关键词:玉米; 幼胚; 活性炭; 转录组 Transcriptome Analysis of Promotive Effects of Active Carbon on Growth and Development of Maize Seedlings from in vitro Cultured Immature Embryos WANG Jin-Ping1,2, SUN Guo-Zhong2,*, WANG Hai-Bo2,* 1College of Agronomy, Agricultural University of Hebei, Baoding 071001, China
2 Institute of Genetics and Physiology, Hebei Academy of Agriculture and Forestry Sciences / Plant Genetic Engineering Center of Hebei Province, Shijiazhuang 050051, China
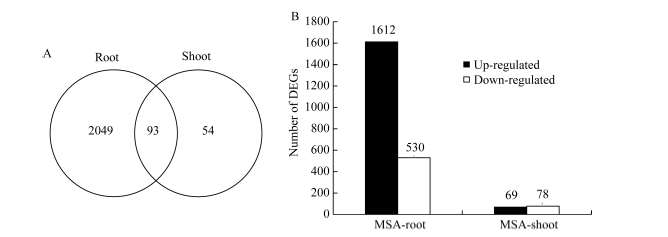
Fund:This study was supported by the Financial Fund Program of Hebei Province (2009055001, F15R25). AbstractImmature embryos from the maize inbred line Chang 7-2 were collected at 14 days after pollination, and cultured on MS or MSA medium (MS medium plus active carbon) for nine days at 24°C. Active carbon significantly accelerated the growth and development of maize seedlings from cultured immature embryos. Using RNA-seq technique, the genes involved in the growth promotive effects of active carbon were analyzed. The presence of active carbon in the medium affected the gene expression in seedlings significantly. Number of up- and down-regulated genes was 1612 and 530 in roots, as well as 69 and 78 in shoots, respectively; indicating that active carbon mainly affects gene expression in roots. GO enrichment analysis showed that differentially expressed genes (DEGs) in roots were mainly involved in DNA packaging, DNA packaging complex and hydrolase activity; the DEGs in shoots were mainly involved in lipid metabolic process, extracellular region and peroxidase activity. The KEGG enrichment analysis showed that the DEGs in roots were significantly associated with energy metabolism, carbohydrate metabolism, lipid metabolism, amino acid metabolism, cell cycle and plant hormone signal transduction. The DEGs in shoots were significantly associated with biosynthesis of ubiquinone and other terpenoid-quinone compounds. Several key genes involved in the cell cycle pathway (i.e., CYC, CDH1, MCM3, PCNA2, and BUB1), signal transduction of auxin ( Aux/IAA) and cytochrome function ( CYP450 oxidase) were significantly up-regulated by active carbon. Ten DEGs were confirmed by Real-time quantitative PCR assay, suggesting that our data and analysis of transcriptome sequencing are reliable.
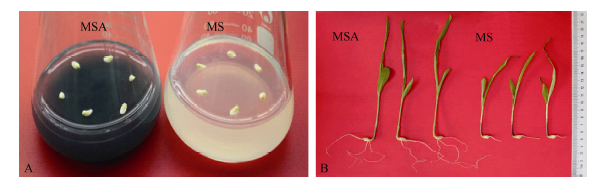
图1 活性炭促进玉米幼胚离体培养成苗 A: 玉米幼胚接种在MS和MSA培养基上; B: 玉米幼胚在MS和MSA培养基上培养9 d后的长势。Fig. 1 Active carbon accelerated the seedlings producing from immature embryos cultured in vitro A: Immature embryos cultured on MS or MSA medium; B: Seedlings developing from immature embryos cultured on MS or MSA for nine days.
表2 Table 2 表2(Table 2)
表2 MS和MSA培养基上玉米幼胚苗性状的比较 Table 2 Comparison of the traits of maize seedlings developing from immature embryos cultured on MS or MSA media
处理 Treatment
根数 Root number
根长 Root length (cm)
株高 Seedling height (cm)
鲜重 Fresh weight (g)
MSA
3.50 ± 0.58 a
9.38 ± 2.42 a
16.15 ± 0.96 a
0.34 ± 0.01 a
MS
1.00 ± 0.00 b
1.48 ± 0.39 b
12.40 ± 1.10 b
0.19 ± 0.01 b
Values within a column followed by different letters are significantly different at the 0.05 probability level. 同一列数值后标不同字母表示在0.05水平上差异显著。
表2 MS和MSA培养基上玉米幼胚苗性状的比较 Table 2 Comparison of the traits of maize seedlings developing from immature embryos cultured on MS or MSA media
图2 活性炭诱导的差异表达基因分布情况 A: 玉米幼胚苗地上部和地下部的DEGs维恩图; B: 玉米幼胚苗地上部和地下部的DEGs数目。Fig. 2 Distribution of DEGs induced by active carbon A: Veen diagram of DEGs in shoots and roots of the maize immature embryo seedlings; B: The total number of DEGs in shoots and roots of maize immature embryo seedlings.
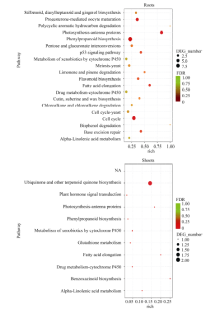
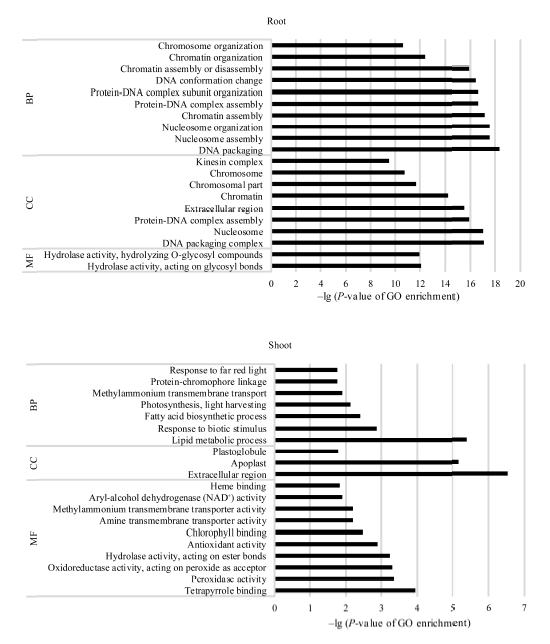
图3 活性炭诱导的玉米胚苗地上部和地下部DEGs中各涉及的20个最显著富集GO分类 BP: 生物学过程; CC: 细胞组分; MF: 分子功能。Fig. 3 Twenty most significantly enriched Gene Ontology (GO) terms of the DEGs induced by active carbon in roots and shoots of maize seedlings BP: biological process; CC: cellular component; MF: molecular function.
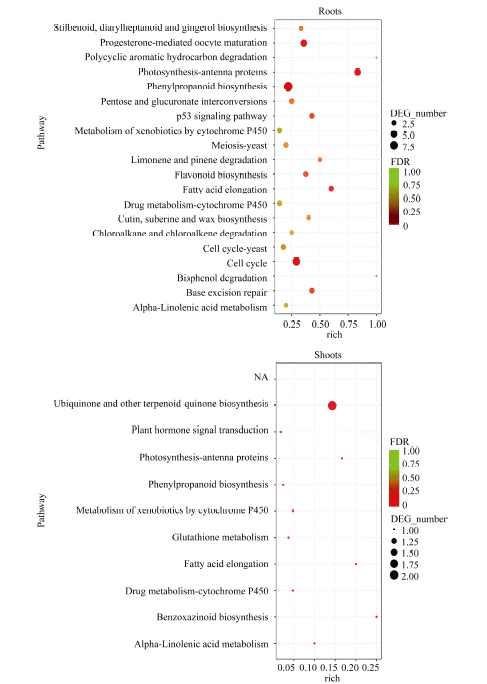
图4 地下部和地上部样品中DEGs富集的KEGG途径 地下部中仅列出前20个显著富集的KEGG pathway。Fig. 4 Enriched KEGG pathways of DEGs in the roots and shoots samples Only twenty most enriched KEGG pathways of DEGs in roots are listed.
微小染色体维持缺陷蛋白 DNA replication licensing factor MCM3 homolog 2
10.24
GRMZM2G009913
有丝分裂检纺锤体监测点蛋白 Mitotic spindle checkpoint protein BUBR1
257.47
外源物质降解代谢 Xenobiotics bio-degradation and metabolism
GRMZM2G057086
细胞色素P450蛋白 Cytochrome P450 (E.C. 1.14.-.-)
55.39
GRMZM2G155502
乙醛脱氢酶 Aldehyde dehydrogenase 3B1 (ALDH 3B1)
184.49
GRMZM2G098346
乙醇脱氢酶 Alcohol dehydrogenase 2 (ADHP2)
0.13
GRMZM2G077206
谷胱氨肽转移酶 Glutathione transferase 27 (GST27)
Inf
GRMZM2G028556
谷胱氨肽转移酶 Glutathione transferase 7 (GST7)
15.85
植物激素信号转导 Plant hormone signal transduction
GRMZM2G006578
bZIP转录因子 Putative bZIP transcription factor superfamily protein (bZIP7)
3.93E-3
GRMZM2G167794
Aux/IAA蛋白 Auxin-responsive protein IAA4
8.11
GRMZM2G366373
Aux/IAA蛋白 Auxin-responsive protein IAA
30.09
MSA/MS represents the ratio of the gene expression level in MSA-roots to the gene expression level in MS-roots; “ Inf” indicates that this gene is only expressed in MSA-roots. MSA/MS表示MSA-roots中基因表达量与MS-roots中的比值; Inf表示该基因在MSA-roots中表达, 在MS-roots中不表达。
表4 细胞周期、植物激素转导和外源物质降解代谢途径中的DEGs Table 4 DEGs functioning in cell cycle, plant hormone signal transduction, xenobiotics bio-degradation and metabolism pathways
图5 受活性炭诱导的10个DEGs的qRT-PCR检测 对MSA-roots中7个上调表达基因和3个下调表达基因进行qRT-PCR验证, 以MSA-roots较MS-roots中基因表达的倍数表示基因的相对表达水平, 以GAPDH 基因为内参, 用 2-∆ ∆ CT方法计算基因相对表达量, 每个基因3个生物学重复检测, 3次技术重复检测。误差线表示标准差, * 表示经qRT-PCR检测, 该基因表达量在MSA-roots和MS-roots间差异显著(单样本t检验, P< 0.05)。Fig. 5 qRT-PCR verification of ten DEGs induced by active carbon Seven DEGs with up-regulated expression and three with down-regulated expression in MSA-roots vs. MS-roots were chosen for qRT-PCR validation. The relative expression level of each gene was expressed as fold change between MSA-roots and MS-roots in the RNA-seq data and qRT-PCR data. Relative gene expressions were normalized by comparison with that of maize GAPDH gene, and analyzed using the 2-∆ ∆ CTmethod. For each of the ten genes, the qRT-PCR assay used three biological replicates, with three technical repeats in each replicate. The bars represent the standard deviation. Asterisks indicate that the transcript level differed significantly between MSA-roots and MS-roots in qRT-PCR assay (single sample t-test, P< 0.05).
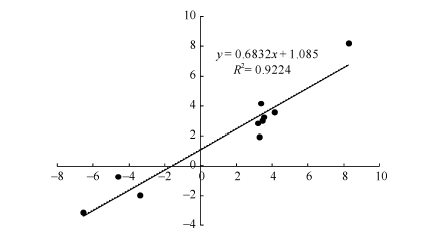
图6 10 DEGs的RNA-seq与qRT-PCR结果相关性分析 散点图表示两平台中基因相对表达量的log2转化值。Fig. 6 Correlation analysis of the expression changes of ten DEGs as revealed by RNA-seq and qRT-PCR, respectively Scatter plots indicate the log2 transformed gene expression values in RNA-seq and qRT-PCR.
王海波, 王彦霞, 赵和. 如何加快作物遗传改良的速度. 河北农业科学, 2003, (7): 50-56Wang HB, Wang YX, ZhaoH. How to accelerate the process of plant genetic modification. J Hebei Agric Sci, 2003, 7: 50-56 (in Chinese with English abstract)[本文引用:1]
[2]
Forster BP, Till BJ, Ghanim A M A, Huynh H O A, Burstmayr H, Caligari P D S. Accelerated plant breeding. CAB Rev, 2014, 9: 1-16[本文引用:1]
[3]
YaoY, ZhangP, Wang HB, Lu ZY, Liu CJ, LiuH, Yan GJ. How to advance up to seven generations of canola ( Brassica napus L. ) per annum for the production of pure line populations. Euphytica, 2016, 209: 113-119[本文引用:1]
[4]
LiuH, ZwerP, Wang HB, Liu CJ, Lu ZY, Wang YX, Yan GJ. A fast generation cycling system for oat and triticale breeding. Plant Breed, 2016, 135: 574-579[本文引用:1]
BoveJ, JullienM, Grappin P. Functional genomics in the study of seed germination. Genome Biol#/magtechI# , 2002, 3: 1002. 1-1002. 5[本文引用:1]
[7]
Galland M, HuguetR, ArcE, CueffG, JobD, RajjouL. Dynamic proteomics emphasizes the importance of selective mRNA translation and protein turnover during Arabidopsis seed germination. Mol Cell Proteomics, 2014, 13: 252-268[本文引用:1]
[8]
RajjouL, GallardoK, DebeaujonI, Vand ekerckhoveJ, JobC, JobD. The effect of α-amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosynthesized mRNAs during germination. Plant Physiol, 2004, 134: 1598-1613[本文引用:1]
[9]
AbrahamZ, Fernández RI, MartinezM, DiazI, CarboneroP, Vicente-CarbajosaJ. A developmental switch of gene expression in the barley seed mediated by HvVP1(Viviparous 1) and HvGAMYB interactions. Plant Physiol, 2016, 170: 2146-2158[本文引用:1]
[10]
MiransariM, Smith DL. Plant hormones and seed germination. Environ Exp Bot, 2014, 99: 110-121[本文引用:1]
[11]
UmezawaT, NakashimaK, MiyakawaT, KuromoriT, TanokuraM, ShinozakiK, Yamaguchi-ShinozakiK. Molecular basis of the core regulatory network in ABA responses: sensing, signaling and transport. Plant Cell Physiol, 2010, 51: 1821-1839[本文引用:1]
[12]
WeitbrechtK, MüllerK, Leubner-MetzgerG. First off the mark: early seed germination. J Exp Bot, 2011, 62: 3289-3309[本文引用:1]
[13]
SunT. Gibberellin-GID1-DELLA: a pivotal regulatory module for plant growth and development. Plant Physiol, 2010, 154: 567-570[本文引用:1]
Thomas TD. The role of activated charcoal in plant tissue culture. Biotechnol Adv, 2008, 26: 618-631[本文引用:1]
[16]
MittalP, DeviR, Gosal SS. Effect of genotypes and activated charcoal on high frequency in vitro plant regeneration in sugarcane. Ind J Biotechnol, 2016, 15: 261-265[本文引用:1]
[17]
FridborgG, PedersénM, Land ström LE, ErikssonT. The effect of activated charcoal on tissue cultures: adsorption of metabolites inhibiting morphogenesis. Physiol Plant, 1978, 43: 104-106[本文引用:1]
[18]
Manchand aP, Gosal SS. Effect of activated charcoal, carbon sources and gelling agents on direct somatic embryogenesis and regeneration in sugarcane via leaf roll segments. Sugar Tech, 2012, 14: 168-173[本文引用:1]
[19]
Nguyen TV, Thu TT, ClaeysM, AngenonG. Agrobacterium-mediated transformation of sorghum ( Sorghum bicolor(L. ) Moench) using an improved in vitro regeneration system. Plant Cell Tissue Organ Cult, 2007, 91: 155-164[本文引用:1]
[20]
EbertA, TaylorF, BlakeJ. Changes of 6-benzylaminopurine and 2, 4-dichlorophenoxyacetic acid concentrations in plant tissue culture media in the presence of activated charcoal. Plant Cell Tissue Organ Cult, 1993, 33: 157-162[本文引用:1]
[21]
EbertA, Taylor HF. Assessment of the changes of 2, 4-dichlorophenoxyacetic acid concentrations in plant tissue culture media in the presence of activated charcoal. Plant Cell Tissue Organ Cult, 1990, 20: 165-172[本文引用:1]
[22]
Halhouli KA, Darwish NA, Al-Dhoon N M. Effects of pH and inorganic salts on the adsorption of phenol from aqueous systems on activated decolorizing charcoal. Separat Sci Technol, 1995, 30: 3313-3324[本文引用:1]
[23]
Nissen SJ, Sutter EG. Stability of IAA and IBA in nutrient medium to several tissue culture procedures. Hortscience, 1990, 25: 800-802[本文引用:1]
[24]
WangZ, GersteinM, SnyderM. RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet, 2009, 10: 57-63[本文引用:1]
[25]
TrapnellC, Hendrickson DG, SauvageauM, GoffL, RinnJ, PachterL. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat Biotechnol, 2013, 31: 46-53[本文引用:1]
[26]
CostaV, AngeliniC, De FeisI, CiccodicolaA. Uncovering the complexity of transcriptomes with RNA-Seq. J Biomed Biotechnol, 2010, 2010: 853916[本文引用:1]
[27]
MurashigeT, SkoogF. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant, 1962, 15: 473-497[本文引用:1]
[28]
Aken BL, AylingS, BarrellD, ClarkeL, CurwenV, FairleyS, Fernand ez BanetJ, BillisK, García GirónC, HourlierT, HoweK, HähäriA, KokocinskiF, Martin FJ, Murphy DN, NagR, RuffierM, SchusterM, Tang YA, Vogel JH, WhiteS, ZadissaA, FlicekP, Searle SM. The Ensembl gene annotation system. Database(Oxford), 2016, 2016: baw093[本文引用:1]
[29]
TrapnellC, PachterL, Salzberg SL. TopHat: discovering splice junctions with RNA-seq. Bioinformatics, 2009, 25: 1105-1111[本文引用:1]
[30]
TrapnellC, RobertsA, GoffL, PerteaG, KimD, Kelley DR, PimentelH, SalzbergS, RinnJ, PachterL. Differential gene and transcript expression analysis of RNA—seq experiments with TopHat and Cufflinks. Nat Protocols, 2012, 7: 562-578[本文引用:1]
[31]
AndersS, Pyl PT, HuberW. HTSeq: a Python framework to work with high-throughput sequencing data. Bioinformatics, 2015, 31: 166-169[本文引用:1]
[32]
Love MI, HuberW, AndersS. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol, 2014, 15: 550[本文引用:1]
[33]
WangL, FengZ, WangX, WangX, ZhangX. DEGseq: an R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics, 2010, 26: 136-138[本文引用:2]
[34]
AshburnerM, Ball CA, Blake JA, BotsteinD, ButlerH, Cherry JM, Davis AP, DolinskiK, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-TarverL, KasarskisA, LewisS, Matese JC, Richardson JE, RingwaldM, Rubin GM, SherlockG. Gene ontology: tool for the unification of biology. Nat Genet, 2000, 25: 25-29[本文引用:1]
[35]
KanehisaM, GotoS. KEGG: Kyoto encyclopedia of genes and genomes. Nucl Acids Res, 2000, 28: 27-30[本文引用:1]
[36]
KanehisaM, GotoS, SatoY, KawashimaM, FurumichiM, TanabeM. Data, information, knowledge and principle: back to metabolism in KEGG. Nucl Acids Res, 2014, 42(database issue): D199-D205[本文引用:1]
MagaG, HübscherU. Proliferating cell nuclear antigen (PCNA): a dancer with many partners. J Cell Sci, 2003, 116: 3051-3060[本文引用:1]
[41]
GeorgescuR, LangstonL, O’Donnell M. A proposal: Evolution of PCNA’s role as a marker of newly replicated DNA. DNA Repair, 2015, 29: 4-15[本文引用:1]
[42]
StrzalkaW, ZiemienowiczA. Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation. Ann Bot, 2011, 107: 1127-1140[本文引用:1]
[43]
LabibK, Tercero JA, Diffley J F X. Uninterrupted MCM2-7 function required for DNA replication fork progression. Science, 2000, 288: 1643-1647[本文引用:1]
[44]
WeiL, ZhaoX. A new MCM modification cycle regulates DNA replication initiation. Nat Struct Mol Biol, 2016, 23: 209-216[本文引用:1]
[45]
HelariuttaY, FukakiH, Wysocka-DillerJ, NakajimaK, JungJ, SenaG, HauserM, Benfey PN. The SHORT-ROOT gene controls radial patterning of the Arabidopsis root through radial signaling. Cell, 2000, 101: 555-567[本文引用:1]
[46]
GrimpletJ, Agudelo-RomeroP, Teixeira RT, Teixeira RT, Martinez-Zapater J M, Fortes A M. Structural and functional analysis of the GRAS gene family in grapevine indicates a role of GRAS proteins in the control of development and stress responses. Front Plant Sci, 2016, 7: 353[本文引用:1]
[47]
Woodward AW, BartelB. Auxin: regulation, action, and interaction. Ann Bot, 2005, 95: 707-735[本文引用:1]
[48]
ZhaoY. Auxin biosynthesis and its role in plant development. Annu Rev Plant Biol, 2010, 61: 49-64[本文引用:1]
[49]
AbelS, TheologisA. Early genes and auxin action. Plant Physiol, 1996, 111: 9-17[本文引用:1]
[50]
Rivas-San VicenteM, PlasenciaJ. Salicylic acid beyond defence: its role in plant growth and development. J Exp Bot, 2011, 62: 3321-3338[本文引用:1]
[51]
Jand aM, Ruelland E. Magical mystery tour: salicylic acid signalling. Environ Exp Bot, 2015, 114: 117-128[本文引用:1]
[52]
JohnsonC, BodenE, AriasJ. Salicylic acid and NPR1 induce the recruitment of trans-activating TGA factors to a defense gene promoter in Arabidopsis. Plant Cell, 2003, 15: 1846-1858[本文引用:1]
 , 孙果忠
, 孙果忠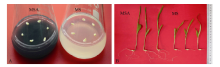






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}