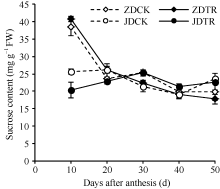
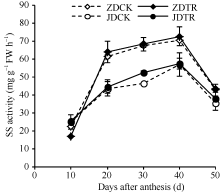
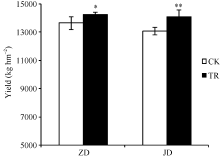
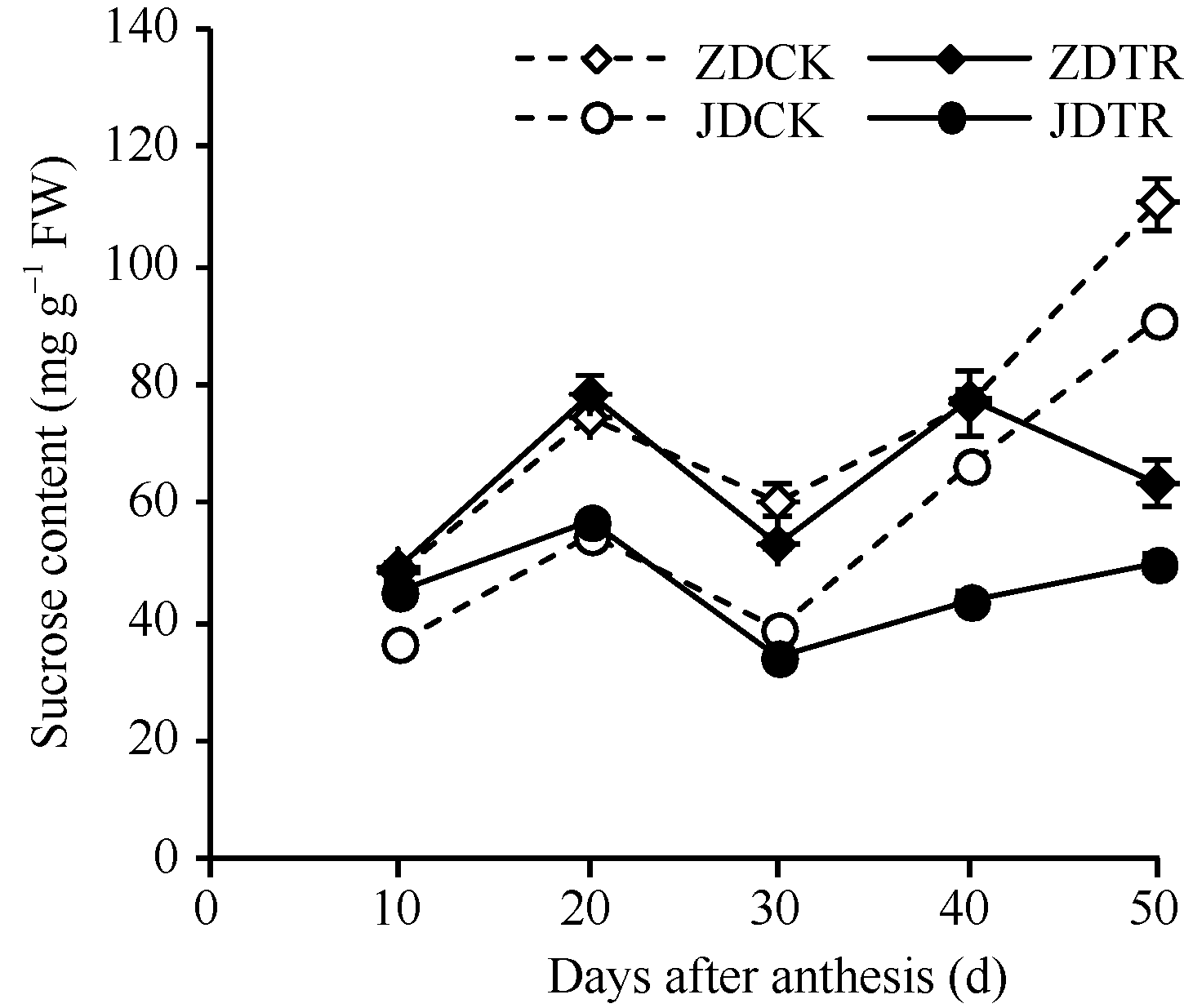
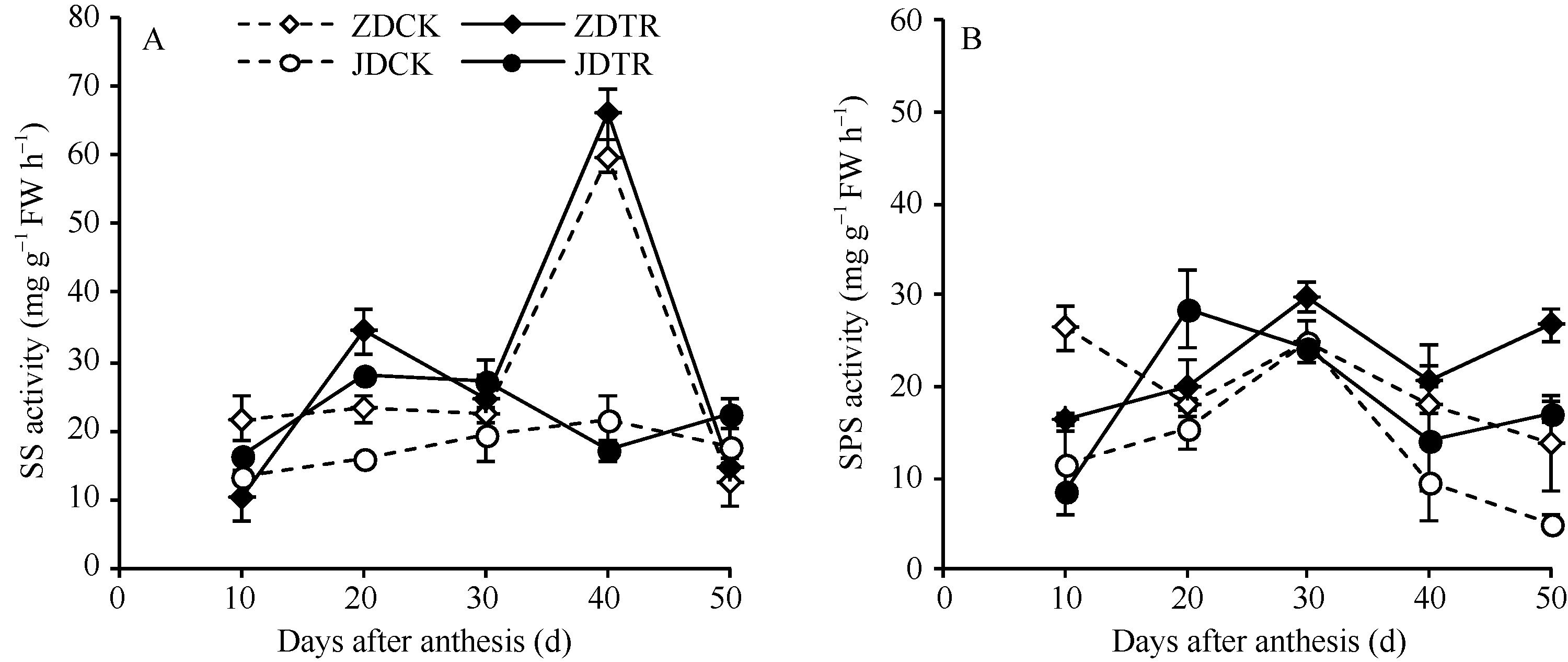
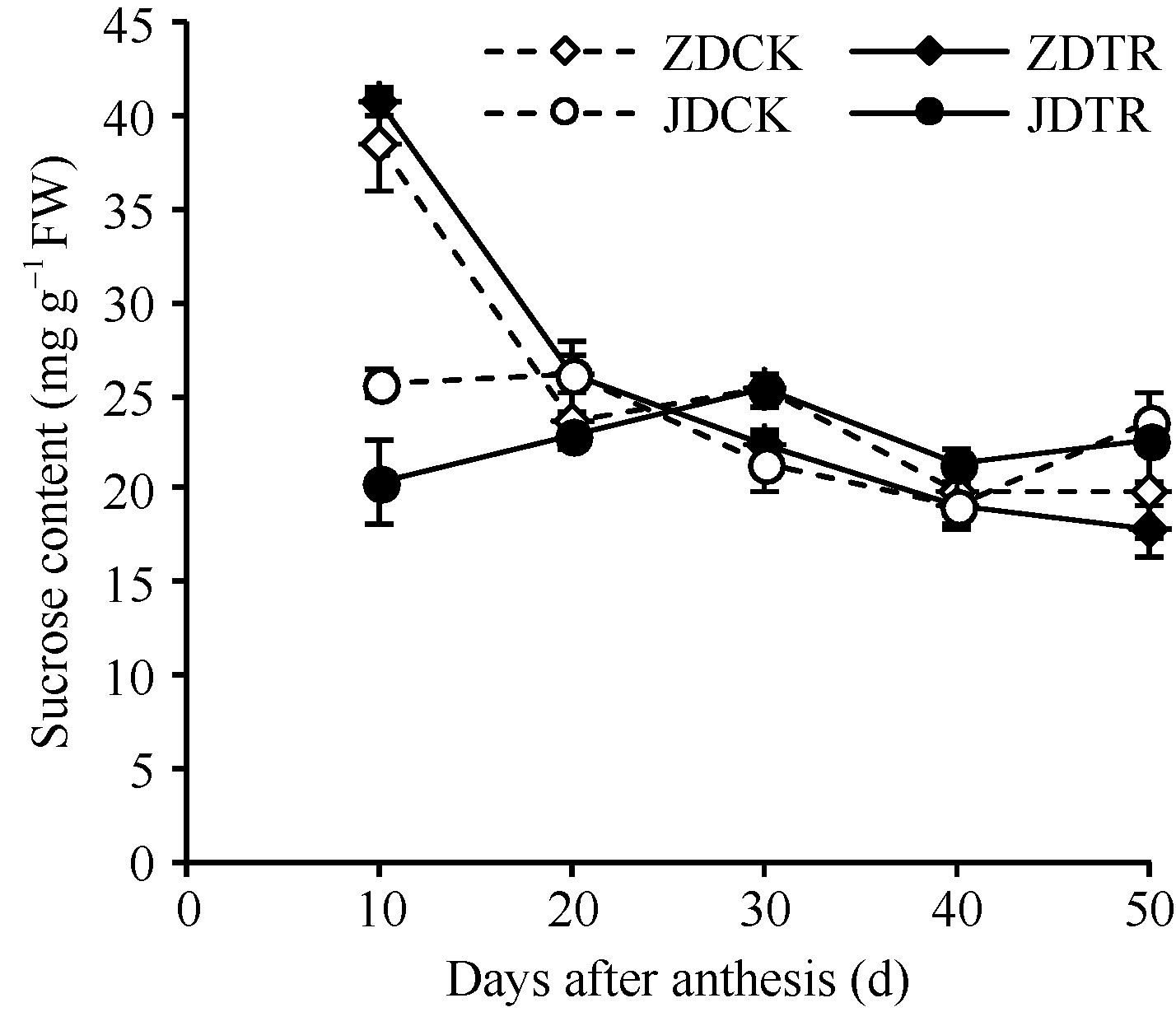
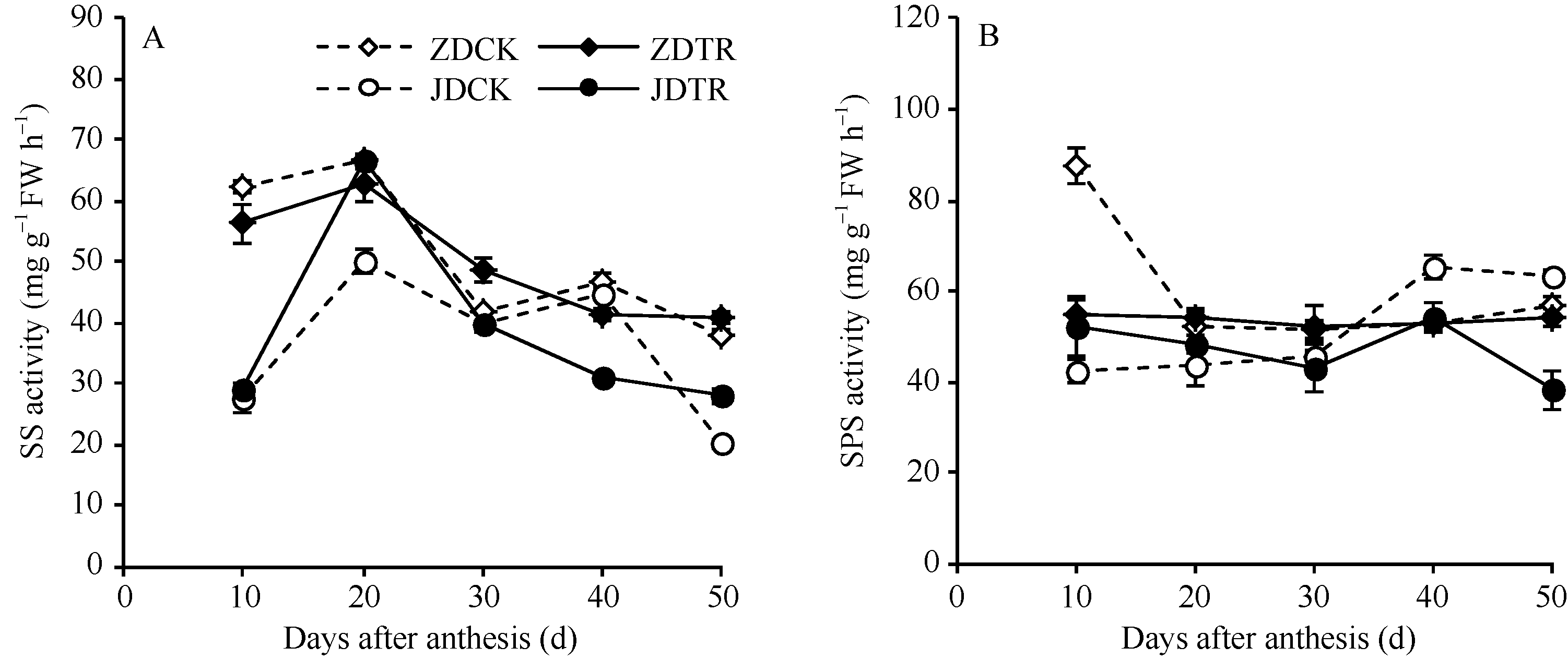
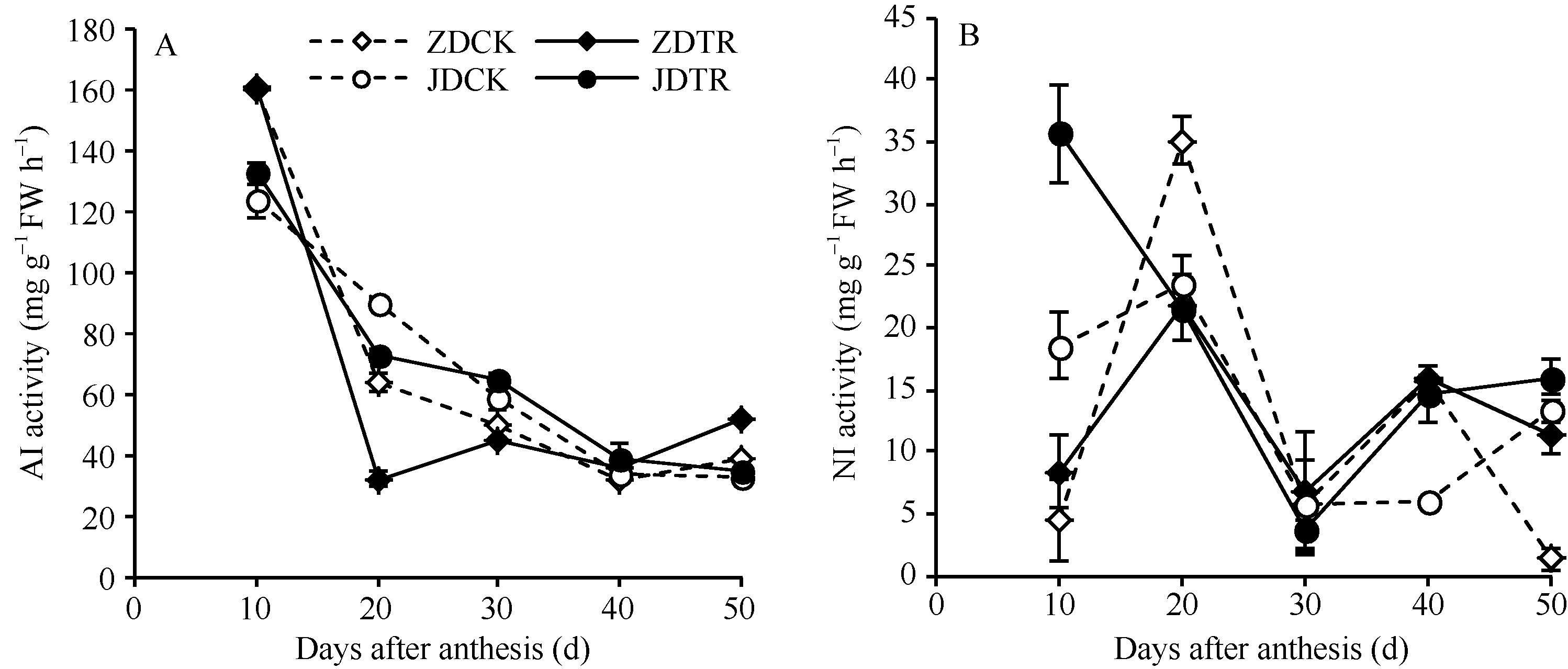
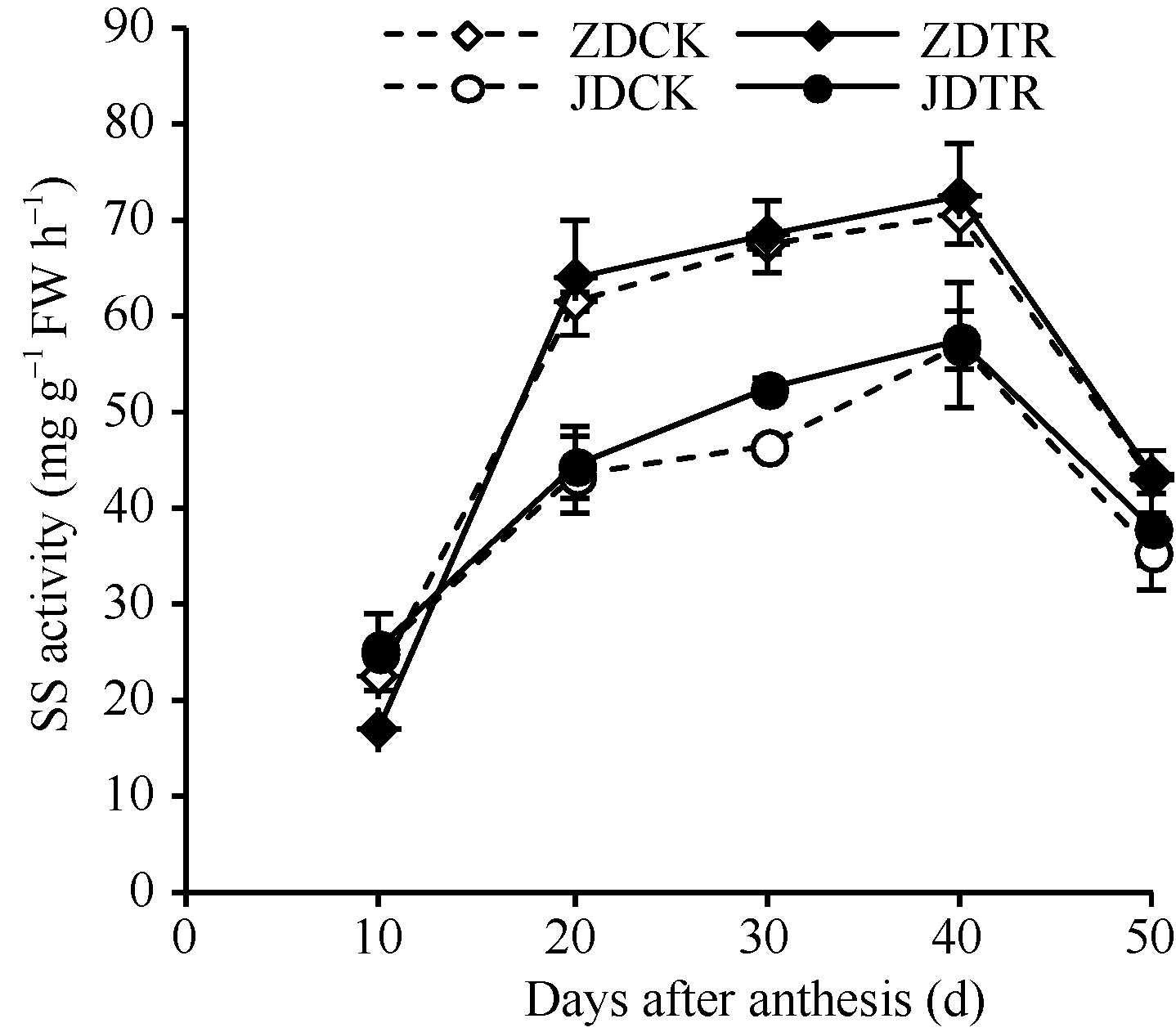
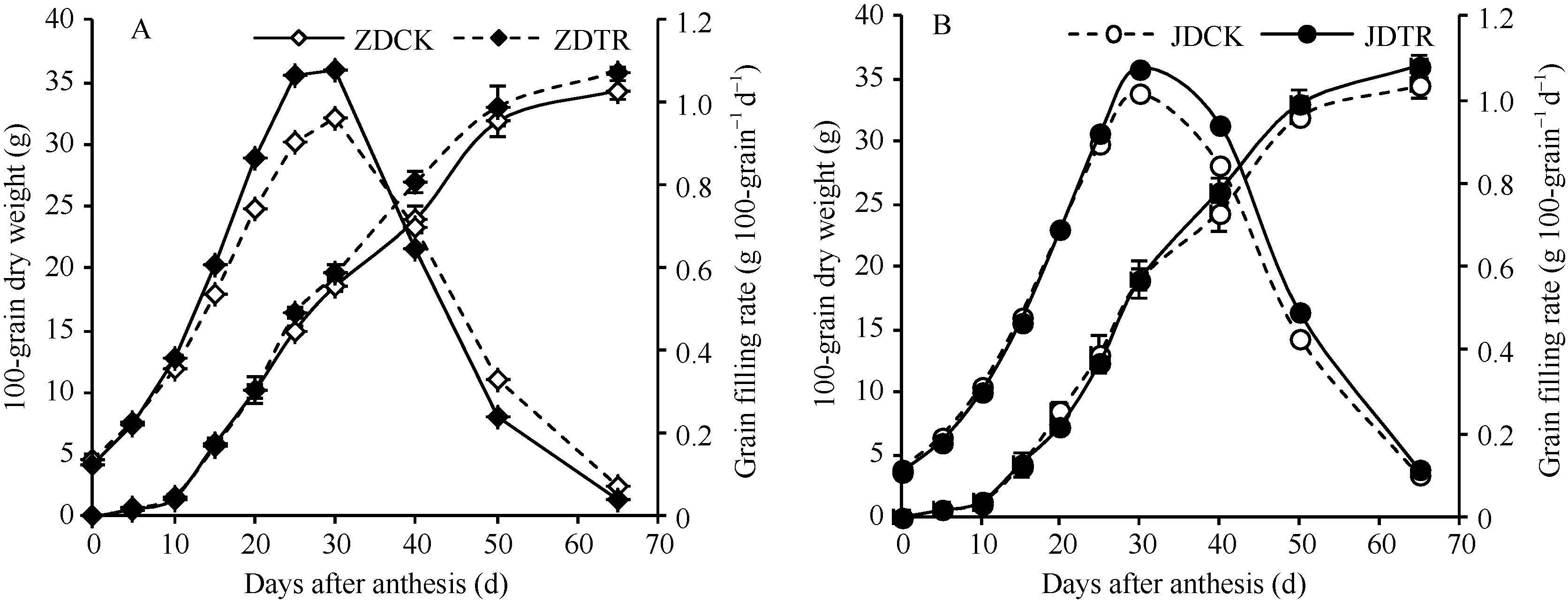
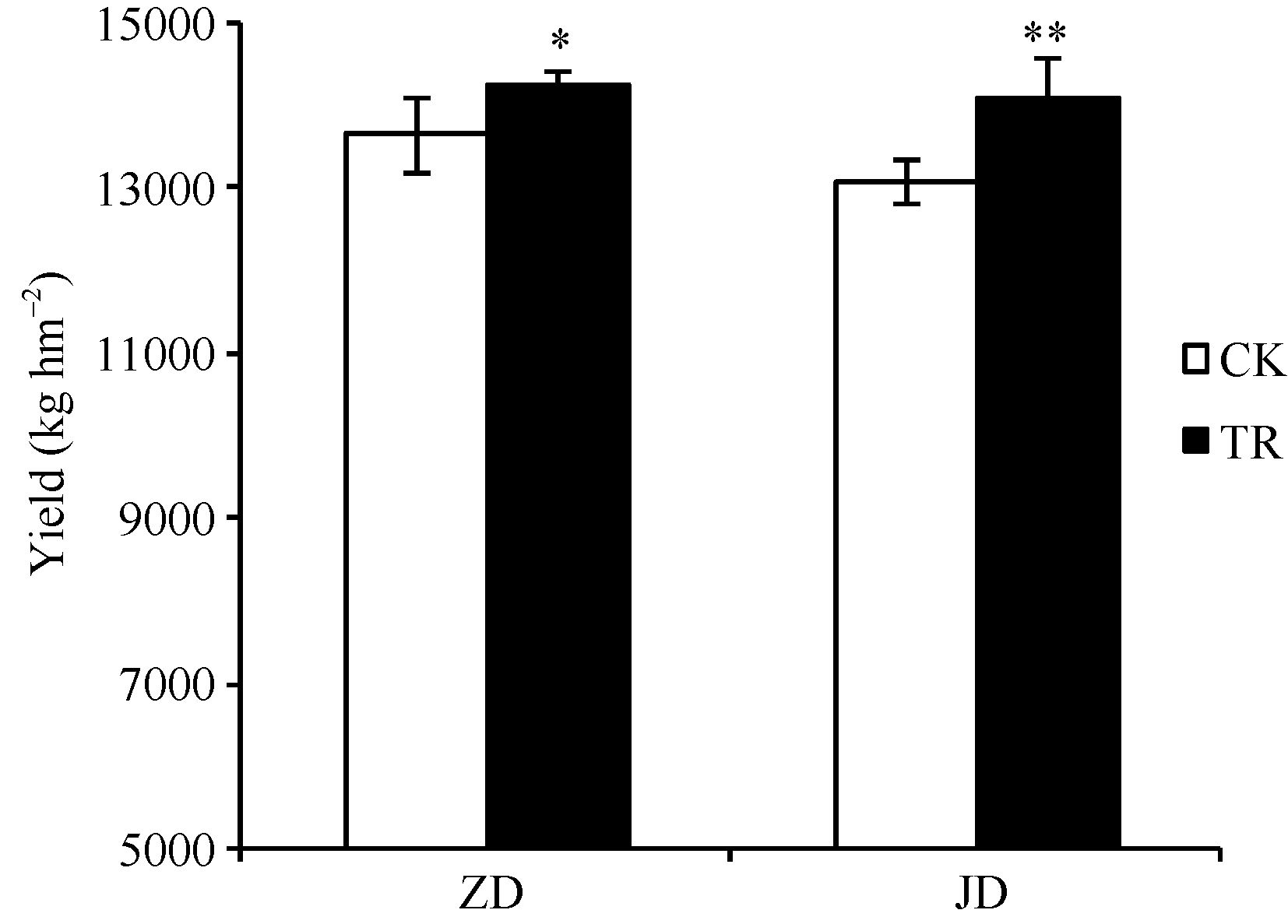
关键词:春玉米; 乙矮合剂; 聚糠萘合剂; 蔗糖; 蔗糖代谢关键酶 Effect of Plant Growth Regulators on Key Enzymes in Sucrose Metabolism of Ear Leaf and Grain at Filling Stage of Spring Maize LI Guang-Yan, WANG Qing-Yan, XU Yan-Li, LU Lin, Jiao Liu, DONG Xue-Rui, DONG Zhi-Qiang* Institute of Crop Science, Chinese Academy of Agricultural Sciences / Key Laboratory of Crop Ecophysiology and Cultivation, Beijing 100081, China Fund:This study was supported by the National Natural Science Foundation of China (31470087), the National Key Technology R&D Program of China (2012BAD04B01-07, 2012BAD4B02) and Science and Technology Innovation Project of Chinese Academy of Agricultural Sciences AbstractIn order to explore the new cultivation techniques, chemical control was established in this study. We conducted a field experiment using two varieties of Zhongdan 909 and Jidan 35 with treatments of leaf-spraying ethylene-chlormequat-potassium (ECK) in V6 and PASP-KT-NAA (PKN) in V9. The results showed that ECK & PKN increased the activities of sucrose synthetase (SS) and sucrose phosphate synthetase (SPS) and sucrose content during early grain filling stage (10-20 d after anthesis) in ear leaf, and decreased sucrose content in ear leaf during late grain filling stages (30-50 d after anthesis), especially it improved the ability of sucrose synthesis at the late filling stage (50 d after anthesis). ECK & PKN also decreased SS (synthesis) activity in grain during middle and late grain filling stages and increased SS (resolving) activity, promoting sucrose decomposition ability, with less effect on SS (synthesis) and SPS activities in grain. The yields of Zhongdan 909 and Jidan 35 compared with their own control were respectively increased by 4.61% and 7.78%. The study indicated that ECK & PKN enhance sucrose synthesis in leaf and decomposition ability in grain, and promote the translocation of carbon assimilation to grain, resulting in high grain-filling rate, high grain weight, and high yield.
Keyword:Spring maize; ECK; PKN; Sucrose; Key enzymes of sucrose metabolism Show Figures Show Figures
夏叔芳, 于新建, 张振清. 叶片光合产物输出的抑制与淀粉和蔗糖的积累. 植物生理学报, 1981, 7: 136-141Xia SF, Yu XJ, Zhang ZQ. Inhibition of photosynthetic product output and accumulation of starch and sucrose in leaves. J Plant Physiol, 1981, 7: 136-141 (in Chinese)[本文引用:1]
[2]
张明方, 李志凌. 高等植物中与蔗糖代谢相关的酶. 植物生理学报, 2002, 38: 289-295Zhang MF, Li ZL. Sucrose-metabolizing enzymes in higher plants. J Plant Physiol, 2002, 38: 289-295 (in Chinese)[本文引用:1]
[3]
Jang JC, SheenJ. Sugar sensing in higher plants. Plant Cell, 1997, 6: 1665-1679[本文引用:1]
[4]
赵福成, 景立权, 闫发宝, 陆大雷, 王桂跃, 陆卫平. 灌浆期高温胁迫对甜玉米籽粒糖分积累和蔗糖代谢相关酶活性的影响. 作物学报, 2013, 39: 1644-1651Zhao FC, Jing LQ, Yan FB, Lu DL, Wang GY, Lu WP. Effects of heat stress during grain filling on sugar accumulation and enzyme activity associated with sucrose metabolism in sweet corn. Acta Agron Sin, 2013, 39: 1644-1651 (in Chinese with English abstract)[本文引用:1]
[5]
覃鸿妮, 蔡一林, 孙海燕, 王久光, 王国强, 刘志斋. 种植密度对不同株型玉米蔗糖代谢和淀粉合成相关酶活性的影响. 中国生态农业学报, 2010, 18: 1183-1188Qin HN, Cai YL, Sun HY, Wang JG, Wang GQ, Liu ZZ. Effects of planting density on sucrose metabolism and activities of enzymes related to starch synthesis in maize hybrids with different plant types. Chin J Eco-Agric, 2010, 18: 1183-1188 (in Chinese with English abstract)[本文引用:3]
[6]
赵宏伟, 孔宇, 于秋竹, 杨亮. 密度对饲用玉米蔗糖合成和积累的影响. 玉米科学, 2009, 17(5): 72-76Zhao HW, KongY, Yu QZ, YangL. Effects of density on sucrose synthesize and accumulation of forage maize. J Maize Sci, 2009, 17(5): 72-76 (in Chinese with English abstract)[本文引用:3]
[7]
王俊忠, 黄高宝, 张超男, 杨亚军, 赵会杰, 朱晓燕, 马培芳. 施氮量对不同肥力水平下夏玉米碳氮代谢及氮素利用率的影响. 生态学报, 2009, 29: 2045-2052Wang JZ, Huang GB, Zhang CN, Yang YJ, Zhao HJ, Zhu XY, Ma PF. Influence of nitrogen fertilizer rate on carbon-nitrogen metabolism and nitrogen use efficiency of summer maize under high and medium yield levels. Acta Ecol Sin, 2009, 29: 2045-2052 (in Chinese with English abstract)[本文引用:2]
[8]
曹玉军, 赵宏伟, 王晓慧, 魏雯雯, 张磊, 王立春, 王永军. 施钾对甜玉米产量、品质及蔗糖代谢的影响. 植物营养与肥料学报, 2011, 29: 881-887Cao YJ, Zhao HW, Wang XH, Wei WW, ZhangL, Wang LC, Wang YJ. Effects of potassium fertilization on yield, quality and sucrose metabolism of sweet maize. Plant Nutr Fert Sci, 2011, 29: 881-887 (in Chinese with English abstract)[本文引用:2]
[9]
王庆燕, 管大海, 潘海波, 李建民, 段留生, 张明才, 李召虎. 油菜素内酯对春玉米灌浆期叶片光合功能与产量的调控效应. 作物学报, 2015, 41: 1557-1563Wang QY, Guan DH, Pan HB, Li JM, Duan LS, Zhang MC, Li ZH. Effect of brassinolide on leaf photosynthetic function and yield in spring maize filling stage. Acta Agron Sin, 2015, 41: 1557-1563 (in Chinese with English abstract)[本文引用:2]
[10]
刘鹏, 胡昌浩, 董树亭, 王空军, 张吉旺, 张保仁. 甜质型与普通型玉米籽粒发育过程中糖代谢相关酶活性的比较. 中国农业科学, 2005, 38: 52-58LiuP, Hu CH, Dong ST, Wang KJ, Zhang JW, Zhang BR. Comparison of enzymes activity associated with sucrose metabolism in the developing grains between sweet corn and normal corn. Sci Agric Sin, 2005, 38: 52-58 (in Chinese with English abstract)[本文引用:3]
[11]
董学会, 李建民, 何钟佩, 段留生, 李召虎. 30%己乙水剂对玉米叶片光合酶活性与同化物分配的影响. 玉米科学, 2006, 14(4): 93-96Dong XH, Li JM, He ZP, Duan LS, Li ZH. Effects of 30% Hex-ethyl-aqua on photosynthetic enzyme activities and as simulation distribution in maize plants. J Maize Sci, 2006, 14(4): 93-96 (in Chinese with English abstract)[本文引用:2]
[12]
卢霖, 董志强, 董学瑞, 焦浏, 李光彦, 高娇. 乙矮合剂对不同密度夏玉米花粒期叶片氮素同化与早衰的影响. 作物学报, 2015, 41: 1870-1879LuL, Dong ZQ, Dong XR, JiaoL, Li GY, GaoJ. Effects of Ethylene-Chlormequat-Potassium on leaf nitrogen assimilation after anthesis and early senescence under different planting densities. Acta Agron Sin, 2015, 41: 1870-1879 (in Chinese with English abstract)[本文引用:2]
[13]
徐田军, 董志强, 高娇, 陈传晓, 焦浏, 解振兴. 聚糠萘水剂对不同积温带玉米叶片衰老和籽粒灌浆速率的影响. 作物学报, 2012, 38: 1698-1709Xu TJ, Dong ZQ, GaoJ, Chen CX, JiaoL, Xie ZX. Effect of PASP-KT-NAA on leaf senescence and grain filling rate during the grain-filling period in different temperature zones. Acta Agron Sin, 2012, 38: 1698-1709 (in Chinese with English abstract)[本文引用:2]
[14]
高娇, 董志强, 徐田军, 陈传晓, 焦浏, 卢霖, 董学瑞. 聚糠萘水剂对不同积温带玉米花后叶片氮同化的影响. 生态学报, 2014, 34: 2938-2947GaoJ, Dong ZQ, Xu TJ, Chen CX, JiaoL, LuL, Dong XR. Effects of PASP-KT-NAA on maize leaf nitrogen assimilation after florescence over different temperature gradients. Acta Ecol Sin, 2014, 34: 2938-2947 (in Chinese with English abstract)[本文引用:1]
[15]
陈传晓, 董志强, 高娇, 徐田军, 焦浏, 卢霖, 张凤路. 聚糠萘水剂对不同积温带春玉米灌浆期光合性能的影响. 玉米科学, 2013, 21(3): 66-70Chen CX, Dong ZQ, GaoJ, Xu TJ, JiaoL, LuL, Zhang FL. Effects of PASP-KT-NAA on the photosynthetic performances of different maize cultivars in different accumulated temperature zones. J Maize Sci, 2013, 21(3): 66-70 (in Chinese with English abstract)[本文引用:1]
[16]
赵智中, 张上隆, 徐昌杰, 陈昆松, 刘拴桃. 蔗糖代谢相关酶在温州蜜柑果实糖积累中的作用. 园艺学报, 2001, 28: 112-118Zhao ZZ, Zhang SL, Xu CJ, Chen KS, Liu ST. The role of sucrose metabolizing enzymes in accumulation of sugars in Satsuma mand arin. Acta Hortic Sin, 2001, 28: 112-118 (in Chinese)[本文引用:1]
[17]
陈传永, 王荣焕, 赵久然, 徐田军, 王元东, 刘秀芝, 刘春阁, 裴志超, 成广雷, 陈国平. 不同生育时期遮光对玉米籽粒灌浆特性及产量的影响. 作物学报, 2014, 40: 1650-1657Chen CY, Wang RH, Zhao JR, Xu TJ, Wang YD, Liu XZ, Liu CG, Pei ZC, Cheng GL, Chen GP. Effects of shading on grain-filling properties and yield of maize at different growth stages. Acta Agron Sin, 2014, 40: 1650-1657 (in Chinese with English abstract)[本文引用:1]
[18]
Gottwald JR, Krysan PJ, Young JC, Evert RF, Sussman MR. Genetic evidence for the in planta role of phloem-specific plasma membrane sucrose transporters. Proc Natl Acad Sci USA, 2000, 97: 13979-13984[本文引用:1]
[19]
FarrarJ, PollockC, GallagherJ. Sucrose and the integration of metabolism in vascular plants. Plant Sci, 2000, 154: 1-11[本文引用:1]
[20]
陈传晓. 不同积温带春玉米碳代谢机理及化学调控效应的研究. 河北农业大学硕士学位论文, 河北石家庄, 2013Chen CX. Studies on the Mechanism of Carbon Metabolism of Spring Maize and Chemical Regulation Effects under Different Accumulated Temperature Zones. PhD Dissertation of Agricultural University of Hebei, Shijiazhuang, China, 2013 (in Chinese with English abstract)[本文引用:1]
[21]
王文静. 不同穗型冬小麦籽粒灌浆期源库强度及其与淀粉积累的关系. 作物学报, 2004, 30: 916-921Wang WJ. The relationship between source-sink intensity and starch accumulation during grain filling period in two winter wheat cultivars with different spike types. Acta Agron Sin, 2004, 30: 916-921 (in Chinese with English abstract)[本文引用:1]
[22]
EschrichW, EschrichB. Control of phloem unloading by source activities and light. Plant Physiol Biochem, 1992, 25: 625-634[本文引用:1]
[23]
Dickinson CD, AltabellaT, Chrispeels MJ. Slow-growth phenotype of transgenic tomoto expressing apoplastic invertase. Plant Physiol, 1991, 95: 420-425[本文引用:1]
[24]
WeberH, BorisjukL, WobusU. Sugar import and metabolism during seed development. Trends Plant Sci, 1997, 2: 169-174[本文引用:1]
[25]
Cheng WH, Chourey PS. Sucrose phosphate synthase expression at the cell and tissue level is coordinated with sucrose sink-to-source transitions in maize leaf. Plant Physiol, 1996, 111: 1021-1029[本文引用:1]
[26]
张海艳, 董树亭, 高荣岐, 李玉全. 玉米籽粒淀粉积累及相关酶活性分析. 中国农业科学, 2008, 41: 2174-2181Zhang HY, Dong ST, Gao RQ, Li YQ. Starch accumulation and enzymes activities associated with starch synthesis in maize kernels. Sci Agric Sin, 2008, 41: 2174-2181 (in Chinese with English abstract)[本文引用:1]
[27]
Hubbard NL, Huber SC, Pharr DM. Sucrose phosphate synthase and acid invertase as determinants of sucrose concentration in developing muskmelon (Cucumis melo L. ) fruits. Plant Physiol, 1989, 91: 1527-1534[本文引用:1]
[28]
张超男, 赵会杰, 王俊忠, 付晓记. 不同施肥方式对夏玉米碳水化合物代谢关键酶活性的影响. 植物营养与肥料学报, 2008, 14: 54-58Zhang CN, Zhao HJ, Wang JZ, Fu XJ. Effect of different fertilization methods on activities of key enzymes in carbohydrate metabolism of summer maize. Plant Nutr Fert Sci, 2008, 14: 54-58 (in Chinese with English abstract)[本文引用:1]
[29]
陈洋, 赵宏伟. 氮素用量对春玉米穗位叶蔗糖合成关键酶活性的影响. 玉米科学, 2008, 16(1): 115-118ChenY, Zhao HW. Effect of nitrogen application on activities of key enzymes of sucrose synthesis in the leaf located near the ear of spring maize. J Maize Sci, 2008, 16(1): 115-118 (in Chinese with English abstract[本文引用:1]
[30]
李合生. 现代植物生理学(第2版). 北京: 高等教育出版社, 2006. pp 207-208Li H S. Modern Plant Physiology, 2nd edn. Beijing: Higher Education Press, 2006. pp 207-208(in Chinese)[本文引用:1]
[31]
BorkovecV, ProchazkaS. Pre-anthesis interaction of cytokinins and ABA in the transport of 14C-sucrose to the ear of winter wheat (Triticum aestivum L. ). J Agron Crop Sci, 1992, 169: 229-235[本文引用:1]
 , 王庆燕, 许艳丽, 卢霖, 焦浏, 董学瑞, 董志强
, 王庆燕, 许艳丽, 卢霖, 焦浏, 董学瑞, 董志强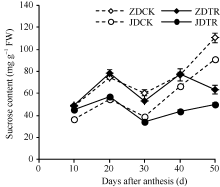





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}