关键词:籼粳杂交稻; 灌浆结实期; 不同低温; 激素 Effect of Low Temperature after Flowering on Grain Filling and Plant Hormones Contents in Rice ZENG Yan-Hua1,2,**, ZHANG Yu-Ping2,**, PAN Xiao-Hua1, ZHU De-Feng2,*, XIANG Jing2, CHEN Hui-Zhe2 1Collaborative Innovation Center for the Modernization Production of Double Cropping Rice, Jiangxi Agricultural University / Key Laboratory of Crop Physiology, Ecology and Genetic Breeding, Ministry of Education / Jiangxi Key Laboratory of Crop Physiology, Ecology and Genetic Breeding, Nanchang 330045, China;
2State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 311400, China
Fund:This study was supported by the China Agriculture Research System (CARS-01-09B), Zhejiang Province Natural Science Foundation (Y13C130013), the Jiangxi Key Technology R&D Program (20123BBF60167), and the Doctor Activation Fundamental Research Funds for Jiangxi Agricultural University (9032305504) AbstractLow temperature (LT) after flowering impacts greatly on grain filling, decreasing rice quality through extended filling duration, shortened filling rate, and decreased physiological activity. Indica-japonica hybrid rice has been applied widely in production practice, due to high yield potential. However, there has been little study on the effect of LT on endogenous hormones in indica-japonica hybrid cultivars at filling stage. In the study, four varieties, i.e. two indica-japonica hybrid rice (Yongyou 538, YY538; Yongyou 17, YY17), one indica rice (Zhongzheyou 1, ZZY1) and one japonica rice (Zhejing 88, ZJ88), were treated with those low temperature treatments at different grain filling phases, to compare endogenous hormone levels in treatments, aiming to reveal the influence of low temperature on grain filling and the response of endogenous hormone in grains to low temperature. The effect of LT on grain filling after different flowering stage appeared significantly at early grain filling stage, while a little difference was observed at later filling stage, compared with CK. Furthermore, the influence on grain filling was found higher in Yongyou 17 and Zhongzheyou 1 than in Yongyou 538 and Zhejing 88. Compared with the contract, LT after flowering at early stage significantly decreased grain filling and prolonged grain filling duration in Yongyou 17, but not in Yongyou 538. Based on the effect of LT on grain filling process and its relation with endogenous hormones contents, it might be deduced that the normal grain filling should be ascribed to the significantly reduced GA3 content and the slightly increased IAA, ZR contents in developing grains of Yongyou 538 at early and middle grain-filling stage. On the contrary, the significantly reduced IAA, ZR contents and the significantly increased GA3, ABA contents in Yongyou17 could restrain grain development. It should enhance grain filling of rice encounted LT by selected measures adjusting hormone in practice management.
Keyword: Indica-japonica hybrid rice; Grain-filling stage; Different low temperature; Hormones Show Figures Show Figures
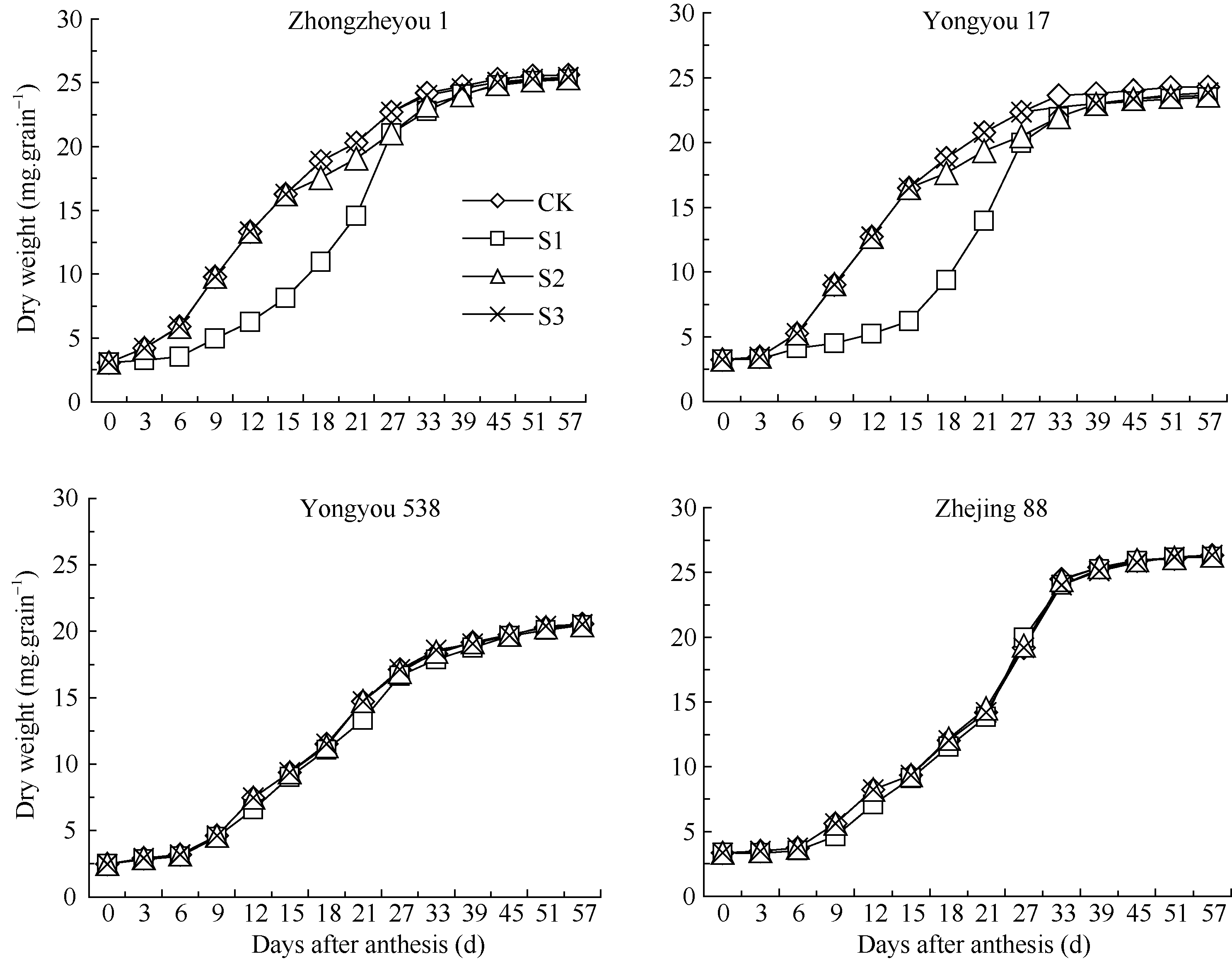
图1 灌浆期温度处理下不同品种籽粒重量动态变化S1、S2、S3为花后灌浆前、中、后期时段的低温处理。Fig. 1 Dynamics of grains weight of varieties at grain-filling stageS1, S2, S3 indicate low temperature from 0 to 15 days, from 15 to 30 days, and from 30 days after flowering to maturity, respectively.
表1 Table 1 表1(Table 1)
表1 灌浆期温度处理下籽粒灌浆过程的Richards方程参数 Table 1 Parameters of Richards equation for grain filling
品种 Variety
处理 Treatment
A
B
K
N
R2
中浙优1号 Zhongzheyou 1
CK
25.38 a
1.87
0.14
0.45
0.999
S1
24.75 c
1894.44
0.29
3.09
0.998
S2
25.12 bc
0.31
0.11
0.12
0.997
S3
25.20 b
2.08
0.14
0.48
0.999
甬优17 Yongyou 17
CK
23.98 a
10.60
0.20
1.03
0.999
S1
23.09 c
327747.90
0.46
5.22
0.996
S2
23.23 bc
1.93
0.15
0.44
0.996
S3
23.32 b
18.90
0.23
1.28
0.999
甬优538 Yongyou 538
CK
20.05 a
16.70
0.16
1.18
0.998
S1
20.05 a
11.62
0.14
1.03
0.998
S2
20.04 a
16.88
0.16
1.19
0.998
S3
20.07 a
19.29
0.17
1.24
0.998
浙粳88 Zhejing 88
CK
26.24 a
455.22
0.22
2.76
0.999
S1
26.10 a
430.14
0.22
2.56
0.999
S2
26.29 a
257.37
0.21
2.47
0.999
S3
26.28 a
175.36
0.19
2.30
0.999
A indicates final grain weight; B, K, and N indicate equation parameters of Richards; R2 indicates determination coefficient. Values followed by different letters are significantly different at the 0.05 probability level. S1, S2, S3 indicate low temperature from 0 to 15 days, from 15 to 30 days, and from 30 days after flowering to maturity, respectively. A为籽粒最终重量(mg); B、K和N为Richards方程参数; R2为决定系数。标以不同字母的值在5%水平上差异显著。S1、S2、S3为花后灌浆前、中、后期时段的低温处理。
表1 灌浆期温度处理下籽粒灌浆过程的Richards方程参数 Table 1 Parameters of Richards equation for grain filling
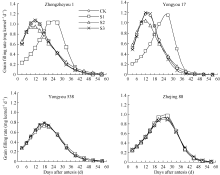
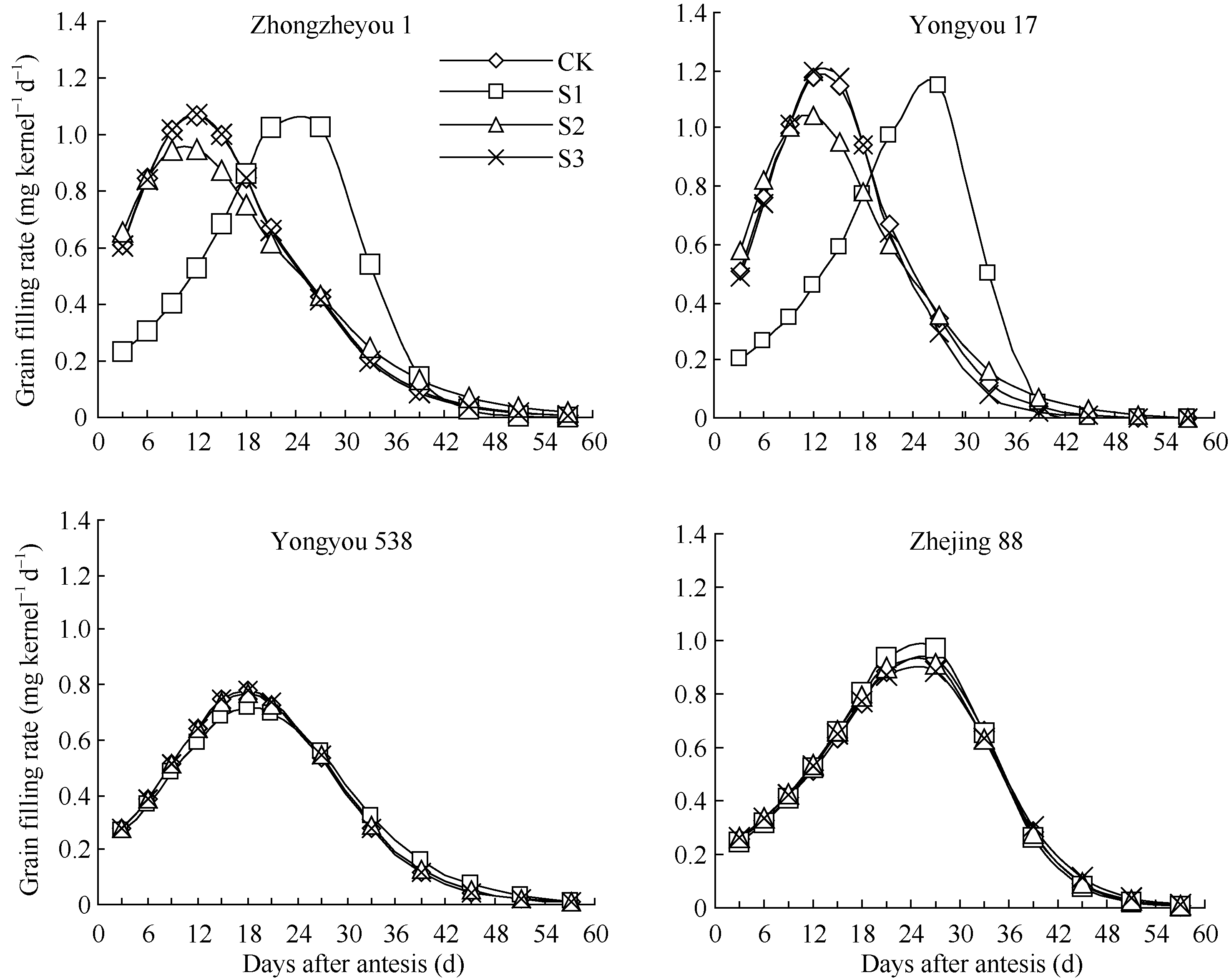
图2 灌浆期温度处理下不同品种籽粒灌浆速率变化S1、S2、S3为花后灌浆前、中、后期时段的低温处理。Fig. 2 Dynamics of grain filling rate of varieties in low temperature treatmentsS1, S2, S3 indicate low temperature from 0 to 15 days, from 15 to 30 days, and from 30 days after flowering to maturity, respectively.
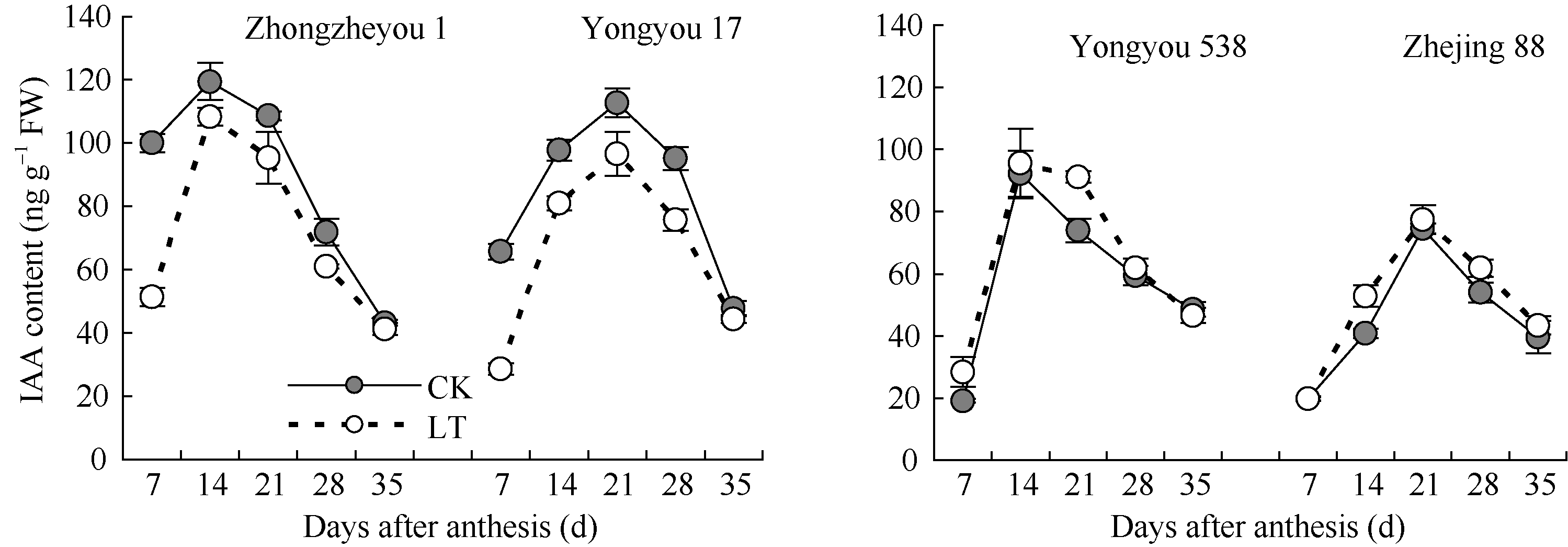
图3 灌浆期低温对籽粒IAA含量的影响LT: 低温处理, 包括花后灌浆前期第7、第14天, 灌浆中期第21、第28天, 灌浆后期第35天。Fig. 3 Effect of low temperature during grain-filling on IAA content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment.
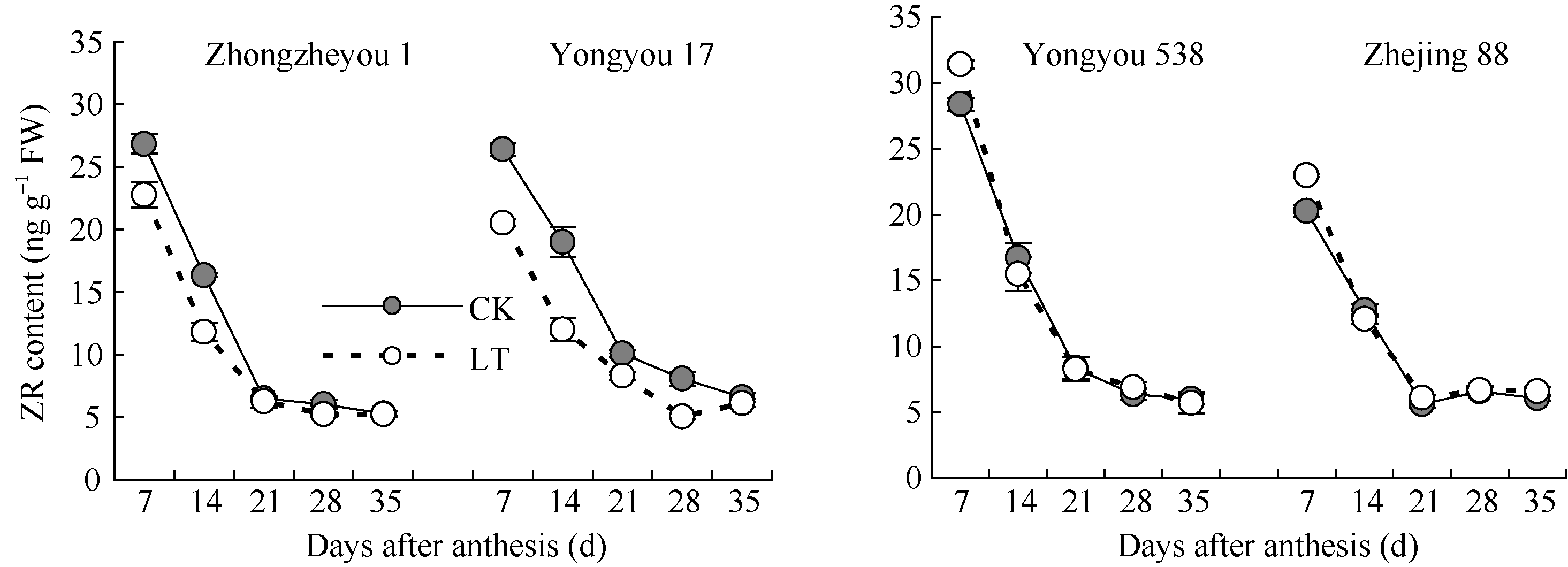
图4 灌浆期低温对籽粒ZR含量的影响LT: 低温处理, 包括花后灌浆前期第7、14天, 灌浆中期第21、28天, 灌浆后期第35天。Fig. 4 Effect of low temperature during grain-filling on ZR content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment.
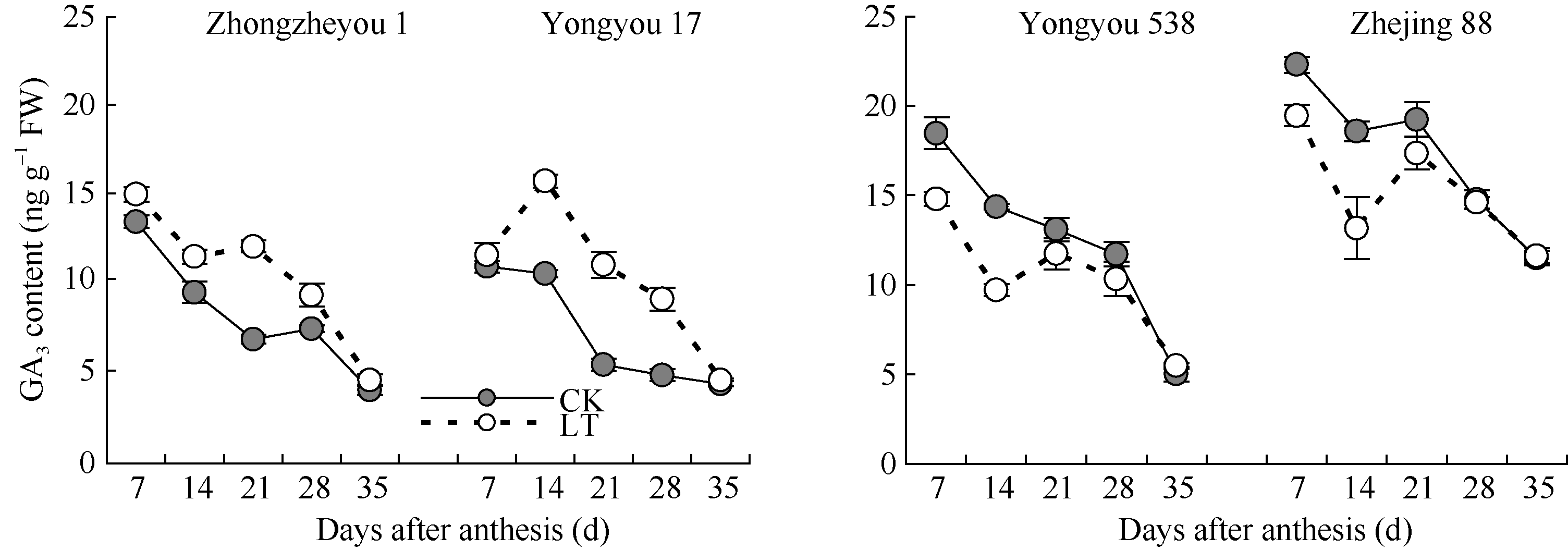
图5 灌浆期低温对籽粒GA3含量的影响LT: 低温处理, 包括花后灌浆前期第7、第14天, 灌浆中期第21、第28天, 灌浆后期第35天。Fig. 5 Effect of low temperature during grain-filling on GA3 content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment.
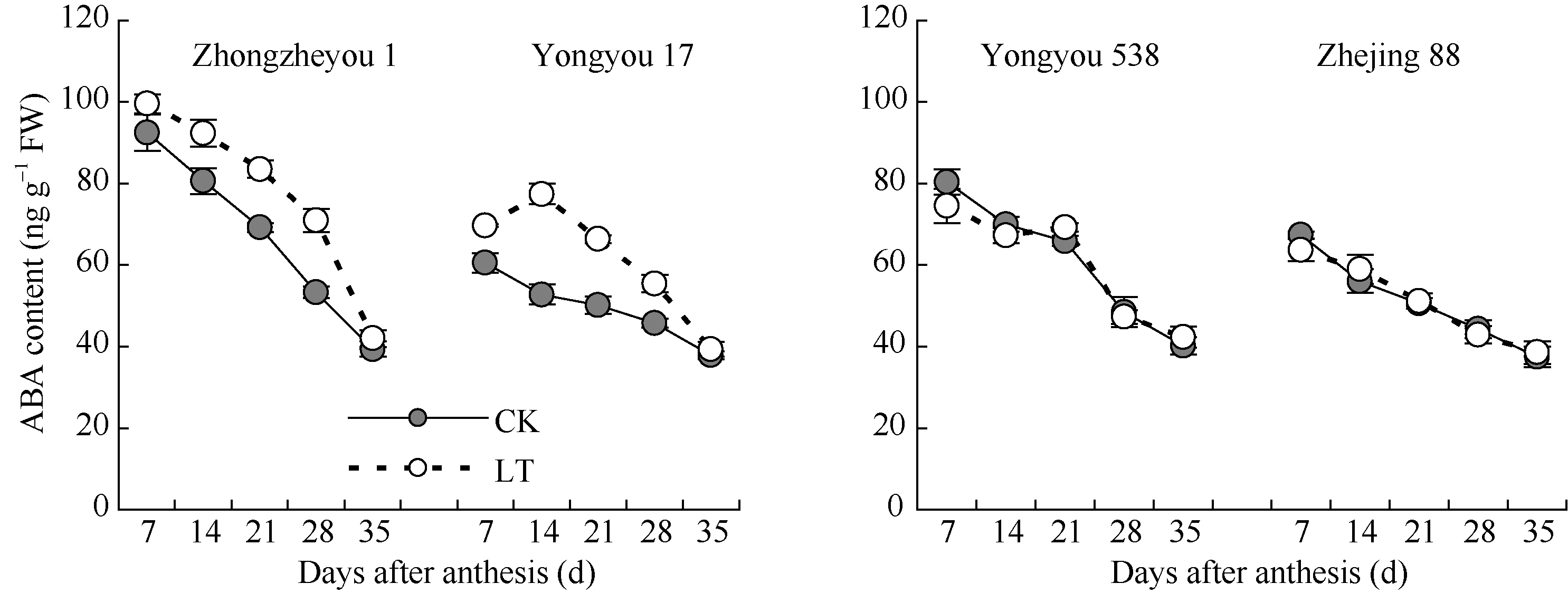
图6 灌浆期低温对籽粒ABA含量的影响LT: 低温处理, 包括花后灌浆前期第7、14天, 灌浆中期第21、28天, 灌浆后期第35天。Fig. 6 Effect of low temperature during grain-filling on ABA content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment.
表2 花后不同温度处理下水稻籽粒灌浆与籽粒中内源激素含量变化的相关分析 Table 2 Correlation coefficients between grain filling and changes of endogenous hormone contents in different temperature treatments
指标 Index
IAA
ZR
GA3
ABA
籽粒灌浆 Grain filling
IAA
1.000
ZR
-0.185
1.000
GA3
-0.266
0.461* *
1.000
ABA
0.300
0.601* *
0.390*
1.000
籽粒灌浆 Grain filling
0.189
-0.843* *
-0.683* *
-0.684* *
1.000
* P < 0.05, * * P < 0.01.
表2 花后不同温度处理下水稻籽粒灌浆与籽粒中内源激素含量变化的相关分析 Table 2 Correlation coefficients between grain filling and changes of endogenous hormone contents in different temperature treatments
刘吉峰, 丁裕国, 江志红. 全球变暖加剧对极端气候概率影响的初步探讨. 高原气象, 2007, 26: 837-842Liu JF, Ding YG, Jiang ZH. The influence of aggravated global warming on the probability of extreme climatic event. Plateau Meteorol, 2007, 26: 837-842 (in Chinese with English abstract)[本文引用:1]
[2]
MarianiL, Parisi SG, ColaG. Space and time behavior of climatic hazard of low temperature for single rice crop in the mid latitude. Int J Climatol, 2009, 29: 1862-1871[本文引用:1]
[3]
王丰, 程方民, 刘奕, 钟连进, 张国平. 不同温度下灌浆期水稻籽粒内源激素含量的动态变化. 作物学报, 2006, 32: 25-29WangF, Cheng FM, LiuY, Zhong LJ, Zhang GP. Dynamic changes of plant hormones in developing grains at rice filling stage under different temperatures. Acta Agron Sin, 2006, 32: 25-29 (in Chinese with English abstract)[本文引用:6]
[4]
金正勋, 杨静, 钱春荣, 刘海英, 金学泳, 秋太权. 灌浆成熟期温度对水稻籽粒淀粉合成关键酶活性及品质的影响. 中国水稻科学, 2005, 19: 377-380Jin ZX, YangJ, Qian CR, Liu HY, Jin XY, Qiu TQ. Effects of temperature during grain filling period on activities of key enzymes for starch synthesis and rice grain quality. Chin J Rice Sci, 2005, 19: 377-380 (in Chinese with English abstract)[本文引用:1]
[5]
程方民, 钟连进, 孙宗修. 灌浆结实期温度对早籼水稻籽粒淀粉合成代谢的影响. 中国农业科学, 2003, 36: 492-501Cheng FM, Zhong LJ, Sun ZX. Effect of temperature at grain-filling stage on starch biosynthetic metabolism in developing rice grains of early-indica. Sci Agric Sin, 2003, 36: 492-501 (in Chinese with English abstract)[本文引用:2]
[6]
金正勋, 钱春荣, 杨静, 刘海英. 水稻灌浆成熟期籽粒谷氨酰胺合成酶活性变化及其与稻米品质关系的初步研究. 中国水稻科学, 2007, 21: 103-106. Jin ZX, Qian CR, YangJ, Liu HY. Changes of activity of glutamine synthetase during grain filling and its influence on rice grain quality. Chin J Rice Sci, 2007, 21: 103-106 (in Chinese with English abstract)[本文引用:1]
[7]
Mohammed AR, TarpleyL. Effects of high night temperature and spikelet position on yield related parameters of rice (Oryza sativa L. ) plants. Eur J Agron, 2010, 33: 117-123[本文引用:1]
[8]
Mohammed AR, TarpleyL. High night temperature and plant growth regulator effects on spikelet sterility, grain characteristics and yield of rice (Oryza sativa L. ) plants. Can J Plant Sci, 2011, 91: 283-291[本文引用:1]
[9]
曹云英. 高温对水稻产量与品质的影响及其生理机制. 扬州大学硕士学位论文, 江苏扬州, 2009Cao YY. Effect of High Temperature on the Quality and Quantity of Rice Yield and Its Physiological Mechanism. MS Thesis of Yangzhou University, Yangzhou, China, 2009[本文引用:1]
[10]
张桂莲. 两个早稻品种耐热性生理及分子遗传基础的研究, 湖南农业大学博士论文, 湖南长沙, 2006Zhang GL. Studies on Physiological and Molecular Genetics Basis of Heat Tolerance of Two Early Rice Cultivars. PhD Dissertation of Hunan Agricultural University, Changsha, China, 2006 (in Chinese with English abstract)[本文引用:1]
[11]
许光利. 水稻灌桨结实期高温弱光对籽粒脂类代谢的影响. 四川农业大学硕士学位论文, 四川雅安, 2011Xu GL. Effects of High-temperature and Weak-light on Lipid Metabolism During Rice Grain-filling Stage. MS Thesis of Sichuan Agricultural University, Ya’an, China, 2011[本文引用:1]
[12]
赵丽晓, 张萍, 王若男, 王璞, 陶洪斌. 花后前期高温对玉米强弱势籽粒生长发育的影响. 作物学报, 2014, 40: 1839-1845Zhao LX, ZhangP, Wang RN, WangP, Tao HB. Effect of high temperature after flowering on growth and development of superior and inferior maize kernels. Acta Agron Sin, 2014, 40: 1839-1845 (in Chinese with English abstract)[本文引用:5]
[13]
袁莉民, 常二华, 徐伟, 王志琴, 杨建昌. 结实期低温对杂交水稻胚乳结构的影响. 作物学报, 2006, 32: 96-102Yuan LM, Chang EH, XuW, Wang ZQ, Yang JC. Effects of low temperature during grain filling on the structure of endosperm in hybrid rice. Acta Agron Sin, 2006, 32: 96-102 (in Chinese with English abstract)[本文引用:3]
[14]
付景, 王志琴, 杨建昌. 抽穗-灌浆期低温与弱光对超级稻结实率和生理性状的影响. 扬州大学学报(农业与生命科学版), 2014, 35(4): 68-74FuJ, Wang ZQ, Yang JC. Effects of low temperature and weak light during heading and grain filling on the seed-setting rate and physiological traits of super rice. J Yangzhou Univ (Agric & Life Sci Edn), 2014, 35(4): 68-74 (in Chinese with English abstract)[本文引用:2]
[15]
耿立清, 王嘉宇, 陈温福. 孕穗-灌浆期低温对水稻穗部性状的影响. 华北农学报, 2009, 24(3): 107-111Geng LQ, Wang JY, Chen WF. Effect of low temperature on panicle characters of rice during booting and grain filling period. Acta Agric Boreali-Sin, 2009, 24(3): 107-111 (in Chinese with English abstract)[本文引用:1]
[16]
杨杰, 翟虎渠, 王才林, 仲维功, 邹江石, 池桥宏, 万建民. 水稻籼粳亚种间杂种低温花粉不育的QTL分析. 遗传学报, 2005, 32: 507-514YangJ, Zhai HQ, Wang CL, Zhong WG, Zou JS, IkehashiH, Wan JM. QTL analysis of low-temperature-sensitive pollen sterility in indica-japonica hybrid rice (Oryza sativa L. ). Acta Genet Sin, 2005, 32: 507-514 (in Chinese with English abstract)[本文引用:1]
[17]
花劲, 周年兵, 张军, 张洪程, 霍中洋, 周培建, 程飞虎, 李国业, 黄大山, 陈忠平, 陈国梁, 戴其根, 许轲, 魏海燕, 高辉, 郭保卫. 双季晚稻甬优系列籼粳杂交稻超高产结构与群体形成特征. 中国农业科学, 2015, 48: 1023-1034HuaJ, Zhou NB, ZhangJ, Zhang HC, Huo ZY, Zhou PJ, Cheng FH, Li GY, Huang DS, Chen ZP, Chen GL, Dai QG, XuK, Wei HY, GaoH, Guo BW. The structure and formation characteristics of super-high yield population with late Yongyou series of indica-japonica hybrid rice in double-cropping rice area. Sci Agric Sin, 2015, 48: 1023-1034 (in Chinese with English abstract)[本文引用:1]
[18]
韦还和, 姜元华, 赵可, 许俊伟, 张洪程, 戴其根, 霍中洋, 许轲, 魏海燕, 郑飞. 甬优系列杂交稻品种的超高产群体特征. 作物学报, 2013, 39: 2201-2210Wei HH, Jiang YH, ZhaoK, Xu JW, Zhang HC, Dai QG, Huo ZY, XuK, Wei HY, ZhengF. Characteristics of super-high yield population in Yongyou series of hybrid rice. Acta Agron Sin, 2013, 39: 2201-2210 (in Chinese with English abstract)[本文引用:1]
[19]
姜元华, 许俊伟, 赵可, 韦还和, 孙建军, 张洪程, 戴其根, 霍中洋, 许轲, 魏海燕, 郭保卫. 甬优系列籼粳杂交稻根系形态与生理特征. 作物学报, 2015, 41: 89-99Jiang YH, Xu JW, ZhaoK, Wei HH, Sun JJ, Zhang HC, Dai QG, Huo ZY, XuK, Wei HY, Guo BW. Root system morphological and physiological characteristics of indica-japonica hybrid rice of Yongyou series. Acta Agron Sin, 2015, 41: 89-99 (in Chinese with English abstract)[本文引用:1]
[20]
曾研华, 张玉屏, 王亚梁, 向镜, 陈惠哲, 朱德峰. 甬优系列杂交稻组合开花期耐冷性评价. 中国水稻科学, 2015, 29: 291-298Zeng YH, Zhang YP, Wang YL, XiangJ, Chen HZ, Zhu DF. Evaluation of chilling resistance in Yongyou series of hybrid rice at flowering stage. Chin J Rice Sci, 2015, 29: 291-298 (in Chinese with English abstract)[本文引用:2]
[21]
朱庆森, 曹显祖, 骆亦其. 水稻籽粒灌浆的生长分析. 作物学报, 1988, 14: 183-193Zhu QS, Cao XZ, Luo YQ. Growth analysis on the process of grain filling in rice. Acta Agron Sin, 1988, 14: 183-193 (in Chinese with English abstract)[本文引用:1]
[22]
顾世梁, 朱庆森, 杨建昌, 彭少兵. 不同水稻材料籽粒灌浆特性的分析. 作物学报, 2001, 27: 7-14Gu SL, Zhu QS, Yang JC, Peng SB. Analysis on grain filling characteristics for different rice types. Acta Agron Sin, 2001, 27: 7-14 (in Chinese with English abstract)[本文引用:1]
[23]
Yang JC, Zhang JH, Wang ZQ, Zhu QS, WangW. Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant Physiol, 2001, 127: 315-323[本文引用:1]
[24]
高亮之, 金之庆. 全球气候变化和中国的农业. 江苏农业学报, 1994, 10(2): 1-10Gao LZ, Jin ZQ. Global climate change and China’s agriculture. Jiangsu J Agric Sci, 1994, 10(2): 1-10 (in Chinese with English abstract)[本文引用:1]
[25]
AhmedN, MaekawaM, Tetlow IJ. Effects of low temperature on grain filling, amylose content, and activity of starch biosynthesis enzymes in endosperm of basmati rice. Aust J Agric Res, 2008, 59: 599-604[本文引用:2]
[26]
朱碧岩, 程方民, 吴永常, 方辉. 稻米粒重形成规律与结实期温度的关系. 西北农业大学学报, 1996, 24(4): 53-58Zhu BY, Cheng FM, Wu YC, FangH. A study on the dynamic changes of rice seed weight and the temperature effect during the full heading stage. Acta Univ Agric Boreali-occident, 1996, 24(4): 53-58 (in Chinese with English abstract)[本文引用:1]
[27]
Brenner ML, CheikhN. The role of hormones in photosynthate partitioning and seed filling. In: Davies P J ed. Plant Hormones. Netherland s: Springer Kluwer Academic Publishers, 1995. pp 649-670[本文引用:1]
[28]
RookF, CorkeF, CardR, MunzG, SmithC, Bevan MW. Impaired sucrose-induction mutants reveal the modulation of sugar-induced starch biosynthetic gene expression by abscisic acid signalling. Plant J, 2001, 26: 421-433[本文引用:1]
[29]
王艳芳, 崔震海, 阮燕晔, 马兴林, 关义新, 张立军. 不同类型春玉米灌浆期间籽粒中内源激素IAA、GA、ZR、ABA含量的变化. 植物生理学通讯, 2006, 42: 225-228Wang YF, Cui ZH, Ruan RY, Ma XL, Guan YX, Zhang LJ. Changes in endogenous hormone of IAA, GA, ZR and ABA in kernels during grain-filling stage in different types of spring maize (Zea mays L. ). Plant Physiol Commun, 2006, 42: 225-228 (in Chinese with English abstract)[本文引用:1]
[30]
徐云姬, 顾道健, 张博博, 张耗, 王志琴, 杨建昌. 玉米果穗不同部位籽粒激素含量及其与胚乳发育和籽粒灌浆的关系. 作物学报, 2013, 39: 1452-1461Xu YJ, Gu DJ, Zhang BB, ZhangH, Wang ZQ, Yang JC. Hormone contents in kernels at different positions on an ear and their relationship with endosperm development and kernel filling in maize. Acta Agron Sin, 2013, 39: 1452-1461 (in Chinese with English abstract)[本文引用:1]
[31]
RookF, CorkerF, CardR. Impaired sucrose-induction mutants reveal the modulation of sugar-induced starch biosynthetic gene expression by abscisic acid signaling. Plant J, 2001, 26: 421-433[本文引用:2]
[32]
Yang JC, ZhangH. Hormones in the grains and roots in relation to post-anthesis development of inferior and superior spikelets in japonica/indica hybrid rice. Plant Physiol Biochem, 2009, 47: 195-204[本文引用:1]
[33]
王兴, 于晶, 杨阳, 苍晶, 李卓夫. 低温条件下不同抗寒性冬小麦内源激素的变化. 麦类作物学报, 2009, 29: 827-831WangX, YuJ, YangY, CangJ, Li ZF. Changes of endogenous hormones of winter wheat varieties with different cold-resistances under low temperature. J Triticeae Crops, 2009, 29: 827-831 (in Chinese with English abstract)[本文引用:1]
[34]
盛婧, 陶红娟, 陈留根. 灌浆结实期不同时段温度对水稻结实与稻米品质的影响. 中国水稻科学, 2007, 21: 396-402ShengJ, Tao HJ, Chen LG. Response of seed-setting and grain quality of rice to temperature at different time during grain filling period. Chin J Rice Sci, 2007, 21: 396-402 (in Chinese with English abstract)[本文引用:1]
 , 向镜
, 向镜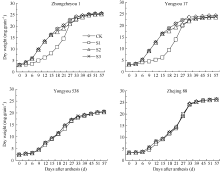





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}