关键词:大豆; 大豆胞囊线虫; 黄淮地区; 生理小种 Race Distribution of Soybean Cyst Nematode in the Main Soybean Producing Area of Huang-Huai Rivers Valley LIAN Yun1,**, WANG Jin-She1,**, LI Hai-Chao1, WEI He1, LI Jin-Ying1, WU Yong-Kang1, LEI Chen-Fang1, ZHANG Hui1, WANG Shu-Feng1, GUO Jian-Qiu2, LI Yue-Xia2, LI Zhi-Hui3, JIN Qiao-Ling3, XU Shu-Xia4, ZHANG Zhi-Min4, YANG Cai-Yun5, YU Hui-Yong5, GENG Zhen6, SHU Wen-Tao6, LU Wei-Guo1,* 1Zhengzhou Subcenter of National Soybean Improvement Center / Key Laboratory of Oil Crops in Huang-Huai Valleys of Ministry of Agriculture / Institute of Industrial Crops, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China
2 Luoyang Academy of Agriculture and Forestry Sciences, Luoyang 471023, China
3 Luohe Academy of Agricultural Sciences, Luohe 462300, China
4 Anyang Academy of Agricultural Sciences, Anyang 455000, China
5 Puyang Academy of Agricultural Sciences, Puyang 457000, China
6 Zhoukou Academy of Agricultural Sciences, Zhoukou 466001, China
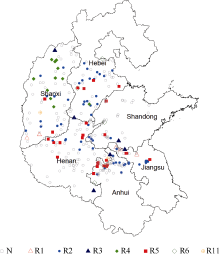
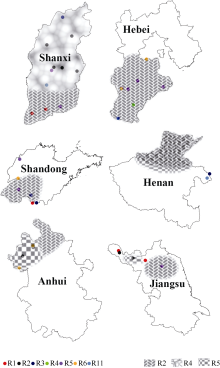
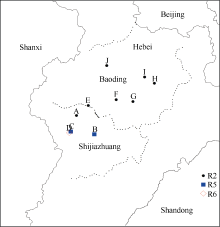
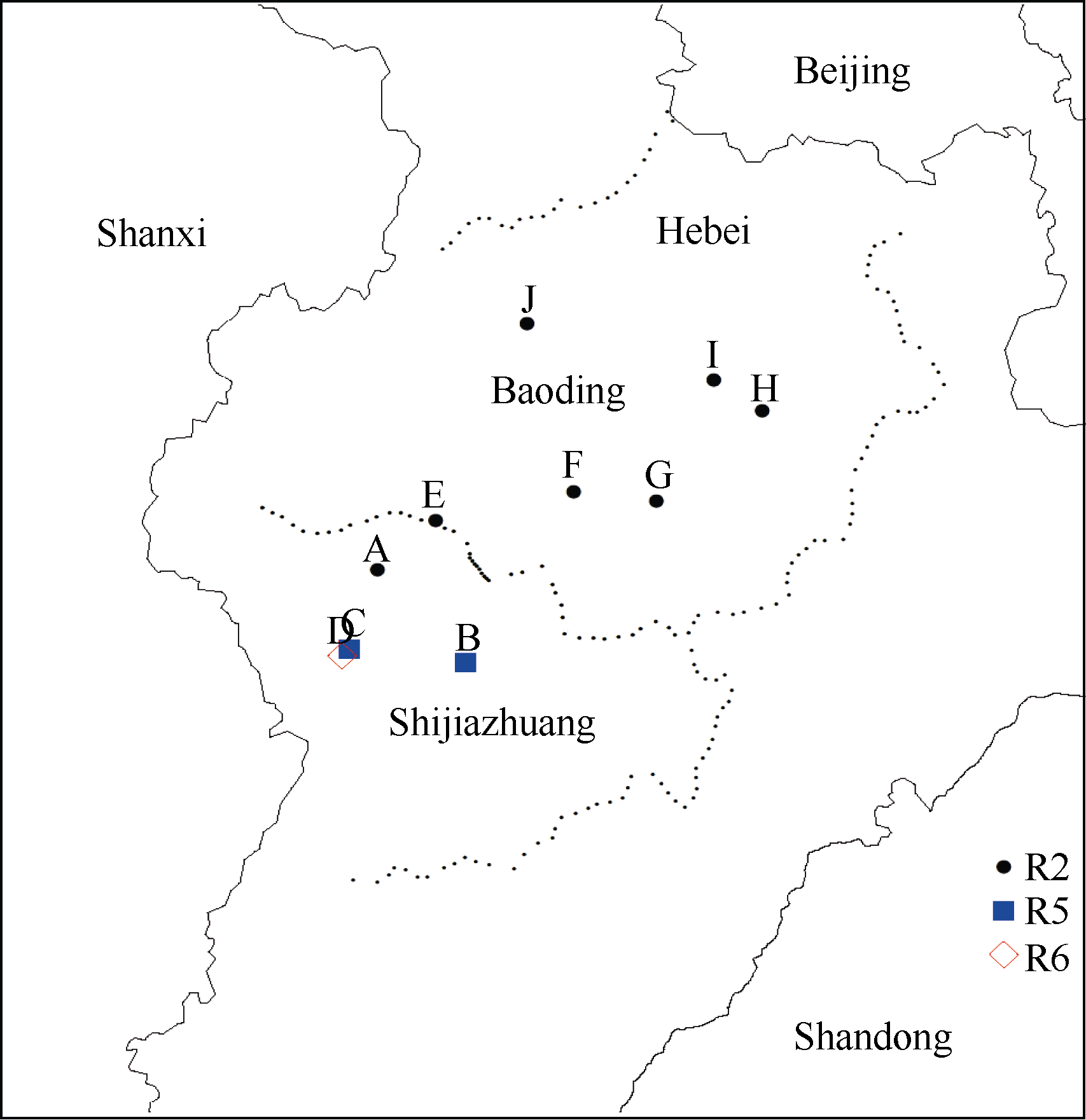
Fund:This study was supported by the National Natural Science Foundation of China (31371652), the Special Fund for Scientific Research and Development of Henan Academy of Agricultural Sciences (20137905), the China Agriculture Research System (CARS-004), and the Science and Technology Project of Henan Province (162102110141) AbstractSoybean cyst nematode (SCN) disease, caused by Heterodera glycines, occurs widely in the Huang-Huai Rivers Valley. Resistant cultivar is the most important solution to ocntrol this disease and reduce yield loss. Knowledge of SCN race distribution in soybean producing area is basis for SCN resistant breeding. A four-year survey from 2012 to 2015 was carried out to understand the diversity and distribution of Heterodera glycines races in the Huang-Huai Rivers Valley. A distribution map of races was constructed based on Riggs model. A total of 322 soil samples with SCN infection were collected. Seven H. glycines races were identified in 112 (34.8%) samples from six provinces, including five races described previously. Race 2 was the leading race, accounting for 50.9% in the 112 soil samples; race 5 was next to race 2 with the ratio of 23.2%; and the remaining types were races 4, 3, 1, 6, and 11, accounting for 9.8%, 5.4%, 4.5%, 4.5%, and 1.8%, respectively. The distribution of races varied greatly among the six provinces. According to frequencies from high to low, they were races 5, 2, 3, and 11 in Henan, races 2, 5, 6, 3, and 4 in Hebei, races 2, 5, 6, and 3 in Anhui, races 2, 4, 5, 1, 3, and 11 in Shanxi, races 2, 3, 5, 1, and 6 in Shandong, and races 2, 5, and 1 in Jiangsu. These results indicate that race 2 is the predominant race in the Huang-Huai Rivers Valley, followed by race 5, and the most virulent race 4 is centered in Shanxi province. Therefore, the SCN resistant breeding in this region should be mainly targeted towards race 2 with consideration of race 5 simultaneously, and race 4 also needs attention in some areas. In this study, races 3, 6, and 11 were newly detected in the Huang-Huai Rivers Valley. We also found that the race components and their distributions varied compared to the survey conducted in the same region in 2001-2003.
Keyword:Soybean; Soybean cyst nematode; Huang-Huai Rivers Valley; Race Show Figures Show Figures
Wrather JA, Koenning SR. Estimates of disease effects on soybean yields in the United States 2003 to 2005. J Nematol, 2006, 38: 173-180[本文引用:1]
[2]
Niblack TL, Tylka GL, Riggs RD. Nematode pathogens of soybean. Wilcox J R. Soybeans: Improvement, Production, and Uses. Madison, WI, USA: American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America, 2004. pp 821-851[本文引用:1]
[3]
Niblack TL, Wrather JA, Heinz RD, Donald PA. Distribution and virulence phenotypes of Heterodera glycines in Missouri. Plant Dis, 2003, 87: 929-932[本文引用:1]
[4]
Willson HR, Riedel RM, Eisley JB, Young CE, Jasinski JR, Wheeler TA, Kauffman PH, Pierson PE, Stuart MC. Distribution and virulence phenotypes of Heterodera glycines in Missouri. J Nematol, 1996, 28: 599-603[本文引用:1]
[5]
Mitchum MG, Wrather JA, Heinz RD, Shannon JG, DanekasG. Variability in distribution and virulence phenotypes of Heterodera glycinesin Missouri during 2005. Plant Dis, 2007, 91: 1473-1476[本文引用:1]
[6]
何艳琴, 闫晓燕, 吴存祥, 杨中路, 年海. 中国大豆新品种动态. 北京: 中国农业科学技术出版社, 2013He YQ, Yan XY, Wu CX, Yang ZL, NianH. The dynamic of new soybean variety in China. Beijing: China Agricultural Science and Technology Press, 2013[本文引用:4]
[7]
练云, 卢为国. 大豆抗SCN机制及抗病相关基因研究进展. 中国油料作物学报, 2013, 35: 727-732LianY, Lu WG. Advances on resistance mechanism and gene to SCN in soybean. Chin J Oil Crop Sci, 2013, 35: 727-732 (in Chinese with English abstract)[本文引用:1]
[8]
卢为国, 盖钧镒, 李卫东. 黄淮地区大豆胞囊线虫(Heterodera glycines Ichinohe)生理小种的抽样调查与研究. 中国农业科学, 2006, 39: 306-312Lu WG, Gai JY, Li WD. Sampling survey and identification of races of soybean cyst nematode (Heterodera glycines Ichinohe) in Huang-Huai Valleys. Sci Agric Sin, 2006, 39: 306-312 (in Chinese with English abstract)[本文引用:11]
[9]
张磊. 安徽淮北地区大豆胞囊线虫生理小种研究初报. 大豆科学, 1988, 7: 251-254ZhangL. Preliminary research on races of soybean cyst nematode (Heterodera glycines Ichinohe) in Huaibei area of Anhui province. Soybean Sci, 1988, 7: 251-254[本文引用:2]
[10]
陈品三, 张东生, 陈森玉. 大豆胞囊线虫7号生理小种的研究初报. 中国农业科学, 1987, 20(2): 94Chen PS, Zhang DS, Chen SY. First report on a new physiological race (race 7) of soybean cyst nematode (Heterodera glycines). Sci Agric Sin, 1987, 20(2): 94 (in Chinese with English abstract)[本文引用:2]
[11]
刘汉起, 商邵刚, 甄鸿杰, 霍虹, 吴和礼, 姚振纯, 李秀兰. 大豆胞囊线虫生理小种研究初报. 大豆科学, 1985, 4: 131-136Liu HQ, Shang SG, Zhen HJ, HuoH, Wu HL, Yao ZC, Li XL. Study on physiological races of soybean cyst nematode (Heterodera glycines). Soybean Sci, 1985, 4: 131-136 (in Chinese with English abstract)[本文引用:2]
[12]
李卫东, 贺春林, 田保明. 河南省大豆胞囊线虫分布及生理小种鉴定. 华北农学报, 1991, 6(增刊1): 111-114Li WD, He CL, Tian BM. Sampling survey and identification of races of soybean cystnematode (Heterodera glycines Ichinohe) in the province of Henan. Acta Agric Boreali-Sin, 1991, 6(S1): 111-114 (in Chinese with English abstract)[本文引用:2]
[13]
邢邯, 赵经荣, 战明奎, 李佩廷, 盖钧镒. 山东省大豆胞囊线虫生理小种的鉴定. 中国油料作物学报, 1997, 19(4): 61-65XingH, Zhao JR, Zhan MK, Li PT, Gai JY. Identification of races of soybean cystnematode (Heterodera glycines Ichinohe) from Shand ong province. Oil Crops China, 1997, 19(4): 61-65 (in Chinese with English abstract)[本文引用:2]
[14]
李莹, 王志, 焦广音, 常汝镇. 中国大豆遗传资源对大豆胞囊线虫4号生理小种的抗性鉴定研究. 中国农业科学, 1991, 24(5): 64-69LiY, WangZ, Jiao GY, Chang RZ. Studies on resistance of soybean germplasm resources to race 4 of soybean cyst nematode. Sci Agric Sin, 1991, 24(5): 64-69 (in Chinese with English abstract)[本文引用:2]
[15]
王衍桐, 彭德良, 陈受宜. 灰布支黑豆对大豆孢囊线虫(Heterodera glycines) 14号小种的抗性遗传. 遗传学报, 2000, 27: 146-150Wang YT, Peng DL, Chen SY. Inheritance of resistance to Heterodera glycines race 14 in Huibuzhi black bean. Acta Genet Sin, 2000, 27: 146-150 (in Chinese with English abstract)[本文引用:2]
[16]
Riggs RD, Schmitt DP. Complete characterization of the race scheme for Heterodera glycines. J Nemato, 1988, 20: 392-395[本文引用:3]
[17]
练云, 魏荷, 王金社, 雷晨芳, 李海朝, 武永康, 卢为国. 影响大豆胞囊线虫生理小种鉴定因素探讨. 分子植物育种, 2015, 13: 1259-1264LianY, WeiH, Wang JS, Lei CF, Li HC, Wu YK, Lu WG. A study on the factors that influence the race-identification of soybean cyst nematode. Mol Plant Breed, 2015, 13: 1259-1264 (in Chinese with English abstract)[本文引用:2]
[18]
王金社, 卢为国, 李金英, 练云, 魏荷, 李海朝, 雷晨芳. 植物病虫害表型数据采集系统. 专利号: 2014SR060158, 中国, 2014Wang JS, Lu WG, Li JY, LianY, WeiH, Li HC, Lei CF. The data acquisition system on the phenotype of plant diseases and insect pests. Patent number: 2014SR060158, China, 2014[本文引用:1]
[19]
Team RC. R: a language and environment for statistical computing. R Found Statist Comput, 2014[本文引用:1]
[20]
Niblack TL, Arelli PR, Noel GR, Opperman CH, Orf JH, Schmitt DP, Shannon JG, Tylka GL. A revised classification scheme for genetically diverse populations of Heterodera glycines. J Nematol, 2002, 34: 279-288[本文引用:1]
[21]
Afzal AJ, SrourA, SainiN, HemmatiN, El Shemy H A, Lightfoot D A. Recombination suppression at the dominant Rhg1/Rfs2 locus underlying soybean resistance to the cyst nematode. Theor Appl Genet, 2011, 124: 1027-1039[本文引用:1]
[22]
Yuan CP, Li YH, Liu ZX, Guan RX, Chang RZ, Qiu LJ. DNA sequence polymorphism of the Rhg4 cand idate gene conferring resistance to soybean cyst nematode in Chinese domesticated and wild soybeans. Mol Breed, 2012, 30: 1155-1162[本文引用:1]
[23]
Glover KD, WangD, Arelli PR, Carlson SR, Cianzio SR, Diers BW. Near isogenic lines confirm a soybean cyst nematode resistance gene from PI 88788 on linkage group. J Crop Sci, 2004, 44: 936-941[本文引用:1]
[24]
Vuong TD, Sleper DA, Shannon JG, Nguyen HT. Novel quantitative trait loci for broad-based resistance to soybean cyst nematode (Heterodera glycines Ichinohe) in soybean PI 567516C. Thoer Appl Genet, 2010, 121: 1253-1266[本文引用:1]
[25]
LiuX, LiuS, JamaiA, BendahmaneA, Lightfoot DA, Mitchum MG, MeksemK. Soybean cyst nematode resistance in soybean is independent of the Rhg4 locus LRR-RLK gene. Funct Integr Genomics, 2011, 11: 539-549[本文引用:1]
[26]
卢为国, 盖钧镒, 郑永战, 李卫东. 大豆遗传图谱的构建和抗胞囊线虫(Heterodera glycines Ichinohe)的QTL分析. 作物学报, 2006, 32: 1272-1279Lu WG, Gai JY, Zheng YZ, Li WD. Construction of a soybean genetic linkage map and mapping QTLs resistant to soybean cyst nematode (Heterodera glycines Ichinohe). Acta Agron Sin, 2006, 32: 1272-1279 (in Chinese with English abstract)[本文引用:1]
[27]
Concibido VC, Diers BW, Arelli PR. A decade of QTL mapping for cyst nematode resistance in soybean. Crop Sci, 2004, 44: 1121-1131[本文引用:1]
[28]
Afzal AJ, NatarajanA, SainiN, Iqbal MJ, GeislerM, El Shemy H A, Mungur R, Willmitzer L, Lightfoot D A. The nematode resistance allele at the rhg1 locus alters the proteome and primary metabolism of soybean roots. Plant Physiol, 2009, 151: 1264-1280[本文引用:1]




{kind=link}
{kind=link}
{kind=link}