9 d, 11 d and 14 d were on chromosomes 4, 2 and 11, accounting for 14.1%, 17.3% and 21.5% of the phenotypic variation, respectively. QTLs for CSTI and WI were on chromosomes 10 and 1, respectively. Two pairs of epistatic loci were identified, but none of the epistatic loci involved the single-effect QTLs. The RM528–RM340 interval on chromosome 6 interacted with the RM278–RM3919B interval on chromosome 9 for CSTI, and the epistatic interaction accounted for 17.7% of the phenotypic variation. A pair of epistatic loci was identified for WI, the RM246–RM5461 interval on chromosome 1 interacted with the ISA–RM447 interval on chromosome 8, which accounted for 22.6% of the phenotypic variation.
DOI:10.3969/j.issn.1001��7216.2013.04.007URL [��������: 3]
��������ϵ�Ǹ��ױ�9311�����壩�;����ձ��磨���壩�����ĵ�Ƭ�δ���ϵȺ�壬��ѿ�ں����ڷֱ���е��´�������ѿ�ڴ�����Ļָ��ɻ��ʺ����ھ�Ҷ�̶���Ϊ�����Ե�����ָ��������������ۡ�����������ձ�����ѿ�ں����������Լ�����ǿ��9311����ͬ����ϵѿ�ں�����������Ҳ���ڼ��������죬ѿ�ں����������Խ�ǿ�Ĵ���ϵX724��X732��X733�Ϻõر�����9311��ũ����״�����������״��һ�����������9311��������18��ѿ��������QTL��5������������QTL���ֲ���12��Ⱦɫ���ϵ�20������Ƭ���У���12Ⱦɫ���RM1261-RM519��RM17����ͬʱ����ѿ�ں�����������QTL��ѿ��������QTL qCTP9��qCTP11.2��qCTP12.1������������QTL qCTS1.1��qCTS1.2���нϴ����ЧӦ����Ӧ����ϵ�����Խ�ǿ��
DOI:10.3969/j.issn.1001��7216.2013.04.007URL [��������: 3]
��������ϵ�Ǹ��ױ�9311�����壩�;����ձ��磨���壩�����ĵ�Ƭ�δ���ϵȺ�壬��ѿ�ں����ڷֱ���е��´�������ѿ�ڴ�����Ļָ��ɻ��ʺ����ھ�Ҷ�̶���Ϊ�����Ե�����ָ��������������ۡ�����������ձ�����ѿ�ں����������Լ�����ǿ��9311����ͬ����ϵѿ�ں�����������Ҳ���ڼ��������죬ѿ�ں����������Խ�ǿ�Ĵ���ϵX724��X732��X733�Ϻõر�����9311��ũ����״�����������״��һ�����������9311��������18��ѿ��������QTL��5������������QTL���ֲ���12��Ⱦɫ���ϵ�20������Ƭ���У���12Ⱦɫ���RM1261-RM519��RM17����ͬʱ����ѿ�ں�����������QTL��ѿ��������QTL qCTP9��qCTP11.2��qCTP12.1������������QTL qCTS1.1��qCTS1.2���нϴ����ЧӦ����Ӧ����ϵ�����Խ�ǿ��
[��������: 3]
[��������: 3]
DOI:10.3724/SP.J.1259.2015.00338URL [��������: 3]
ˮ��(Oryza sativa)ѿ�������������������������в��ɺ��ӵ���Ҫ������״, �����Ŵ������ĸ��źͻ������ص�Ӱ��; ���õ�Ƭ�δ���ϵ(SSSLs)�ܼ����Ŵ������ĸ��š����о���85����Ƭ�δ���ϵΪ����, �������ױ�Ϊ��½��4��, �����ױ�Ϊ�ձ��硣ͨ�������ط��������Dunnett’s���رȽ�, ������Ƭ�δ���ϵ�������ױ�֮��ѿ�������ԵIJ���, ���Դ���Ƭ���ϵ�ѿ������QTL���м�������P≤0.001Ϊ��ֵ����8��ѿ������QTL, �ֱ�ֲ��ڵ�1��6��8��9��10��Ⱦɫ����, ����4��QTLͨ��������ͼ��������λ����ЩQTL����ЧӦ������Ϊ��Ч����, ��2����ȼ������ЧӦֵ�ı仯��Χ�ֱ�Ϊ14%-44%��10%-45%, ����ЧӦ�ٷ��ʵı仯��Χ�ֱ�Ϊ700% -2 200%��500%-2 250%, ����qCTP9-2��2����ȼ�ļ���ЧӦ�����, �ֱ�Ϊ44%��45%���о�����Խ�һ������������µ�ˮ��ѿ������QTL������Ҫ���塣
DOI:10.3724/SP.J.1259.2015.00338URL [��������: 3]
ˮ��(Oryza sativa)ѿ�������������������������в��ɺ��ӵ���Ҫ������״, �����Ŵ������ĸ��źͻ������ص�Ӱ��; ���õ�Ƭ�δ���ϵ(SSSLs)�ܼ����Ŵ������ĸ��š����о���85����Ƭ�δ���ϵΪ����, �������ױ�Ϊ��½��4��, �����ױ�Ϊ�ձ��硣ͨ�������ط��������Dunnett’s���رȽ�, ������Ƭ�δ���ϵ�������ױ�֮��ѿ�������ԵIJ���, ���Դ���Ƭ���ϵ�ѿ������QTL���м�������P≤0.001Ϊ��ֵ����8��ѿ������QTL, �ֱ�ֲ��ڵ�1��6��8��9��10��Ⱦɫ����, ����4��QTLͨ��������ͼ��������λ����ЩQTL����ЧӦ������Ϊ��Ч����, ��2����ȼ������ЧӦֵ�ı仯��Χ�ֱ�Ϊ14%-44%��10%-45%, ����ЧӦ�ٷ��ʵı仯��Χ�ֱ�Ϊ700% -2 200%��500%-2 250%, ����qCTP9-2��2����ȼ�ļ���ЧӦ�����, �ֱ�Ϊ44%��45%���о�����Խ�һ������������µ�ˮ��ѿ������QTL������Ҫ���塣
DOI:10.1007/s11032-016-0520-9URL [��������: 3]
DOI:10.1007/s00299-017-2247-4URLPMID:29322237 [��������: 3]
KEY MESSAGE: A region containing three genes on chromosome 1 of indica rice was associated with cold tolerance at the bud burst stage; these results may be useful for breeding cold-tolerant lines. Low temperature at the bud burst stage is one of the major abiotic stresses limiting rice growth, especially in regions where rice seeds are sown directly. In this study, we investigated cold tolerance of rice at the bud burst stage and conducted a genome-wide association study (GWAS) based on the 5K rice array of 249 indica rice varieties widely distributed in China. We improved the method to assess cold tolerance at the bud burst stage in indica rice, and used severity of damage (SD) and seed survival rate (SR) as the cold-tolerant indices. Population structure analysis demonstrated that the Chinese indica panel was divided into three subgroups. In total, 47 significant single-nucleotide polymorphism (SNP) loci associated with SD and SR, were detected by association mapping based on mixed linear model. Because some loci overlapped between SD and SR, the loci contained 13 genome intervals and most of them have been reported previously. A major QTL for cold tolerance on chromosome 1 at the position of 31.6 Mb, explaining 13.2% of phenotypic variation, was selected for further analysis. Through LD decay, GO enrichment, RNA-seq data, and gene expression pattern analyses, we identified three genes (LOC_Os01g55510, LOC_Os01g55350 and LOC_Os01g55560) that were differentially expressed between cold-tolerant and cold-sensitive varieties, suggesting they may be candidate genes for cold tolerance. Together, our results provide a new method to assess cold tolerance in indica rice, and establish the foundation for isolating genes related to cold tolerance that could be used in rice breeding.
DOI:10.1101/gr.089516.108URLPMID:19420380 [��������: 1]
The next-generation sequencing technology coupled with the growing number of genome sequences opens the opportunity to redesign genotyping strategies for more effective genetic mapping and genome analysis. We have developed a high-throughput method for genotyping recombinant populations utilizing whole-genome resequencing data generated by the Illumina Genome Analyzer. A sliding window approach is designed to collectively examine genome-wide single nucleotide polymorphisms for genotype calling and recombination breakpoint determination. Using this method, we constructed a genetic map for 150 rice recombinant inbred lines with an expected genotype calling accuracy of 99.94% and a resolution of recombination breakpoints within an average of 40 kb. In comparison to the genetic map constructed with 287 PCR-based markers for the rice population, the sequencing-based method was approximately 20x faster in data collection and 35x more precise in recombination breakpoint determination. Using the sequencing-based genetic map, we located a quantitative trait locus of large effect on plant height in a 100-kb region containing the rice
DOI:10.3390/ijms21041284URL [��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1093/bioinformatics/btq565URLPMID:20966004 [��������: 1]
MOTIVATION: R/qtl is free and powerful software for mapping and exploring quantitative trait loci (QTL). R/qtl provides a fully comprehensive range of methods for a wide range of experimental cross types. We recently added multiple QTL mapping (MQM) to R/qtl. MQM adds higher statistical power to detect and disentangle the effects of multiple linked and unlinked QTL compared with many other methods. MQM for R/qtl adds many new features including improved handling of missing data, analysis of 10,000 s of molecular traits, permutation for determining significance thresholds for QTL and QTL hot spots, and visualizations for cis-trans and QTL interaction effects. MQM for R/qtl is the first free and open source implementation of MQM that is multi-platform, scalable and suitable for automated procedures and large genetical genomics datasets. AVAILABILITY: R/qtl is free and open source multi-platform software for the statistical language R, and is made available under the GPLv3 license. R/qtl can be installed from http://www.rqtl.org/. R/qtl queries should be directed at the mailing list, see http://www.rqtl.org/list/. CONTACT: kbroman@biostat.wisc.edu.
DOI:10.3969/j.issn.1001��7216.2014.06.005URL [��������: 2]
Ϊ��ʾˮ�����Ժ�������ϵ������Ե��Ŵ��������ԹG������½129�����ӽ������Ļؽ������Խ�ϵȺ��Ϊ���ϣ�����������Ӧ����245�����ӱ����ɵ��Ŵ�ͼ��ˮ�����Ժ�������������״���з���������10�����Ƹ�ƽ��ֱ�����ܸ�����ƽ�����������ƽ��������������״��QTL����ЩQTL�ֲ��ڵ�1��2��5��9��11Ⱦɫ���ϣ����ڵ�9��11Ⱦɫ���ϳɴطֲ�����������Ϊ10.7%~28.5%���������“ѡ����ͼ”���Զ���ЧQTL��������֤���������У�λ�ڵ�11Ⱦɫ���ϱ��C477��G320B֮��(5 461 121-6 686 166 bp)��ͬʱ���Ƹ������������������QTL qRL11.1��qASA11��qRN11.1���·��ֵ�λ�㣬����һ�����о���Ӧ�ü�ֵ��
DOI:10.3969/j.issn.1001��7216.2014.06.005URL [��������: 2]
Ϊ��ʾˮ�����Ժ�������ϵ������Ե��Ŵ��������ԹG������½129�����ӽ������Ļؽ������Խ�ϵȺ��Ϊ���ϣ�����������Ӧ����245�����ӱ����ɵ��Ŵ�ͼ��ˮ�����Ժ�������������״���з���������10�����Ƹ�ƽ��ֱ�����ܸ�����ƽ�����������ƽ��������������״��QTL����ЩQTL�ֲ��ڵ�1��2��5��9��11Ⱦɫ���ϣ����ڵ�9��11Ⱦɫ���ϳɴطֲ�����������Ϊ10.7%~28.5%���������“ѡ����ͼ”���Զ���ЧQTL��������֤���������У�λ�ڵ�11Ⱦɫ���ϱ��C477��G320B֮��(5 461 121-6 686 166 bp)��ͬʱ���Ƹ������������������QTL qRL11.1��qASA11��qRN11.1���·��ֵ�λ�㣬����һ�����о���Ӧ�ü�ֵ��
DOI:10.1371/journal.pone.0017595URLPMID:21390234 [��������: 1]
Huge efforts have been invested in the last two decades to dissect the genetic bases of complex traits including yields of many crop plants, through quantitative trait locus (QTL) analyses. However, almost all the studies were based on linkage maps constructed using low-throughput molecular markers, e.g. restriction fragment length polymorphisms (RFLPs) and simple sequence repeats (SSRs), thus are mostly of low density and not able to provide precise and complete information about the numbers and locations of the genes or QTLs controlling the traits. In this study, we constructed an ultra-high density genetic map based on high quality single nucleotide polymorphisms (SNPs) from low-coverage sequences of a recombinant inbred line (RIL) population of rice, generated using new sequencing technology. The quality of the map was assessed by validating the positions of several cloned genes including GS3 and GW5/qSW5, two major QTLs for grain length and grain width respectively, and OsC1, a qualitative trait locus for pigmentation. In all the cases the loci could be precisely resolved to the bins where the genes are located, indicating high quality and accuracy of the map. The SNP map was used to perform QTL analysis for yield and three yield-component traits, number of tillers per plant, number of grains per panicle and grain weight, using data from field trials conducted over years, in comparison to QTL mapping based on RFLPs/SSRs. The SNP map detected more QTLs especially for grain weight, with precise map locations, demonstrating advantages in detecting power and resolution relative to the RFLP/SSR map. Thus this study provided an example for ultra-high density map construction using sequencing technology. Moreover, the results obtained are helpful for understanding the genetic bases of the yield traits and for fine mapping and cloning of QTLs.
DOI:10.1073/pnas.1306579110URLPMID:23940322 [��������: 1]
The growing world population and shrinkage of arable land demand yield improvement of rice, one of the most important staple crops. To elucidate the genetic basis of yield and uncover its associated loci in rice, we resequenced the core recombinant inbred lines of Liang-You-Pei-Jiu, the widely cultivated super hybrid rice, and constructed a high-resolution linkage map. We detected 43 yield-associated quantitative trait loci, of which 20 are unique. Based on the high-density physical map, the genome sequences of paternal variety 93-11 and maternal cultivar PA64s of Liang-You-Pei-Jiu were significantly improved. The large recombinant inbred line population combined with plentiful high-quality single nucleotide polymorphisms and insertions/deletions between parental genomes allowed us to fine-map two quantitative trait loci, qSN8 and qSPB1, and to identify days to heading8 and lax panicle1 as candidate genes, respectively. The quantitative trait locus qSN8 was further confirmed to be days to heading8 by a complementation test. Our study provided an ideal platform for molecular breeding by targeting and dissecting yield-associated loci in rice.
DOI:10.3389/fpls.2017.01223URLPMID:28747923 [��������: 1]
Mapping major quantitative trait loci (QTL) responsible for rice seed germinability under low temperature (GULT) can provide valuable genetic source for improving cold tolerance in rice breeding. In this study, 124 rice backcross recombinant inbred lines (BRILs) derived from a cross indica cv. Changhui 891 and japonica cv. 02428 were genotyped through re-sequencing technology. A bin map was generated which includes 3057 bins covering distance of 1266.5 cM with an average of 0.41 cM between markers. On the basis of newly constructed high-density genetic map, six QTL were detected ranging from 40 to 140 kb on Nipponbare genome. Among these, two QTL qCGR8 and qGRR11 alleles shared by 02428 could increase GULT and seed germination recovery rate after cold stress, respectively. However, qNGR1 and qNGR4 may be two major QTL affecting indica Changhui 891germination under normal condition. QTL qGRR1 and qGRR8 affected the seed germination recovery rate after cold stress and the alleles with increasing effects were shared by the Changhui 891 could improve seed germination rate after cold stress dramatically. These QTL could be a highly valuable genetic factors for cold tolerance improvement in rice lines. Moreover, the BRILs developed in this study will serve as an appropriate choice for mapping and studying genetic basis of rice complex traits.
DOI:10.1186/s12864-015-2242-5URLPMID:26691201 [��������: 1]
BACKGROUND: To safeguard the food supply for the growing human population, it is important to understand and exploit the genetic basis of quantitative traits. Next-generation sequencing technology performs advantageously and effectively in genetic mapping and genome analysis of diverse genetic resources. Hence, we combined re-sequencing technology and a bin map strategy to construct an ultra-high-density bin map with thousands of bin markers to precisely map a quantitative trait locus. RESULTS: In this study, we generated a linkage map containing 1,151,856 high quality SNPs between Mo17 and B73, which were verified in the maize intermated B73 x Mo17 (IBM) Syn10 population. This resource is an excellent complement to existing maize genetic maps available in an online database (iPlant, http://data.maizecode.org/maize/qtl/syn10/ ). Moreover, in this population combined with the IBM Syn4 RIL population, we detected 135 QTLs for flowering time and plant height traits across the two populations. Eighteen known functional genes and twenty-five candidate genes for flowering time and plant height trait were fine-mapped into a 2.21-4.96 Mb interval. Map expansion and segregation distortion were also analyzed, and evidence for inadvertent selection of early flowering time in the process of mapping population development was observed. Furthermore, an updated integrated map with 1,151,856 high-quality SNPs, 2,916 traditional markers and 6,618 bin markers was constructed. The data were deposited into the iPlant Discovery Environment (DE), which provides a fundamental resource of genetic data for the maize genetic research community. CONCLUSIONS: Our findings provide basic essential genetic data for the maize genetic research community. An updated IBM Syn10 population and a reliable, verified high-quality SNP set between Mo17 and B73 will aid in future molecular breeding efforts.
URLPMID:26593310 [��������: 1]
DOI:10.1093/jxb/ers205URLPMID:22859680 [��������: 1]
The productivity of sorghum is mainly determined by agronomically important traits. The genetic bases of these traits have historically been dissected and analysed through quantitative trait locus (QTL) mapping based on linkage maps with low-throughput molecular markers, which is one of the factors that hinder precise and complete information about the numbers and locations of the genes or QTLs controlling the traits. In this study, an ultra-high-density linkage map based on high-quality single nucleotide polymorphisms (SNPs) generated from low-coverage sequences (~0.07 genome sequence) in a sorghum recombinant inbred line (RIL) population was constructed through new sequencing technology. This map consisted of 3418 bin markers and spanned 1591.4 cM of genome size with an average distance of 0.5 cM between adjacent bins. QTL analysis was performed and a total of 57 major QTLs were detected for eight agronomically important traits under two contrasting photoperiods. The phenotypic variation explained by individual QTLs varied from 3.40% to 33.82%. The high accuracy and quality of this map was evidenced by the finding that genes underlying two cloned QTLs, Dw3 for plant height (chromosome 7) and Ma1 for flowering time (chromosome 6), were localized to the correct genomic regions. The close associations between two genomic regions on chromosomes 6 and 7 with multiple traits suggested the existence of pleiotropy or tight linkage. Several major QTLs for heading date, plant height, numbers of nodes, stem diameter, panicle neck length, and flag leaf width were detected consistently under both photoperiods, providing useful information for understanding the genetic mechanisms of the agronomically important traits responsible for the change of photoperiod.
[��������: 1]
[��������: 1]
[��������: 3]
DOI:10.1007/s00425-007-0548-5URLPMID:17549515 [��������: 3]
Rice (Oryza sativa L.) plant is sensitive to chilling, particularly at early stages of seedling development. Here a novel cold-inducible gene, designated OsCOIN (Oryza sativa cold-inducible), was isolated and characterized. Results showed that OsCOIN protein, a RING finger protein, was localized in both nuclear and cytoplasm membrane. OsCOIN is expressed in all rice organs and strongly induced by low temperature, ABA, salt and drought. Over-expression of OsCOIN in transgenic rice lines significantly enhanced their tolerance to cold, salt and drought, accompanied by an up-regulation of OsP5CS expression and an increase of cellular proline level.
DOI:10.1007/s11103-013-0092-6URLPMID:23780733 [��������: 3]
Plants can cope with adverse environmental conditions through the activation of stress response signalling pathways, in which the proteasome seems to play an important role. However, the mechanisms underlying the proteasome-mediated stress response in rice are still not fully understood. To address this issue, we have identified a rice E3-ubiquitin ligase, OsHOS1, and characterized its role in the modulation of the cold stress response. Using a RNA interference (RNAi) transgenic approach we found that, under cold conditions, the RNAi::OsHOS1 plants showed a higher expression level of OsDREB1A. This was correlated with an increased amount of OsICE1, a master transcription factor of the cold stress signalling. However, the up-regulation of OsDREB1A was transient and the transgenic plants did not show increased cold tolerance. Nevertheless, we could confirm the interaction of OsHOS1 with OsICE1 by Yeast-Two hybrid and bi-molecular fluorescence complementation in Arabidopsis protoplasts. Moreover, we could also determine through an in vitro degradation assay that the higher amount of OsICE1 in the transgenic plants was correlated with a lower amount of OsHOS1. Hence, we could confirm the involvement of the proteasome in this response mechanism. Taken together our results confirm the importance of OsHOS1, and thus of the proteasome, in the modulation of the cold stress signalling in rice.
DOI:10.1371/journal.pone.0047275URLPMID:23077584 [��������: 1]
The clustered genes C-repeat (CRT) binding factor (CBF)1/dehydration-responsive element binding protein (DREB)1B, CBF2/DREB1C, and CBF3/DREB1A play a central role in cold acclimation and facilitate plant resistance to freezing in Arabidopsis thaliana. Rice (Oryza sativa L.) is very sensitive to low temperatures; enhancing the cold stress tolerance of rice is a key challenge to increasing its yield. In this study, we demonstrate chilling acclimation, a phenomenon similar to Arabidopsis cold acclimation, in rice. To determine whether rice CBF/DREB1 genes participate in this cold-responsive pathway, all putative homologs of Arabidopsis DREB1 genes were filtered from the complete rice genome through a BLASTP search, followed by phylogenetic, colinearity and expression analysis. We thereby identified 10 rice genes as putative DREB1 homologs: nine of these were located in rice genomic regions with some colinearity to the Arabidopsis CBF1-CBF4 region. Expression profiling revealed that six of these genes (Os01g73770, Os02g45450, Os04g48350, Os06g03670, Os09g35010, and Os09g35030) were similarly expressed in response to chilling acclimation and cold stress and were co-expressed with genes involved in cold signalling, suggesting that these DREB1 homologs may be involved in the cold response in rice. The results presented here serve as a prelude towards understanding the function of rice homologs of DREB1 genes in cold-sensitive crops.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}