关键词:同位素示踪; 氮效率; 水稻; 氮肥; 碳氮代谢 Effects of Postponing Nitrogen Topdressing on Post-anthesis Carbon and Nitrogen Metabolism in Rice Cultivars with Different Nitrogen Use Efficiencies SUN Yong-Jian1, SUN Yuan-Yuan2, YAN Feng-Jun1, YANG Zhi-Yuan1, XU Hui1, LI Yue1, WANG Hai-Yue1, MA Jun1,* 1 Rice Research Institute, Sichuan Agricultural University / Key Laboratory of Crop Physiology, Ecology, and Cultivation in Southwest, Ministry of Agriculture, Wenjiang 611130, China
2 Institute of Plateau Meteorology, China Meteorological Administration, Chengdu 610072, China
Fund:This study was supported by National Key Research and Development Program of China (2016YFD0300506), Scientific Research Fund of Sichuan Provincial Education Department (16ZA0044), Open Project of Key Laboratory of Crop Physiology, Ecology, and Cultivation in Southwest, Chinese Ministry of Agriculture (201303), the National Science and Technology Project of Food Production of China (2013BAD07B13), and the Rice Breeding Project in Sichuan Province of China (2016NYZ0051). AbstractThe optimal nitrogen (N) managements and the selection of genotypes with high N use efficiency (NUE) play a vital role in rice production aiminged at high yield and high NUE. Two rice cultivars, one with high-NUE (Dexiang 4103) and the other with low-NUE (Yixiang 3724) were used in pot and field experiments in 2013 and 2014. The total N fertilizer applied was 180 kg ha-1 of urea and three treatments were included: 1) 50% basal dressing, 30% topdressing at 7 d after transplanting (DAT), and 20% topdressing at 4th leaves emerged from the top (N1), 2) 30% basal dressing, 30% topdressing at 7 DAT, 40% topdressing was split into two equal applications at 4th and 2nd leaves emerged from the top, 3) 20% basal dressing, 20% topdressing at 7 DAT, 60% topdressing was split into two equal applications at 4th and 2nd leaves emerged from the top, respectively. Double isotope tracing technique of13C and15N and physiological-biochemical analysis were used to study the accumulation, translocation, distribution of N and photosynthate, and the correlation between morphology and physiological-biochemical characteristics and their relationships with grain yield. There were significant effects of cultivars and N application modes on grain yield, as well as the absorption and translocation of N and photosynthate from full-heading to maturity stage. Compared with N1and N3, N2 treatment with high-NUE was the best model in this paper referred as the variety and N application coupling model, which could improve N accumulation in rice plant after anthesis, increase photosynthetic rate, activities of ribulose 1,5-bisphosphate carboxylase, and glutamine synthetase in flag leaves, promote accumulation and translocation of photosynthate and N, and then improve the yield and NUE. Double isotope labeling results showed that the accumulation amount of photosynthetic products and N in rice plant with high-NUE was 7.78-12.75 mg13C plant-1 and 15.14-18.78 mg15N plant-1 higher, the translocation amount of photosynthate and N in leaves with high-NUE was 1.70-2.93 mg13C plant-1, 2.21-4.55 mg15N plant-1 higher, the translocation amount of photosynthate and N in leaf sheaths with high-NUE was 1.70-2.93 mg13C plant-1, 0.05-1.14 mg15N plant-1 higher than those with low-NUE, respectively. From full-heading to maturity stage,13C photosynthate in the spike with high-NUE and low-NUE respectively increased by 31.04-44.68 mg13C plant-1(accounting for 42.04%-46.38% of total amount of13C) and 24.94-34.26 mg13C plant-1 (accounting for 36.45%-41.36% of total amount of13C), while,15N accumulation in the spike with high-NUE and low-NUE respectively increased by 35.56-46.58 mg15N plant-1(accounting for 61.82%-82.93% of total amount of15N) and 27.37-31.57 mg15N plant-1 (accounting for 58.04%-68.31% of total amount of15N). Compared with low-NUE, the high NUE rice cultivar is more beneficial to the accumulation and translocation of photosynthate, with higher N absorption capacity, stronger N translocation ability, and stronger C and N metabolism capacity in leaves, satisfying the N demand of plants during their grain forming stage, which is the important reason for high-NUE rice cultivar further to increase yield and NUE. From, the information of C/N ratio in different organs after-anthesis, combined with the results of high yield and high NUE in different varieties under N2 treatment, from full-heading to maturity stage, the C/N increasing range in leaf and panicle is the same as the reducing range in leaf sheaths and roots. Therefore, C/N ratio in different organs from full-heading to maturity stage might be a candidate indicator for high yield and high NUE in rice production.
Keyword:Isotope tracer; N use efficiency; Rice; N fertilizer; Carbon and N metabolism Show Figures Show Figures
表1 试验田耕层土壤(0~20 cm)理化性状 Table 1 Physicochemical characteristics of soil (0-20 cm) in the experiments
年份 Year
全氮 Total N (g kg-1)
有机质 Organic matter (g kg-1)
速效养分 Available nutrient (mg kg-1)
pH
容重 Bulk density (g cm-3)
N
P
K
2013
1.87
21.1
98.0
35.3
95.5
6.41
1.24
2014
1.91
22.3
103.8
33.6
102.4
6.44
1.28
表1 试验田耕层土壤(0~20 cm)理化性状 Table 1 Physicochemical characteristics of soil (0-20 cm) in the experiments
表2 Table 2 表2(Table 2)
表2 供试品种产量表现及氮利用率 Table 2 Yield performance and N use efficiency of two experimental rice cultivars
氮效率类型 N use efficiency (NUE) type
品种 Cultivar
生育期 Stage growth (d)
对应最佳氮肥水平 Optimum N application (kg N hm-2)
稻谷产量 Grain yield (kg hm-2)
氮肥回收利用率 N recovery efficiency (%)
氮肥生理利用率 N physiological efficiency (kg kg-1)
氮高效 High-NUE
德香4103 Dexiang 4103
150.2
180
10397.6 a
50.61 a
29.67 a
氮低效 Low-NUE
宜香3724 Yixiang 3724
150.0
180
9340.1 b
41.65 b
22.10 b
Values within a column followed by different letters are significantly different at P< 0.05. Data are averaged across 2011 and 2012 years. 同栏内标以不同字母的值在5%水平上差异显著。表中数据为2011年和2012年试验的平均值。
表2 供试品种产量表现及氮利用率 Table 2 Yield performance and N use efficiency of two experimental rice cultivars
表3 氮肥后移对不同氮效率水稻产量及构成因素的影响(盆栽试验2013-2014年) Table 3 Effects of postponing N topdressing on yield and its components in rice cultivars with different NUE in the pot experiments (2013-2014)
品种 Cultivar
处理 Treatment
有效穗 Effective panicles (plant-1)
穗粒数 Spikelets (No. panicle-1)
总颖花数 Total spikelets (plant-1)
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
稻谷产量 Grain yield (g pot-1)
2013
2014
德香4103
N0
10.08 e
155.43 bc
1565.96 e
88.79 a
30.71 e
87.32 e
85.00 e
Dexiang 4103
N1
11.64 bc
165.81 a
1930.53 b
86.90 bc
30.92 e
103.14 bc
105.39 bc
N2
12.14 a
169.52 a
2057.38 a
88.57 a
31.56 d
110.97 a
113.68 a
N3
12.08 ab
165.47 a
1998.79 ab
87.74 b
31.32 de
106.86 ab
109.16 ab
平均Average
11.48
164.06
1888.16
88.00
31.13
102.07
103.31
宜香3724
N0
9.87 e
148.21 d
1462.24 f
86.27 c
33.13 bc
82.84 e
83.47 e
Yixiang 3724
N1
11.49 cd
151.83 cd
1744.60 d
85.14 d
33.75 b
98.59 c
100.52 c
N2
11.57 c
158.80 b
1836.76 c
86.36 c
34.45 a
105.92 b
106.13 b
N3
11.03 d
154.90 bc
1707.77 d
83.33 e
33.06 c
93.40 d
95.18 d
平均Average
10.99
153.44
1687.84
85.28
33.60
95.19
96.33
F值 F-value
C
7.07* *
26.15* *
51.27* *
6.96*
93.59* *
43.11* *
53.58* *
N
27.49* *
5.64*
60.06* *
16.93* *
4.52*
159.32* *
188.00* *
C× N
1.86
3.54*
4.04*
0.79
1.03
5.02*
4.12*
Values within a column followed by different letters are significantly different at P< 0.05. * Significant at P< 0.05; * * Significant at P< 0.01. Data of yield components are the average across two years. C: cultivar; N: N fertilizer treatment; C× N: cultivar and N fertilizer treatment interaction. 同栏内标以不同字母的值在5%水平上差异显著。* , * * 分别表示在0.05和0.01水平上差异显著。表中产量构成因素数据为两年试验的平均值。C: 品种; N: 施肥处理; C× N: 品种与施肥处理互作。
表3 氮肥后移对不同氮效率水稻产量及构成因素的影响(盆栽试验2013-2014年) Table 3 Effects of postponing N topdressing on yield and its components in rice cultivars with different NUE in the pot experiments (2013-2014)
表4 Table 4 表4(Table 4)
表4 氮肥后移对不同氮效率水稻产量及构成因素的影响(大田试验2013-2014年) Table 4 Effects of postponing N topdressing on yield and its components in rice cultivars with different NUE in the field experiments (2013-2014)
品种 Cultivar
处理 Treatment
有效穗 Effective panicles (× 104 hm-2)
每穗粒数 Spikelets (No. panicle-1)
总颖花数 Total spikelets (× 106 hm-2)
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
稻谷产量 Grain yield (kg hm-2)
2013
2014
德香4103
N0
189.55 e
150.52 cd
285.31 e
87.17 a
31.45 cd
7745.05 d
7702.87 e
Dexiang 4103
N1
212.06 b
163.16 b
345.99 b
85.88 b
32.03 c
9329.52 b
9423.14 bc
N2
222.48 a
170.22 a
378.70 a
86.80 a
32.10 c
10371.16 a
10460.61 a
N3
212.09 b
173.41 a
367.78 a
82.31 e
31.21 d
9402.48 b
9865.56 ab
平均Average
209.04
164.33
344.45
85.54
31.70
9212.05
9363.05
宜香3724
N0
180.37 f
142.73 e
257.44 f
84.88 c
34.37 b
7454.03 d
7514.79 e
Yixiang 3724
N1
204.51 cd
147.33 de
301.30 de
83.15 de
35.16 ab
8655.20 c
8968.69 cd
N2
207.04 bc
155.61 bc
320.93 c
83.27 d
35.88 a
9444.98 b
9556.02 bc
N3
198.97 c
155.01 cd
312.44 cd
80.34 f
34.37 b
8568.62 c
8535.77 d
平均Average
197.72
150.17
298.03
82.91
34.94
8530.71
8643.82
F值 F-value
C
7.41*
19.36* *
49.36* *
3.88*
63.27* *
38.92* *
40.96* *
N
9.58* *
6.36* *
41.00* *
6.49*
5.82*
76.02* *
82.03* *
C× N
0.19
3.98*
10.21* *
0.11
0.64
8.20* *
4.83*
Values within a column followed by different letters are significantly different at P < 0.05. * Significant at P< 0.05; * * Significant at P< 0.01. Data of yield components are the average across two years. Abbreviations are the same as those given in Table 3. 同栏内标以不同字母的值在5%水平上差异显著。* , * * 分别表示在0.05和0.01水平上差异显著。表中产量构成因素数据为两年试验的平均值。缩写同表3。
表4 氮肥后移对不同氮效率水稻产量及构成因素的影响(大田试验2013-2014年) Table 4 Effects of postponing N topdressing on yield and its components in rice cultivars with different NUE in the field experiments (2013-2014)
表5 Table 5 表5(Table 5)
表5 氮肥后移对不同氮效率水稻氮素累积利用效率的影响(盆栽2014年) Table 5 Effects of postponing N topdressing on N accumulation and N use efficiency (NUE) in rice cultivars with different NUE in the pot experiment (2014)
品种 Cultivar
处理 Treatment
齐穗期氮素积累量 FTNA (g pot-1)
成熟期氮素总积累量 MTNA (g pot-1)
氮素干物质 生产效率 NMPE (kg kg-1)
氮素稻谷 生产效率 NGPE (kg kg-1)
氮肥回收 利用率 NRE (%)
氮肥农艺 利用率 NAE (kg kg-1)
氮肥生理 利用率 NPE (kg kg-1)
德香4103
N0
1.04 e
1.24 e
126.96 a
68.49 a
—
—
—
Dexiang 4103
N1
1.88 a
2.13 b
95.89 d
49.48 bc
44.46 bc
10.20 cd
22.93 c
N2
1.87 ab
2.23 a
96.09 d
51.10 b
49.18 a
14.34 a
29.16 a
N3
1.77 c
2.18 ab
99.37 cd
50.12 bc
46.85 ab
12.08 b
25.78 b
平均Average
1.64
1.94
104.58
54.80
46.83
12.21
25.96
宜香3724
N0
1.04 e
1.22 e
128.33 a
68.14 a
—
—
—
Yixiang 3724
N1
1.77 c
2.02 cd
99.59 cd
49.70 bc
39.88 d
8.53 d
21.38 c
N2
1.78 bc
2.09 bc
100.38 bc
50.90 b
43.01 c
11.33 bc
26.35 b
N3
1.67 d
1.99 d
103.62 b
47.79 c
38.34 d
5.86 e
15.27 d
平均Average
1.56
1.83
107.98
54.13
40.41
8.57
21.00
F值 F-value
C
15.28* *
23.17* *
4.84*
2.45
48.83* *
104.28* *
78.69* *
N
97.69* *
80.11* *
43.10* *
68.43* *
11.83* *
50.15* *
62.43* *
C× N
1.79
4.46*
0.20
0.52
4.98*
17.89* *
12.65* *
Values within a column followed by different letters are significantly different at P < 0.05. * * Significant at P < 0.05; * * Significant at P < 0.01. FTNA: total N accumulation at full-heading; MTNA: total N accumulation at maturity; NMPE: N dry matter production efficiency; NGPE: N grain production efficiency; NRE: N recovery efficiency; NAE: N agronomy efficiency; NPE: N physiological efficiency. Abbreviations are the same as those given in Table 3. 同栏内标以不同字母的数据在5%水平上差异显著。* , * * 分别表示在0.05和0.01水平上差异显著。缩写同表3。
表5 氮肥后移对不同氮效率水稻氮素累积利用效率的影响(盆栽2014年) Table 5 Effects of postponing N topdressing on N accumulation and N use efficiency (NUE) in rice cultivars with different NUE in the pot experiment (2014)
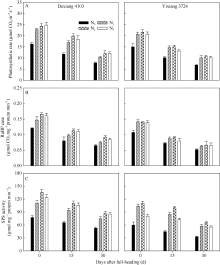
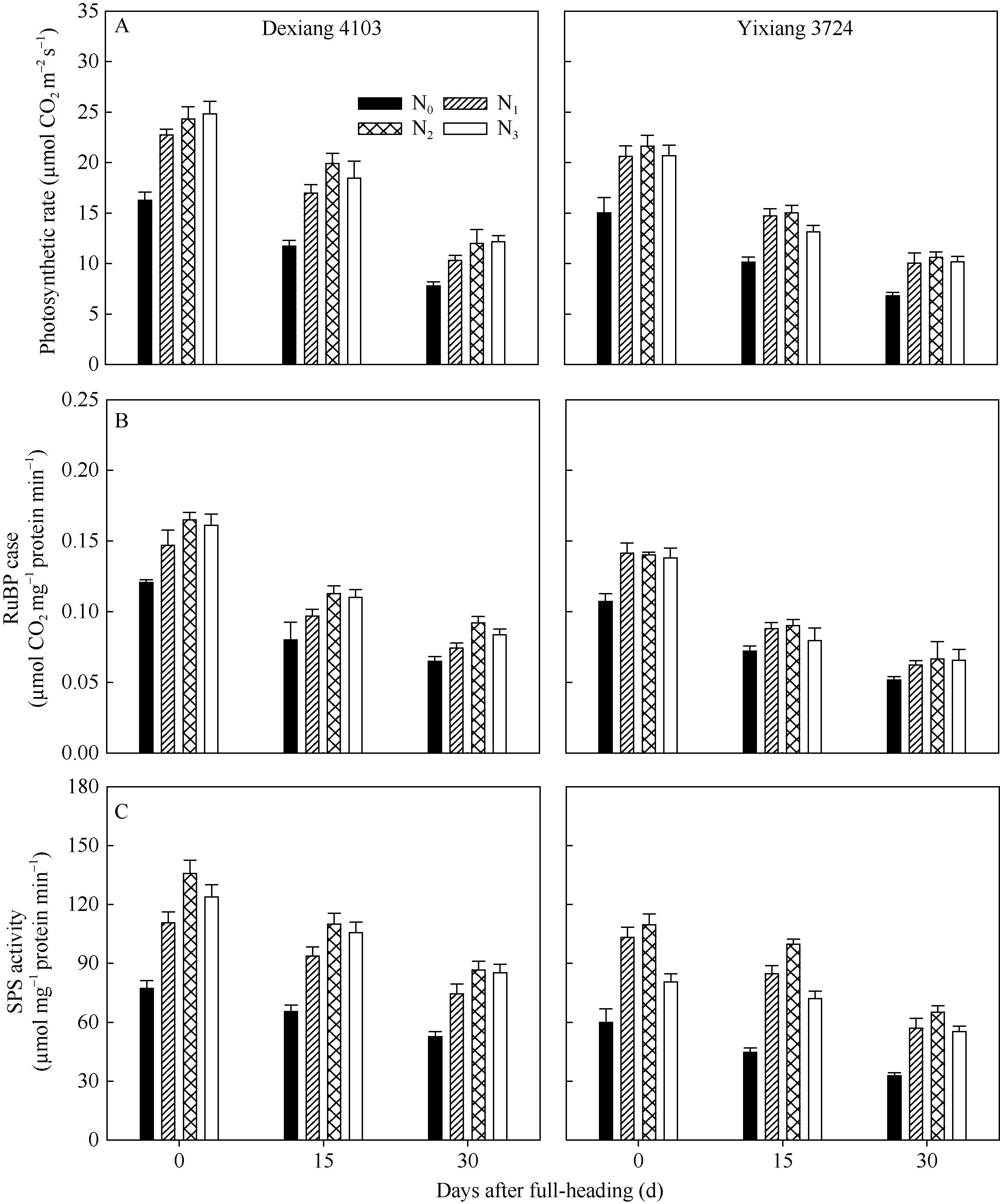
图1 氮肥后移对不同氮效率水稻剑叶光合速率(A)、RuBP羧化酶(B)和SPS(C)活性的影响(盆栽, 2014年)Fig. 1 Effects of postponing N topdressing on photosynthetic rate (A), RuBPCase (B) and SPS (C) activity of leaves in rice cultivars with different NUE in the pot experiment (2014)
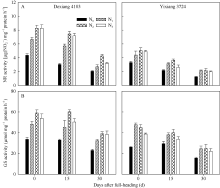
图2 氮肥后移对不同氮效率水稻剑叶NR (A)和GS (B)活性的影响(盆栽, 2014年)Fig. 2 Effects of postponing N topdressing on NR (A) and GS (B) activity of leaves in rice cultivars with different NUE in the pot experiments (2014)
表6 Table 6 表6(Table 6)
表6 氮肥后移对不同氮效率水稻花后各营养器官13C同化物累积与转运的影响(mg 13C 株-1) (盆栽, 2014年) Table 6 Effects of postponing N topdressing on post-anthesis accumulation and translocation of 13C in rice cultivars with different NUE (mg 13C plant-1) in the pot experiment (2014)
品种 Cultivar
处理 Treatment
齐穗期Full-heading stage
成熟期Maturity stage
叶 Leaf
茎鞘 Stem
穗 Spike
根 Root
叶 Leaf
茎鞘 Stem
穗 Spike
根 Root
德香4103
N0
13.92 d
38.33 de
9.66 de
5.01 e
6.04 e
15.02 e
40.69 d
5.16 cd
Dexiang 4103
N1
25.11 b
49.16 b
10.09 cd
6.29 d
12.61 cd
22.96 b
48.62 bc
6.46 b
N2
26.69 a
54.05 a
12.24 a
7.10 c
13.91 a
23.02 b
56.92 a
6.21 b
N3
26.64 a
48.26 b
11.41 b
8.99 a
13.94 a
23.62 a
51.59 b
6.20 b
平均Average
23.09
47.45
10.85
6.85
11.62
21.16
49.45
6.01
宜香3724
N0
13.11 d
33.92 f
7.81 f
5.46 e
6.93 e
15.10 e
32.75 e
5.52 c
Yixiang 3724
N1
22.70 c
40.32 d
9.46 de
7.07 cd
11.90 d
21.27 d
39.39 d
6.99 a
N2
24.29 b
43.44 c
10.49 c
7.52 bc
12.89 bc
23.16 ab
44.76 c
4.94 d
N3
23.24 c
36.62 ef
8.95 e
8.02 b
13.44 ab
22.25 c
36.96 d
4.19 e
平均Average
20.84
38.58
9.18
7.02
11.29
20.45
38.46
5.41
F值 F-value
C
16.67* *
177.69* *
66.36* *
1.39
1.91
2.72
226.58* *
27.71* *
N
105.72* *
98.26* *
28.04* *
90.17* *
194.80* *
80.18* *
62.71* *
33.28* *
C× N
0.94
8.99* *
3.45*
3.14
3.10
1.24
4.13*
29.34* *
Values within a column followed by different letters are significantly different at P< 0.05. * Significant at P< 0.05; * * Significant at P< 0.01. Abbreviations are the same as those given in Table 3. 同栏内标以不同字母的值在5%水平上差异显著。* , * * 分别表示在0.05和0.01水平上差异显著。缩写同表3。
表6 氮肥后移对不同氮效率水稻花后各营养器官13C同化物累积与转运的影响(mg 13C 株-1) (盆栽, 2014年) Table 6 Effects of postponing N topdressing on post-anthesis accumulation and translocation of 13C in rice cultivars with different NUE (mg 13C plant-1) in the pot experiment (2014)
表7 Table 7 表7(Table 7)
表7 氮肥后移对不同氮效率水稻花后各营养器官15N累积与分配的影响(mg 15N 株-1) (盆栽, 2014年) Table 7 Effects of postponing N topdressing on post-anthesis accumulation and distribution of 15N in rice cultivars with different NUE (mg 15N plant-1) in the pot experiment (2014)
品种 Cultivar
处理 Treatment
齐穗期Full-heading stage
成熟期Maturity stage
叶 Leaf
茎鞘 Stem
穗 Spike
根 Root
15N总量 Total 15N
叶 Leaf
茎鞘 Stem
穗 Spike
根 Root
15N总量 Total 15N
德香4103
N0
—
—
—
—
—
—
—
—
—
—
Dexiang 4103
N1
28.13 a
18.68 a
10.92 a
2.35 d
60.08 a
14.49 a
15.98 a
48.06 b
2.21 cd
80.73 a
N2
26.78 a
16.08 ab
10.32 ab
2.98 ab
56.16 ab
11.11 bc
13.27 b
56.90 a
2.88 b
84.17 a
N3
21.96 b
13.81 bc
8.70 c
3.20 a
47.67 c
10.30 cd
12.01 c
45.26 b
3.85 a
71.42 b
平均Average
25.63
16.19
9.98
2.84
54.63
11.97
13.75
50.07
2.98
78.77
宜香3724
N0
—
—
—
—
—
—
—
—
—
—
Yixiang 3724
N1
23.05 b
15.10 bc
8.42 c
2.66 c
49.23 bc
11.61 b
12.53 bc
36.99 d
1.96 d
63.09 c
N2
22.70 b
12.41 cd
9.32 bc
2.74 bc
47.16 c
11.58 b
10.74 d
40.89 c
2.18 cd
65.39 c
N3
18.84 c
10.95 d
7.29 d
2.98 ab
40.06 d
10.05 d
9.21 e
34.66 d
2.36 c
56.28 d
平均Average
21.53
12.82
8.34
2.79
45.48
11.08
10.83
37.51
2.16
61.59
F 值 F-value
C
53.27* *
94.64* *
56.18* *
0.56
59.24* *
10.53* *
147.73* *
115.30* *
136.64* *
105.91* *
N
32.87* *
57.14* *
28.61* *
25.74* *
29.25* *
37.21* *
20.59* *
23.04* *
90.25* *
15.35* *
C× N
1.02
0.55
4.19*
3.21
0.62
13.85* *
22.35* *
5.31*
34.00* *
4.14*
Values within a column followed by different letters are significantly different at P< 0.05. * Significant at P< 0.05. * * Significant at P< 0.01. Abbreviations are the same as those given in Table 3. 同栏内标以不同字母的值在5%水平上差异显著。* , * * 分别表示在0.05和0.01水平上差异显著。缩写同表3。
表7 氮肥后移对不同氮效率水稻花后各营养器官15N累积与分配的影响(mg 15N 株-1) (盆栽, 2014年) Table 7 Effects of postponing N topdressing on post-anthesis accumulation and distribution of 15N in rice cultivars with different NUE (mg 15N plant-1) in the pot experiment (2014)
表8 氮肥后移对不同氮效率水稻花后各营养器官总碳氮比的影响(盆栽2014年) Table 8 Effects of postponing N topdressing on post-anthesis total carbon nitrogen ratio of different organs in rice cultivars with different NUE in the pot experiment (2014)
品种 Cultivar
处理 Treatment
齐穗期Full-heading stage
成熟期Maturity stage
叶 Leaf
茎鞘 Stem
根 Root
穗 Spike
叶 Leaf
茎鞘 Stem
根 Root
穗 Spike
德香4103
N0
20.45 a
97.04 a
31.08 bc
18.79 c
46.12 ab
73.71 a
59.34 a
69.03 a
Dexiang 4103
N1
15.78 de
50.89 f
29.01 cd
20.10 b
43.75 bc
37.86 b
36.72 b
56.66 b
N2
17.92 c
59.22 de
30.59 bc
23.92 a
36.63 de
30.39 c
15.14 e
46.10 c
N3
20.39 a
72.40 c
39.21 a
24.60 a
30.91 f
28.74 c
12.10 e
36.27 d
平均Average
18.64
69.89
32.47
21.85
39.35
42.68
30.83
52.02
宜香3724
N0
19.78 ab
80.16 b
29.82 cd
18.76 c
55.36 a
67.87 a
63.83 a
65.11 a
Yixiang 3724
N1
15.05 e
43.15 g
27.58 d
20.00 b
40.12 cd
38.98 b
26.91 c
58.61 b
N2
16.20 d
55.35 ef
33.28 b
23.93 a
32.84 ef
28.49 c
17.06 de
48.24 c
N3
19.04 b
63.24 d
37.00 a
24.17 a
31.16 f
36.98 b
21.21 d
39.58 d
平均Average
17.52
60.48
31.92
21.72
39.87
43.08
32.25
52.89
F 值 F-value
C
9.06* *
47.12* *
0.70
0.10
0.40
0.18
3.61
0.63
N
36.17* *
168.00* *
40.59* *
24.93* *
110.80* *
198.45* *
227.67* *
133.55* *
C× N
4.64*
3.96*
2.77
3.15
13.68* *
10.00* *
28.87* *
2.20
Values within a column followed by different letters are significantly different at P< 0.05. * Significant at P< 0.05; * * Significant at P< 0.01. Abbreviations are the same as those given in Table 3. 同栏内标以不同字母的值在5%水平上差异显著。* , * * 分别表示在0.05和0.01水平上差异显著。缩写同表3。
表8 氮肥后移对不同氮效率水稻花后各营养器官总碳氮比的影响(盆栽2014年) Table 8 Effects of postponing N topdressing on post-anthesis total carbon nitrogen ratio of different organs in rice cultivars with different NUE in the pot experiment (2014)
Haefele SM, Jabbar S M A, Siopongco J D L C, Tirol-Padre A, Amarante S T, Sta Cruz P C, Cosico W C. Nitrogen use efficiency in selected rice ( Oryza sativaL. ) genotypes under different water regimes and nitrogen levels. Field Crops Res, 2008, 107: 137-146[本文引用:1]
[2]
孙永健, 孙园园, 徐徽, 李玥, 严奉君, 蒋明金, 马均. 水氮管理模式对不同氮效率水稻氮素利用特性及产量的影响. 作物学报, 2014, 40: 1639-1649Sun YJ, Sun YY, XuH, LiY, Yan FJ, Jiang MJ, MaJ. Effects of water-nitrogen management patterns on nitrogen utilization characteristics and yield in rice cultivars with different nitrogen use efficiencies. Acta Agron Sin, 2014, 40: 1639-1649 (in Chinese with English abstract)[本文引用:6]
[3]
KumarR, Sarawgi AK, RamosC, Amarante ST, Ismail AM, Wade LJ. Partitioning of dry matter during drought stress in rainfed lowland rice. Field Crops Res, 2006, 96: 455-465[本文引用:4]
[4]
Lu YH, WatanabeA, KimuraM. Input and distribution of photosynthesized carbon in a flooded soil. Global Biogeochem Cycles, 2002, 16: 321-328[本文引用:5]
[5]
MaeT, OhiraK. The remobilization of nitrogen related to leaf growth and senescence in rice plants ( Oryza sativa L. ). Plant Cell Physiol, 1981, 22: 1067-1074[本文引用:4]
[6]
Sun YJ, MaJ, Sun YY, XuH, Yang ZY, Liu SJ, Jia XW, Zheng HZ. The effects of different water and nitrogen managements on yield and nitrogen use efficiency in hybrid rice of China. Field Crops Res, 2012, 127: 85-98[本文引用:3]
[7]
黄见良, 邹应斌, 彭少兵, Buresh RJ. 水稻对氮素的吸收、分配及其在组织中的挥发损失. 植物营养与肥料学报, 2004, 10: 579-583Huang JL, Zou YB, Peng SB, Buresh RJ. Nitrogen uptake, distribution by rice and its losses from plant tissues. Plant Nutr Fert Sci, 2004, 10: 579-583 (in Chinese with English abstract)[本文引用:4]
[8]
KrappA, Saliba-ColombaniV, Daniel-VedeleF. Analysis of C and N metabolisms and of C/N interactions using quantitative genetics. Photosynth Res, 2005, 83: 251-263[本文引用:3]
[9]
林晶晶, 李刚华, 薛利红, 张巫军, 许慧阁, 王绍华, 杨林章, 丁艳锋. 15N示踪的水稻氮肥利用率细分. 作物学报, 2014, 40: 1424-1434Lin JJ, Li GH, Xue LH, Zhang WJ, Xu HG, Wang SH, Yang LZ, Ding YF. Subdivision of nitrogen use efficiency of rice based on 15N tracer. Acta Agron Sin, 2014, 40: 1424-1434 (in Chinese with English abstract)[本文引用:5]
[10]
闫川, 洪晓富, 阮关海, 余守武, 王绍华, 丁艳锋. 大穗型水稻13C光合产物的积累与分配. 核农学报, 2014, 28: 1282-1287YanC, Hong XF, Ruan GH, Yu SW, Wang SH, Ding YF. Studies on the accumulation and transformation of assimilation product of heavy panicle type rice using 13C labeling technique. Acta Agric Nucl Sin, 2014, 28: 1282-1287 (in Chinese with English abstract)[本文引用:4]
[11]
严建民, 翟虎渠, 万建民, 焦德茂, 张荣铣. 亚种间重穗型杂交稻光合产物的运转特性及其生理机制. 中国农业科学, 2003, 36: 502-507Yan JM, Zhai HQ, Wan JM, Jiao DM, Zhang RX. Transportation characteristics of assimilate and physiologic mechanisms in subspecific heavy ear hybrid rice ( Oryza sativa L. ). Sci Agric Sin, 2003, 36: 502-507 (in Chinese with English abstract)[本文引用:5]
[12]
曾建敏, 崔克辉, 黄见良, 贺帆, 彭少兵. 水稻生理生化特性对氮肥的反应及与氮利用效率的关系. 作物学报, 2007, 33: 1168-1176Zeng JM, Cui KH, Huang JL, HeF, Peng SB. Responses of physio-biochemical properties to N-fertilizer application and its relationship with nitrogen use efficiency in rice ( Oryza sativa L. ). Acta Agron Sin, 2007, 33: 1168-1176 (in Chinese with English abstract)[本文引用:6]
[13]
叶利庭, 宋文静, 吕华军, 栗艳霞, 沈其荣, 张亚丽. 不同氮效率水稻生育后期氮素积累转运特征. 土壤学报, 2010, 47: 303-310Ye LT, Song WJ, Lyu HJ, Li YX, Shen QR, Zhang YL. Accumulation and translocation of nitrogen at late-growth stage in rices different in cultivar nitrogen use efficiency. Acta Pedol Sin, 2010, 47: 303-310 (in Chinese with English abstract)[本文引用:6]
[14]
Broadbent FE, De Datta S K, Laureles E V. Measurement of nitrogen utilization efficiency in rice genotypes. Agron J, 1987, 79: 786-791[本文引用:1]
[15]
李粹芳, 李立人. 分光光度法与14C标记法测定RuBP羧化酶的活性的比较. 植物生理学通讯, 1989, (1): 49-50Li CF, Li LR. Comparison between the spectrophotometric method and 14C-labelled method for measuring RuBPCase activity. Plant Physiol Commun, 1989, (1): 49-50 (in Chinese with English abstract)[本文引用:2]
王维, 蔡一霞, 蔡昆争, 张建华, 杨建昌, 朱庆森. 土壤水分亏缺对水稻茎秆贮藏碳水化合物向籽粒运转的调节. 植物生态学报, 2005, 29: 819-828WangW, Cai YX, Cai KZ, Zhang JH, Yang JC, Zhu QS. Regulation of soil water deficits on stem stored carbohydrate remobilization to grain of rice. Acta Phytoecol Sin, 2005, 29: 819-828 (in Chinese with English abstract)[本文引用:1]
[18]
李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000. pp 125-127Li HS. Experimental Principle and Technique for Plant Physiology and Biochemistry. Beijing: Higher Education Press, 2000. pp 125-127(in Chinese)[本文引用:1]
[19]
Lea PJ, Blackwell RD, Chen FL. Enzymes of primary metabolism. In: Harborne J B. Methods in Plant Biochemistry. Vol. 3. New York: Academic Press, 1990. pp 260-273[本文引用:1]
[20]
田纪春, 陈建省, 王延训, 张永祥. 氮素追肥后移对小麦籽粒产量和旗叶光合特性的影响. 中国农业科学, 2001, 34: 101-103Tian JC, Chen JS, Wang YX, Zhang YX. Effects of delayed-nitrogen application on grain yield and photosynthetic characteristics in flag leaves of wheat cultivars. Sci Agric Sin, 2001, 34: 101-103 (in Chinese with English abstract)[本文引用:1]
[21]
本庄一雄. 米のタンパク含量に関する研究: 第2報施肥条件のちがいが玄米のタンパク質含有率およびタンパク質総量に及ぼす影響. 日本作物學會紀事, 1971, 40: 190-196HonjyoK. Studies on protein content in rice grain: II. Effects of the fertilization on protein content and protein production in paddy grain. Jpn J Crop Sci, 1971, 40: 190-196 (in Japanese with English abstract)[本文引用:2]
[22]
王永锐, 周洁. 杂交水稻始穗期氮钾营养对剑叶生理特性的影响. 中国水稻科学, 1997, 11: 165-169Wang YR, ZhouJ. Effects of N, K supply at initial stage of panicle emerging on physiological traits in flag leaf of hybrid rice Shanyou 63. C hin J Rice Sci, 1997, 11: 165-169 (in Chinese with English abstract)[本文引用:2]
[23]
宋建民, 田纪春, 赵世杰. 植物光合碳和氮代谢之间的关系及其调节. 植物生理学通讯, 1998, 34: 230-236Song JM, Tian JC, Zhao SJ. The relationship between photosynthetic carbon and nitrogen metabolism and its regulation in plants. Plant Physiol Commun, 1998, 34: 230-236 (in Chinese with English abstract)[本文引用:1]
[24]
杨建昌, 王志琴, 朱庆森. 水稻产量源库关系的研究. 扬州大学学报(农业与生命科学版), 1993, (3): 47-53Yang JC, Wang ZQ, Zhu QS. Studies on yield source and sink relationships in rice. J Yangzhou Univ(Agric & Life Sci Edn), 1993, (3): 47-53 (in Chinese with English abstract)[本文引用:1]
[25]
Moore PA, Gilmour JT, Wells BR. Seasonal patterns of growth and soil nitrogen uptake by rice. Soil Sci Soc Am J, 1981, 45: 875-879[本文引用:1]
[26]
蒋德安, 陆庆, 翁晓燕, 郑炳松, 奚海福. 水稻光合关键酶类在光合日变化中的作用. 作物学报, 2001, 27: 301-307Jiang DA, LuQ, Weng XY, Zheng BS, Xi HF. Role of key enzymes for photosynthesis in the diurnal change of photosynthetic rate in rice. Acta Agron Sin, 2001, 27: 301-307 (in Chinese with English abstract)[本文引用:1]
[27]
Peng SB, Huang JH, Zhong XH, Yang JC, Wang GH, Zou YB, Zhang FS, Zhu QS, BureshR, WittC. Challenge and opportunity in improving fertilizer-nitrogen use efficiency of irrigated rice in China. Agric Sci China, 2002, 1: 776-785[本文引用:2]
[28]
胡健, 杨连新, 周娟, 王余龙, 朱建国. 开放式空气CO2浓度增高和施氮量对水稻结实期叶片内肽酶活力的影响. 中国水稻科学, 2008, 22: 155-160HuJ, Yang LX, ZhouJ, Wang YL, Zhu JG. Effect of free air CO2 enrichment (FACE) and nitrogen level on endopeptidase activities in rice leaves during grain filling stage. C hin J Rice Sci, 2008, 22: 155-160 (in Chinese with English abstract)[本文引用:2]
[29]
孙永健, 孙园园, 李旭毅, 郭翔, 马均. 水氮互作下水稻氮代谢关键酶活性与氮素利用的关系. 作物学报, 2009, 35: 2055-2063Sun YJ, Sun YY, Li XY, GuoX, MaJ. Relationship of activities of key enzymes involved in nitrogen metabolism with nitrogen utilization in rice under water-nitrogen interaction. Acta Agron Sin, 2009, 35: 2055-2063 (in Chinese with English abstract)[本文引用:2]
[30]
WeigeltK, KusterH, RuttenT, FaitA, Fernie AR, MierschO, WasternackC. ADP-glucose pyrophosphorylase-deficient pea embryos reveal specific transcriptional and metabolic changes of carbon-nitrogen metabolism and stress responses. Plant Physiol, 2009, 149: 395-411[本文引用:1]



{kind=link}
{kind=link}