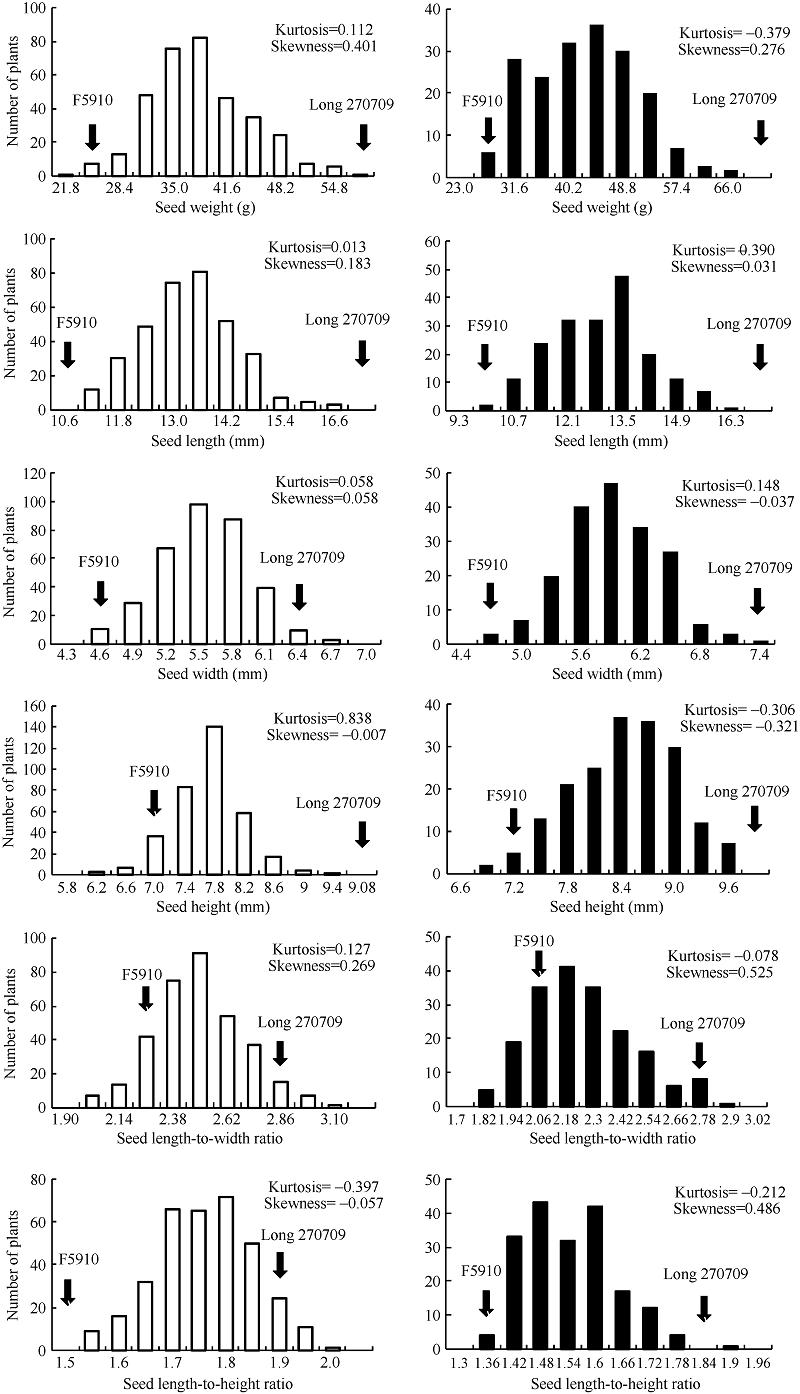
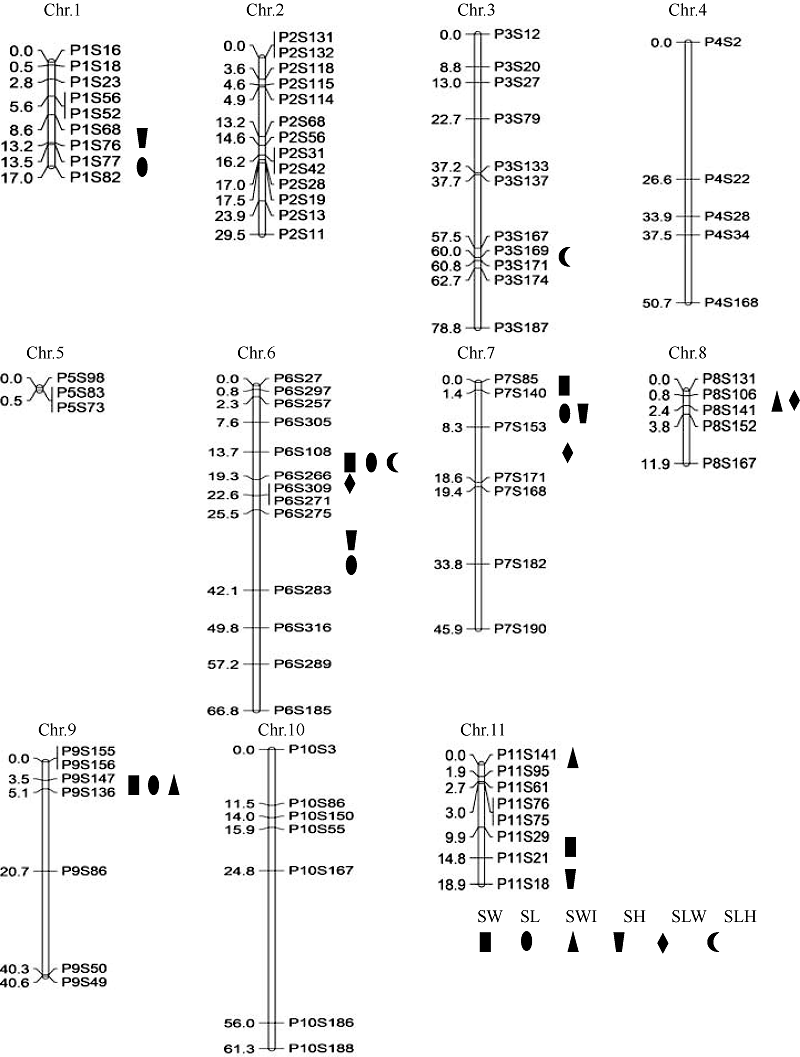
关键词:普通菜豆; 籽粒大小; 粒形; 数量性状位点(QTL) QTL Mapping for Seed Size and Shape in Common Bean GENG Qing-He, WANG Lan-Fen, WU Jing, WANG Shu-Min* Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund:This study was supported by the National Natural Science Foundation of China (31471559), the China Agriculture Research System (CARS-09), and the National Key Technology R&D Program of China (2013BAD01B05-2-4) AbstractCommon bean is one of the most important food legumes worldwide. Seed size and shape have a great effect on yield and seed quality. This research used an F2 population derived from a cross of Andean cultivar Long 270709 × Mesoamerican cultivar F5910 planted in Harbin, Heilongjiang and Beijing respectively to analyze the correlation among seed weight, seed length, seed width, seed height, seed length-to-width ratio and seed length-to-height ratio, and to detect QTL related to these traits. The correlation analysis showed that seed weight had a significant correlation with other traits related to seed size and shape. Software IciMapping 4.1 based on inclusive composite interval mapping was used to identify the additive QTL for seed size and shape. A total of 38 QTL were detected on 10 chromosomes except the first chromosome in Harbin environment with the PVE between 2.39% and 17.37%. A total of 21 QTL were detected on seven chromosomes (Chr.1, Chr.3, Chr.6, Chr.7, Chr.8, Chr.9, Chr.11) in Beijing environment with the PVE between 5.92% and 22.53%. Among them, four pairs of QTL were detected on the same marker interval, including seed weight QTL SW7 and SW7’, seed weight QTL SW6.1 and SW6’, seed length QTL SL6.1 and SL6.1’, and seed height QTL SH11 and SH11’. The PVE of SW7, SW6.1, SL6.1, SW6’, and SL6.1’ was larger than 10%.
Keyword:Common bean; Seed size; Seed shape; Quantitative trait locus (QTL) Show Figures Show Figures
表2 籽粒性状间的相关性分析 Table 2 Correlation coefficients for seed traits in common bean
性状 Trait
百粒重 SW
粒长 SL
粒宽 SWI
粒厚 SH
长宽比 SLW
长厚比 SLH
百粒重 SW
0.867* *
0.725* *
0.761* *
0.166* *
0.449* *
粒长 SL
0.886* *
0.445* *
0.760* *
0.556* *
0.658* *
粒宽 SWI
0.723* *
0.431* *
0.426* *
-0.493* *
0.185* *
粒厚 SH
0.854* *
0.772* *
0.493* *
0.344* *
0.013
长宽比 SLW
0.262*
0.630* *
-0.424* *
0.351* *
0.464* *
长厚比 SLH
0.440* *
0.715* *
0.123
0.110
0.608* *
The data at upper right corner are from the F2population planted in Harbin, Heilongjiang, and these at lower left corner are from the F2population planted in Beijing. * * Significant at the 0.01 probability level; * significant at the 0.05 probability level. SW: seed weight; SL: seed length; SWI: seed width; SH: seed height; SLW: seed length-to-width ratio; SLH: seed length-to-height ratio. 表右上角为在黑龙江哈尔滨种植的F2各性状间的相关系数, 左下角为在北京昌平种植的F2各性状间的相关系数; * * 表示0.01的显著水平, * 表示0.05的显著水平。
表2 籽粒性状间的相关性分析 Table 2 Correlation coefficients for seed traits in common bean
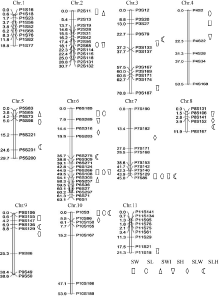
2.2 遗传图谱构建利用哈尔滨种植的346株F2和北京昌平种植的188株F2分别构建了连锁图谱(图2和图3)。346株F2构建的遗传图谱包括100个SSR标记, 分布于11条染色体上, 每条染色体上的标记个数5~17个, 连锁图谱总长439.88 cM, 标记间平均遗传距离4.40 cM, 有2对标记间的遗传距离大于20.00 cM, 分别位于第10染色体P10S167~P10S186 (31.91 cM)和第4染色体P4S2~P4S22 (22.28 cM), 其余标记遗传距离均小于20 cM。188株F2构建的遗传图谱包括88个SSR标记, 与前者相比, 第5、第6、第7、第9和第11染色体分别缺少了4、4、2、1、1个标记, 为F2基因型缺失较严重的标记, 其余标记均与前者相同, 每条染色体上的标记个数3~13个, 连锁图谱总长421.94 cM, 标记间平均遗传距离4.80 cM, 标记间遗传距离大于20 cM的2对标记同样位于第10染色体P10S167~P10S186 (31.18 cM)和第4染色体P4S2~ P4S22 (26.57 cM), 其余标记的遗传距离均小于20 cM。 图2 Fig. 2
张赤红, 曹永生, 宗绪晓, 王志刚, 王述民. 普通菜豆种质资源形态多样性鉴定与分类研究. 中国农业科学, 2005, 38: 27~32Zhang CH, Cao YS, Zong XX, Wang ZG, Wang SM. Morphological diversity and classification of common bean (Phaseolus vulgaris L. ) germplasm resource in China. Sci Agric Sin, 2005, 38: 27-32 (in Chinese with English abstract)[本文引用:1]
[2]
Pérez-VegaE, PañedaA, Rodríguez-SuárezC, CampaA, GiraldezR, Ferreira JJ. Mapping of QTLs for morpho-agronomic and seed quality traits in a RIL population of common bean (Phaseolus vulgaris L. ). Theor Appl Genet, 2010, 120: 1367-1380[本文引用:4]
[3]
SchmutzJ, McClean P E, Mamidi S, Wu G A, Cannon S B, Grimwood J, Jenkins J, Shu S Q, Song Q J, Chavarro C, Torres-Torres M, Geffroy V, oghaddam S M, Gao D Y, Abernathy B, Barry K, Blair M, Brick M A, Chovatia M, Gepts P, Goodstein D M, Gonzales M, Hellsten U, Hyte D L, Jia G F, Kelly J D, Kudrna D, Lee R, Richard M M S, Miklas P N, Osorno J M, Rodrigues J, Thareau V, Urrea C A, Wang M, Yu Y, Zhang M, Wing R A, Cregan P B, Rokhsar D S, Jackson S A. A reference genome for common bean and genome-wide analysis of dual domestications. Nat Genet, 2014, 46: 707-713[本文引用:3]
[4]
Fan CC, Xing YZ, Mao HL, Lu TT, HanB, Xu CG, Li XH, Zhang QF. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet, 2006, 112: 1164-1171[本文引用:3]
[5]
Song XJ, HuangW, ShiM, Zhu MZ, Lin HX. A QTL for rice grain width and weight encodes a previously unknown RING- type E3 ubiquitin ligase. Nat Genet, 2007, 39: 623-630[本文引用:3]
[6]
ShomuraA, IzawaT, EbanaK, EbitaniT, KanegaeH, KonishiS, YanoM. Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet, 2008, 40: 1023-1028[本文引用:2]
[7]
Weng JF, Gu SH, Wan XY, GaoH, GuoT, SuN, Lei CL, ZhangX, Cheng ZJ, Guo XP, Wang JL, JiangL, Zhai HQ, Wan JM. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res, 2008, 18: 1199-1209[本文引用:1]
[8]
Wang ET, Wang JJ, Zhu XD, HaoW, Wang LY, LiQ, Zhang LX, HeW, Lu BR, Lin HX, MaH, Zhang GQ, He ZH. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat Genet, 2008, 40: 1370-1374[本文引用:1]
[9]
Li YB, Fan CC, Xing YZ, Jiang YH, Luo LJ, SunL, ShaoD, Xu CJ. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet, 2011, 43: 1266-1269[本文引用:1]
[10]
Wang SK, WuK, Yuan QB, Liu XY, Liu ZB, Lin XY, Zeng RZ, Zhu H T. Control of grain size, shape and quality by OsSPL16 in rice. 2012, Nat Genet, 44: 950-954[本文引用:1]
[11]
SaxK. The association of size differences with seed-coat pattern and pigmentation in PHASEOLUS VULGARIS. Genetics, 1923, 8: 552-560[本文引用:2]
[12]
Vallejos CE, Chase CD. Linkage between isozyme markers and a locus affecting seed size in Phaseolus vulgaris L. Theor Appl Genet, 1991, 81: 413-419[本文引用:1]
[13]
Blair MW, IriarteG, BeebeS. QTL analysis of yield traits in an advanced backcross population derived from a cultivated Andean × wild common bean (Phaseolus vulgaris L. ) cross. Theor Appl Genet, 2006, 112: 1149-1163[本文引用:4]
[14]
Yuste-Lisbona FJ, González AN, CapelC, Garcia-AlcazarM, CapelJ, Ron A MD, LozanoR, SantallaM. Genetic analysis of single-locus and epistatic QTLs for seed traits in an adapted× nuña RIL population of common bean (Phaseolus vulgaris L. ). Theor Appl Genet, 2014, 127: 897-912[本文引用:4]
[15]
VlasovaA, Capella-GutiérrezS, Rendón-AnayaM, Hernández- Oñate M E. Minoche A, Erb I, Câmara F, Prieto-Barja P, Corvelo A, Sanseverino W, Westergaard G, Dohm J C, Pappas Jr G J, Saburido-Alvarez S, Kedra D, Gonzalez I, Cozzuto L, Gómez-Garrido J, Aguilar-Morón M A, Andreu N, Aguilar O, Garcia-Mas J, Zehnsdorf M, Vázquez M P, Delgado-Salinas A, Delaye L, Lowy E, Mentaberry A, Vianello-Brondani R P, García J, Alioto T, Sánchez F, Himmelbauer H, Santalla M, Notredame C, Gabaldón T, Herrera-Estrella A, Guigó R. Genome and transcriptome analysis of the Mesoamerican common bean and the role of gene duplications in establishing tissue and temporal specialization of genes. Genome Biol, 2016, 17: 1-18[本文引用:1]
[16]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[17]
Li HH, Ye GY, Wang JK. A modified algorithm for the improvement of composite interval mapping. Genetics, 2007, 175: 361-374[本文引用:2]
[18]
MengL, Li HH, Zhang LY, Wang JK. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental population. Crop J, 2015, 3: 269-283[本文引用:2]
[19]
Wan XY, Wan JM, JiangL, Wang JK, Zhai HQ, Weng JF, Wang HL, Lei CL, Wang JK, ZhangX. QTL analysis for rice grain length and fine mapping of an identified QTL with stable and major effects. Theor Appl Genet, 2006, 112: 1258-1270[本文引用:1]
[20]
高用明, 朱军, 宋佑胜, 何慈信, 石春海, 邢永忠. 水稻永久F2群体抽穗期QTL的上位性及其与环境互作效应的分析. 作物学报, 2004, 30: 849-854Gao YM, ZhuJ, Song YS, He CX, Shi CH, Xing YZ. Use of permanent F2 population to analyze epistasis and their interaction effects with environments for QTLs controlling heading date in rice. Acta Agron Sin, 2004, 30: 849-854 (in Chinese with English abstract)[本文引用:1]
[21]
Hoyos-VillegasV, SongQ, Wright EM, Beebe SE, Kelly DJ. Joint linkage QTL mapping for yield and agronomic traits in a composite map of three common bean RIL populations. Crop Sci, 2016, 56: 1-18[本文引用:1]
[22]
Tanksley SD. Mapping ploygenes. Annu Rev Genet, 1993, 27: 205-233[本文引用:1]
[23]
陈秀华, 于丽娟, 罗黎明, 陈洪梅, 刘丽. 玉米分子标记辅助育种及标记开发研究进展. 中国农业科技导报, 2016, 18(1): 26-31Chen XH, Yu LJ, Luo LM, Chen HM, LiuL. Research progress on maize molecular marker-assisted breeding and marker development. J Agric Sci Technol, 2016, 18(1): 26-31 (in Chinese with English abstract)[本文引用:1]
[24]
赵洪波, 李明丽, 鲁绍雄, 连林生, 李国治. 群体规模和性状遗传力对F2设计下QTL定位效果的影响. 云南农业大学学报, 2007, 22(2): 159-163Zhao HB, Li ML, Lu SX, Lian LS, Li GZ. Study on the effects of population size and trait heritability on the accuracy of QTL mapping under F2 design. J Yunnan Agric Univ, 2007, 22(2): 159-163 (in Chinese with English abstract)[本文引用:1]
[25]
GroosC, RobertN, BervasE, CharmetG. Genetic analysis of grain protein-content, grain yield and thousand -kernel weight in bread wheat. Theor Appl Genet, 2003, 106: 1032-1040[本文引用:1]
[26]
张坤普, 徐宪斌, 田纪春. 小麦籽粒产量及穗部相关性状的QTL定位. 作物学报, 2009, 35: 270-278Zhang KP, Xu XB, Tian JC. QTL mapping for grain yield and spike related traits in common wheat. Acta Agron Sin, 2009, 35: 270-278 (in Chinese with English abstract)[本文引用:1]
[27]
李文福, 刘宾, 彭涛, 袁倩倩, 韩淑晓, 田纪春. 利用DH和IF2两个群体进行小麦粒重、粒型和硬度的QTL分析. 中国农业科学, 2012, 45: 3453-3462Li WF, LiuB, PengT, Yuan QQ, Han SX, Tian JC. Detection of QTL for kernel weight, grain size, and grain hardness in wheat using DH and immortalized F2 population. Sci Agric Sin, 2012, 45: 3453-3462 (in Chinese with English abstract)[本文引用:1]
[28]
覃鸿妮, 晏萌, 王召辉, 郭莹, 王辉, 孙海燕, 刘志斋, 蔡一林. 玉米籽粒中花色苷和黑色素含量的QTL分析. 作物学报, 2012, 38: 275-284Qin HN, YanM, Wang ZH, GuoY, WangH, Sun HY, Liu ZZ, Cai YL. QTL mapping for anthocyanin and melanin contents in maize kernel. Acta Agron Sin, 2012, 38: 275-284 (in Chinese with English abstract)[本文引用:1]
[29]
郑德波, 杨小红, 李建生, 严建兵, 张士龙, 贺正华, 黄益勤. 基于SNP标记的玉米株高及穗位高QTL定位. 作物学报, 2013: 549-556Zheng DB, Yang XH, Li JS, Yan JB, Zhang SL, He ZH, Huang YQ. QTL identification for plant height and ear height based on SNP mapping in maize (Zea mays L. ). Acta Agron Sin, 2013, 39: 549-556 (in Chinese with English abstract)[本文引用:1]
[30]
TuberosaR, SalviS, Sanguineti MC, Land iP, MaccaferriM, ContiS. Mapping QTLs regulating morpho-physiological traits and yield: case studies, shortcomings and perspectives in drought-stressed maize. Ann Bot, 2002, 89: 941-963[本文引用:1]
 , 王兰芬, 武晶, 王述民
, 王兰芬, 武晶, 王述民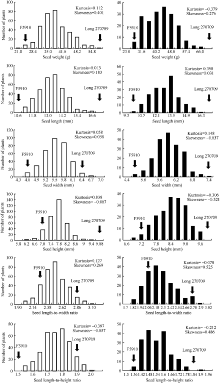



{kind=link}
{kind=link}
{kind=link}