关键词:水稻; 高温胁迫; 颖花退化; 花药发育; 籽粒充实 Effect of Heat Stress on Spikelet Degeneration and Grain Filling at Panicle Initiation Period of Rice WANG Ya-Liang**, ZHANG Yu-Ping**, ZHU De-Feng*, XIANG Jing, WU Hui, CHEN Hui-Zhe, ZHANG Yi-Kai China National Rice Research Institute / State Key Laboratory of Rice Biology, Hangzhou 310006, China Fund:This study was supported by Special Fund for Agro-Scientific Research in the Public Interest (201203029) and the China Agriculture Research System (CARS-01-26) AbstractPot experiment was conducted to investigate the effects of heat stress on rice spikelets degeneration and grain filling at panicle initiation stage. A tolerant (Huanghuazhan) and a susceptible (Fengliangyou 6) cultivars were treated in phytotron with high temperature (10:00-15:00, 40℃) and normal temperature (10:00-15:00, 32℃) for one, three, five, seven, and nine days at last leaf emergence stage, with these grown under natural condition as control. The results indicated that the ratio of spikelet degeneration increased greatly with increasing heat-treating time. The ratio of spikelet degeneration of Huanghuazhan increased 45.8% and 62.9% after nine days under high temperature as compared with normal temperature treatment (control), and the ratio of Fengliangyou6 increased 81.7% and 136.1% in the same treatment, showing higher ratio of spikelet degeneration in Fengliangyou6 than Huanghuazhan. The pollen activity significantly decreased after one day under high temperature stress, and anther formation significantly inhibited after five days under high temperature stress. Bad anther formation and poor pollen activity under heat stress resulting in seed-setting rate significantly decreased. Spikelet length and grain filling of two cultivars decreased due to heat stress effect, resulting in a significant reduction of grain weight, with a larger reduction rate in Fengliangyou 6 than in Huanghuazhan. Grain filling reduced significantly after five days of high temperature treatment. According to the study, heat stress at last leaf emergence stage may sharply decrease rice yield, and corresponding cultivation measures should be strengthened in production.
Keyword:Rice; Heat stress; Spikelet degeneration; Anther formation; Grain filling Show Figures Show Figures
表1 高温处理对水稻颖花退化和结实率的影响 Table 1 Effect of high temperature on rice spikelets degeneration and seed-setting rate
处理 Treatment temperature
处理天数 Treatment days
黄华占 Huanghuazhan
丰两优6号 Fengliangyou 6
穗粒数 Number of spikelets per panicle (Nr.)
退化率 Ratio of spikelets degeneration (%)
结实率 Seed-setting rate (%)
穗粒数 Number of spikelets per panicle
退化率 Ratio of spikelets degeneration (%)
结实率 Seed-setting rate (%)
对照CK
255.8 ± 6.1 a
36.8 ± 0.9 ef
88.2 ± 5.4 a
266.8 ± 4.5 a
24.1 ± 2.7 cd
85.6 ± 4.1 ab
HT
1 d
251.5 ± 18.8 a
31.8 ± 3.4 g
92.5 ± 8.3 a
257.6 ± 18.2 ab
19.8 ± 1.1 d
79.7 ± 5.4 b
3 d
227.5 ± 7.3 bcd
43.0 ± 1.9 c
75.7 ± 8.6 b
254.7 ± 7.4 ab
26.5 ± 4.7 cd
66.7 ± 8.3 c
5 d
191.1 ± 4.4 ef
50.1 ± 3.4 b
52.0 ± 7.3 c
196.3 ± 30.9 c
28.4 ± 2.0 cd
29.8 ± 8.6 d
7 d
178.9 ± 18.0 f
58.9 ± 4.0 a
12.7 ± 2.2 d
149.2 ± 8.6 d
48.9 ± 4.4 b
10.0 ± 7.3 e
9 d
174.9 ± 9.8 f
59.6 ± 3.4 a
4.4 ± 4.8 d
118.3 ± 13.3 e
56.9 ± 8.2 a
2.5 ± 2.2 e
NT
1 d
247.6 ± 2.9 abc
36.5 ± 0.8 ef
83.6 ± 3.8 ab
250.5 ± 25.9 ab
26.4 ± 4.2 cd
86.2 ± 0.4 ab
3 d
241.1 ± 14.3 abc
36.1 ± 3.3 f
92.3 ± 3.4 a
247.3 ± 22.8 ab
26.1 ± 1.4 cd
90.2 ± 3.4 a
5 d
226.3 ± 12.6 bcd
38.2 ± 2.3 def
88.7 ± 1.0 a
234.9 ± 17.3 ab
28.5 ± 2.6 c
92.1 ± 1.1 a
7 d
212.3 ± 8.5 de
42.4 ± 1.8 cd
91.5 ± 3.2 a
228.0 ± 6.8 b
29.1 ± 2.6 c
94.5 ± 3.2 a
9 d
228.5 ± 25.8 bcd
40.9 ± 1.2 cde
91.9 ± 1.1 a
225.9 ± 9.4 bc
31.3 ± 5.5 c
93.3 ± 1.2 a
CK is the treatment under natural condition; HT is the treatment in high temperature phytotron; NT is the treatment in normal temperature phytotron. Values followed by different small letters within the same column are significantly different at the 0.05 probability level among treatments. 对照为自然环境条件下的处理; HT表示高温人工气候箱处理; NT表示适温人工气候箱处理; 同列不同小写字母表示处理间差异显著(P< 0.05)。
表1 高温处理对水稻颖花退化和结实率的影响 Table 1 Effect of high temperature on rice spikelets degeneration and seed-setting rate
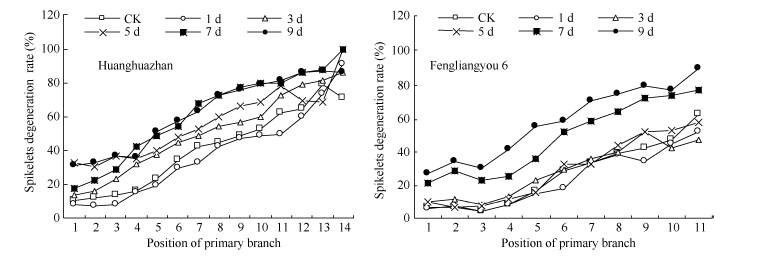
图1 高温处理对不同部位枝梗上的退化率的影响 对照为自然环境条件下的处理; 图中稻穗一次枝梗部位顺序依次从稻穗顶部到基部。Fig. 1 Effect of high temperature on degenerated spikelet ratio of different primary branches CK is the treatment under natural condition; Order of primary branches is from panicle top to bottom.
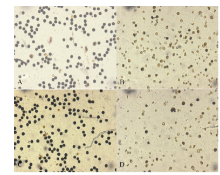
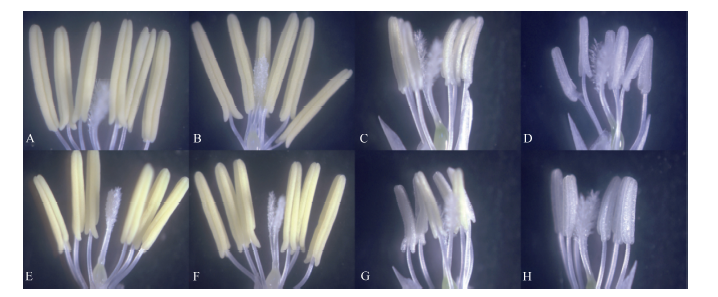
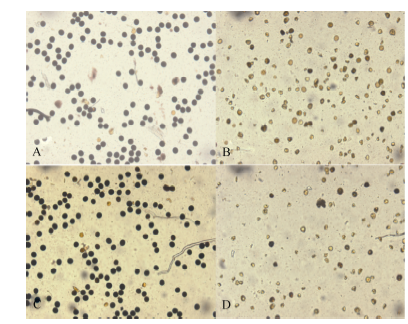
图2 高温处理花药性状的分类 A~D: 黄华占品种的花药; E~H: 丰两优6号的花药; A, E: 对照; B, F: 适温处理; C~D, G~H: 高温处理; A、B和E、F是正常的花药; C和G是部分充实花药; D和H是不充实的花药。Fig. 2 Classification of anther characters under high temperature A-D: Huanghuazhan’ s anther; E-H: Fengliangyou 6’ s anther; A and E: CK; B and F: normal temperature treatment; C-D and G-H: high temperature treatment; A-B and E-F: normal anther; C and G: partially filled anther; D and H: non-filled anther.
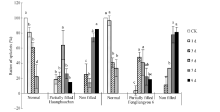
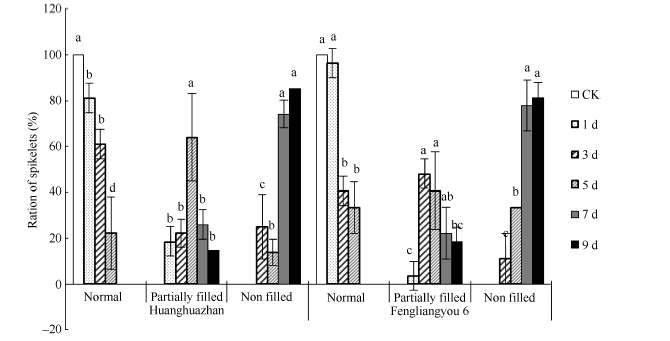
图3 高温处理后不同类型花药颖花比例 对照(CK)为自然环境条件下的处理; 不同小写字母表示处理间差异显著(P< 0.05)。Fig. 3 Ratio of spikelets with different anther characters under high temperature CK is the treatment under natural condition. Bars superscripted by different small letters are significantly different at the 0.05 probability level among treatments.
表2 高温处理对花药大小的影响 Table 2 Effect of high temperature on the size of anther (mm)
处理天数 Treatment days
黄华占Huanghuzhan
丰两优6号 Fengliangyou 6
长 Length
宽 Width
长 Length
宽 Width
对照CK
2.25 ± 0.15 a
0.43 ± 0.02 a
2.10 ± 0.10 b
0.38 ± 0.03 b
1 d
2.20 ± 0.06 a
0.43 ± 0.01 a
2.40 ± 0.05 a
0.42 ± 0.02 a
3 d
2.27 ± 0.04 a
0.43 ± 0.01 a
1.84 ± 0.25 c
0.34 ± 0.02 c
5 d
1.74 ± 0.17 b
0.35 ± 0.02 b
1.82 ± 0.04 c
0.33 ± 0.00 c
7 d
1.59 ± 0.08 b
0.33 ± 0.02 bc
1.54 ± 0.11 d
0.31 ± 0.01 c
9 d
1.39 ± 0.03 c
0.30 ± 0.02 c
1.58 ± 0.12 d
0.31 ± 0.01 c
CK is the treatment under natural condition; Values followed by different small letters within the same column are significantly different at 0.05 probability level among treatments. 对照为自然环境条件下的处理; 同列不同小写字母表示处理间差异显著(P< 0.05)。
表2 高温处理对花药大小的影响 Table 2 Effect of high temperature on the size of anther (mm)
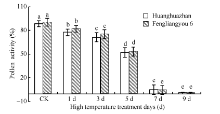
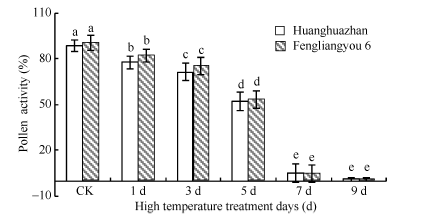
图4 高温处理对花粉活力的影响 对照为自然环境条件下的处理; 不同小写字母表示处理间差异显著(P< 0.05)。Fig. 4 Effect of high temperature on pollen viability CK is the treatment under natural condition; Bars superscripted by different small letters are significantly different at the 0.05 probability level among treatments.
图5 不同花药的花粉性状比较 A: 黄华占正常花药的花粉; B: 黄华占充实性差花药的花粉; C: 丰两优6号正常花药的花粉; D: 丰两优6号充实性差花药的花粉。Fig. 5 Comparison of pollen character in different anthers A: pollen of normal anther in Huanghuazhan; B: pollen of partially filled anther in Huanghuazhan; C: pollen of normal anther in Fengliangyou 6; D: pollen of partially filled anther in Fengliangyou 6.
表3 高温处理对颖花大小的影响 Table 3 Effect of high temperature on size of spikelets (mm)
处理温度 Treatment temperature
处理天数 Treatment days
黄华占Huanghuazhan
丰两优6号 Fengliangyou 6
长 Length
宽 Width
长 Length
宽 Width
对照CK
8.7 ± 0.1 a
2.1 ± 0.0 b
9.0 ± 0.1 a
2.5 ± 0.0 b
HT
1 d
8.1 ± 0.1 cde
2.1 ± 0.0 b
8.6 ± 0.1 b
2.4 ± 0.0 b
3 d
8.0 ± 0.1 e
2.1 ± 0.0 b
8.1 ± 0.1 c
2.4 ± 0.0 b
5 d
8.1 ± 0.1 de
2.2 ± 0.0 a
7.9 ± 0.1 d
2.5 ± 0.1 b
7 d
8.0 ± 0.2 b
2.2 ± 0.0 a
7.5 ± 0.1 e
2.5 ± 0.1 b
9 d
7.8 ± 0.2 f
2.1 ± 0.1 b
6.8 ± 0.2 f
2.6 ± 0.1 a
NT
1 d
8.3 ± 0.1 bc
2.1 ± 0.0 b
8.9 ± 0.0 a
2.5 ± 0.0 b
3 d
8.2 ± 0.1 bcd
2.1 ± 0.0 b
8.6 ± 0.1 b
2.5 ± 0.0 b
5 d
8.4 ± 0.0 b
2.2 ± 0.0 a
8.7 ± 0.1 b
2.6 ± 0.1 a
7 d
8.1 ± 0.1 de
2.2 ± 0.0 a
8.6 ± 0.1 b
2.6 ± 0.1 a
9 d
8.6 ± 0.0 a
2.2 ± 0.0 a
8.7 ± 0.1 b
2.6 ± 0.1 a
CK is the treatment under natural condition; HT is the treatment in high temperature phytotron; NT is the treatment in normal temperature phytotron; Values followed by different small letters within the same column are significantly different at the 0.05 probability level among treatments. 对照为自然环境条件下的处理; HT表示高温人工气候箱处理; NT表示适温人工气候箱处理; 同列不同小写字母表示处理间差异显著(P< 0.05)。
表3 高温处理对颖花大小的影响 Table 3 Effect of high temperature on size of spikelets (mm)
表4 高温处理对比粒重和千粒重的影响 Table 4 Effect of high temperature on ratio of grain weight to area and 1000-grain weight
处理温度 Treatment temperature
处理天数 Treatment days
黄华占 Huanghuazhan
丰两优6号 Fengliangyou 6
比粒重 Ratio of grain weight to area (mg mm-2)
千粒重 1000-grain weight (g)
比粒重 Ratio of grain weight to area (mg mm-2)
千粒重 1000-grain weight (g)
对照CK
1.4 ± 0.0 a
19.4 ± 0.5 ab
1.6 ± 0.0 a
26.8 ± 0.7 ab
HT
1 d
1.3 ± 0.0 b
18.6 ± 0.4 c
1.5 ± 0.0 ab
24.8 ± 0.5 c
3 d
1.4 ± 0.0 a
18.1 ± 0.2 c
1.5 ± 0.0 ab
22.9 ± 0.3 d
5 d
1.2 ± 0.0 c
17.1 ± 0.3 d
1.4 ± 0.0 b
21.9 ± 0.4 e
7 d
1.2 ± 0.0 c
15.7 ± 0.8 e
1.3 ± 0.0 c
18.1 ± 0.9 f
9 d
1.2 ± 0.0 c
16.2 ± 0.6 e
1.2 ± 0.1 d
15.3 ± 1.5 g
NT
1 d
1.4 ± 0.0 a
19.4 ± 0.3 b
1.6 ± 0.0 a
27.1 ± 8.9 a
3 d
1.4 ± 0.0 a
19.7 ± 0.3 ab
1.5 ± 0.1 ab
26.4 ± 8.6 b
5 d
1.4 ± 0.0 a
19.6 ± 0.3 ab
1.6 ± 0.0 a
27.4 ± 8.7 b
7 d
1.4 ± 0.0 a
20.1 ± 0.9 a
1.5 ± 0.0 ab
26.1 ± 8.6 b
9 d
1.4 ± 0.0 a
19.9 ± 0.4 ab
1.5 ± 0.0 ab
26.9 ± 8.7 b
CK is the treatment under natural condition; HT is the treatment in high temperature phytotron; NT is the treatment in normal temperature phytotron; Values followed by different small letters within the same column are significantly different at the 0.05 probability level among treatments. 对照为自然环境条件下的处理; HT表示高温人工气候箱处理; NT表示适温人工气候箱处理; 同列不同小写字母表示处理间差异显著(P< 0.05)。
表4 高温处理对比粒重和千粒重的影响 Table 4 Effect of high temperature on ratio of grain weight to area and 1000-grain weight
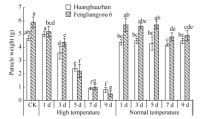
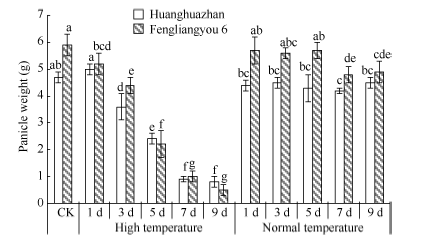
图6 高温处理对单穗重的影响 对照(CK)为自然环境条件下的处理; 不同小写字母表示处理间差异显著(P< 0.05)。Fig. 6 Effect of high temperature on rice panicle weight CK is the treatment under natural condition; Bars superscripted by different small letters are significantly difference at the 0.05 probability level among treatments.
表5 Table 5 表5(Table 5)
表5 高温处理穗部性状的相关性分析 Table 5 Correlation coefficients of panicle traits under high temperature
品种 Cultivar
指标 Index
处理天数 Treatment days (d)
穗重 Panicle weight (g)
穗粒数 Number of spikelets
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
黄华占 Huanghuanzhan
处理天数 Treatment days
1.00
穗重 Panicle weight (g)
-0.98* *
1.00
穗粒数 Number of spikelets
-0.99* *
0.98* *
1.00
结实率 Seed-setting rate (%)
-0.97* *
0.98* *
0.97* *
1.00
千粒重 1000-grain weight (g)
-0.95* *
0.97* *
0.94* *
0.95* *
1.00
丰两优6号 Fengliangyou 6
处理天数Treatment days
1.00
穗重 Panicle weight (g)
-0.98* *
1.00
穗粒数 Number of spikelets
-0.97* *
0.97*
1.00
结实率 Seed-setting rate (%)
-0.98* *
0.99* *
0.98* *
1.00
千粒重 1000-grain weight (g)
-0.98* *
0.96* *
0.96* *
0.95* *
1.00
* P< 0.05, * * P< 0.01.
表5 高温处理穗部性状的相关性分析 Table 5 Correlation coefficients of panicle traits under high temperature
李勇, 杨晓光, 叶清, 陈阜. 全球气候变暖对中国种植制度可能影响: IX. 长江中下游地区单双季稻高低温灾害风险及其产量影响. 中国农业科学, 2013, 46: 3997-4006LiY, Yang XG, YeQ, ChenF. The possible effects of global warming on cropping systems in China: IX. The risk of high and low temperature disasters for single and double rice and its impacts on rice yield in the Middle-lower Yangtze plain. Sci Agric Sin, 2013, 46: 3997-4006 (in Chinese with English abstract)[本文引用:1]
[2]
谢晓金, 李秉柏, 王琳, 戴秦如, 申双和. 长江中下游地区高温时空分布及水稻花期的避害对策. 中国农业气象, 2010, 31: 144-150Xie XJ, Li BB, WangL, Dai QR, Shen SH. Spatial and temporal distribution of high temperature and strategists rice florescence harm in the lower-middle reaches of Yangtze River. Chin J Agrometeorol, 2010, 31: 144-150 (in Chinese with English abstract)[本文引用:1]
[3]
谢志清, 杜银, 高苹, 曾燕. 江淮流域水稻高温热害灾损变化及应对策略. 气象, 2013, 39: 774-784Xie ZQ, DuY, GaoP, ZengY. Impact of high-temperature on single cropping rice over Yangtze-Huaihe river valley and response measures. Meteorol Monthly, 2013, 39: 774-784 (in Chinese with English abstract)[本文引用:1]
[4]
IPCC. Climate Change 2013: the Physical Science Basis. Oslo: Inter Government Panel on Climate Change, 2013[本文引用:1]
[5]
DasS, KrishnanP, NayakM, RamakrishnanB. High temperature stress effects on pollens of rice (Oryza sativa L. ) genotypes. Environ Exp Bot, 2014, 101: 36-46[本文引用:2]
[6]
Rang ZW, Jagadish S V K, Zhou Q M, Craufurd P Q, Heuer S. Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ Exp Bot, 2011, 70: 58-65[本文引用:2]
[7]
张桂莲, 张顺堂, 肖浪涛, 唐文帮, 肖应辉, 陈立云. 抽穗开花期高温胁迫对水稻花药、花粉粒及柱头生理特性的影响. 中国水稻科学, 2014, 28: 155-166Zhang GL, Zhang ST, Xiao LT, Tang WB, Xiao YH, Chen LY. Effect of high temperature stress on physiological characteristics of anther, pollen and stigma of rice during heading-flowering stage. Chin J Rice Sci, 2014, 28: 155-166 (in Chinese with English abstract)[本文引用:2]
[8]
Shrivastava PR, SaxenaP, Xalxo MS, Verulkar SB. Effect of high temperature at different growth stages on rice yield and grain quality traits. J Rice Res, 2012, 5: 29-42[本文引用:3]
[9]
Prasad P VV, Boote KJ, Allen LH. Adverse high temperature effects on pollen viability, seed-set, seed yield and harvest index of grain-sorghum [Sorghum bicolor (L. ) Moench] are more severe at elevated carbon dioxide due to higher tissue temperatures. Agric For Meteorol, 2006, 139: 237-251[本文引用:2]
[10]
唐恬, 金荣花, 彭相瑜, 牛若芸. 2013年夏季我国南方区域性高温天气的极端性分析. 气象, 2014, 40: 1207-1215TangT, Jin RH, Peng XY, Niu NY. Analysis on extremely high temperature over southern china in summer 2013. Meteorol Monthly, 2014, 40: 1207-1215 (in Chinese with English abstract)[本文引用:1]
[11]
邓运, 田小海, 吴晨阳, 松井勤, 肖波. 热害胁迫条件下水稻花药发育异常的早期特征. 中国生态农业学报, 2010, 18: 377-383DengY, Tian XH, Wu CY, MatsuT, XiaoB. Early signs of heat stress-induced abnormal development of anther in rice. Chin J Eco-agric, 2010, 18: 377-383 (in Chinese with English abstract)[本文引用:2]
[12]
曹云英, 段骅, 杨立年, 王志琴, 周少川, 杨建昌. 减数分裂期高温胁迫对耐热性不同水稻品种产量的影响及其生理原因. 作物学报, 2008, 34: 2134-2142Cao YY, DuanH, Yang LN, Wang ZQ, Zhou SC, Yang JC. Effect of heat-stress meiosis on grain yield of rice cultivars differing in heat-tolerance and its physiological mechanism. Acta Agron Sin, 2008, 34: 2134-2142 (in Chinese with English abstract)[本文引用:2]
[13]
AbikoM, AkibayashiK, SakataT, KimuraM, KiharaM, ItohK, AsamizuE, SatoS, TakahashiH, HigashitaniA. High-temperature induction of male sterility during barley (Hordeum vulgare L. ) anther development is mediated by transcriptional inhibition. Sex Plant Reprod, 2005, 18: 91-100[本文引用:1]
[14]
OshinoT, AbikoM, SaitoR, IchiishiE, EndoM, Kawagishi- KobayashiM, HigashitaniA. Premature progression of anther early developmental programs accompanied by comprehensive alterations in transcription during high-temperature injury in barley plants. Mol Genet Genomics, 2007, 278: 31-42[本文引用:1]
[15]
王亚梁, 张玉屏, 曾研华, 武辉, 向镜, 陈惠哲, 张义凯, 朱德峰. 水稻穗分化期高温对颖花分化及退化的影响. 中国农业气象, 2015, 36: 724-731Wang YL, Zhang YP, Zeng YH, WuH, XiangX, Chen HZ, Zhang YK, Zhu DF. Effect of high temperature stress on rice spikelet differentiation and degeneration during panicle initiation stage. Chin J Agrometeorol, 2015, 36: 724-731 (in Chinese with English abstract)[本文引用:1]
[16]
杨弘远. 水稻生殖生物学. 杭州: 浙江大学出版社, 2005. pp 22-23Yang H Y. Rice Reproduction Biology. Hangzhou: Zhejiang University Press, 2005. pp 22-23(in Chinese)[本文引用:1]
柳新伟, 孟亚利, 周治国, 曹卫星. 水稻颖花分化与退化的动态特征. 作物学报, 2005, 31: 451-455Liu XW, Meng YL, Zhou ZG, Cao WX. Dynamic characteristic of floret differentiation and degeneration in rice. Acta Agron Sin, 2005, 31: 451-455 (in Chinese with English abstract)[本文引用:1]
[19]
杨洪建, 杨连新, 黄建晔, 刘红江, 董桂春, 颜士敏, 朱建国, 王余龙. FACE对武香粳14颖花分化与退化的影响. 作物学报, 2006, 32: 1076-1082Yang HJ, Yang LX, Huang JY, Liu HJ, Dong GC, Yan SM, Zhu JG, Wang YL. Effect of free-air CO2 enrichment on spikelet differentiation and degeneration of japonica rice (Oryza sativa L. ) cultivar Wuxiangjing 14. Acta Agron Sin, 2006, 32: 1076-1082 (in Chinese with English abstract)[本文引用:1]
[20]
戚华雄, 杜雪树, 李进波. 水稻颖花退化的遗传研究进展. 湖北农业科学, 2014, 53: 5905-5907Qi HX, Du XS, Li JB. Genetic studies of floret degeneration in rice (Oryza sativa L. ). Hubei Agric Sci, 2014, 53: 5905-5097 (in Chinese with English abstract)[本文引用:1]
[21]
魏育明, 郑有良. 内源激素对小麦可育小花数的调控. 四川农业大学学报, 1998, 16: 289-293Wei YM, Zheng YL. The regulation of phytohormone in determination of fertile floret number in wheat. J Sichuan Agric Univ, 1998, 16: 289-293 (in Chinese with English abstract)[本文引用:1]
[22]
杨建昌, 刘凯, 张慎凤, 王学明, 王志琴, 刘立军. 水稻减数分裂期颖花中激素对水分胁迫的响应. 作物学报, 2008, 34: 111-118Yang JC, LiuK, Zhang SF, Wang XM, Wang ZQ, Liu LJ. Hormones in rice spikelets in responses to water stress during meiosis. Acta Agron Sin, 2008, 34: 111-118 (in Chinese with English abstract)[本文引用:1]
[23]
Prasad P VV, Boote KJ, Allen LH, Sheehy JE, Thomas J MG. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res, 2006, 95: 398-411[本文引用:1]
[24]
Mohammed AR, TarpleyL. High nighttime temperatures affect rice productivity through altered pollen germination and spikelet fertility. Agric For Meteorol, 2009, 149: 999-1008[本文引用:1]
[25]
MatsuiT, OmasaK, HorieT. High temperature at flowering inhibits swelling of pollen grains, a driving force thecae dehiscence in rice. Plant Prod Sci, 2000, 3: 430-434[本文引用:1]
[26]
霍中洋, 叶全宝, 李华, 张洪程, 戴其根, 许轲. 水稻源库关系研究进展. 中国农学通报, 2002, 18(6): 72-77Huo ZY, Ye QB, LiH, Zhang HC, Dai QG, XuK. Advances of research on source-sink relationship of rice. Chin Agric Sci Bull, 2002, 18(6): 72-77 (in Chinese with English abstract)[本文引用:1]
[27]
陈惠哲. 水稻物质运转规律及其产量形成的研究. 中国农业科学院硕士学位论文, 北京, 2007Chen HZ. Study on Matter Translation Characteristic and Yield Formation in Rice (Oryza sativa L. ). MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2007 (in Chinese with English abstract)[本文引用:1]
[28]
张卫星. 超级稻品种穗生长发育和灌浆结实对水分亏缺的响应及其机理. 中国农业科学院博士学位论文, 北京, 2008Zhang WX. Study on Response of Panicle Growth and Development, Grain Filling under Water Deficit and Its Mechanism in Super Rice (Oryza sativa L. ) Varieties. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2008 (in Chinese with English abstract)[本文引用:1]
[29]
郑建初, 张彬, 陈留根, 杜群, 秦永生, 宋健, 张卫建. 抽穗期高温对水稻产量构成要素和稻米品质的影响及其基因型差异. 江苏农业科学, 2005, 21: 249-254Zheng JC, ZhangB, Chen LG, DuQ, Qin YS, SongJ, Zhang WJ. Genotypic differences in effects of high air temperature in field on rice yield component and grain quality during heading stage. Jiangsu J Agric Sci, 2005, 21: 249-254 (in Chinese with English abstract)[本文引用:1]
[30]
王亚梁, 张玉屏, 朱德峰, 向镜, 陈惠哲, 张义凯. 水稻器官形态和干物质积累对穗分化不同时期高温的响应. 中国水稻科学, 2016, 30: 161-169Wang YL, Zhang YP, Zhu DF, XiangJ, Chen HZ, Zhang YK. Response of rice organ morphology and dry matter accumulation to high temperature at different panicle initiation stages. Chin J Rice Sci, 2006, 30: 161-169 (in Chinese with English abstract)[本文引用:1]
[31]
上海植物生理研究所人工气候室. 高温对早稻开花结实的影响及其防治: II. 早稻开花期高温对开花结实的影响. 植物学报, 1976, 18: 323-329Laboratory ofPhytotron, Shanghai Institute of Plant Physiology. The influence of high temperature on the flowering and fruiting of high temperature on the flowering and fruiting of early rice and its control: II. The influence of high temperature on the flowering-fruiting of early rice during the flowering stage. Acta Bot Sin, 1976, 18: 323-329 (in Chinese with English abstract)[本文引用:1]
[32]
熊洪, 冉茂林, 徐富贤, 洪松. 南方稻区再生稻研究进展及发展. 作物学报, 2000, 26: 297-304XiongH, Ran ML, Xu FX, HongS. Achievements and developments of rationing rice in south of China. Acta Agron Sin, 2000, 26: 297-304 (in Chinese with English abstract)[本文引用:1]
[33]
张彬, 郑建初, 黄山, 田云录, 彭兰, 卞新民, 张卫建. 抽穗期不同灌水深度下水稻群体与大气的温度差异. 应用生态学报, 2008, 19: 87-92ZhangB, Zheng JC, HuangS, Tian YL, PengL, Bian XM, Zhang WJ. Temperature differences of air-rice plant under different irrigated water depths at spiking stage. Chin J Appl Ecol, 2008, 19: 87-92 (in Chinese with English abstract)[本文引用:1]
[34]
张彩霞. 水杨酸减轻水稻高温伤害的机理研究. 中国农业科学院硕士学位论文, 北京, 2015Zhang CX. The Mechanism of Salicylic Acid in Alleviating Damage on the Rice Induced by Heat Stress. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2015 (in Chinese with English abstract)[本文引用:1]
 , 向镜, 武辉, 陈惠哲, 张义凯
, 向镜, 武辉, 陈惠哲, 张义凯


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}