 ,1,2,*, 王瑞,1,2,*
,1,2,*, 王瑞,1,2,*Genome-wide association of roots, hypocotyls and fresh weight at germination stage under as stress in Brassica napus L.
QU Cun-Min1,2,**, MA Guo-Qiang1,2,**, ZHU Mei-Chen1,2, HUANG Xiao-Hu1,2, JIA Le-Dong1,2, WANG Shu-Xian1,2, ZHAO Hui-Yan1,2, XU Xin-Fu1,2, LU Kun1,2, LI Jia-Na,1,2,*, WANG Rui,1,2,*通讯作者:
第一联系人:
收稿日期:2018-07-5接受日期:2018-10-8网络出版日期:2018-11-06
| 基金资助: |
Received:2018-07-5Accepted:2018-10-8Online:2018-11-06
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (2787KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
曲存民, 马国强, 朱美晨, 黄小虎, 贾乐东, 王书贤, 赵会彦, 徐新福, 卢坤, 李加纳, 王瑞. 砷胁迫下甘蓝型油菜苗期根、下胚轴和鲜重的全基因组关联分析[J]. 作物学报, 2019, 45(2): 175-187. doi:10.3724/SP.J.1006.2019.84093
QU Cun-Min, MA Guo-Qiang, ZHU Mei-Chen, HUANG Xiao-Hu, JIA Le-Dong, WANG Shu-Xian, ZHAO Hui-Yan, XU Xin-Fu, LU Kun, LI Jia-Na, WANG Rui.
随着工农业和城市化进程的发展, 以及化肥农药的不合理利用, 使我国农田土壤日益恶化。砷是广泛存在于自然界的一种微量元素, 有剧毒且有致癌作用, 同时会抑制植物生长, 从而严重影响作物产量, 且可食部分砷的积累会对食物链造成污染[1,2]。植物响应重金属元素胁迫的机制包括阻止和控制重金属的吸收、体内螯合解毒、体内区室化分隔以及代谢平衡等生物学过程[3]。在土壤中, 砷主要以砷酸盐和亚砷酸盐的形式存在, 在有氧条件下, 植物从土壤中吸收利用的主要是砷酸盐, 而在厌氧条件下, 亚砷酸盐的吸收占主导地位[4]。研究表明, 磷酸盐转运蛋白(Pht)和水通道蛋白的亚家族(NIPs)是砷酸盐进入植物体的主要通道蛋白, 拟南芥中NIP1;1、NIP1;2、NIP3;1、NIP5;1和NIP7;1都是参与As3+吸收和转运的重要基因[5,6,7], 在水稻中, OsNIP3;2参与侧根对As3+吸收和转运[8]; OsNIP1;1和OsNIP3;3均具有运输As3+的能力, 过量表达后水稻地上部的砷含量显著受到抑制[9]。然而大多数植物体内对重金属的解毒途径是以谷胱甘肽为还原剂, 在砷酸盐氧化还原酶类作用下将体内的砷酸盐还原成亚砷酸盐, 从而降低其毒性[4]。在拟南芥中, 还原酶ATQ1/HAC1突变显著增加了其对砷酸盐的敏感性, 同时显著降低了亚砷酸盐和砷酸盐的比率, 从而使As3+从根部流出的能力降低[10]。另外, 植物体内的金属硫蛋白(metallotioneins, MTs) 和植物螯合蛋白 (PCs) 等与重金属形成螯合物质, 并在ATP结合盒(ABC)转运蛋白作用下转移到液泡中, 能够缓解As3+对植物细胞的毒害[11]。同时, 为了应对胁迫, 植物体内产生的抗氧化酶(CAT、POD、SOD、APX)及非酶抗氧化剂[谷胱甘肽(GSH)、抗坏血酸(AsA)等]能够消除自由基, 抵抗ROS对细胞的损伤, 同时会主动积累一些可溶性溶质, 如可溶性蛋白、可溶性糖等来降低胞内渗透势, 以保证重金属胁迫条件下水分的正常供应, 维持细胞正常的生理功能[11]。
十字花科植物是用于植物修复重金属污染土壤的理想物种[12,13]。油菜作为良好的冬闲田作物, 因具有生长速度快、生物量高、对重金属有较强的耐受性及吸收积累能力等特点, 被认为是修复土壤重金属污染的优良作物之一, 但研究主要集中于Cd、Cu、Zn等重金属离子方面[12,14-15]。此外, 宋俊英等[16]通过对不同甘蓝型油菜和芥菜型油菜品种的水培试验筛选获得砷排异型品种, 证明低浓度的砷胁迫在一定程度上能够促进排异型油菜的生长, 并增加其产量, 但具体的分子机制有待进一步分析。为解析砷胁迫下影响甘蓝型油菜耐砷性的关键位点和候选基因, 本研究对140份甘蓝型油菜在砷胁迫后的相对根长、相对下胚轴长和相对鲜重进行了全基因组关联分析, 确定显著关联的SNP标记和候选区间, 进一步筛选控制性状变异的候选基因, 并通过实时荧光定量PCR验证候选基因在砷胁迫下表达的特性, 明确其基本的生物学功能。本研究为甘蓝型油菜耐砷胁迫的油菜资源的鉴定提供分子标记, 对于砷污染土壤的修复及砷污染土壤上农产品的安全生产具有重要意义。
1 材料与方法
1.1 供试材料
140份甘蓝型油菜材料(附表1)的遗传背景来源广泛, 其中国内品种131份, 大部分在长江流域的重庆、四川、湖北、湖南等地种植, 国外品种9份, 主要在加拿大和德国种植。上述材料均由西南大学重庆市油菜工程技术研究中心保存提供。1.2 试验设计及表型数据考察
随机选取10份材料, 分别用不同浓度的砷酸钠溶液(0、2.5、5、7.5、10、15和20 mg L-1)预处理, 以获得最佳处理浓度(15 mg L-1)。然后选取每份材料健康饱满的100粒种子, 共分为2组。以蒸馏水为对照, 分别将材料播种于培养盘中, 并用保鲜膜封口。将培养盘置培养间, 培养条件为昼夜温度为25℃, 光照/黑暗时间为16 h/8 h, 光照强度为100 μmol m-2 s-1, 相对湿度为60%[17]。培养7 d后, 选取每份材料长势一致的幼苗5株照相, 用AdobeScan移动应用程序读取根和下胚轴长, 用万分之一天平分别称量5株幼苗的鲜重和干重。各表型性状的相对值 = 处理组测定值/对照组测定值[18], 其中相对根长(relative root length)、相对下胚轴长(relative hypocotyl length)和相对鲜重(relative fresh weight)分别用RRL、RHL和RFW表示。
1.3 供试材料基因型、群体结构及亲缘关系分析
参照卢坤等[19]方法, 利用油菜60K SNP芯片对140份甘蓝型油菜材料进行SNP基因型分析, 最终获得32,542个在甘蓝型油菜基因组中具有唯一位置的SNP 标记(MAF < 0.05)用于群体性状的关联分析。基于贝叶斯数学模型, 利用Structure V2.3.4软件[20]对140份材料的关联群体进行群体结构分析。假设群体内存在的亚群数目K的范围为1~10, 运用该软件对每个K值进行5次模拟运算, 将模拟参数迭代(length of burn-in period)和蒙特卡罗迭代(markov chain monte carlo MCMC)均设置为10,000次循环, 并在混合模型下运算。最后根据STRUCTUREV 2.3.4软件运算得到的后验概率值和2个连续的后验概率值的变化速率(ΔK)来确定群体中存在的亚群数目[21]。利用SPAGeDi v1.4 软件进行亲缘关系(kinship)分析, 并计算亲缘关系值的矩阵(K矩阵)[22]。
1.4 全基因组关联分析与LD区间分析
参照Wang等[23]方法, 利用R语言程序包的MRMLM (Multi-Locus Random-SNP-Effect Mixed Linear Model)方法进行砷胁迫相关性状 GWAS分析, 参数设置均为默认值[23,24]。显著关联SNP标记阈值以1/标记数设定为1/32,542 = 3.0E-5, 同时采用Haploview 4.2计算显著关联SNP所在染色体上的LD区间, 设定HW阈值(Hardy Weinberg P-value cutoff)为0.001, 非缺失标记的比例为75%, MAF为0.05, 参照卢坤等[19]方法进行, 最终以显著关联的SNP所在的单倍型块作为候选基因预测区间, SNP标记未在单倍型块内的, 则以标记两侧100 kb侧翼序列作为LD候选区域, 用于候选基因的预测和功能注释。
1.5 候选基因预测与qRT-PCR分析
根据已知的甘蓝型油菜基因组测序数据库(http://www.genoscope.cns.fr/blat-server/cgi-bin/colza/webBlat)信息[25], 筛选获得LD区间内的候选基因, 利用Geneious 4.8.5软件进行本地BlastP分析, 与拟南芥进行BlastP比对分析的阈值E-value ≤ 1E-10, 最终以获得的同源性最高的拟南芥功能基因注释候选基因功能[19]。为进一步明确候选基因的功能, 利用qRT-PCR方法, 检测砷胁迫后候选基因在根、下胚轴和子叶中表达量变化差异。参照Zhou等[26]方法提取根、下胚轴和叶片总RNA, 合成cDNA和进行qRT-PCR扩增, 反应结束后, 根据参照基因BnACT7用2-ΔΔCT法计算目的基因相对表达量, 3次重复。候选基因扩增的特异性引物源自qPrimerDB数据库[27] (表1)。
Table 1
表1
表1候选基因qRT-PCR特异性引物序列
Table 1
| 基因名 Gene name | 基因号 Gene ID | 引物序列 Sequences of primers (5'-3') |
|---|---|---|
| BnaABCG25 | BnaA02g15690D | F: GTGGCACTCAGATTATCGAGACGTACA R: AGTCCGAGTCCTTGGGATGCTAAAA |
| BnaGSTU10 | BnaA07g31660D | F: ATCGATGAAACCTGGAAGAACT R: GAATCTCTCTCTAGCCACTTCC |
| BnaGST16 | BnaA02g20640D | F: GCAGGTATCAAAGTTTTCGGACACG R: GTGGGATATGTTCTTGGAGTCGGCT |
| BnaPHT3;3 | BnaA06g25890D | F: GTGAAGCAACAGAAAGTAACGGTGGT R: CCGCCGCTGTTGTTGTTGTTAC |
| BnaPHT1;9 | BnaA07g32730D, BnaA07g32740D, BnaA07g32750D | F: CTCCACGGCCGTGACCTCTTC R: AACCCCATTATCTGAATCTTGACTCG |
| BnaGSTU25 | BnaA09g45070D, BnaA09g45080D, BnaA09g45090D | F: TCCTTCCTTCTGATCCTTACCA R: GGACACTCTGCTTCGATGCTAA |
| BnaGSTU12 | BnaC06g31020D, BnaC06g31030D | F: TCAGATCCTTCCATCCTCCCCTCA R: CCACCATAGCCCCACAAGCGAT |
| BnaGSTU11 | BnaC06g31040D | F: CCCCGTCCATAAACAGATCCCC R: CGCTATTGCCATCTTTCTTTCCTCC |
| BnaUBC21 | BnaA06g27860D, BnaA09g04490D | F: CCTCTGCAGCCTCCTCAAGT R: CATATCTCCCCTGTCTTGAAATGC |
| BnaACTIN7 | BnaA03g55890D, BnaC02g00690D, BnaA10g22340D | F: CCCTGGAATTGCTGACCGTA R: TGGAAAGTGCTGAGGGATGC |
新窗口打开|下载CSV
2 结果与分析
2.1 砷胁迫相关性状的表型变异分析
在正常和砷胁迫处理下(表2), 油菜根长的变异幅度分别介于1.41~15.80 cm和0.61~13.39 cm之间, 变异系数分别为25.91%和28.91%, 其相关系数为0.986 (P<0.01); 下胚轴长变异幅度分别介于2.68~ 7.99 cm和1.75~6.48 cm之间, 变异系数分别为16.87%和17.79%, 其相关系数为0.955 (P<0.01); 鲜重变异幅度分别为0.026~0.093 g和0.019~0.087 g之间, 变异系数分别为21.93%和26.68%, 相关系数为0.996 (P<0.01), 上述结果表明, 甘蓝型油菜发芽期根长、下胚轴长和鲜重在砷胁迫后受到不同程度的抑制作用, 以相对根长、相对下胚轴长和相对鲜重作为油菜受抑制程度的衡量指标, 其变异系数分别为31.09%、20.62%和20.27%, 说明受砷胁迫后供试材料在萌发期存在较大的性状变异。Table 2
表2
表2砷胁迫下甘蓝型油菜苗期性状统计分析
Table 2
| 性状 Trait | 材料数Number of accessions | 均值±标准差Mean±SD | 最小值Minimum | 中位数Median | 最大值Maximum | 偏度 Skewness | 峰度 Kurtosis | 变异系数 CV (%) | 相关系数Correlation coefficient |
|---|---|---|---|---|---|---|---|---|---|
| CRL | 140 | 8.898±2.306 | 1.410 | 9.065 | 15.800 | -0.135 | 0.958 | 25.91 | 0.986** |
| TRL | 137 | 7.823±2.226 | 0.610 | 7.767 | 13.386 | -0.571 | 1.053 | 28.91 | |
| RRL | 137 | 0.903±0.280 | 0.061 | 0.876 | 1.686 | 0.409 | 0.950 | 31.09 | |
| CHL | 140 | 5.735±0.968 | 2.680 | 5.695 | 7.990 | -0.338 | 1.036 | 16.87 | 0.995** |
| THL | 137 | 4.453±0.792 | 1.755 | 4.473 | 6.477 | -0.155 | 1.060 | 17.79 | |
| RHL | 137 | 0.793±0.163 | 0.257 | 0.779 | 1.437 | 0.816 | 3.165 | 20.62 | |
| CFW | 140 | 0.056±0.012 | 0.026 | 0.055 | 0.093 | 0.382 | 0.379 | 21.93 | 0.996** |
| TFW | 138 | 0.047±0.013 | 0.019 | 0.048 | 0.087 | -0.409 | 2.227 | 26.68 | |
| RFW | 138 | 0.863±0.174 | 0.160 | 0.867 | 1.326 | -0.318 | 1.461 | 20.27 |
新窗口打开|下载CSV
统计分析(图1)表明, 各性状值均呈连续性变异, 符合多基因控制的数量性状遗传特点, 适于用GWAS方法进行有效的基因定位分析。
图1
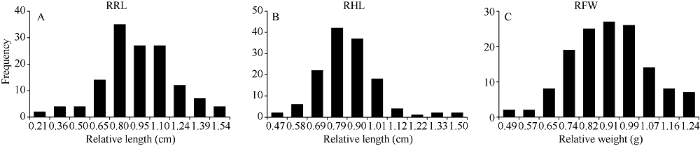 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1砷胁迫下甘蓝型油菜相对根长、下胚轴和鲜重的频次分布
Fig. 1Frequency distribution of RRL, RHL, and RFW of B. napus under As stress treatment
2.2 耐砷胁迫相关性状的全基因组关联分析
用MRMLM模型对60K SNP芯片获得的基因型数据与140份甘蓝型油菜耐砷性指标进行全基因组关联分析(图2), 共获得15个RRL的显著关联SNP位点, 分别位于A02、A03、A04、A05、A06、A07、A08、C02、C03、C05、C06、C07和C08染色体, 单个SNP可解释表型变异的17.31%~24.39% (图2-A和表3); 20个与RHL性状显著关联的SNP位点, 分别位于A01、A02、A03、A07、C02和C04染色体上, 单个SNP位点可解释18.04%~33.82%的表型变异(图2-B和表3); 35个与RFW紧密关联的SNP位点, 分别位于A01、A02、A07、A09、C02、C04和C07染色体上, 其中, 在A02染色体上检测到20个成簇分布, 单个SNP可解释表型变异的20.19%~25.06% (图2-C和表3)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2砷胁迫下甘蓝型油菜相对根长、相对下胚轴长和相对鲜重全基因组关联分析的曼哈顿图
Fig. 2Manhattan plots of GWAS for RRL, RHL, and RFW of rapeseed under As stress treatment
Table 3
表3
表3砷胁迫下甘蓝型油菜相关性状的显著关联SNPs
Table 3
| 性状 Trait | 显著性SNP位点 Significant SNP | 染色体 Chr. | SNP数目 No. of SNP | 区间 Interval (bp) | 等位基因 Allele | P值 P-value | 贡献率 R2 (%) |
|---|---|---|---|---|---|---|---|
| RHL | Bn-A01-p1145674 | A01 | 1 | 649099-849099 | A/G | 2.98E-06 | 19.08 |
| RFW | Bn-A01-p24425211 | A01 | 1 | 20123238-20323238 | C/G | 3.23E-06 | 20.72 |
| RFW | Bn-A02-p8277939 | A02 | 2 | 5250663-5523057 | T/C | 3.51E-06 | 20.70 |
| RFW | Bn-A02-p9422112 | A02 | 4 | 6241447-6919284 | T/C | 8.37E-06 | 20.77 |
| RFW | Bn-A02-p11041638 | A02 | 2 | 7804288-7830004 | A/G | 1.55E-05 | 22.92 |
| RFW | Bn-A02-p11750740 | A02 | 2 | 8461461-8497275 | A/G | 1.99E-05 | 20.19 |
| RFW | Bn-A02-p12278577 | A02 | 3 | 8993675-9490338 | T/C | 2.47E-05 | 22.60 |
| RFW, RHL | Bn-A02-p13019236 | A02 | 3 | 9728636-10424361 | T/G | 6.46E-09 | 21.60 |
| RFW | Bn-A05-p9116236 | A02 | 2 | 12983734-13031064 | T/C | 6.46E-07 | 20.49 |
| RRL, RHL, RW | Bn-A02-p23842763 | A02 | 7 | 19237258-22535850 | T/C | 2.22E-06 | 25.04 |
| RHL | Bn-A03-p8733974 | A03 | 1 | 7923113-8123113 | A/G | 9.44E-06 | 24.37 |
| RRL | Bn-A03-p19129006 | A03 | 2 | 18067901-19247585 | A/G | 1.76E-05 | 20.14 |
| RRL | Bn-A04-p13050240 | A04 | 1 | 13637904-13837904 | T/G | 2.92E-05 | 24.23 |
| RRL | Bn-A05-p2711208 | A05 | 1 | 2708636-2908636 | A/G | 2.98E-05 | 24.39 |
| RDW | Bn-A05-p15227777 | A05 | 1 | 11606312-11806312 | T/C | 3.17E-05 | 19.00 |
| RRL | Bn-A06-p16406113 | A06 | 2 | 17919297-18408096 | T/G | 2.74E-06 | 20.53 |
| RFW, RHL, RRL | Bn-A07-p17382019 | A07 | 6 | 19282162-22517611 | A/G | 6.28E-06 | 21.22 |
| RHL | Bn-A07-p21643815 | A07 | 4 | 23186702-23580640 | A/G | 8.69E-06 | 18.16 |
| RRL | Bn-A08-p17193797 | A08 | 1 | 14579373-14779373 | A/C | 1.66E-05 | 18.82 |
| RFW | Bn-A09-p32954471 | A09 | 5 | 30697653-31316771 | A/G | 1.75E-05 | 20.40 |
| RFW | Bn-A02-p10745711 | C02 | 1 | 13769735-13969735 | A/G | 2.37E-05 | 23.40 |
| RHL, RFW | Bn-C13944312-p158 | C02 | 1 | 15912037-16112037 | T/C | 2.39E-05 | 18.05 |
| RRL | Bn-scaff_16300_1-p870231 | C02 | 1 | 22806371-23006371 | T/C | 2.45E-05 | 18.92 |
| RHL | Bn-scaff_17109_1-p557859 | C02 | 1 | 41708473-41908473 | A/G | 2.46E-05 | 21.79 |
| RRL | Bn-scaff_16182_1-p319123 | C03 | 1 | 51869206-52069206 | T/C | 2.47E-05 | 22.17 |
| RHL | Bn-scaff_16534_1-p1804261 | C04 | 1 | 4356230-4556230 | A/C | 2.52E-05 | 22.65 |
| RFW | Bn-scaff_20567_1-p64644 | C04 | 1 | 20723951-20923951 | A/G | 2.57E-05 | 23.27 |
| RRL | Bn-scaff_16770_1-p684639 | C05 | 1 | 35142768-35342768 | T/G | 2.75E-05 | 17.31 |
| RRL | Bn-scaff_17454_1-p87022 | C06 | 1 | 8199084-8399084 | A/C | 2.78E-05 | 20.28 |
| RHL | Bn-scaff_15743_1-p353143 | C06 | 1 | 27442608-27642608 | T/C | 2.87E-05 | 20.77 |
| RHL | Bn-scaff_16874_1-p269409 | C06 | 4 | 31639304-36835784 | A/C | 2.88E-05 | 26.28 |
| RFW | Bn-scaff_28403_1-p154436 | C07 | 1 | 8625086-8825086 | T/C | 3.14E-05 | 23.05 |
| RRL | Bn-scaff_16110_1-p1058284 | C07 | 1 | 43787584-43987584 | C/G | 3.21E-05 | 22.86 |
| RRL | Bn-scaff_18602_1-p278628 | C08 | 1 | 16338368-16538368 | T/G | 2.34E-05 | 21.11 |
新窗口打开|下载CSV
在检测到的显著关联SNP位点中, 在A02、A07和C02染色体上存在与RRL、RHL和RFW重合的LD连锁区间, 对这些区间的功能注释表明, 可能存在影响甘蓝型油菜参与响应砷胁迫相关的基因位点。
2.3 响应砷胁迫相关候选基因分析
根据已公布的油菜“Darmor-bzh”基因组信息[25], 分别将确定的LD置信区间和未在LD区间内的显著连锁SNP标记上下游各100 kb侧翼序列作为候选区间, 通过本地BlastP将其候选基因蛋白序列比对到拟南芥中进行基因的注释, 筛选出目标基因组区段内与砷相关的油菜同源基因。结果共注释了61个与重金属胁迫或代谢相关的候选基因, 主要包括PHT3;3、PHT1;9、GSTU、SUC1、OTC5、NRAMP1、ZIP12等(表4)。其中在A02染色体上, 与RRL、RHL和RFW均显著关联的LD区间(20.77~23.58 Mb)内注释了一个与重金属转运相关的候选基因(BnaA02g31250D), 而在A02染色体关联SNP标记覆盖的5.25~13.03 Mb候选区段内, 还包括重要候选基因BnaA02g20640D和BnaA02g16380D, 其功能分别为谷胱甘肽转移酶(Glutathione S-Transferase 16, GST16[28])和种子特异性的水孔通道蛋白(ALPHA-TONOPLAST Intrinsic Protein, TIP3[29]), 以及注释基因BnaA02g15690D与种子萌发相关(A. thaliana ATP-Binding Cassette G25, AtABCG25[30])等。在A07染色体与RRL、RHL和RFW均显著关联的LD区间(19.28~23.58 Mb)内, 共注释到16个相关候选基因(表4), 其中4个串联重复基因BnaA07g26010D、BnaA07g26020D、BnaA07g26030D和BnaA07g26040D, 与拟南芥MATE基因家族蛋白为同源基因, 距Bn-A07-p20935217标记下游78.98~ 90.77 kb存在3个串联重复基因BnaA07g32730D、BnaA07g32740D和BnaA07g32750D, 与拟南芥AT1G76430 (phosphate transporter 1, PHT1;9)为同源基因, 其注释功能为与砷离子跨膜运输及磷酸盐离子运输相关(表4), 为本研究的重要候选基因。同时, 还包括GSTU10、GSTU21和GLYI6等相关基因的同源基因(表4), 而在C02染色体的LD置信区间内并未发现注释的相关功能的候选基因。Table 4
表4
表4甘蓝型油菜砷胁迫下相关性状候选基因
Table 4
| 关联性状 Associated trait | 染色体 Chr. | 油菜基因编号 B. napus code | 位置 Position (bp) | 拟南芥基因编号 Arabidopsis AGI No. | 功能描述 Functional description |
|---|---|---|---|---|---|
| RHL | A01 | BnaA01g01280D | 662364 | AT4G36430 | 过氧化物酶超家族蛋白 Peroxidase superfamily protein |
| BnaA01g01600D | 848378 | AT4G35970 | 抗坏血酸过氧化物酶(APX5) ascorbate peroxidase 5 (APX5) | ||
| RFW, RHW, RRL | A02 | BnaA02g10160D | 5151058 | AT5G53650 | 未知功能蛋白 unknown protein |
| BnaA02g10840D | 5596986 | AT4G25630 | 纤维蛋白(FIB2) fibrillarin 2 (FIB2) | ||
| BnaA02g11800D | 6188790 | AT5G49890 | 氯离子通道C (CLC-C) chloride channel C (CLC-C) | ||
| BnaA02g12080D | 6320966 | AT1G65410 | NAP蛋白(NAP11) non-intrinsic ABC protein 11 (NAP11) | ||
| BnaA02g12270D | 6486908 | AT1G65820 | 谷胱甘肽s-转移酶 microsomal glutathione s-transferase | ||
| BnaA02g12430D | 6638194 | AT1G66200 | 谷氨酰胺合成酶(GSR2) glutamine synthase clone F11 (GSR2) | ||
| BnaA02g14910D | 8543407 | AT3G29670 | HXXXD 型酰基转移酶 HXXXD-type acyl-transferase family protein | ||
| BnaA02g15690D | 9144596 | AT1G71960 | ATP绑定蛋白类(ABCG25) ATP-binding casette family G25 (ABCG25) | ||
| BnaA02g16380D | 9732696 | AT1G73190 | α-内在蛋白(TIP3) TIP3 | ||
| BnaA02g20570D | 12940679 | AT4G02480 | AAA型ATP酶蛋白 AAA-type ATPase family protein | ||
| BnaA02g20640D | 13009933 | AT2G02930 | 谷胱甘肽s-转移酶(GSTF3) glutathione S-transferase F3 (GSTF3) | ||
| BnaA02g31250D | 22577390 | AT5G27690 | 重金属运输 解毒超家族蛋白 Heavy metal transport detoxification superfamily protein | ||
| BnaA02g31310D | 22604742 | AT3G05580 | 钙调磷酸酶家族蛋白 Calcineurin-like metallo-phosphoesterase superfamily protein | ||
| RHL, RRL | A03 | BnaA03g17000D | 7971502 | AT2G37130 | 过氧化物酶超家族蛋白 Peroxidase superfamily protein |
| BnaA03g17020D | 7983649 | AT2G37170 | 质膜内在蛋白(PIP2B) plasma membrane intrinsic protein 2 (PIP2B) | ||
| BnaA03g17030D | 7987203 | AT2G37170 | 质膜内在蛋白(PIP2B) plasma membrane intrinsic protein 2 (PIP2B) | ||
| BnaA03g17160D | 8044155 | AT2G37300 | 未知功能蛋白 unknown protein | ||
| BnaA03g38620D | 19192627 | AT2G14580 | 基本抗病相关蛋白(PRB1) basic pathogenesis-related protein 1 (PRB1) | ||
| RRL | A05 | BnaA05g05230D | 2731629 | AT4G00430 | 跨膜蛋白(TMP-C) TRANSMEMBRANE PROTEIN C (TMP-C) |
| RRL | A06 | BnaA06g25890D | 17916831 | AT2G17270 | 磷酸盐转运蛋白 phosphate transporter 3 |
| BnaA06g25960D | 17949890 | AT5G66110 | 重金属运输 解毒超家族蛋白 Heavy metal transport detoxification superfamily protein | ||
| BnaA06g26040D | 17979799 | AT5G23310 | 铁超氧化物歧化酶 Fe superoxide dismutase 3 (FSD3) | ||
| RFW, RHL, RRL | A07 | BnaA07g26010D | 19187837 | AT1G64820 | 伴侣排出的家族蛋白 MATE efflux family protein |
| BnaA07g26020D | 19193050 | AT1G64820 | 伴侣排出的家族蛋白 MATE efflux family protein | ||
| BnaA07g26030D | 19199115 | AT1G64820 | 伴侣排出的家族蛋白 MATE efflux family protein | ||
| BnaA07g26040D | 19203891 | AT1G66760 | 伴侣排出的家族蛋白 MATE efflux family protein | ||
| BnaA07g26290D | 19358162 | AT1G67280 | 乙二醛酶 博来霉素抗性蛋白 加双氧酶蛋白 Glyoxalase Bleomycin resistance protein Dioxygenase superfamily protein | ||
| BnaA07g26330D | 19381145 | AT2G18330 | AAA型ATP酶蛋白 AAA-type ATPase family protein | ||
| BnaA07g27500D | 19996403 | AT1G68850 | 过氧化物酶超级家族蛋白 Peroxidase superfamily protein | ||
| BnaA07g31660D | 22053195 | AT1G74590 | 谷胱甘肽s-转移酶(GSTU10) glutathione S-transferase TAU 10 (GSTU10) | ||
| BnaA07g32730D | 22592486 | AT1G76430 | 磷酸盐转运蛋白 phosphate transporter 1 | ||
| BnaA07g32740D | 22596590 | AT1G76430 | 磷酸盐转运蛋白 phosphate transporter 1 | ||
| BnaA07g32750D | 22608377 | AT1G76430 | 磷酸盐转运蛋白 phosphate transporter 1 | ||
| BnaA07g34210D | 23311592 | AT1G78360 | 谷胱甘肽s-转移酶(GSTU21) glutathione S-transferase TAU 21 (GSTU21) | ||
| BnaA07g34260D | 23358782 | AT1G78610 | 电导率敏感通道蛋白(MSL6) mechanosensitive channel of small conductance-like 6 (MSL6) | ||
| BnaA07g34490D | 23426237 | AT1G78900 | 液泡ATP酶(VHA-A) vacuolar ATP synthase subunit A (VHA-A) | ||
| BnaA07g34850D | 23620541 | AT1G79360 | 阳离子转运蛋白(2-Oct) organic cation carnitine transporter 2 (2-Oct) | ||
| BnaA07g34890D | 23632496 | AT1G79410 | 阳离子转运蛋白(5-Oct) organic cation carnitine transporter5 (5-Oct) | ||
| RFW | A09 | BnaA09g44820D | 30722833 | AT1G17810 | β-脂质体内在蛋白(BETA-TIP) beta-tonoplast intrinsic protein (BETA-TIP) |
| BnaA09g44870D | 30765654 | AT2G04040 | 解毒蛋白 TX1 | ||
| BnaA09g44980D | 30813852 | AT1G17500 | ATPase E1-E2型家族蛋白 ATPase E1-E2 type family protein haloacid dehalogenase-like hydrolase family protein | ||
| BnaA09g45070D | 30894204 | AT1G17180 | 谷胱甘肽s-转移酶(GSTU25) glutathione S-transferase TAU 25 (GSTU25) | ||
| BnaA09g45080D | 30902768 | AT1G17180 | 谷胱甘肽s-转移酶(GSTU25) glutathione S-transferase TAU 25 (GSTU25) | ||
| BnaA09g45090D | 30909957 | AT1G17180 | 谷胱甘肽s-转移酶(GSTU25) glutathione S-transferase TAU 25 (GSTU25) | ||
| BnaA09g45650D | 31200643 | AT4G18593 | 双特异性抗病蛋白 dual specificity protein phosphatase-related | ||
| BnaA09g45870D | 31257749 | AT1G14040 | 维持磷锌平衡基因 PHO1;H3 | ||
| BnaA09g45890D | 31281439 | AT1G14040 | 维持磷锌平衡基因 PHO1;H3 | ||
| RRL | C03 | BnaC03g62740D | 51991951 | AT4G18593 | 双特异性抗病蛋白 dual specificity protein phosphatase-related |
| RHL | C04 | BnaC04g06200D | 4438087 | AT2G39350 | ABC-2型转运蛋白 ABC-2 type transporter family protein |
| BnaC04g06210D | 4439047 | AT3G55090 | ABC-2型转运蛋白 ABC-2 type transporter family protein | ||
| RHL | C06 | BnaC06g31020D | 31680823 | AT1G69920 | 谷胱甘肽s-转移酶(GSTU12) glutathione S-transferase TAU 12 (GSTU12) |
| BnaC06g31030D | 31683577 | AT1G69920 | 谷胱甘肽s-转移酶(GSTU12) glutathione S-transferase TAU 12 (GSTU12) | ||
| BnaC06g31040D | 31689290 | AT1G69930 | 谷胱甘肽s-转移酶(GSTU11) glutathione S-transferase TAU 11 (GSTU11) | ||
| BnaC06g32880D | 32943807 | AT1G71880 | 糖基转运蛋白(SUC1) sucrose-proton symporter 1 (SUC1) | ||
| BnaC06g39760D | 36750159 | AT1G79360 | 阳离子转运蛋白(2-Oct) organic cation carnitine transporter 2 (2-Oct) | ||
| BnaC06g39810D | 36768217 | AT1G79410 | 阳离子转运蛋白(5-Oct) organic cation carnitine transporter5 (5-Oct) | ||
| BnaC06g39820D | 36770807 | AT1G79410 | 阳离子转运蛋白(5-Oct) organic cation carnitine transporter5 (5-Oct) | ||
| BnaC06g40180D | 36932959 | AT1G80830 | 吞噬细胞抗性蛋白(NRAMP1) natural resistance-associated macrophage protein 1 (NRAMP1) | ||
| RRL | C08 | BnaC08g10950D | 16367664 | AT5G62160 | 锌转运蛋白(ZIP12) zinc transporter 12 precursor (ZIP12) |
新窗口打开|下载CSV
另外, 候选基因中在A03、A09和C06染色体上存在重要的与重金属离子相关的串联重复基因, 其中A03染色体的2个串联重复基因BnaA03g17020D和BnaA03g17030D与拟南芥水通道活性蛋白AT2G37170 (PIP2B)同源; 在A09染色体的2个串联重复基因BnaC04g06200D和BnaC04g06210D与拟南芥AT2G39350 (ABCG1)和AT3G55090 (ABCG16)为同源基因, 都属于ABC转运蛋白, 3个串联重复基因BnaA09g45070D、BnaA09g45080D和BnaA09g 45090D与拟南芥中的AT1G17180 (GSTU25)为同源基因, 具有谷胱甘肽转移酶活性; C06染色体上的3个串联重复基因均与谷胱甘肽转移酶活性相关, 分别与拟南芥AT1G69920 (GSTU12)和AT1G69930 (GSTU11)为同源基因, 而BnaC06g39810D和BnaC06g39820D与拟南芥AT1G79410 (OTC5)为同源基因, 具有碳水化合物跨膜转运活性。因此, 本研究为深入解析甘蓝型油菜响应砷胁迫的分子机制提供了基础。
2.4 砷胁迫下候选基因的差异表达分析
通过qRT-PCR方法分析了LD候选区间内8个关键候选基因在砷胁迫下的表达模式(图3), 这些候选基因在甘蓝型油菜根、下胚轴和叶中具有不同的表达模式。其中6个基因家族成员在根中的表达量显著高于对照, 这些基因与砷离子的吸收密切相关。基因PHT1;9和GSTU11在下胚轴中显著下调表达, 基因PHT3;3在所有材料中显著上调表达, 但叶中均表现为显著下调表达, 基因PHT1;9、GSTU11和PHT3;3可能为油菜响应砷胁迫的重要基因。图3
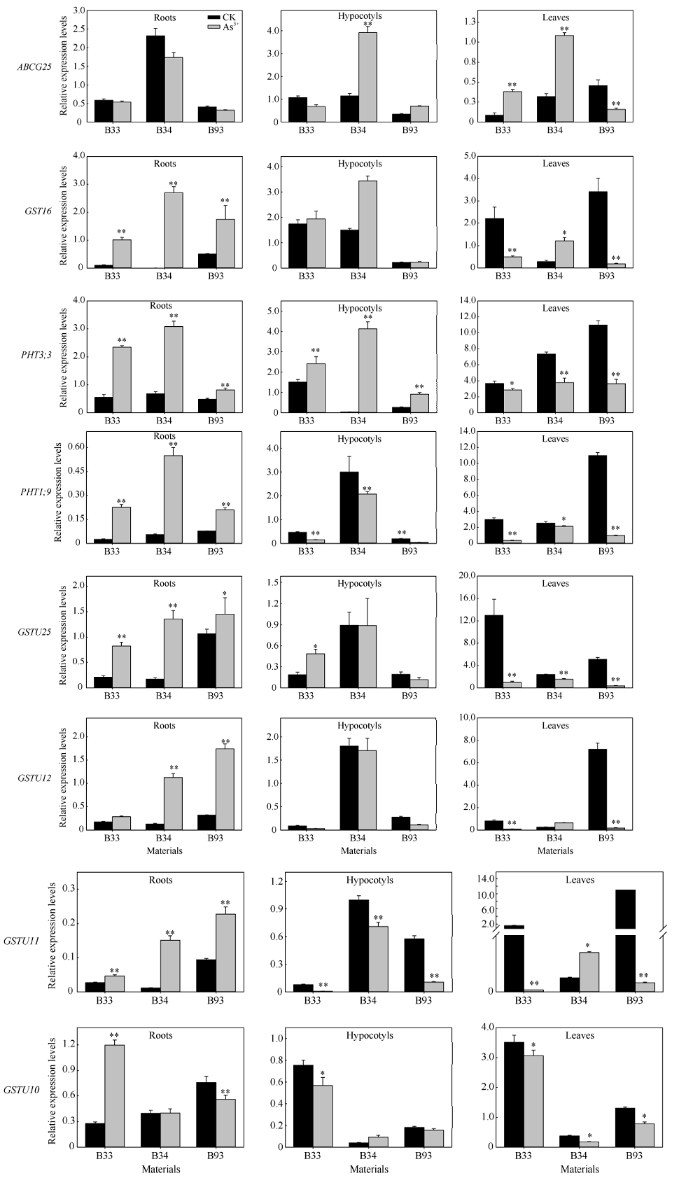 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3砷胁迫下候选基因在甘蓝型油菜中的表达模式分析
误差线表示平均值的标准差(n=3); *和**分别表示在0.05和0.01水平上差异显著。
Fig. 3Expression patterns of candidate genes in B. napus under As stress
Error bar represents the standard error of the mean (n=3); * and ** indicate significant difference at the 0.05 and 0.01 probability levels, respectively.
3 讨论
在自然界中, 砷作为一种非必须微量元素, 被认为是I级致癌物, 对动植物的生长发育和人们的健康产生危害[31]。十字花科植物对重金属镉、铅、锌、汞、砷等均具有较强的耐受性, 但对于甘蓝型油菜响应砷胁迫分子机制的研究相对缺乏。随着油菜60K基因芯片开发及甘蓝型油菜基因组测序的完成[25], 通过GWAS分析并结合基因组信息挖掘油菜重要数量性状的候选基因在油菜研究中已成为常规手段[17,19,32-34]。本研究通过对正常和砷胁迫条件下RRL、RHL和RFW 3个性状的全基因组关联分析, 共检测到67个显著关联的SNP位点, 分别分布在油菜的15条染色体上。其中, A02和A07的关联SNP标记分别与Chen等[35]报道的镉离子相关性状的关联SNP标记位点相近, 关联区间距离分别为202.81 kb和404.37 kb, 很可能为同一位点; 其余位点则与本研究的位点未能重叠, 可能与考察的性状及鉴定的方法存在差异相关联, 也可能是由于甘蓝型油菜响应镉和砷胁迫的分子作用机制存在差异性。通过关联SNP标记在油菜基因组中的物理位置及确定的关联LD置信区间, 根据已公布的甘蓝型油菜“Darmor-bzh”基因组信息[25], 我们共注释了61个可能与响应耐砷胁迫相关的候选基因。在植物中, 研究表明磷酸盐和砷酸盐采用相同吸收系统[36]。同时, 砷酸盐主要通过磷酸盐转运蛋白(Pht)进入植物体, 且与磷酸盐是化学类似物, 在提高磷酸盐含量同时可减少砷的吸收[37,38,39], 从而降低砷对油菜的毒害作用。在A07染色体SNP标记Bn-A07-p20935217下游78.98~90.77 kb区域内注释了3个串联重复基因BnaA07g32730D、BnaA07g32740D和BnaA07g32750D, 与拟南芥AT1G76430 (PHT1;9)为同源基因, 该基因与砷的吸收转运与磷酸盐存在紧密关联性[39,40]。此外, 在A06染色体上SNP标记Bn-A06-p16406113上游24~2254 bp的区间内筛选到一个基因BnaA06g25890D, 其拟南芥同源基因为AT2G17270 (PHT3;3)[41,42], 均具有磷酸盐离子跨膜转运活性。同时在砷胁迫后, PHT1;9和PHT3;3在根中的表达量显著升高, 说明这2个基因可能与砷离子的吸收相关联。另外, PHT3;3在下胚轴中的表达量也显著升高, 说明该基因可能是砷离子吸收转运相关的重要影响因子。
植物在受到高盐、重金属等胁迫时, 体内的植物ATP结合盒(ABC)转运蛋白在离子吸收、累积、转运和外排过程中发挥重要作用[43]。本研究在关联候选区间内注释的6个ABC转运蛋白相关的候选基因(BnaA02g10160D、BnaA02g12080D、BnaA02g15690D、BnaA03g17160D、BnaC04g06200D和BnaC04g06210D)分别与拟南芥AT5G53650 (ABCA)、AT1G65410 (ABCI13)、AT1G71960 (ABCG25)、AT2G37300 (ABCI16)、AT2G39350 (ABCG1)和AT3G55090 (ABCG16)为同源基因。在砷胁迫后, 本研究发现ABCG25在叶中的表达量显著升高, 而在根和下胚轴中的变化不明显(图4), 说明该基因可能参与了砷离子的转运, 其具体的机理有待进一步验证。另外, 谷胱甘肽-S-转移酶(gluthione S-transferase, GST)是植物体内重要的解毒酶类物质, 也是植物螯合肽(Phytochelatin, PC)合成前体, 对重金属有较大的亲和力和重金属离子鳌合能力, 是植物自身解毒机制形成的重要因子[44]。同时, 高浓度的GSH可提高植物体对重金属的耐受能力, 对植物抗重金属过程中的作用进行了广泛研究[45,46], 但是在甘蓝型油菜中的相关报道还较少。本研究在A07, A09和C06染色体的关联LD区间内注释到7个编码谷胱甘肽转移酶相关的候选基因 (BnaA07g31660D、BnaA09g45070D、BnaA09g45080D、BnaA09g45090D、BnaC06g31020D、BnaC06g31030D和BnaC06g31040D)和1个谷胱甘肽巯基转移酶活性相关基因(BnaA02g12270D)。在砷胁迫下, 本研究中, 除GSTU10外, GSTU11、GSTU12、GSTU16和GSTU25在根中显著上调表达(图4), 说明这些基因功能可能与根对砷离子的吸收相关联。然而GSTU11在下胚轴中、GSTU10和GSTU25在叶中均显著下调表达(图4), 说明它们可能与砷离子的吸收转运及降解相关联。因此, 进一步深入开展上述关联候选基因的功能分析将有助于揭示甘蓝型油菜发芽期适应砷胁迫的响应机制, 为甘蓝型油菜重金属耐受性新品种的选育提供理论基础。
4 结论
共获得70个油菜砷胁迫性状相关的显著关联SNP标记位点, 其中与相对根长、相对下胚轴长和相对鲜重显著关联的位点分别为15、20和35个。在显著关联的候选区间内共注释到61个砷胁迫相关的候选基因, 其中PHT3;3和PHT1;9可能是参与甘蓝型油菜砷离子吸收转运的重要候选基因。附表 请见网络版: 1) 本刊网站http://zwxb.chinacrops.org/; 2) 中国知网http://www.cnki.net/; 3) 万方数据http://c.wanfangdata.com.cn/Periodical-zuowxb. aspx。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.3389/fphys.2012.00182URLPMID:22685440 [本文引用: 1]

The two forms of inorganic arsenic, arsenate (AsV) and arsenite (AsIII), are easily taken up by the cells of the plant root. Once in the cell, AsV can be readily converted to AsIII, the more toxic of the two forms. AsV and AsIII both disrupt plant metabolism, but through distinct mechanisms. AsV is a chemical analog of phosphate that can disrupt at least some phosphate-dependent aspects of metabolism. AsV can be translocated across cellular membranes by phosphate transport proteins, leading to imbalances in phosphate supply. It can compete with phosphate during phosphorylation reactions, leading to the formation of AsV adducts that are often unstable and short-lived. As an example, the formation and rapid autohydrolysis of AsV-ADP sets in place a futile cycle that uncouples photophosphorylation and oxidative phosphorylation, decreasing the ability of cells to produce ATP and carry out normal metabolism. AsIII is a dithiol reactive compound that binds to and potentially inactivates enzymes containing closely spaced cysteine residues or dithiol co-factors. Arsenic exposure generally induces the production of reactive oxygen species that can lead to the production of antioxidant metabolites and numerous enzymes involved in antioxidant defense. Oxidative carbon metabolism, amino acid and protein relationships, and nitrogen and sulfur assimilation pathways are also impacted by As exposure. Readjustment of several metabolic pathways, such as glutathione production, has been shown to lead to increased arsenic tolerance in plants. Species- and cultivar-dependent variation in arsenic sensitivity and the remodeling of metabolite pools that occurs in response to As exposure gives hope that additional metabolic pathways associated with As tolerance will be identified.
DOI:10.17521/cjpe.2006.0092URL [本文引用: 1]
植物适应重金属元素胁迫的机制包括阻止和控制重金属的吸收、体内螯合解毒、体内区室化分隔以及代谢平衡等。近年来,随着分子生物学技术在生态学研究中的深入应用,控制这些过程的分子生态机理逐渐被揭示出来。菌根、根系分泌物以及细胞膜是控制重金属进入植物根系细胞的主要生理单元。外生菌根能显著提高寄主植物的重金属耐性,根系分泌物通过改变根际pH、改变金属物质的氧化还原状态和形成络合物等机理减少植物对重金属的吸收。目前,控制菌根和根系分泌物重金属抗性的分子生态机理还不清楚。但细胞膜跨膜转运器已得到深入研究,相关金属离子转运器被鉴定和分离,一些控制基因如铁锌控制运转相关蛋白(ZIP)类、自然抵抗相关巨噬细胞蛋白(Nramp)类、P1B_typeATPase类基因已被发现和克隆。金属硫蛋白(MTs)、植物螯合素(PCs)、有机酸及氨基酸等是植物体内主要的螯合物质,它们通过螯合作用固定金属离子,降低其生物毒性或改变其移动性。与MTs合成相关的MT_like基因已经被克隆,PCs合成必需的植物螯合素合酶(PCS),即γ_Glu_Cys二肽转肽酶(γ_ECS)的编码基因已经被克隆,控制麦根酸合成的氨基酸尼克烟酰胺(NA)在重金属耐性中的作用和分子机理也被揭示出来。ATP结合转运器(ABC)和阳离子扩散促进器(CDF)是植物体内两种主要膜转运器,通过它们和其它跨膜方式,重金属被分隔贮藏于液泡内。控制这些蛋白转运器合成的基因也已经被克隆,在植物中的表达证实其与重金属的体内运输和平衡有关。热休克蛋白(HSP)等蛋白类物质的产生是一种重要的体内平衡机制,其分子机理有待进一步研究。重金属耐性植物在这些环节产生了相关响应基因或功能蛋白质,分子克隆和转基因技术又使它们在污染治理上得到了初步的应用。
DOI:10.17521/cjpe.2006.0092URL [本文引用: 1]
植物适应重金属元素胁迫的机制包括阻止和控制重金属的吸收、体内螯合解毒、体内区室化分隔以及代谢平衡等。近年来,随着分子生物学技术在生态学研究中的深入应用,控制这些过程的分子生态机理逐渐被揭示出来。菌根、根系分泌物以及细胞膜是控制重金属进入植物根系细胞的主要生理单元。外生菌根能显著提高寄主植物的重金属耐性,根系分泌物通过改变根际pH、改变金属物质的氧化还原状态和形成络合物等机理减少植物对重金属的吸收。目前,控制菌根和根系分泌物重金属抗性的分子生态机理还不清楚。但细胞膜跨膜转运器已得到深入研究,相关金属离子转运器被鉴定和分离,一些控制基因如铁锌控制运转相关蛋白(ZIP)类、自然抵抗相关巨噬细胞蛋白(Nramp)类、P1B_typeATPase类基因已被发现和克隆。金属硫蛋白(MTs)、植物螯合素(PCs)、有机酸及氨基酸等是植物体内主要的螯合物质,它们通过螯合作用固定金属离子,降低其生物毒性或改变其移动性。与MTs合成相关的MT_like基因已经被克隆,PCs合成必需的植物螯合素合酶(PCS),即γ_Glu_Cys二肽转肽酶(γ_ECS)的编码基因已经被克隆,控制麦根酸合成的氨基酸尼克烟酰胺(NA)在重金属耐性中的作用和分子机理也被揭示出来。ATP结合转运器(ABC)和阳离子扩散促进器(CDF)是植物体内两种主要膜转运器,通过它们和其它跨膜方式,重金属被分隔贮藏于液泡内。控制这些蛋白转运器合成的基因也已经被克隆,在植物中的表达证实其与重金属的体内运输和平衡有关。热休克蛋白(HSP)等蛋白类物质的产生是一种重要的体内平衡机制,其分子机理有待进一步研究。重金属耐性植物在这些环节产生了相关响应基因或功能蛋白质,分子克隆和转基因技术又使它们在污染治理上得到了初步的应用。
DOI:10.1016/j.tplants.2017.09.015URLPMID:29056439 [本文引用: 2]
Abstract Arsenic is toxic to all life forms and is a potent carcinogen. Its accumulation in crop plants and subsequent consumption poses a serious threat to public health worldwide. Recent developments have enhanced our understanding of the molecular mechanisms governing arsenic uptake, detoxification, and accumulation in plants. In particular, the identification of plant arsenate reductase enzymes and emerging details of the processes underlying arsenic distribution and deposition in the seed will prove invaluable in the development of new strategies to mitigate this threat. Here we provide an outline of these recent developments and suggest new molecular mechanisms that could be employed to reduce arsenic in crops.
[本文引用: 1]
DOI:10.1016/j.molp.2015.01.005URLPMID:25732589 [本文引用: 1]
We report that AtNIP3;1, a member of NIP (nodulin 26-like intrinsic protein) family, functions as the arsenite transporter in Arabidopsis. We demonstrate that AtNIP3;1 plays an important role in both the uptake and root-to-shoot translocation of arsenite under arsenite stress conditions. The findings are important for understanding the molecular mechanisms underlying arsenic distribution in plants.
[本文引用: 1]
DOI:10.1093/jxb/erx165URLPMID:28505352 [本文引用: 1]
Abstract Previous studies have shown that the Nodulin 26-like intrinsic membrane protein (NIP) Lsi1 (OsNIP2;1) is involved in arsenite [As(III)] uptake in rice (Oryza sativa). However, the role of other rice NIPs in As(III) accumulation in planta remains unknown. In the present study, we investigated the role OsNIP3;2 in As(III) uptake in rice. When expressed in Xenopus laevis oocytes, OsNIP3;2 showed a high transport activity for As(III). Quantitative real-time RT-PCR showed that the expression of OsNIP3;2 was suppressed by 5 0008M As(III), but enhanced by 20 and 100 0008M As(III). Transgenic rice plants expressing OsNIP3;2pro-GUS showed that the gene was predominantly expressed in the lateral roots and the stele region of the primary roots. Transient expression of OsNIP3;2:GFP fusion protein in rice protoplasts showed that the protein was localized in the plasma membrane. Knockout of OsNIP3;2 significantly decreased As concentration in the roots, but had little effect on shoot As concentration. Synchrotron microfocus X-ray fluorescence showed decreased As accumulation in the stele of the lateral roots in the mutants compared with wild-type. Our results indicate that OsNIP3;2 is involved in As(III) uptake by lateral roots, but its contribution to As accumulation in the shoots is limited. 0008 The Author 2017. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
URL [本文引用: 1]
DOI:10.1371/journal.pbio.1002009URLPMID:4251824 [本文引用: 1]
Inorganic arsenic is a carcinogen, and its ingestion through foods such as rice presents a significant risk to human health. Plants chemically reduce arsenate to arsenite. Using genome-wide association (GWA) mapping of loci controlling natural variation in arsenic accumulation in Arabidopsis thaliana allowed us to identify the arsenate reductase required for this reduction, which we named High Arsenic Content 1 (HAC1). Complementation verified the identity of HAC1, and expression in Escherichia coli lacking a functional arsenate reductase confirmed the arsenate reductase activity of HAC1. The HAC1 protein accumulates in the epidermis, the outer cell layer of the root, and also in the pericycle cells surrounding the central vascular tissue. Plants lacking HAC1 lose their ability to efflux arsenite from roots, leading to both increased transport of arsenic into the central vascular tissue and on into the shoot. HAC1 therefore functions to reduce arsenate to arsenite in the outer cell layer of the root, facilitating efflux of arsenic as arsenite back into the soil to limit both its accumulation in the root and transport to the shoot. Arsenate reduction by HAC1 in the pericycle may play a role in limiting arsenic loading into the xylem. Loss of HAC1-encoded arsenic reduction leads to a significant increase in arsenic accumulation in shoots, causing an increased sensitivity to arsenate toxicity. We also confirmed the previous observation that the ACR2 arsenate reductase in A. thaliana plays no detectable role in arsenic metabolism. Furthermore, ACR2 does not interact epistatically with HAC1, since arsenic metabolism in the acr2 hac1 double mutant is disrupted in an identical manner to that described for the hac1 single mutant. Our identification of HAC1 and its associated natural variation provides an important new resource for the development of low arsenic-containing food such as rice.
DOI:10.13523/j.cb.20150914URL [本文引用: 2]
Heavy metals are a class of pollutants that may produce certainly toxic actions towards plants, plants have evolved mechanisms of heavy metal stress tolerance in long-term evolution. Based on plant heavy metal tolerance, making a brief overview that the study of tolerance mechanisms of plant response to heavy metal stress at home and abroad in recent years. The molecular mechanism that stress effects of heavy metals on plants, plant antioxidant systems, osmotic adjustment substance including proline, soluble sugar or protein, and different types of gene families under heavy metal are mainly discussed. In order to improve the ability of plant resistance to heavy metals and help phytoremediation.
DOI:10.13523/j.cb.20150914URL [本文引用: 2]
Heavy metals are a class of pollutants that may produce certainly toxic actions towards plants, plants have evolved mechanisms of heavy metal stress tolerance in long-term evolution. Based on plant heavy metal tolerance, making a brief overview that the study of tolerance mechanisms of plant response to heavy metal stress at home and abroad in recent years. The molecular mechanism that stress effects of heavy metals on plants, plant antioxidant systems, osmotic adjustment substance including proline, soluble sugar or protein, and different types of gene families under heavy metal are mainly discussed. In order to improve the ability of plant resistance to heavy metals and help phytoremediation.
DOI:10.1007/s11356-016-6176-5URLPMID:26884243 [本文引用: 2]
This paper analyses the capacity of the rape (Brassica napus) to extract Cd and Zn from the soil and the effect of these metals on the morphometric parameters of the plant (length, weight, surface are
[本文引用: 1]
URL [本文引用: 1]
DOI:10.1016/j.agee.2012.04.006URL [本文引用: 1]
To estimate plant resistance to Cadmium Chloride (CdCl) stress for phytoremediation purposes, the effect of cadmium (Cd) phytotoxicity was assessed on total soluble protein, chlorophyll (Chl) content and antioxidant enzymes in the leaves of three different Brassica napus (B. napus) cultivars, Mohican, Reg. Cob and Okapi. Plants were exposed to three levels of CdCl(0.75, 1.5 and 2.25 mM) in irrigation water. A reduction in protein and Chl content was noted for all treatments in the three cultivars. Generally, superoxide dismutase (SOD) and ascorbate peroxidase (APX) activities were increased with 0.75 mM CdCland then decreased at higher concentrations. SOD activity was enhanced up to 1.5 mM CdCl2 concentration in Mohican cultivar. Moreover, APX activity of Okapi cultivar was increased at a much higher rate of CdCllevels compared to Mohican and Reg. Cob cultivars. Different concentrations of CdClinduced a reduction in the catalase (CAT) activity of Mohican and Reg. Cob. However, this activity was increased with 0.75 mM CdClin Okapi and then decreased with higher concentrations. These results indicate that B. napuscultivars have different tolerances to CdClstress and in consequence, different phytoremediation efficiencies. Moreover, because Okapi possesses a higher antioxidant enzyme activity than the other two cultivars, it is suggested that it is probably the most tolerant cultivar to CdClstress.
DOI:10.7666/d.y1805482URL [本文引用: 1]
砷是广泛存在于自然界的一种微量元素,有剧毒并且有致癌作用。进入环境系统的砷,通过植物积累进入动物和人的体内,给粮食安全生产、人们生活和健康带来了威胁。油菜是我国南方广泛种植的大田作物,在砷污染地区有不同程度的种植。本文在总结了国内外砷对植物生物有效性研究的基础上,通过水培和土壤培养试验相结合的方式,以不同耐性芸薹属植物品种为材料,应用生理生化研究法并结合砷的形态分析技术、根系分泌物收集,深入研究了不同耐性油菜品种对砷胁迫的反应和初步机理。主要研究结果如下: 1.应用水培试验对73个甘蓝型油菜品种和197个芥菜型油菜品种进行筛选。试验结果表明,砷排异型品种有J176、J296、J286、J110、J085、J245、J148、J200、J062、J039、J045、J208、J059、071、041、097,特点是地上部和地下部生物量均较对照增大,砷的冠根比很小,砷主要集中在根系;砷排异敏感型品种有J008、J010、J069、J022、J214、J282、001、025,具有生物量较对照小、砷主要集中在地下部的特点;砷低吸收型品种主要有J179、J135、J098、J237、007、075、091,其生物量较对照增加,但植株砷累积量较少。从这31个品种中选出有代表性的砷排异型品种和砷排异敏感型品种6个,分别为排异型品种J200、J062、071、041和排异敏感型品种J022、025用于进一步进行模拟砷污染土壤盆栽试验的机理研究。 2.在土壤盆栽条件下,研究了砷胁迫对不同耐性油菜生长及产量的影响。结果表明,砷排异型品种(J200、J062、071、041,其中前2个为芥菜型,后两个为甘蓝型)的生物量均出现砷处理高于空白处理的趋势;而排异敏感型品种(J022、025)的生物量及产量则呈现砷处理低于空白处理的趋势。砷排异型品种(J200、J062、071、041)的产量和荚果数均出现上升趋势,其中J200砷处理的产量较空白出现显著性增加,J062单株荚果数增加了47.2%,而砷排异敏感型品种(J022、025)的产量和荚果数均出现下降趋势;同时,砷处理下所有品种的千粒重均有所上升,其中品种J200较对照提高了30.6%。比较不同耐性品种的反应和砷累积情况,可知在土壤砷浓度约100mg/kg时,适合种植排异型油菜。 3.采用水培试验研究了不同耐砷性油菜根系对砷的吸收动力学和根系分泌物反应。试验结果表明,对三价砷来说,敏感型品种的Imax大、Cmin和Km小,而排异型品种的Imax小、Cmin和Km大,均达到显著性差异,敏感型品种表现出较排异型品种更强的三价砷吸附亲和力,而对五价砷的离子亲和力则差异不显著。油菜根系分泌的有机酸主要是草酸和苹果酸。砷胁迫下,排异型品种较敏感型品种分泌的有机酸增加趋势较明显,J062分泌的草酸和苹果酸较对照分别增加了44.5%和30.5%;071分泌的草酸和苹果酸较对照分别增加了45.0%和15.5%。 4.采用砷价态分离技术研究了不同耐砷性油菜体内砷含量和砷价态的变化。结果表明:进入植物体内的砷,主要存在于根系,转移到地上部分的砷含量很低。砷主要以无机三价态和五价态存在于油菜的根系和叶片中,且三价态的砷含量高于五价态砷的含量,其在叶片和根系中的含量分别达到70%和80%以上。敏感型品种(J022、025)中的As(V)显著高于排异型品种,受到的毒害性较大。 5.不同耐砷性油菜酶系统、非酶系统和光合系统对砷污染的反应。结果表明,砷胁迫下,两类不同耐性油菜品种通过酶系统(SOD和CAT)和非酶系统(AsA、GSH和MDA)的反应抵抗或降低砷对作物体本身的伤害。砷处理下,排异型油菜的酶系统和非酶系统均非常活跃,共同作用抵抗砷的毒害;而敏感型的自我调节功能较差,受害严重,025的SOD、MDA含量差异显著,分别出现51.7%的下降和43.61%的增加。 砷胁迫下6个品种的叶绿素含量都有所增加,其中薹期排异型品种的叶绿素a/b出现上升,而敏感型的叶绿素a/b出现下降。砷胁迫提高了排异型品种叶片的光合速率,促进其光合作用,敏感型品种则出现光合速率的下降。
DOI:10.7666/d.y1805482URL [本文引用: 1]
砷是广泛存在于自然界的一种微量元素,有剧毒并且有致癌作用。进入环境系统的砷,通过植物积累进入动物和人的体内,给粮食安全生产、人们生活和健康带来了威胁。油菜是我国南方广泛种植的大田作物,在砷污染地区有不同程度的种植。本文在总结了国内外砷对植物生物有效性研究的基础上,通过水培和土壤培养试验相结合的方式,以不同耐性芸薹属植物品种为材料,应用生理生化研究法并结合砷的形态分析技术、根系分泌物收集,深入研究了不同耐性油菜品种对砷胁迫的反应和初步机理。主要研究结果如下: 1.应用水培试验对73个甘蓝型油菜品种和197个芥菜型油菜品种进行筛选。试验结果表明,砷排异型品种有J176、J296、J286、J110、J085、J245、J148、J200、J062、J039、J045、J208、J059、071、041、097,特点是地上部和地下部生物量均较对照增大,砷的冠根比很小,砷主要集中在根系;砷排异敏感型品种有J008、J010、J069、J022、J214、J282、001、025,具有生物量较对照小、砷主要集中在地下部的特点;砷低吸收型品种主要有J179、J135、J098、J237、007、075、091,其生物量较对照增加,但植株砷累积量较少。从这31个品种中选出有代表性的砷排异型品种和砷排异敏感型品种6个,分别为排异型品种J200、J062、071、041和排异敏感型品种J022、025用于进一步进行模拟砷污染土壤盆栽试验的机理研究。 2.在土壤盆栽条件下,研究了砷胁迫对不同耐性油菜生长及产量的影响。结果表明,砷排异型品种(J200、J062、071、041,其中前2个为芥菜型,后两个为甘蓝型)的生物量均出现砷处理高于空白处理的趋势;而排异敏感型品种(J022、025)的生物量及产量则呈现砷处理低于空白处理的趋势。砷排异型品种(J200、J062、071、041)的产量和荚果数均出现上升趋势,其中J200砷处理的产量较空白出现显著性增加,J062单株荚果数增加了47.2%,而砷排异敏感型品种(J022、025)的产量和荚果数均出现下降趋势;同时,砷处理下所有品种的千粒重均有所上升,其中品种J200较对照提高了30.6%。比较不同耐性品种的反应和砷累积情况,可知在土壤砷浓度约100mg/kg时,适合种植排异型油菜。 3.采用水培试验研究了不同耐砷性油菜根系对砷的吸收动力学和根系分泌物反应。试验结果表明,对三价砷来说,敏感型品种的Imax大、Cmin和Km小,而排异型品种的Imax小、Cmin和Km大,均达到显著性差异,敏感型品种表现出较排异型品种更强的三价砷吸附亲和力,而对五价砷的离子亲和力则差异不显著。油菜根系分泌的有机酸主要是草酸和苹果酸。砷胁迫下,排异型品种较敏感型品种分泌的有机酸增加趋势较明显,J062分泌的草酸和苹果酸较对照分别增加了44.5%和30.5%;071分泌的草酸和苹果酸较对照分别增加了45.0%和15.5%。 4.采用砷价态分离技术研究了不同耐砷性油菜体内砷含量和砷价态的变化。结果表明:进入植物体内的砷,主要存在于根系,转移到地上部分的砷含量很低。砷主要以无机三价态和五价态存在于油菜的根系和叶片中,且三价态的砷含量高于五价态砷的含量,其在叶片和根系中的含量分别达到70%和80%以上。敏感型品种(J022、025)中的As(V)显著高于排异型品种,受到的毒害性较大。 5.不同耐砷性油菜酶系统、非酶系统和光合系统对砷污染的反应。结果表明,砷胁迫下,两类不同耐性油菜品种通过酶系统(SOD和CAT)和非酶系统(AsA、GSH和MDA)的反应抵抗或降低砷对作物体本身的伤害。砷处理下,排异型油菜的酶系统和非酶系统均非常活跃,共同作用抵抗砷的毒害;而敏感型的自我调节功能较差,受害严重,025的SOD、MDA含量差异显著,分别出现51.7%的下降和43.61%的增加。 砷胁迫下6个品种的叶绿素含量都有所增加,其中薹期排异型品种的叶绿素a/b出现上升,而敏感型的叶绿素a/b出现下降。砷胁迫提高了排异型品种叶片的光合速率,促进其光合作用,敏感型品种则出现光合速率的下降。
DOI:10.3864/j.issn.0578-1752.2017.01.002URL [本文引用: 2]
【目的】解析甘蓝型油菜发芽期根和下胚轴发育及耐盐性的调控位点,筛选油菜耐盐性相关的候选基因,可为油菜耐盐性改良提供依据。【方法】以317份具有代表性的甘蓝型油菜自交系为材料,在正常生长和盐胁迫条件下进行沙培鉴定,利用芸薹属60K SNP芯片和全基因组关联分析鉴定正常生长与盐胁迫下甘蓝型油菜发芽期根和下胚轴长度显著关联的SNP,并确定其连锁不平衡区间。通过区间内基因的功能注释及盐胁迫下油菜幼苗根和叶片转录组差异表达基因筛选连锁不平衡区间内的重要候选基因,并以实时荧光定量PCR分析候选基因的组织特异性和盐胁迫诱导表达模式,提高候选基因筛选的准确性。【结果】正常生长和盐胁迫下甘蓝型油菜发芽期下胚轴和根长在不同材料间变异较大,频次分布表明目标性状均为数量性状,受多基因调控。全基因组关联分析模型比较表明,MLM+P+K模型为最优模型。以此模型对目标性状进行全基因组关联分析,检测到45个显著关联SNP,其中40个与下胚轴长度显著关联,5个与根长显著关联,单个SNP解释的表型变异分别为9.12%—14.46%和7.67%—8.93%。重复检测的显著相关SNP中,值得注意的是C04染色体的rs8970,同时与4个性状显著关联,表型贡献率为7.67%—12.35%,是唯一在下胚轴长和根长间重复检测到的显著关联SNP。11个重要关联SNP中有6个位于10—442 kb的连锁不平衡区块中。转录组分析表明,11个连锁不平衡区间共包含447个基因,其中15个受盐胁迫诱导表达。转录组和基因功能注释综合分析表明,BnaSRO1、BnaPAGR2、BnaNPH3、BnaMYB124、BnaSAM-Mtase、BnaBIN2、BnaUMAMIT11、BnaEXPA7、BnaRPT3、BnaEF-hand和BnaF3H很可能为各自区间的候选基因。实时荧光定量PCR结果证实除BnaNPH3外,其他基因均在根或下胚轴中受盐胁迫诱导上调表达。组织特异性分析还发现BnaUMAMIT11、BnaPAGR2和BnaEXPA7主要在萌发的根和下胚轴中特异表达,BnaRPT3、BnaBIN2和BnaMYB124虽然呈组成型表达,但在萌发阶段的下胚轴中表达量最高,证实这些基因很可能参与油菜发芽期根和下胚轴生长发育及耐盐性的调节。【结论】全基因组关联分析共鉴定出45个控制油菜发芽期根和下胚轴发育及耐盐性的显著关联SNP。连锁不平衡、转录组和基因功能注释综合分析初步鉴定出11个重要候选基因。
DOI:10.3864/j.issn.0578-1752.2017.01.002URL [本文引用: 2]
【目的】解析甘蓝型油菜发芽期根和下胚轴发育及耐盐性的调控位点,筛选油菜耐盐性相关的候选基因,可为油菜耐盐性改良提供依据。【方法】以317份具有代表性的甘蓝型油菜自交系为材料,在正常生长和盐胁迫条件下进行沙培鉴定,利用芸薹属60K SNP芯片和全基因组关联分析鉴定正常生长与盐胁迫下甘蓝型油菜发芽期根和下胚轴长度显著关联的SNP,并确定其连锁不平衡区间。通过区间内基因的功能注释及盐胁迫下油菜幼苗根和叶片转录组差异表达基因筛选连锁不平衡区间内的重要候选基因,并以实时荧光定量PCR分析候选基因的组织特异性和盐胁迫诱导表达模式,提高候选基因筛选的准确性。【结果】正常生长和盐胁迫下甘蓝型油菜发芽期下胚轴和根长在不同材料间变异较大,频次分布表明目标性状均为数量性状,受多基因调控。全基因组关联分析模型比较表明,MLM+P+K模型为最优模型。以此模型对目标性状进行全基因组关联分析,检测到45个显著关联SNP,其中40个与下胚轴长度显著关联,5个与根长显著关联,单个SNP解释的表型变异分别为9.12%—14.46%和7.67%—8.93%。重复检测的显著相关SNP中,值得注意的是C04染色体的rs8970,同时与4个性状显著关联,表型贡献率为7.67%—12.35%,是唯一在下胚轴长和根长间重复检测到的显著关联SNP。11个重要关联SNP中有6个位于10—442 kb的连锁不平衡区块中。转录组分析表明,11个连锁不平衡区间共包含447个基因,其中15个受盐胁迫诱导表达。转录组和基因功能注释综合分析表明,BnaSRO1、BnaPAGR2、BnaNPH3、BnaMYB124、BnaSAM-Mtase、BnaBIN2、BnaUMAMIT11、BnaEXPA7、BnaRPT3、BnaEF-hand和BnaF3H很可能为各自区间的候选基因。实时荧光定量PCR结果证实除BnaNPH3外,其他基因均在根或下胚轴中受盐胁迫诱导上调表达。组织特异性分析还发现BnaUMAMIT11、BnaPAGR2和BnaEXPA7主要在萌发的根和下胚轴中特异表达,BnaRPT3、BnaBIN2和BnaMYB124虽然呈组成型表达,但在萌发阶段的下胚轴中表达量最高,证实这些基因很可能参与油菜发芽期根和下胚轴生长发育及耐盐性的调节。【结论】全基因组关联分析共鉴定出45个控制油菜发芽期根和下胚轴发育及耐盐性的显著关联SNP。连锁不平衡、转录组和基因功能注释综合分析初步鉴定出11个重要候选基因。
DOI:10.1023/A:1024553303144URL [本文引用: 1]
Fast and effective glasshouse screening techniques that could identify genetic variation in salinity tolerance were tested. The objective was to produce screening techniques for selecting salt-tolerant progeny in breeding programs in which genes for salinity tolerance have been introduced by either conventional breeding or genetic engineering. A set of previously unexplored tetraploid wheat genotypes, from five subspecies of Triticum turgidum, were used in a case study for developing and validating glasshouse screening techniques for selecting for physiologically based traits that confer salinity tolerance. Salinity tolerance was defined as genotypic differences in biomass production in saline versus non-saline conditions over prolonged periods, of 3 4 weeks. Short-term experiments (1 week) measuring either biomass or leaf elongation rates revealed large decreases in growth rate due to the osmotic effect of the salt, but little genotypic differences, although there were genotypic differences in long-term experiments. Specific traits were assessed. Na+ exclusion correlated well with salinity tolerance in the durum subspecies, and K+/Na+ discrimination correlated to a lesser degree. Both traits were environmentally robust, being independent of root temperature and factors that might influence transpiration rates such as light level. In the other four T. turgidum subspecies there was no correlation between salinity tolerance and Na+ accumulation or K+/Na+ discrimination, so other traits were examined. The trait of tolerance of high internal Na+ was assessed indirectly, by measuring chlorophyll retention. Five landraces were selected as maintaining green healthy leaves despite high levels of Na+ accumulation. Factors affecting field performance of genotypes selected by trait-based techniques are discussed.
DOI:10.3724/SP.J.1006.2016.000344URL [本文引用: 4]
角果是油菜重要的光合作用和种子存储器官,对油菜产量具有重要贡献。本研究以412份具有代表性的甘蓝型油菜品种(系)为材料,利用芸薹属60K Illumina Infinium SNP芯片对其基因型分析,并对油菜结角高度和角果层厚度进行全基因组关联分析。结果共检测到16个显著关联的SNP,其中重庆环境下分别检测到2个和4个SNP与结角高度和结角层厚度显著关联,单个SNP解释的表型变异为5.61%~5.69%和5.94%~6.31%。云南环境下分别检测到5个和1个显著关联的SNP,单个标记解释的表型变异为12.66%~13.97%和22.43%。对2个环境的结角高度差和结角层厚度差共检测到3个和1个与性状显著相关的SNP,它们对表型变异的解释率分别为17.33%~20.32%和29.05%。其中,环境间结角厚度差的关联SNP与重庆环境结角层厚度的1个显著关联SNP位于同一LD区间。各显著关联标记LD区段的多个基因调节植物细胞组织发生、花分生组织发育、角果数目和多器官发育,如NSN1、TPST和SACl等,它们可能通过上述功能影响油菜花序或角果的生长发育,导致结角高度或结角层厚度差异。本研究发掘的这些位点和候选基因可作为影响油菜结角高度和角果层厚度的重要候选区域和基因,为揭示油菜结角性状的遗传基础和分子机制,提高油菜单位面积产量奠定了基础。
DOI:10.3724/SP.J.1006.2016.000344URL [本文引用: 4]
角果是油菜重要的光合作用和种子存储器官,对油菜产量具有重要贡献。本研究以412份具有代表性的甘蓝型油菜品种(系)为材料,利用芸薹属60K Illumina Infinium SNP芯片对其基因型分析,并对油菜结角高度和角果层厚度进行全基因组关联分析。结果共检测到16个显著关联的SNP,其中重庆环境下分别检测到2个和4个SNP与结角高度和结角层厚度显著关联,单个SNP解释的表型变异为5.61%~5.69%和5.94%~6.31%。云南环境下分别检测到5个和1个显著关联的SNP,单个标记解释的表型变异为12.66%~13.97%和22.43%。对2个环境的结角高度差和结角层厚度差共检测到3个和1个与性状显著相关的SNP,它们对表型变异的解释率分别为17.33%~20.32%和29.05%。其中,环境间结角厚度差的关联SNP与重庆环境结角层厚度的1个显著关联SNP位于同一LD区间。各显著关联标记LD区段的多个基因调节植物细胞组织发生、花分生组织发育、角果数目和多器官发育,如NSN1、TPST和SACl等,它们可能通过上述功能影响油菜花序或角果的生长发育,导致结角高度或结角层厚度差异。本研究发掘的这些位点和候选基因可作为影响油菜结角高度和角果层厚度的重要候选区域和基因,为揭示油菜结角性状的遗传基础和分子机制,提高油菜单位面积产量奠定了基础。
[本文引用: 1]
DOI:10.1111/mec.2005.14.issue-8URL [本文引用: 1]
DOI:10.1046/j.1471-8286.2002.00305.xURL [本文引用: 1]
DOI:10.1038/srep19444URLPMID:4726296 [本文引用: 2]
Abstract Genome-wide association studies (GWAS) have been widely used in genetic dissection of complex traits. However, common methods are all based on a fixed-SNP-effect mixed linear model (MLM) and single marker analysis, such as efficient mixed model analysis (EMMA). These methods require Bonferroni correction for multiple tests, which often is too conservative when the number of markers is extremely large. To address this concern, we proposed a random-SNP-effect MLM (RMLM) and a multi-locus RMLM (MRMLM) for GWAS. The RMLM simply treats the SNP-effect as random, but it allows a modified Bonferroni correction to be used to calculate the threshold p value for significance tests. The MRMLM is a multi-locus model including markers selected from the RMLM method with a less stringent selection criterion. Due to the multi-locus nature, no multiple test correction is needed. Simulation studies show that the MRMLM is more powerful in QTN detection and more accurate in QTN effect estimation than the RMLM, which in turn is more powerful and accurate than the EMMA. To demonstrate the new methods, we analyzed six flowering time related traits in Arabidopsis thaliana and detected more genes than previous reported using the EMMA. Therefore, the MRMLM provides an alternative for multi-locus GWAS.
DOI:10.1371/journal.pcbi.1005357URLPMID:5308866 [本文引用: 1]
by Cox Lwaka Tamba, Yuan-Li Ni, Yuan-Ming ZhangGenome-wide association study (GWAS) entails examining a large number of single nucleotide polymorphisms (SNPs) in a limited sample with hundreds ...
[本文引用: 4]
DOI:10.3390/genes8100288URLPMID:5664138 [本文引用: 1]
The basic region/leucine zipper motif (bZIP) transcription factor family is one of the largest families of transcriptional regulators in plants. bZIP genes have been systematically characterized in some plants, but not in rapeseed (Brassica napus). In this study, we identified 247BnbZIPgenes in the rapeseed genome, which we classified into 10 subfamilies based on phylogenetic analysis of their deduced protein sequences. TheBnbZIPgenes were grouped into functional clades withArabidopsisgenes with similar putative functions, indicating functional conservation. Genome mapping analysis revealed that theBnbZIPsare distributed unevenly across all 19 chromosomes, and that some of these genes arose through whole-genome duplication and dispersed duplication events. All expression profiles of 247 bZIP genes were extracted from RNA-sequencing data obtained from 17 differentB.napusZS11 tissues with 42 various developmental stages. These genes exhibited different expression patterns in various tissues, revealing that these genes are differentially regulated. Our results provide a valuable foundation for functional dissection of the differentBnbZIPhomologs inB.napusand its parental lines and for molecular breeding studies ofbZIPgenes inB.napus.
DOI:10.1093/nar/gkx725URLPMID:5753361 [本文引用: 1]
Abstract Real-time quantitative polymerase chain reaction (qPCR) is one of the most important methods for analyzing the expression patterns of target genes. However, successful qPCR experiments rely heavily on the use of high-quality primers. Various qPCR primer databases have been developed to address this issue, but these databases target only a few important organisms. Here, we developed the qPrimerDB database, founded on an automatic gene-specific qPCR primer design and thermodynamics-based validation workflow. The qPrimerDB database is the most comprehensive qPCR primer database available to date, with a web front-end providing gene-specific and pre-computed primer pairs across 147 important organisms, including human, mouse, zebrafish, yeast, thale cress, rice and maize. In this database, we provide 3331426 of the best primer pairs for each gene, based on primer pair coverage, as well as 47760359 alternative gene-specific primer pairs, which can be conveniently batch downloaded. The specificity and efficiency was validated for qPCR primer pairs for 66 randomly selected genes, in six different organisms, through qPCR assays and gel electrophoresis. The qPrimerDB database represents a valuable, timesaving resource for gene expression analysis. This resource, which will be routinely updated, is publically accessible at http://biodb.swu.edu.cn/qprimerdb. The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
.
[本文引用: 1]
DOI:10.1093/jxb/erv244URLPMID:4507774 [本文引用: 1]
The tonoplast intrinsic proteins TIP3;1 and TIP3;2 are specifically expressed during seed maturation and localized to the seed protein storage vacuole membrane. However, the function and physiological roles of TIP3s are still largely unknown. The seed performance of TIP3 knockdown mutants was analysed using the controlled deterioration test. The tip3;1/tip3;2 double mutant was affected in seed longevity and accumulated high levels of hydrogen peroxide compared with the wild type, suggesting that TIP3s function in seed longevity. The transcription factor ABSCISIC ACID INSENSITIVE 3 (ABI3) is known to be involved in seed desiccation tolerance and seed longevity. TIP3 transcript and protein levels were significantly reduced in abi3-6 mutant seeds. TIP3;1 and TIP3;2 promoters could be activated by ABI3 in the presence of abscisic acid (ABA) in Arabidopsis protoplasts. TIP3 proteins were detected in the protoplasts transiently expressing ABI3 and in ABI3-overexpressing seedlings when treated with ABA. Furthermore, ABI3 directly binds to the RY motif of the TIP3 promoters. Therefore, seed-specific TIP3s may help maintain seed longevity under the expressional control of ABI3 during seed maturation and are members of the ABI3-mediated seed longevity pathway together with small heat shock proteins and late embryo abundant proteins.
DOI:10.1038/ncomms9113URLPMID:4569717 [本文引用: 1]
Seed germination is a key developmental process that has to be tightly controlled to avoid germination under unfavourable conditions. Abscisic acid (ABA) is an essential repressor of seed germination. In Arabidopsis, it has been shown that the endosperm, a single cell layer surrounding the embryo, synthesizes and continuously releases ABA towards the embryo. The mechanism of ABA transport from the endosperm to the embryo was hitherto unknown. Here we show that four AtABCG transporters act in concert to deliver ABA from the endosperm to the embryo: AtABCG25 and AtABCG31 export ABA from the endosperm, whereas AtABCG30 and AtABCG40 import ABA into the embryo. Thus, this work establishes that radicle extension and subsequent embryonic growth are suppressed by the coordinated activity of multiple ABA transporters expressed in different tissues. Seed germination is repressed by release of the plant hormone abscisic acid (ABA) to the embryo from the surrounding endosperm tissue. Here Kang et al. characterize four different ABA transporters and propose that they act in concert to control ABA release and regulate germination.
DOI:10.1016/j.plaphy.2016.03.032URLPMID:27038600 [本文引用: 1]
61Silicon mitigates arsenic stress through the changes in root traits.61Si addition increased As uptake with simultaneous alleviation of As toxicity.61Histofluorescent staining showed decrease in As induced ROS damage by Si addition.61As02+02Si treatment improved root traits through decrease in MDA and H2O2level.61Altered level of antioxidant enzymes and stress modulators were also observed.
URLPMID:25914704 [本文引用: 1]
Abstract Rapid and uniform seed germination is a crucial prerequisite for crop establishment and high yield levels in crop production. A disclosure of genetic factors contributing to adequate seed vigor would help to further increase yield potential and stability. Here we carried out a genome-wide association study in order to define genomic regions influencing seed germination and early seedling growth in oilseed rape (Brassica napus L.). A population of 248 genetically diverse winter-type B. napus accessions was genotyped with the Brassica 60k SNP Illumina genotyping array. Automated high-throughput in vitro phenotyping provided extensive data for multiple traits related to germination and early vigor, such as germination speed, absolute germination rate and radicle elongation. The data obtained indicate that seed germination and radicle growth are strongly environmentally dependent, but could nevertheless be substantially improved by genomic-based breeding. Conditions during seed production and storage were shown to have a profound effect on seed vigor, and a variable manifestation of seed dormancy appears to contribute to differences in germination performance in B. napus. Several promising positional and functional candidate genes could be identified within the genomic regions associated with germination speed, absolute germination rate, radicle growth and thousand seed weight. These include B. napus orthologs of the Arabidopsis thaliana genes SNOWY COTYLEDON 1 (SCO1), ARABIDOPSIS TWO-COMPONENT RESPONSE REGULATOR (ARR4), and ARGINYL-t-RNA PROTEIN TRANSFERASE 1 (ATE1), which have been shown previously to play a role in seed germination and seedling growth in A. thaliana.
DOI:10.1186/s12864-015-1607-0URLPMID:25962630
Background Harvest index (HI), the ratio of grain yield to total biomass, is considered as a measure of biological success in partitioning assimilated photosynthate to the harvestable product. While crop production can be dramatically improved by increasing HI, the underlying molecular genetic mechanism of HI in rapeseed remains to be shown. Results In this study, we examined the genetic architecture of HI using 35,791 high-throughput single nucleotide polymorphisms (SNPs) genotyped by the Illumina BrassicaSNP60 Bead Chip in an association panel with 155 accessions. Five traits including plant height (PH), branch number (BN), biomass yield per plant (BY), harvest index (HI) and seed yield per plant (SY), were phenotyped in four environments. HI was found to be strongly positively correlated with SY, but negatively or not strongly correlated with PH. Model comparisons revealed that the A???D test (ADGWAS model) could perfectly balance false positives and statistical power for HI and associated traits. A total of nine SNPs on the C genome were identified to be significantly associated with HI, and five of them were identified to be simultaneously associated with HI and SY. These nine SNPs explained 3.42% of the phenotypic variance in HI. Conclusions Our results showed that HI is a complex polygenic phenomenon that is strongly influenced by both environmental and genotype factors. The implications of these results are that HI can be increased by decreasing PH or reducing inefficient transport from pods to seeds in rapeseed. The results from this association mapping study can contribute to a better understanding of natural variations of HI, and facilitate marker-based breeding for HI.
DOI:10.1093/dnares/dsu002URLPMID:24510440 [本文引用: 1]
Association mapping can quickly and efficiently dissect complex agronomic traits. Rapeseed is one of the most economically important polyploid oil crops, although its genome sequence is not yet published. In this study, a recently developed 60K Brassica Infinium?? SNP array was used to analyse an association panel with 472 accessions. The single-nucleotide polymorphisms (SNPs) of the array were in silico mapped using ???pseudomolecules??? representative of the genome of rapeseed to establish their hypothetical order and to perform association mapping of seed weight and seed quality. As a result, two significant associations on A8 and C3 of Brassica napus were detected for erucic acid content, and the peak SNPs were found to be only 233 and 128 kb away from the key genes BnaA.FAE1 and BnaC.FAE1. BnaA.FAE1 was also identified to be significantly associated with the oil content. Orthologues of Arabidopsis thaliana HAG1 were identified close to four clusters of SNPs associated with glucosinolate content on A9, C2, C7 and C9. For seed weight, we detected two association signals on A7 and A9, which were consistent with previous studies of quantitative trait loci mapping. The results indicate that our association mapping approach is suitable for fine mapping of the complex traits in rapeseed.<br>
DOI:10.3389/fpls.2018.00375URL [本文引用: 1]
Cadmium is a potentially toxic heavy metal to human health. Rapeseed (Brassica napusL.), a vegetable and oilseed crop, might also be a Cd hyperaccumulator, but there is little information on this trait in rapeseed. We evaluated Cd accumulation in different oilseed accessions and employed a genome-wide association study to identify quantitative trait loci (QTLs) related to Cd accumulation. A total of 419B. napusaccessions and inbred lines were genotyped with a 60K Illumina Infinium SNP array of Brassica. Wide genotypic variations in Cd concentration and translocation were found. Twenty-five QTLs integrated with 98 single-nucleotide polymorphisms (SNPs) located at 15 chromosomes were associated with Cd accumulation traits. These QTLs explained 3.49 7.57% of the phenotypic variation observed. Thirty-two candidate genes were identified in these genomic regions, and they were 0.33 497.97 kb away from the SNPs. We found orthologs ofArabidopsis thalianalocated near the significant SNPs on theB. napusgenome, including NRAMP6 (natural resistance-associated macrophage protein 6), IRT1 (iron-regulated transporter 1), CAD1 (cadmium-sensitive 1), and PCS2 (phytochelatin synthase 2). Of them, four candidate genes were verified by qRT-PCR, the expression levels of which were significantly higher after exposure to Cd than in the controls. Our results might facilitate the study of the genetic basis of Cd accumulation and the cloning of candidate Cd accumulation genes, which could be used to help reduce Cd levels in edible plant parts and/or create more efficient hyperaccumulators.
[本文引用: 1]
DOI:10.1111/j.1365-313X.2004.02161.xURLPMID:15272879 [本文引用: 1]
Of the mineral nutrients essential for plant growth, phosphorus plays the widest diversity of roles and a lack of phosphorus has profound effects on cellular metabolism. At least eight members of the Arabidopsis Pht1 phosphate (Pi) transporter family are expressed in roots and Pht1;1 and Pht1;4 show the highest transcript levels. The spatial and temporal expression patterns of these two genes show extensive overlap. To elucidate the in planta roles of Pht1;1 and Pht1;4, we identified loss-of-function mutants and also created a double mutant, lacking both Pht1;1 and Pht1;4. Consistent with their spatial expression patterns, membrane location and designation as high-affinity Pi transporters, Pht1;1 and Pht1;4 contribute to Pi transport in roots during growth under low-Pi conditions. In addition, during growth under high-Pi conditions, the double mutant shows a 75% reduction in Pi uptake capacity relative to wildtype. Thus, Pht1;1 and Pht1;4 play significant roles in Pi acquisition from both low- and high-Pi environments.
DOI:10.1104/pp.111.174805URLPMID:21628630 [本文引用: 1]
Phosphorus (P) remobilization in plants is required for continuous growth and development. The Arabidopsis (Arabidopsis thaliana) inorganic phosphate (Pi) transporter Pht1;5 has been implicated in mobilizing stored Pi out of older leaves. In this study, we used a reverse genetics approach to study the role of Pht1;5 in Pi homeostasis. Under low-Pi conditions, Pht1; 5 loss of function (pht1; 5-1) resulted in reduced P allocation to shoots and elevated transcript levels for several Pi starvation-response genes. Under Pi-replete conditions, pht1;5-1 had higher shoot P content compared with the wild type but had reduced P content in roots. Constitutive overexpression of Pht1;5 had the opposite effect on distribution: namely, lower levels in shoots compared with the wild type but higher P content in roots. Pht1;5 overexpression also resulted in altered Pi remobilization, as evidenced by a greater than 2-fold increase in the accumulation of Pi in siliques, premature senescence, and an increase in transcript levels of genes involved in Pi scavenging. Furthermore, Pht1;5 overexpressors exhibited increased root hair formation and reduced primary root growth that could be rescued by the application of silver nitrate (ethylene perception inhibitor) or aminoethoxyvinylglycine (ethylene biosynthesis inhibitor), respectively. Together, these data indicate that Pht1;5 plays a critical role in mobilizing Pi from P source to sink organs in accordance with developmental cues and P status. The study also provides evidence for a link between Pi and ethylene signaling pathways.
[本文引用: 2]
[本文引用: 1]
DOI:10.1371/journal.pone.0043530URLPMID:22937061 [本文引用: 1]
The mitochondrial phosphate transporter (MPT) plays crucial roles in ATP production in plant cells. ThreeMPTgenes have been identified inArabidopsis thaliana. Here we report that the mRNA accumulations ofAtMPTswere up-regulated by high salinity stress inA. thalianaseedlings. And the transgenic lines overexpressingAtMPTsdisplayed increased sensitivity to salt stress compared with the wild-type plants during seed germination and seedling establishment stages. ATP content and energy charge was higher in overexpressing plants than those in wild-typeA. thalianaunder salt stress. Accordingly, the salt-sensitive phenotype of overexpressing plants was recovered after the exogenous application of atractyloside due to the change of ATP content. Interestingly, Genevestigator survey and qRT-PCR analysis indicated a large number of genes, including those related to gibberellin synthesis could be regulated by the energy availability change under stress conditions inA. thaliana. Moreover, the exogenous application of uniconazole to overexpressing lines showed that gibberellin homeostasis was disturbed in the overexpressors. Our studies reveal a possible link between the ATP content mediated by AtMPTs and gibberellin metabolism in responses to high salinity stress inA. thaliana.
[本文引用: 1]
DOI:10.1002/j.1460-2075.1992.tb05431.xURLPMID:1396551 [本文引用: 1]
Abstract In response to heavy metal stress, plants and certain fungi, such as the fission yeast Schizosaccharomyces pombe, synthesize small metal-binding peptides known as phytochelatins. We have identified a cadmium sensitive S. pombe mutant deficient in the accumulation of a sulfide-containing phytochelatin-cadmium complex, and have isolated the gene, designated hmt1, that complements this mutant. The deduced protein sequence of the hmt1 gene product shares sequence identity with the family of ABC (ATP-binding cassette)-type transport proteins which includes the mammalian P-glycoproteins and CFTR, suggesting that the encoded product is an integral membrane protein. Analysis of fractionated fission yeast cell components indicates that the HMT1 polypeptide is associated with the vacuolar membrane. Additionally, fission yeast strains harboring an hmt1-expressing multicopy plasmid exhibit enhanced metal tolerance along with a higher intracellular level of cadmium, implying a relationship between HMT1 mediated transport and compartmentalization of heavy metals. This suggests that tissue-specific overproduction of a functional hmt1 product in transgenic plants might be a means to alter the tissue localization of these elements, such as for sequestering heavy metals away from consumable parts of crop plants.
DOI:10.17521/cjpe.2005.0066URL [本文引用: 1]
由于重金属污染日益严重,重金属在土壤_植物系统中的行为引起了人们的高度重视。高等植物对重金属的耐性与积累性,已经成为污染生态学研究的热点。近年来,由于分子生态学等学科的发展,有关植物对重金属的解毒和耐性机理、重金属离子富集机制的研究取得了较大进展。高等植物对重金属的耐性和积累在种间和基因型之间存在很大差异。根系是重金属等土壤污染物进入植物的门户。根系分泌物改变重金属的生物有效性和毒性,并在植物吸收重金属的过程中发挥重要作用。土壤中的大部分重金属离子都是通过金属转运蛋白进入根细胞,并在植物体内进一步转运至液泡贮存。在重金属胁迫条件下植物螯合肽(PC)的合成是植物对胁迫的一种适应性反应。耐性基因型合成较多的PC ,谷胱甘肽(GSH)是合成PC的前体,重金属与PC螯合并转移至液泡中贮存,从而达到解毒效果。金属硫蛋白(MTs)与PC一样,可以与重金属离子螯合,从而降低重金属离子的毒性。该文从分子水平上论述了根系分泌物、金属转运蛋白、MTs、PC、GSH在重金属耐性及超积累性中的作用,评述了近10年来这方面的研究进展,并在此基础上提出存在的问题和今后研究的重点。
DOI:10.17521/cjpe.2005.0066URL [本文引用: 1]
由于重金属污染日益严重,重金属在土壤_植物系统中的行为引起了人们的高度重视。高等植物对重金属的耐性与积累性,已经成为污染生态学研究的热点。近年来,由于分子生态学等学科的发展,有关植物对重金属的解毒和耐性机理、重金属离子富集机制的研究取得了较大进展。高等植物对重金属的耐性和积累在种间和基因型之间存在很大差异。根系是重金属等土壤污染物进入植物的门户。根系分泌物改变重金属的生物有效性和毒性,并在植物吸收重金属的过程中发挥重要作用。土壤中的大部分重金属离子都是通过金属转运蛋白进入根细胞,并在植物体内进一步转运至液泡贮存。在重金属胁迫条件下植物螯合肽(PC)的合成是植物对胁迫的一种适应性反应。耐性基因型合成较多的PC ,谷胱甘肽(GSH)是合成PC的前体,重金属与PC螯合并转移至液泡中贮存,从而达到解毒效果。金属硫蛋白(MTs)与PC一样,可以与重金属离子螯合,从而降低重金属离子的毒性。该文从分子水平上论述了根系分泌物、金属转运蛋白、MTs、PC、GSH在重金属耐性及超积累性中的作用,评述了近10年来这方面的研究进展,并在此基础上提出存在的问题和今后研究的重点。
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}