 ,1,*, 景蕊莲,2,*
,1,*, 景蕊莲,2,*Genetic diversity assessment in derivative offspring of Mazhamai and Xiaobaimai wheat
BAI Yan-Ming1,2, LI Long2, WANG Hui-Yan1,2, LIU Yu-Ping2, WANG Jing-Yi2, MAO Xin-Guo2, CHANG Xiao-Ping2, SUN Dai-Zhen,1,*, JING Rui-Lian,2,*通讯作者:
第一联系人:
收稿日期:2019-02-1接受日期:2019-04-15网络出版日期:2019-04-22
| 基金资助: |
Received:2019-02-1Accepted:2019-04-15Online:2019-04-22
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1421KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
白彦明, 李龙, 王绘艳, 柳玉平, 王景一, 毛新国, 昌小平, 孙黛珍, 景蕊莲. 蚂蚱麦和小白麦衍生系的遗传多样性分析[J]. 作物学报, 2019, 45(10): 1468-1477. doi:10.3724/SP.J.1006.2019.91012
BAI Yan-Ming, LI Long, WANG Hui-Yan, LIU Yu-Ping, WANG Jing-Yi, MAO Xin-Guo, CHANG Xiao-Ping, SUN Dai-Zhen, JING Rui-Lian.
小麦是世界第二大粮食作物, 满足全球约35%人口对粮食的需求[1,2,3], 同时也是我国的主要口粮作物。黄淮冬麦区和北部冬麦区是我国的小麦主产区, 在保障国家“口粮绝对安全”中占有重要地位[4]。自20世纪40年代以来, 黄淮冬麦区以陕西关中地区的小麦地方品种蚂蚱麦、北部冬麦区以山西平遥的地方品种小白麦为原始骨干亲本, 分别育成系列大面积推广的优良品种和骨干亲本, 形成两大衍生系及两者的混合衍生系, 在我国北方小麦品种改良中发挥了重要作用[5]。这表明蚂蚱麦和小白麦中存在着有利基因并在子代中能高频传递。然而, 由于相同或相似优良亲本在小麦育种进程中的重复利用, 致使新品种遗传基础日益狭窄[6]。与其他作物类似, 在现代育种计划中, 小麦经过人为驯化和选择, 遗传多样性逐渐下降, 不仅使育种工作难以取得突破性进展, 而且降低了小麦应对不良环境的能力, 增加潜在风险[7]。因此, 通过分子标记研究小麦骨干亲本及其衍生系群体的遗传多样性, 对于高效利用种质资源及合理选配亲本, 拓宽品种的遗传基础具有重要意义[8]。
遗传多样性是植物遗传、育种、保护和进化的重要组成部分, 根据遗传多样性可以了解群体遗传特征, 解析品种之间的亲缘关系[9]。遗传多样性亦是表型-基因型关联分析的前提, 对于关联分析定位结果的准确性至关重要[10]。传统的系谱追踪分析法只能定性判断品种之间的亲缘关系, 加之某些品种的遗传背景比较复杂, 培育过程涉及多个亲本和多代杂交, 为鉴定品种之间的亲缘关系增加了挑战。然而, 随着分子标记技术的发展, 单核苷酸多态性(single nucleotide polymorphism, SNP)标记因其数量多、分布广、密度高等特点[11,12], 已被广泛应用于小麦遗传多样性研究。Chao等[13]利用SNP标记对来自64个国家的429份春小麦品种进行遗传多样性分析, 发现该群体多样性较丰富; 陈广凤等[14]利用SNP标记分析发现, 由我国冬麦区205份小麦育成品种(系)构成的自然群体遗传多样性丰富; 刘新伦等[15]通过对骨干亲本阿夫及其衍生系的遗传分析, 揭示了该衍生系重要性状的演变规律; 曹廷杰等[8]利用SNP标记将河南省近年审定的96个小麦品种分为7个亚群, 并指出河南小麦品种的遗传多样性较低, 迫切需要引入新的种质资源以拓宽遗传基础。
陕西关中蚂蚱麦和山西平遥小白麦是我国的小麦地方品种, 以这两个地方品种为原始亲本, 均培育出了大量的品种。前人曾经研究了蚂蚱麦[16]和小白麦[17]衍生品种的遗传构成, 但对其衍生系进行综合遗传多样性分析的研究尚未见报道。本研究以黄淮冬麦区蚂蚱麦衍生品种(系)和北部冬麦区小白麦衍生品种(系)为材料, 采用小麦660K SNP芯片标记检测基因型, 分析其遗传多样性, 旨在为优异基因资源合理利用和亲本选配提供理论依据。
1 材料与方法
1.1 小麦材料
供试小麦材料149份(附表1), 分为两类: (1)蚂蚱麦、小白麦及其衍生系136份, 包括蚂蚱麦4份(陕西关中地区3份, 山东冠县1份), 关中地区蚂蚱麦衍生品种(系) 87份; 山西平遥的小白麦1份, 小白麦衍生品种(系)103份; 其中同时具有蚂蚱麦和小白麦血缘的衍生品种(系) 59份; (2)大面积推广品种13份, 没有蚂蚱麦和小白麦血缘, 作为对照品种。供试材料分别由中国农业科学院作物科学研究所国家作物种质库及甘肃农业大学杨德龙教授提供。1.2 DNA提取及检测
小麦种子经7 d水培, 取5株幼苗叶片混合为1个样品, 采用CTAB法提取基因组DNA[18], 用1%的琼脂糖凝胶电泳定性检测DNA质量和浓度, 用紫外分光光度计通过NanoDrop 2000软件定量检测DNA浓度及纯度, 将达标的DNA稀释到50 ng μL-1用于SNP标记检测。1.3 SNP芯片检测结果分析
利用小麦660K SNP芯片对小麦材料进行全基因组标记扫描(博奥晶典生物技术有限公司)。该芯片含有630,517个SNP标记, 标记的物理位置来源于中国春基因组测序结果(IWGSC WGA v0.4)。剔除缺失率大于20%和频率小于5%的SNP标记, 获得有效标记用于遗传多样性分析。1.4 遗传多样性分析
利用PowerMarker V3.25软件[19]计算SNP位点的基因多样性(gene diversity, H); 利用TASSEL 5.0软件计算核苷酸多样性指数(nucleotide diversity, π); 采用MEGA 6.0软件构建NJ聚类树, 并通过ITOL在线工具对图形润色; 用NTSYS-PC 2.1软件[20]计算遗传相似系数(genetic similarity, GS)。2 结果与分析
2.1 多态性SNP位点分布
利用小麦660K SNP芯片检测149份材料的基因型, 共获得630,517个SNP位点, 剔除缺失率大于20%及频率小于5%的SNP位点, 最终获得368,376个多态性SNP位点, 分布于21条染色体上。不同染色体上的多态性SNP位点数差异明显, 变幅为2954~43,997个, 其中染色体3B的多态性SNP位点最多, 4D最少; 在A、B、D三个基因组中, 第3同源群的多态性SNP位点数最多, 为67,909个, 第四同源群的多态性SNP位点数最少, 只有28,496个(图1-A, B)。比较3个基因组的多态性SNP位点数目, B基因组的多态性位点数最多, D基因组上的位点数目远少于A和B基因组, 其顺序为B>A>D (图1-C)。图1
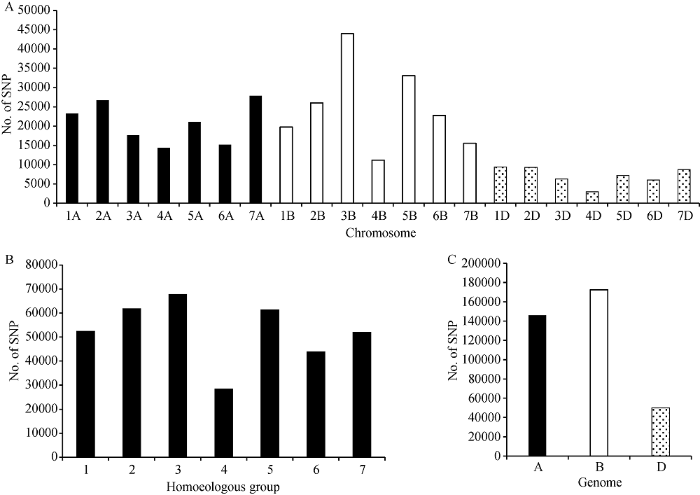 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1多态性SNP标记在染色体(A)、部分同源群(B)和基因组(C)上的分布
Fig. 1Distribution of polymorphic SNP markers on chromosomes (A), homoeologous groups (B), and genomes (C)
2.2 小麦材料SNP位点多态性
对149份小麦材料的SNP位点多态性分析表明, 其基因多样性(H)变幅为0.095~0.500, 平均为0.336。不同材料间的遗传相似系数(GS)变幅为0.335~0.997, 平均为0.619, 95.23%材料的GS<0.74 (图2)。供试材料的核苷酸多样性指数π值变幅为0.272~0.435, 平均为0.340。图2
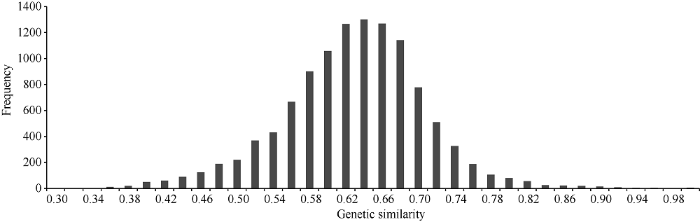 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2供试材料间遗传相似系数频次分布图
Fig. 2Frequency distributions of genetic similarity among accessions
2.3 蚂蚱麦、小白麦衍生系核苷酸多样性随年代和衍生世代变化趋势
根据SNP标记检测结果, 分析蚂蚱麦和小白麦衍生系, 以及大面积推广品种的核苷酸多样性(π)随育成年代的变化趋势, 结果显示1980s及之后育成品种和1960s及之前育成品种π值基本稳定, 差异微小, 而1970s育成品种(系) π值最小, 为0.352 (表1)。分别分析蚂蚱麦和小白麦不同世代衍生系π值变化趋势发现, 蚂蚱麦衍生九代π值最小, 为0.275, 衍生六代π值最大, 为0.355 (表2); 将小白麦与其系选品种燕大1817归为同一世代, 其衍生一代π值最小, 为0.233, 衍生五代π值最大, 为0.375 (表3)。由于不同世代选择的育种亲本不同, 衍生世代和π值间没有必然规律可循, 不同衍生世代的π值可用于比较分析各世代的核苷酸多样性。Table 1
表1
表1不同年代育成材料的核苷酸多样性
Table 1
| 年代 Era | π值 π value | 品种数 No. of accession |
|---|---|---|
| Pre-1960 | 0.389 | 12 |
| 1960s | 0.391 | 26 |
| 1970s | 0.352 | 19 |
| 1980s | 0.387 | 13 |
| 1990s | 0.387 | 19 |
| 2000s | 0.388 | 34 |
| 2010s | 0.388 | 26 |
新窗口打开|下载CSV
Table 2
表2
表2蚂蚱麦各衍生世代材料及相关信息
Table 2
| 衍生世代 Generation | 品种名称 Accession | 品种数 No. of accession | 核苷酸多样性 Nucleotide diversity (π) |
|---|---|---|---|
| 1 | 碧蚂2号 Bima 2 | 1 | |
| 2 | 衡水6404, 邢选1号, 北京8号, 陕农4号, 徐州6号, 淮沭10号, 昌乐5号, 济南2号, 石家庄54, 青春1号, 青春2号, 陕农17-17, 郑州24, 陕合6号, 陕农1号, 陕农18 Hengshui 6404, Xingxuan 1, Beijing 8, Shaannong 4, Xuzhou 6, Huaishu 10, Changle 5, Jinan 2, Shijiazhuang 54, Qingchun 1, Qingchun 2, Shaannong 17-17, Zhengzhou 24, Shaanhe 6, Shaannong 1, Shaannong 18 | 16 | 0.348 |
| 3 | 北京13, 农大198, 济南10号, 冀麦2号, 平凉35, 西峰9号, 冀麦1号 Beijing 13, Nongda 198, Jinan 10, Jimai 2, Pingliang 35, Xifeng 9, Jimai 1 | 7 | 0.305 |
| 4 | 晋麦29, 庆丰1号, ZHM 44, 晋麦47, 长武702, 丰抗13, 平凉21 Jinmai 29, Qingfeng 1, ZHM 44, Jinmai 47, Changwu 702, Fengkang 13, Pingliang 21 | 7 | 0.353 |
| 5 | 运旱20410, 临旱6号, 运旱22-33, 洛旱13, 晋麦79, 晋麦33, 洛旱2号, 丰抗8号, 丰抗7号, 京农79-13, 丰抗10号, 兰天4号, 京437, ZHM 40, ZHM 42, ZHM 43, 洛旱15, 中麦36 Yunhan 20410, Linhan 6, Yunhan 22-33, Luohan 13, Jinmai 79, Jinmai 33, Luohan 2, Fengkang 8, Fengkang 7, Jingnong 79-13, Fengkang 10, Lantian 4, Jing 437, ZHM 40, ZHM 42, ZHM 43, Luohan 15, Zhongmai 36 | 18 | 0.305 |
| 6 | 晋麦100, 晋麦92, 晋太170, 晋麦63, 西峰20, 中麦175, 北京8694, 京冬8号 Jinmai 100, Jinmai 92, Jintai 170, Jinmai 63, Xifeng 20, Zhongmai 175, Beijing 8694, Jingdong 8 | 8 | 0.355 |
| 7 | 陇育5号, 长4640, 长6878, 长6154, 长6452, 晋麦87, 长7016, 临丰615, 洛旱11, 洛旱9号, 洛旱6号, 洛旱3号 Longyu 5, Chang 4640, Chang 6878, Chang 6154, Chang 6452, Jinmai 87, Chang 7016, Linfeng 615, Luohan 11, Luohan 9, Luohan 6, Luohan 3 | 12 | 0.333 |
| 8 | 长武134, 宁冬16, 晋太1310, 晋太114, 长8744, 长6794, 长5222, 运旱137, 长麦6197, 长7050, ZHM 35, 鲁麦14, 衡136 Changwu 134, Ningdong 16, Jintai 1310, Jintai 114, Chang 8744, Chang 6794, Chang 5222, Yunhan 137, Changmai 6197, Chang 7050, ZHM 35, Lumai 14, Heng 136 | 13 | 0.339 |
| 9 | 长6990, 长6388, 长9499, ZHM 41, ZHM 45 Chang 6990, Chang 6388, Chang 9499, ZHM 41, ZHM 45 | 5 | 0.275 |
新窗口打开|下载CSV
Table 3
表3
表3小白麦/燕大1817各衍生世代材料及相关信息
Table 3
| 衍生世代 Generation | 品种名称 Accession | 品种数 No. of accession | 核苷酸多样性Nucleotide diversity (π) |
|---|---|---|---|
| 1 | 农大36, 农大183, 农大311, 农大90, 太原566, 石家庄407, 北京5号, 北京6号, 北京 7号 Nongda 36, Nongda 183, Nongda 311, Nongda 90, Taiyuan 566, Shijiazhuang 407, Beijing 5, Beijing 6, Beijing 7 | 9 | 0.233 |
| 2 | 晋麦1号, 农大155, 农大166, 工农12, 渭麦4号, 科遗29, 旱选1号, 旱选2号, 旱选3号, 晋麦5号, 晋麦8号, 晋麦17, 冀麦32, 延安15, 冀麦6号, 洛农10号, 济南10号 Jinmai 1, Nongda 155, Nongda 166, Gongnong 12, Weimai 4, Keyi 29, Hanxuan 1, Hanxuan 2, Hanxuan 3, Jinmai 5, Jinmai 8, Jinmai 17, Jimai 32, Yanan 15, Jimai 6, Luonong 10, Jinan 10 | 17 | 0.299 |
| 3 | 晋麦44, 晋麦16, 长治515, 东方红3号, 晋麦13, 冀麦9号, 西峰16, 沧麦6005, 北京10号, 北京13, 丰抗8号, 丰抗7号, 京农79-13, 丰抗10号, 衡水6404, 邢选1号, 晋麦29 Jinmai 44, Jinmai 16, Changzhi 515, Dongfanghong 3, Jinmai 13, Jimai 9, Xifeng 16, Cangmai 6005, Beijing 10, Beijing 13, Fengkang 8, Fengkang 7, Jingnong 79-13, Fengkang 10, Hengshui 6404, Xingxuan 1, Jinmai 29 | 17 | 0.323 |
| 4 | 冀麦22, 兰天10号, 冀麦29, 晋麦18, 晋麦51, 兰天9号, 晋麦33, 农大198, 庆丰1号, 北京8694, 京冬8号 Jimai 22, Lantian 10, Jimai 29, Jinmai 18, Jinmai 51, Lantian 9, Jinmai 33, Nongda 198, Qingfeng 1, Beijing 8694, Jingdong 8 | 11 | 0.359 |
| 5 | 晋麦46, 晋麦53, 晋麦92, 晋麦79, 洛旱2号, 兰天4号, 晋麦47 Jinmai 46, Jinmai 53, Jinmai 92, Jinmai 79, Luohan 2, Lantian 4, Jinmai 47 | 7 | 0.375 |
| 6 | 太10604, 运旱20410, 临旱6号, 运旱22-33, 长武134, 晋麦100, 洛旱13, 晋麦63, 西峰20, 京437, ZHM 40, ZHM 42, ZHM 43, 洛旱15, 中麦36 Tai 10604, Yunhan 20410, Linhan 6, Yunhan 22-33, Changwu 134, Jinmai 100, Luohan 13, Jinmai 63, Xifeng 20, Jing 437, ZHM 40, ZHM 42, ZHM 43, Luohan 15, Zhongmai 36 | 15 | 0.304 |
| 7 | 陇育5号, 长4640, 晋太170, 长6878, 长6154, 长6452, 晋麦87, 长7016, 中麦175 Longyu 5, Chang 4640, Jintai 170, Chang 6878, Chang 6154, Chang 6452, Jinmai 87, Chang 7016, Zhongmai 175 | 9 | 0.306 |
| 8 | 宁冬16, 晋太1310, 晋太114, 长8744, 长6794, 长5222, 运旱137, 长麦6197, 长7050, ZHM 35 Ningdong 16, Jintai 1310, Jintai 114, Chang 8744, Chang 6794, Chang 5222, Yunhan 137, Changmai 6197, Chang 7050, ZHM 35 | 10 | 0.329 |
| 9 | 长6990, 长6388, 长9499, ZHM 41, ZHM 45, 鲁麦14 Chang 6990, Chang 6388, Chang 9499, ZHM 41, ZHM 45, Lumai 14 | 6 | 0.259 |
| 10 | ZHM 44 ZHM 44 | 1 |
新窗口打开|下载CSV
2.4 聚类分析
根据SNP标记信息聚类分析表明, 供试材料可分为三大类群, 5个亚群(图3), 即北部冬麦区小白麦衍生系(亚群I和III), 黄淮冬麦区蚂蚱麦衍生系(亚群II), 以及同时具有蚂蚱麦和小白麦血缘的衍生系(亚群IV和V)。图3
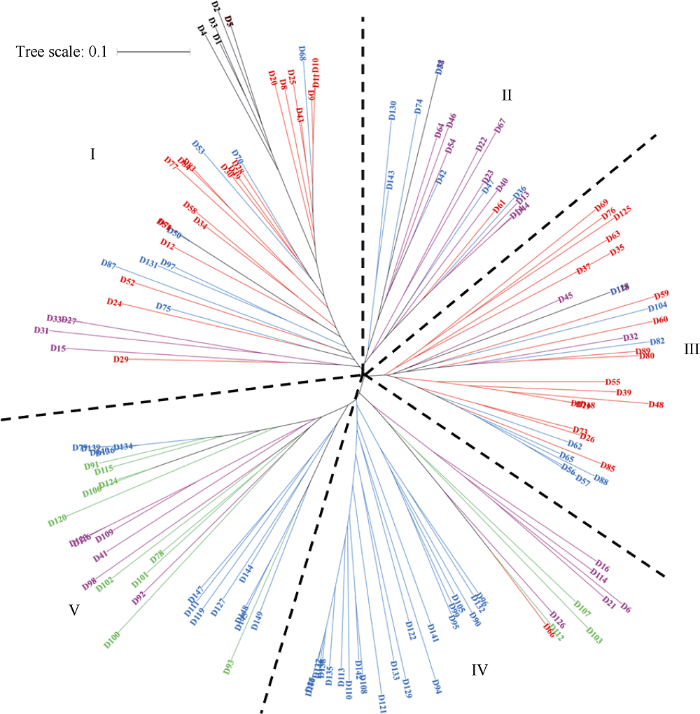 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3149份材料基于多态性SNP的聚类分析图
黑色表示蚂蚱麦和小白麦/燕大1817; 紫色表示蚂蚱麦衍生系; 红色表示小白麦衍生系; 蓝色表示有蚂蚱麦和小白麦共同血缘的衍生品(系); 绿色表示大面积推广品种。
Fig. 3Cluster analysis of 149 accessions based on the polymorphic SNP
The black fonts indicate Mazhamai and Xiaobaimai/Yanda 1817. The purple fonts indicate the derivative varieties (lines) of Mazhamai. The red fonts indicate the derivative varieties (lines) of Xiaobaimai. The blue fonts indicate derivative varieties (lines) with both kinship of Mazhamai and Xiaobaimai. The green fonts indicate commercial varieties.
第I亚群有39份材料, 以小白麦及其衍生系材料为主。包括小白麦及其系选品种燕大1817, 小白麦衍生材料21份, 具有小白麦和蚂蚱麦共同血缘的衍生材料8份, 蚂蚱麦4份, 以及蚂蚱麦衍生材料4份。在这个主要由北部冬麦区小白麦及其衍生系构成的亚群中, 来自黄淮冬麦区的4份蚂蚱麦(D1~D4)与来自北部冬麦区的小白麦(D5)及其系选品种燕大1817 (D7)紧密地聚在一起, 表明6份地方品种之间的遗传相似性很高。小白麦(D5)与其系选品种燕大1817 (D7)的SNP位点一致性高达99.1%, 表明燕大1817与小白麦间只有微小的遗传变异。该亚群平均π值为0.306, 主要为2000年之前的育成品种(表4), 分布于山西、陕西、河北和北京(表5)。
Table 4
表4
表4各亚群材料的育成年代
Table 4
| 育成年代 Released era | 亚群I Subgroup I | 亚群II Subgroup II | 亚群III Subgroup III | 亚群IV Subgroup IV | 亚群V Subgroup V |
|---|---|---|---|---|---|
| Pre-1960 | 11 | — | — | 1 | — |
| 1960s | 11 | 6 | 7 | 2 | — |
| 1970s | 6 | 6 | 6 | — | 1 |
| 1980s | 3 | 3 | 6 | 1 | — |
| 1990s | 6 | 2 | 7 | 2 | 2 |
| 2000s | 1 | — | 2 | 17 | 15 |
| 2010s | 1 | 2 | 2 | 9 | 11 |
| 合计Total | 39 | 19 | 30 | 32 | 29 |
新窗口打开|下载CSV
Table 5
表5
表5各亚群材料的地理来源分布
Table 5
| 来源 Origin | 亚群I Subgroup I | 亚群II Subgroup II | 亚群III Subgroup III | 亚群IV Subgroup IV | 亚群V Subgroup V |
|---|---|---|---|---|---|
| 山西Shanxi | 17 | — | 7 | 15 | 9 |
| 陕西Shaanxi | 7 | 1 | 3 | 4 | 2 |
| 河北Hebei | 5 | 1 | 3 | 4 | 1 |
| 河南Henan | — | 1 | — | 4 | 7 |
| 北京Beijing | 7 | 8 | 12 | 5 | 4 |
| 甘肃Gansu | 1 | 4 | 5 | — | — |
| 江苏Jiangsu | 1 | — | — | — | 1 |
| 山东Shandong | 1 | 3 | — | — | 5 |
| 宁夏Ningxia | — | 1 | — | — | — |
| 合计Total | 39 | 19 | 30 | 32 | 29 |
新窗口打开|下载CSV
第II亚群共19份材料, 除科遗29 (D61)外, 其余均为蚂蚱麦后代碧蚂4号的衍生系, 其中7份材料具有蚂蚱麦和小白麦的共同血缘。该亚群平均π值为0.288, 主要为1960s—1970s的育成品种, 分布于北京、甘肃和山东。
第III亚群有30份材料, 以小白麦衍生一代农大183和农大311及其衍生材料为主, 其中小白麦衍生材料20份, 拥有小白麦和蚂蚱麦共同血缘的衍生材料8份, 另外2份为蚂蚱麦衍生材料平凉21 (D32)和长武702 (D45)。该亚群平均π值为0.305, 以1960s—1990s的育成品种为主, 分布于北京、山西和甘肃。
第IV亚群的32份材料主要是具有蚂蚱麦和小白麦共同血缘的衍生系, 蚂蚱麦单一衍生材料5份, 另外有大面积推广品种3份。该亚群平均π值为0.333, 以2000年之后的育成品种为主, 分布于山西、陕西、河南、河北和北京。
第V亚群的29份材料以蚂蚱麦衍生系为主, 其中晋麦63 (D90)具有蚂蚱麦和小白麦共同血缘, 由其衍生的品种(系) 13份, 只有蚂蚱麦血缘的衍生品种6份, 以及大面积推广品种10份。该亚群平均π值为0.314, 主要为2000年之后的育成品种, 分布于山西、河南、山东和北京。
2.5 同名异种蚂蚱麦、小白麦及燕大1817的遗传相似性
149份供试材料中, 来自黄淮冬麦区的4份蚂蚱麦(D1~D4)与来自北部冬麦区的小白麦(D5)和燕大1817(D7)紧密地聚为一类(图3)。该6份材料两两间GS变幅为0.628~0.989, 平均GS为0.750。其中D5和D7间GS最大, 为0.989; D5、D7与D4的GS最小, 均为0.628。D1~D4间GS变幅为0.738~0.889, 平均为0.802, 其中D1与D4的GS最小, 为0.738, 与D3的GS最大, 为0.889。陕西关中地区3份蚂蚱麦和小白麦、燕大1817间的GS范围在0.635~0.799, 平均GS为0.699, 其中D1与两者的GS最高, D3与两者的GS最低(表6)。该结果表明, 不同地域来源的同名异种蚂蚱麦间的GS有差别。Table 6
表6
表6蚂蚱麦、小白麦及燕大1817间的遗传相似系数
Table 6
| 品种 Accession | D1 (岐山蚂蚱麦) Mazhamai of Qishan | D2 (武功蚂蚱麦) Mazhamai of Wugong | D3 (扶风蚂蚱麦) Mazhamai of Fufeng | D4 (冠县蚂蚱麦) Mazhamai of Guanxian | D5 (小白麦) Xiaobaimai |
|---|---|---|---|---|---|
| D2 (武功蚂蚱麦) Mazhamai of Wugong | 0.747 | ||||
| D3 (扶风蚂蚱麦) Mazhamai of Fufeng | 0.889 | 0.802 | |||
| D4 (冠县蚂蚱麦) Mazhamai of Guanxian | 0.738 | 0.849 | 0.787 | ||
| D5 (小白麦) Xiaobaimai | 0.798 | 0.663 | 0.636 | 0.628 | |
| D7 (燕大1817) Yanda 1817 | 0.799 | 0.664 | 0.635 | 0.628 | 0.989 |
新窗口打开|下载CSV
3 讨论
利用分子标记研究小麦遗传多样性的报道较多[21,22,23,24,25,26], 不过早期研究所用的分子标记多数是SSR、AFLP或者RFLP, 标记数量较少, 难以覆盖全基因组。本研究利用小麦660K SNP标记对149份材料进行全基因组扫描表明, B基因组的多态性标记数最多, D基因组的多态性标记数最少(图1-C), 这与曹廷杰等[8]和贾继增等[27]的研究结果一致。其原因可能是D基因组中基因序列相对保守, 亦或是其最后参与了六倍体小麦的形成, 进化时间较短 [8]。本研究还发现第四同源群多态性标记数最少, 与之对应的4A、4B、4D多态性标记数在三个基因组中也分别最少(图1-A), 与以往研究结果相同, 表明第四同源群的基因组序列相对保守。前人已对多种作物进行了遗传多样性研究, 如小麦[28,29]和水稻[30,31]等研究发现, 由于人工和自然选择的影响, 使得作物遗传多样性逐渐降低。本研究利用SNP标记分析不同世代或年代育成品种(系)的核苷酸多样性(π), 发现蚂蚱麦衍生九代(表2)和小白麦衍生一代(表3)的π值均最小, 可能是这2个衍生世代中的品种分别为姊妹系。然而, 对包括13份大面积推广品种的综合分析发现, 1980s及之后育成品种和1960s及之前育成品种π值基本稳定, 而1970s育成品种的π值最小(表1), 可能是1970s育成品种所用种质材料的遗传基础比较狭窄, 且在品种培育过程中受到了较强的人工选择所致。
聚类分析结果表明, 黄淮冬麦区的同名异种蚂蚱麦与北部冬麦区的小白麦均位于第I亚群, 并且紧密地聚为一小类(图3), 6份地方品种间的平均GS为0.750 (表6), 遗传相似性较高, 其原因可能是在早期较低的生产水平和环境条件下, 通过长期的人工和自然选择, 地方品种等位基因趋同化, 例如植株较高, 茎秆较细, 分蘖力强, 小穗小粒, 耐逆性强等[32]。陕西关中地区蚂蚱麦(D1~D3)与山西平遥小白麦间的平均GS为0.699 (表6), 遗传相似性也较高。第I亚群主要是1980年之前选育的品种, 与近年来的育成品种相比, 早期育成品种与地方品种的性状特征相似性更高, 这在一定程度上较好地解释了蚂蚱麦和小白麦紧密地聚在第I亚群的现象。第II亚群中, 碧蚂4号衍生系聚为一类, 李小军等[16]曾报道碧蚂4号在主要农艺性状上对后代衍生品种具有较高的遗传贡献, 这或许是其衍生品种(系)能够单独聚为一类的原因。但是, 在第III亚群中, 具有蚂蚱麦和小白麦共同血缘的8份材料与小白麦衍生系聚为一类, 而在第IV亚群中, 具有蚂蚱麦和小白麦共同血缘的品种(系)则与蚂蚱麦衍生系聚为一类, 其原因可能是不同地区的育种目标侧重点不同, 因此对蚂蚱麦和小白麦优良基因的人工选择方向有所区别。第V亚群中晋麦63衍生系(D144、D145等)与大面积推广品种聚为一类, 可能是因为晋麦63不仅拥有蚂蚱麦和小白麦的血缘, 同时也导入了多个国外种质的优良基因, 例如胜利麦(美)、早洋麦(美)、伊卡124D (罗)、苏早1号(苏)、小鹅186 (苏)等, 这些优良基因也传递到其衍生后代中, 使其成为重要推广品种。前人研究表明, 大面积推广品种的亲本可以追溯到较多的骨干亲本, 因此也聚合了大量优良基因, 这可能也是大面积推广品种能够聚在同一亚群的原因。
聚类分析结果还表明, 具有共同亲本或同一育种单位选育的品种(系)优先聚为一类(图3)。例如, 以燕大1817、有芒红7号、华北187、晋麦47、晋麦63、北京8号和鲁麦14为亲本的衍生系在各亚群中优先聚类。不过, 聚类分析结果与系谱信息也存在一定差异, 例如, 亚群I和III大部分为小白麦及其衍生系, 亲缘关系较近, 但却被亚群II隔开, 可能是尽管碧蚂4号衍生系拥有较多的蚂蚱麦血缘, 而蚂蚱麦和小白麦的遗传相似性较高, 同时亚群I和II主要是1960s—1970s的育成品种, 且以北京的材料居多, 具有较高的相似性, 因此亚群II优先与亚群I聚在一起。徐州6号(D15)和陕农18 (D16)均为1961年从杂交组合“碧蚂1号/苏联早熟1号”中选育的品种, 但却分别聚在亚群I和亚群IV, 其GS为0.572, 表明两个品种遗传差异较大, 可能是由于江苏省徐州地区与陕西的生态和生产条件差异较大, 育种目标各有侧重, 因此从相同的亲本组合中选出了遗传差异较大的品种。分析各亚群材料的育成年代和地理来源(表4和表5), 发现149份供试材料按育成年代大致可以划分为两大类, 即以2000年为分界点, 之前为一类(I, II, III), 之后为另一类(IV, V), 其原因可能是不同年代的小麦生产水平和主要育种目标各有侧重, 2000年之前以提高产量为主, 之后以产量、品质和抗性的同步提高为主要目标。同时亚群I、II和III的π值均小于亚群IV和V, 表明2000年之后的品种(系)遗传多样性提高。从品种的地理来源上看, 不同地区在品种选育过程中存在基因交流现象, 而同一地区因育种目标、亲本选配及中间亲本材料的不同, 品种间的差异可能较大。本研究149份供试材料的平均基因多样性H为0.336, 遗传相似系数GS为0.619, 核苷酸多样性π值为0.340, 表明蚂蚱麦和小白麦衍生系群体, 以及大面积推广品种的总体遗传多样性水平较低。在今后的育种工作中应进一步加强优异种质资源的引进和应用, 以拓宽新品种的遗传基础, 提高小麦育种水平。
4 结论
黄淮冬麦区的4份蚂蚱麦(D1~D4)与北部冬麦区的小白麦(D5)、燕大1817 (D7)紧密地聚为一类, 6份地方品种之间的遗传相似性很高; 2000年之前的育成品种以蚂蚱麦或小白麦的单一衍生系为主, 2000年之后的多数育成品种同时拥有蚂蚱麦和小白麦血缘, 两大衍生系之间存在基因交流, 提高了遗传多样性; 但蚂蚱麦和小白麦衍生系以及大面积推广品种的总体遗传多样性水平较低, 应加强优异种质资源的引进和应用, 拓宽新品种的遗传基础。附表 请见网络版: 1) 本刊网站http://zwxb.chinacrops.org/; 2) 中国知网http://www.cnki.net/; 3) 万方数据http://c.wanfangdata.com.cn/Periodical-zuowxb. aspx。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}