关键词:普通小麦-黑麦易位系; 农艺性状; 细胞学鉴定; QTL定位; 矮秆基因 Identification of Dwarfing Wheat Germplasm SN224 and Analysis of QTLs for Its Agronomic Traits WANG Xin1,2, MA Ying-Xue1,2, YANG Yang1,2, WANG Dan-Feng1, YIN Hui-Juan1, WANG Hong- Gang1,2,* 1College of Agronomy, Shandong Agricultural University, Tai’an 271018, China
2Shandong Subcentre of National Wheat Improvement Center, Tai’an 271018, China
Fund:This study was supported by the National Key Technology R&D Program of China (2013BAD01B02-8) AbstractSN224 is a dwarfing wheat line derived from the cross between hexaploid triticale (AABBRR, 2 n= 6 x) and common wheat (AABBDD, 2 n= 6 x). We evaluated its cytologic characteristic and main agronomic traits, and analyzed the genetic basis of dwarfing trait in order to use the germplasm in wheat breeding program. This white-grain wheat had compact plant type, spindle-shaped panicle and moderate resistance to powdery mildew ( Blumeria graminis f. sp. tritici, Bgt) and stripe rust ( Puccinia striiformisf. sp. tritici, Pst). There were 42 chromosomes in root tip cells, showing 21 bivalents in pollen mother cells. FISH confirmed that SN224 was a 1BL·1RS translocation line. The plant height was between two parents of F1from a cross between SN224 and Huixianhong and distributed continuously and normally in F2 population. The detection of specific molecular marker for genes Rht-B1b, Rht-D1b, and Rht8 indicated that this line had none of the three dwarfing genes. In the meantime, the introduction of 1RS had no obvious effect on plant height. The F2 population was used to construct a genetic linkage map containing 134 SSR markers which covered a total length of 1332.1 cM. Two major dwarfing QTLs on chromosomes 1B and 4B were detected by additive-inclusive composite interval mapping (ICIM-ADD). QPh1B and QPh4B, located in the Xwmc719- Xgwm18 and Xgwm368- Xmag4284intervals, explained 20.0% and 10.2% of phenotypic variation, respectively. Seven QTLs controlling ear length, panicle number per plant, and kernel number per spike were detected. The QTL in KSUM062- Xmag4284interval contributed to decrease plant height, increase ear length and panicle number per plant.
Keyword:Common wheat-rye translocation line; Agronomic traits; Cytological identification; QTL mapping; Dwarfing gene Show Figures Show Figures
表1 SN224的主要农艺性状特点 Table 1 Main agronomic traits of SN224
性状 Trait
E1
E2
E3
范围Range
均值Mean
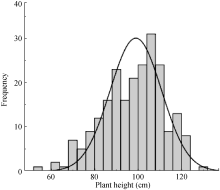
株高 Plant height (cm)
67.6± 2.9
68.0± 6.2
70.1± 2.1
67.6-70.1
68.6
穗长 Spike length (cm)
10.5± 0.9
10.4± 1.1
10.7± 0.7
10.4-10.7
10.5
单株穗数(个) Panicle number per plant
7.0± 2.0
10.2± 5.0
7.3± 2.8
7.0-10.2
8.2
每穗小穗数(个) Spikelet number per spike
23.6± 1.9
23.5± 2.4
22.9± 1.4
22.9-23.6
23.3
每穗穗粒数(粒) Kernel number per spike
58.1± 5.5
56.8± 10.9
60.4± 7.2
56.8-60.4
58.4
千粒重 Thousand-kernel weight (g)
42.2± 0.2
43.0± 1.2
40.8± 2.4
40.8-43.0
42.0
穗下节间长 Internode length below spike (cm)
20.0± 1.7
19.6± 6.2
20.2± 1.1
19.6-20.2
19.9
倒二节间长 Penultimate internode length (cm)
13.0± 0.9
12.8± 1.0
13.6± 1.0
12.8-13.6
13.1
Three field experiments were carried out in the Experimental Station of Shandong Agricultural University (Tai’ an, Shandong) in 2014 and 2015. Three environments were designated E1 for 2014, E2 for moderate soil fertility in 2015, and E3 for high soil fertility in 2015. No significant variations were detected among environments. E1、E2和E3分别指2014年山东农业大学农学实验站(山东泰安)、2015年山东农业大学农学实验站中等肥力和2015年山东农业大学农学实验站高肥力3个环境, 环境间变异不显著。
表1 SN224的主要农艺性状特点 Table 1 Main agronomic traits of SN224
图1 SN224的细胞学鉴定 a: 根尖细胞GISH; b: 花粉母细胞减数分裂MI期GISH; c: 根尖细胞FISH。Fig. 1 Cytological identification of SN224 a: GISH of root tip cell; b: GISH of pollen mother cell at MI stage; c: FISH of root tip cell.
图4 SN224/辉县红F2群体的连锁遗传图 株高、穗长、单株穗数和每穗小穗数相关QTL分别用红、绿、亮蓝和蓝色注出。Fig. 4 Genetic linkage map of SN224/Huixianhong F2 population QTL(s) associated with plant height, spike length, panicle number per plant, and kernel number per spike are marked with red, green, bright blue, and blue bars, respectively.
表2 Table 2 表2(Table 2)
表2 SN224/辉县红F2 QTL的检测结果 Table 2 Detective results of QTL in SN224/Huixianhong F2 population
PengJ, Richards DE, Hartley NM, Murphy GP, Devos KM, Flintham JE, BealesJ, Fish LJ, Worland AJ, PelicaF, SudhakarD, ChristouP, Snape JW, Gale MD, Harberd NP. “Green revolution” genes encode mutant gibberellin response modulators. Nature, 1999, 400: 256-261[本文引用:1]
[2]
HeddenP. The genes of the green revolution. Trends Genet, 2003, 19: 5-9[本文引用:1]
[3]
Saville RJ, GosmanN, Burt CJ, MakepeaceJ, SteedA, CorbittM, Chand lerE, Brown J K M, Boulton M I, Nicholson P. The ‘Green Revolution’ dwarfing genes play a role in disease resistance in Triticum aestivum and Hordeum vulgare. J Exp Bot, 2011, 63: 1271-1283[本文引用:1]
[4]
嵇怡, 缪旻珉, 陈学好. 植物矮生性状的分子遗传研究进展. 分子植物育种, 2006, 4: 753-771JiY, Miao MM, Chen XH. Progress on the molecular genetics of dwarf character in plants. Mol Plant Breed, 2006, 4: 753-771 (in Chinese with English abstract)[本文引用:1]
[5]
CadalenT, SourdilleP, CharmetG, Tixier MH, GayG, BoeufC, BernardS, LeroyP, BernardM. Molecular markers linked to genes affecting plant height in wheat using a doubled-haploid population. Theor Appl Genet, 1998, 96: 933-940[本文引用:1]
[6]
AhmadM, Sorrells ME. Distribution of microsatellite alleles linked to Rht8 dwarfing gene in wheat. Euphytica, 2002, 123: 235-240[本文引用:2]
[7]
WangZ, WuX, RenQ, ChangX, LiR, JingR. QTL mapping for developmental behavior of plant height in wheat (Triticum aestivum L. ). Euphytica, 2010, 174: 447-458[本文引用:1]
[8]
杨松杰, 张晓科, 何中虎, 夏先春, 周阳. 用STS标记检测矮秆基因Rht-B1b和Rht-D1b在中国小麦中的分布. 中国农业科学, 2006, 39: 1680-1688Yang SJ, Zhang XK, He ZH, Xia XC, ZhouY. Distribution of dwarfing genes Rht-B1b and Rht-D1b in Chinese bread wheats detected by STS marker. Sci Agric Sin, 2006, 39: 1680-1688 (in Chinese with English abstract)[本文引用:2]
[9]
贾继增, 丁寿康, 李月华, 张辉. 中国小麦的主要矮秆基因及矮源的研究. 中国农业科学, 1992, 25(1): 1-5Jia JZ, Ding SK, Li YH, ZhangH. Studies of main dwarf genes and dwarf resources on Chinese wheat. Sci Agric Sin, 1992, 25(1): 1-5 (in Chinese with English abstract)[本文引用:1]
[10]
石涛, 王洪刚, 何方, 邓世民, 高居荣. 小麦矮秆新基因的SSR标记. 山东农业科学, 2008, (8): 1-5ShiT, Wang HG, HeF, Deng SM, Gao JR. Identification of SSR markers linked to new wheat dwarf gene. Shand ong Agric Sci, 2008, (8): 1-5 (in Chinese with English abstract)[本文引用:1]
[11]
宗浩, 崔法, 鲍印广, 赵春华, 王玉海, 杜斌, 王庆专, 王洪刚. 小麦矮秆种质系山农495矮秆基因的分子标记定位. 麦类作物学报, 2009, 29: 385-389ZongH, CuiF, Bao YG, Zhao CH, Wang YH, DuB, Wang QZ, Wang HG. Developing molecular markers for the Rht gene in dwarfing germplasm line Shannong 495. J Triticeae Crops, 2009, 29: 385-389 (in Chinese with English abstract)[本文引用:1]
[12]
韩静然, 王长有, 赵宁娟, 吉万全, 赵毓, 刘雪利. 冬小麦新种质N0238D矮秆性状的遗传分析. 西北农业学报, 2010, 19(6): 60-63Han JR, Wang CY, Zhao NJ, Ji WQ, ZhaoY, Liu XL. Genetic analysis of dwarf trait in new winter wheat germplasm N0238D. Acta Agric Boreali-Occident Sin, 2010, 19(6): 60-63 (in Chinese with English abstract)[本文引用:1]
[13]
武军, 马琳, 赵继新, 陈新宏, 刘淑会, 杨群慧. 普通小麦-华山新麦草矮秆种质B62的分子细胞学鉴定. 西北农林科技大学学报(自然科学版), 2010, 38(12): 123-127WuJ, MaL, Zhao JX, Chen XH, Liu SH, Yang QH. Molecular cytology on a dwarf germplasm derived from Triticum aestivum ×Psathyrosatachys huashanica. J Northwest A&F Univ (Nat Sci Edn), 2010, 38(12): 123-127 (in Chinese with English abstract)[本文引用:1]
[14]
杨恩年, 李俊, 杨武云, 邹裕春. 矮秆大穗高产小麦育种亲本SW3243重要农艺性状特性及育种应用效果. 中国农学通报, 2010, 26(12): 114-117Yang EN, LiJ, Yang WY, Zou YC. The agronomic traits and the utilization of dwarf, large spike and high yielding potential wheat breeding parent SW3243. Chin Agric Sci Bull, 2010, 26(12): 114-117 (in Chinese with English abstract)[本文引用:1]
[15]
闫美, 刘如如, 于海涛, 达瓦, 格桑, 李安飞, 孔令让. 小麦矮秆种质山农矮330的农艺性状与矮化特性研究. 山东农业科学, 2011, (8): 14-17YanM, Liu RR, Yu HT, DaW, GeS, Li AF, Kong LR. Agronomic traits and dwarfing characteristics of wheat dwarf germplasm Shannong’ai 330. Shand ong Agric Sci, 2011, (8): 14-17 (in Chinese with English abstract)[本文引用:1]
[16]
付颖, 吴金华, 王长有, 张保军, 吉万全. 小麦新种质N0381D矮秆基因的遗传与SSR标记分析. 麦类作物学报, 2011, 31: 411-415FuY, Wu JH, Wang CY, Zhang BJ, Ji WQ. Genetic and SSR analysis of dwarf gene in new wheat germplasm N0381D. J Triticeae Crops, 2011, 31: 411-415 (in Chinese with English abstract)[本文引用:1]
[17]
王刚, 胡铁柱, 李小军, 董娜, 冯素伟, 李淦, 张立琳, 茹振钢. 小麦新品种百农矮抗58及其亲本矮秆基因的检测. 河南农业科学, 2012, 41(9): 22-25WangG, Hu TZ, Li XJ, DongN, Feng SW, LiG, Zhang LL, Ru ZG. Detection of dwarfing genes in wheat variety AK58 and its parents. J Henan Agric Sci, 2012, 41(9): 22-25 (in Chinese with English abstract)[本文引用:1]
[18]
欧俊梅, 王治斌, 周强, 任勇, 李生荣, 陶军. 糯小麦矮源新种质11-805矮秆性状遗传研究. 中国农学通报, 2014, 30(24): 152-155Ou JM, Wang ZB, ZhouQ, RenY, Li SR, TaoJ. Genetic rule of dwarf trait in new waxy dwarf wheat 11-805. Chin Agric Sci Bull, 2014, 30(24): 152-155 (in Chinese with English abstract)[本文引用:1]
[19]
张明, 吴瑕, 张一铎, 张超, 牛祖彪, 崔淑佳, 杨秋平, 王洪刚. 小麦矮秆种质系山农342-9矮秆基因的分子标记定位. 山东农业科学, 2014, 46(7): 7-10ZhangM, WuX, Zhang YD, ZhangC, Niu ZB, Cui SJ, Yang QP, Wang HG. Molecular mapping of Rht gene in wheat dwarfing germplasm line Shannong 342-9. Shand ong Agric Sci, 2014, 46(7): 7-10 (in Chinese with English abstract)[本文引用:1]
[20]
昝凯, 郑青焕, 敬樊, 陈真真, 白宇浩, 刘洋, 王亮明, 刘淑会, 王中华, 陈新宏. 小麦-大麦矮秆渗入系WB29的分子细胞学鉴定及其矮秆遗传特性分析. 农业生物技术学报, 2015, 23: 1273-1281ZanK, Zheng QH, JingF, Chen ZZ, Bai YH, LiuY, Wang LM, Liu SH, Wang ZH, Chen XH. Molecular cytogenetic identification of a wheat (Triticum aestivum L. )-barley (Hordeum vulgare ssp. distichon Hsü. ) dwarf introgression line WB29 and its dwarf traits genetic analysis. J Agric Biol, 2015, 23: 1273-1281 (in Chinese with English abstract)[本文引用:1]
[21]
崔淑佳, 卢虹, 崔雨, 高居荣, 王洪刚, 李兴锋. 小麦-长穗偃麦草矮秆种质系的鉴定及遗传分析. 核农学报, 2015, 29: 435-441Cui SJ, LuH, CuiY, Gao JR, Wang HG, Li XF. Identification and genetic analysis of dwarf wheat-Thinopyrum ponticum germplasms. J Nucl Agric Sci, 2015, 29: 435-441 (in Chinese with English abstract)[本文引用:1]
[22]
杨秋平, 杨阳, 王鑫, 马莹雪, 张明, 王洪刚. 小麦矮秆种质山农11069-5矮秆基因的遗传分析及分子定位. 分子植物育种, 2015, 13: 71-76Yang QP, YangY, WangX, Ma YX, ZhangM, Wang HG. Genetic analysis and molecular mapping of the Rht gene in dwarf germplasm Shannong 11069-5. Mol Plant Breed, 2015, 13: 71-76 (in Chinese with English abstract)[本文引用:1]
[23]
中华人民共和国农业部. NY/T1301-2007 农作物品种区域试验技术规程—小麦Ministry of Agriculture of the People’s Republic of China. NY/T1301-2007 Technical Procedures for Wheat Variety Regional Trials (in Chinese)[本文引用:1]
[24]
中华人民共和国农业部. NY/T1443. 1-2007小麦抗病虫性评价技术规范——第1部分: 小麦抗条锈病评价技术规范, 2007Ministry of Agriculture of the People`s Republic of China. NY/T1443. 1—2007 Rules for resistance evaluation of wheat to diseases and insect pests Part1: Rule for resistance evaluation of wheat to yellow rust (Puccinia striiformis West. f. sp. tritici Eriks. Et Henn. ), 2007 (in Chinese)[本文引用:1]
[25]
盛宝钦. 用反应型记载小麦苗期白粉病. 植物保护, 1988, (1): 49Sheng BQ. Scoring powdery mildew with infection type at wheat seedling stage. Plant Prot, 1988, (1): 49 (in Chinese)[本文引用:1]
[26]
HeF, XuJ, QiX, BaoY, LiX, ZhaoF, WangH. Molecular cytogenetic characterization of two partial wheat Elytrigia elongata amphiploids resistant to powdery mildew. Plant Breed, 2013, 132: 533-557[本文引用:1]
[27]
BaoY, LiX, LiuS, CuiF, WangH. Molecular cytogenetic characterization of a new wheat-Thinopyrum partial amphiploid resistant to powdery mildew and stripe rust. Cyto Geno Res, 2009, 126: 390-395[本文引用:1]
[28]
TangZ, YangZ, FuS. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119. 2, pTa-535, pTa71, CCS1, and pAWRC. 1 for FISH analysis, J Appl Genet, 2014, 55: 313-318[本文引用:1]
[29]
SteinN, HerrenG, KellerB. A new DNA extraction method for high-throughout marker analysis in a large genome species such as Triticum aestivum. Plant Breed, 2001, 120: 354-356[本文引用:1]
[30]
LiJ, CuiF, DingA, ZhaoC, WangX, WangL, BaoY, QiX, LiX, GaoJ, FengD, WangH. QTL detection of seven quality traits in wheat using two related recombinant inbred line populations. Euphytica, 2002, 183: 207-226[本文引用:1]
[31]
Kosambi DD. The estimation of map distances from recombination values. Ann Eugent, 1944, 12: 172-175[本文引用:1]
[32]
MengL, Li HH, Zhang LY, Wang JK. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J, 2015, 3: 269-283[本文引用:1]
[33]
LiH, Ribaut JM, LiZ, WangJ. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations. Theor Appl Genet, 2008, 116: 243-260[本文引用:1]
[34]
CaoG, ZhuJ, HeC, GaoY, YanJ, WuP. Impact of epistasis and QTL × environment interaction on the developmental behavior of plant height in rice (Oryza sativa L. ). Theor Appl Genet, 2001, 103: 153-160[本文引用:1]
[35]
Liu DC, Gao MQ, Guan RX, Li RZ, Cao SH, Guo XL, Zhang AM, Mapping quantitative trait loci for plant height in wheat (Triticum aestivum L. ) using a F2: 3 population. Acta Genet Sin, 2002, 29: 706-711[本文引用:1]
[36]
毕晓静. 小麦重要农艺性状的遗传分析与QTL定位. 西北农林科技大学硕士学位论文. 陕西杨凌, 2013Bi XJ. Genetic Analysis and QTL of Agronomic Traits in Wheat. MS Thesis of Dissertation of Northwest A&F University, Yangling, China, 2013[本文引用:1]
[37]
陈广凤, 陈建省, 田纪春. 小麦株高相关性状与SNP标记全基因组关联分析. 作物学报, 2015, 41: 1500-1509Chen GF, Chen JS, Tian JC. Genome-wide association analysis between SNP markers and plant height related traits in wheat. Acta Agron Sin, 2015, 41: 1500-1509 (in Chinese with English abstract)[本文引用:1]
[38]
WangY, ChenL, DuY, YangZ, Condon AG, Hu YG. Genetic effect of dwarfing gene Rht13 compared with Rht-D1b on plant height and some agronomic traits in common wheat (Triticum aestivum L. ). Field Crops Res, 2014, 162: 39-47[本文引用:1]
 , 马莹雪
, 马莹雪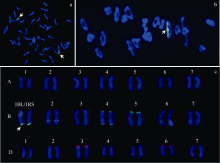






{kind=link}
{kind=link}
{kind=link}
{kind=link}