关键词:棉花(Gossypium hirsutum L.); 花铃期; 渍水; 叶片蔗糖代谢; 铃重 Effects of Waterlogging on Sucrose Metabolism of the Subtending Leaf of Cotton Boll and Boll Weight during Flowering and Boll-forming Stage YANG Chang-Qin1,2, LIU Rui-Xian2, ZHANG Guo-Wei2, XU Li-Hua2, ZHOU Zhi-Guo1,* 1Key Laboratory of Crop Physiology & Ecology in Southern China, Ministry of Agriculture / College of Agriculture, Nanjing Agricultural University, Nanjing 210095, China
2Institute of Industrial Crops, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
Fund: AbstractTwo year experiments were conducted at experimental station of Jiangsu Academy of Agricultural Sciences of Nanjing City with waterlogging treatments for seven days and fourteen days in pools with rainproof shelter at anthesis of the 6th or 8th fruiting branches of cotton NuCOTN 33B, taking well water conditions as control. The results showed that contents of soluble sugar, sucrose and starch of subtending leaves of bolls increased and the ratio of sucrose content to starch content decreased under waterlogging as compared with control and the increase ranges rised with lasting waterlogging duration, indicating that the sucrose exportation of subtending leaves was inhibited and became worse with lasting waterlogging duration. The activities of sucrose phosphate synthase (SPS), sucrose synthase (SS), and acid invertase (AI) increased and got higher with the rise in fruit branch position and waterlogging duration. The contents of soluble sugar, sucrose and starch were positively and significantly correlated with activities of SPS, SS, and AI (P<0.05), indicating that the increased activities of SPS, SS, and AI do not facilitate sucrose transiting out. As a result, the boll weight declined under waterlogging with the rise in fruit branch position and waterlogging duration. Therefore, the increased enzyme activity in sucrose metabolism and the impaired sucrose efflux are the main causes of boll weight decrease under waterlogging.
Keyword:Cotton (Gossypium hirsutum L.); Flowering and boll-forming stage; Waterlogging; Leaves sucrose metabolism; Boll weight Show Figures Show Figures
表1 花铃期渍水对棉花铃重的影响(2007年和2009年) Table 1 Effects of waterlogging on boll weight during flowering and boll-forming stage in 2007 and 2009 (g boll-1)
处理 Treatment
2007
2009
LFB
MFB
UFB
LFB
MFB
UFB
对照 Control
5.05 a
5.54 a
4.96 a
5.10 a
5.88 a
5.25 a
渍水7 d Waterlogging 7 days
—
—
—
4.77 b
5.06 b
4.07 b
渍水14 d Waterlogging 14 days
4.27 b
4.66 b
3.68 b
4.45 c
4.83 c
3.57 c
LFB、MFB和UFB分别代表下部、中部和上部果枝。数值后小写字母表示在0.05水平上差异显著。 LFB, MFB, and UFB stand for low fruiting branch, middle fruiting branch and upper fruiting branch, respectively. Values followed by different letters are significantly different at P<0.05.
表1 花铃期渍水对棉花铃重的影响(2007年和2009年) Table 1 Effects of waterlogging on boll weight during flowering and boll-forming stage in 2007 and 2009 (g boll-1)
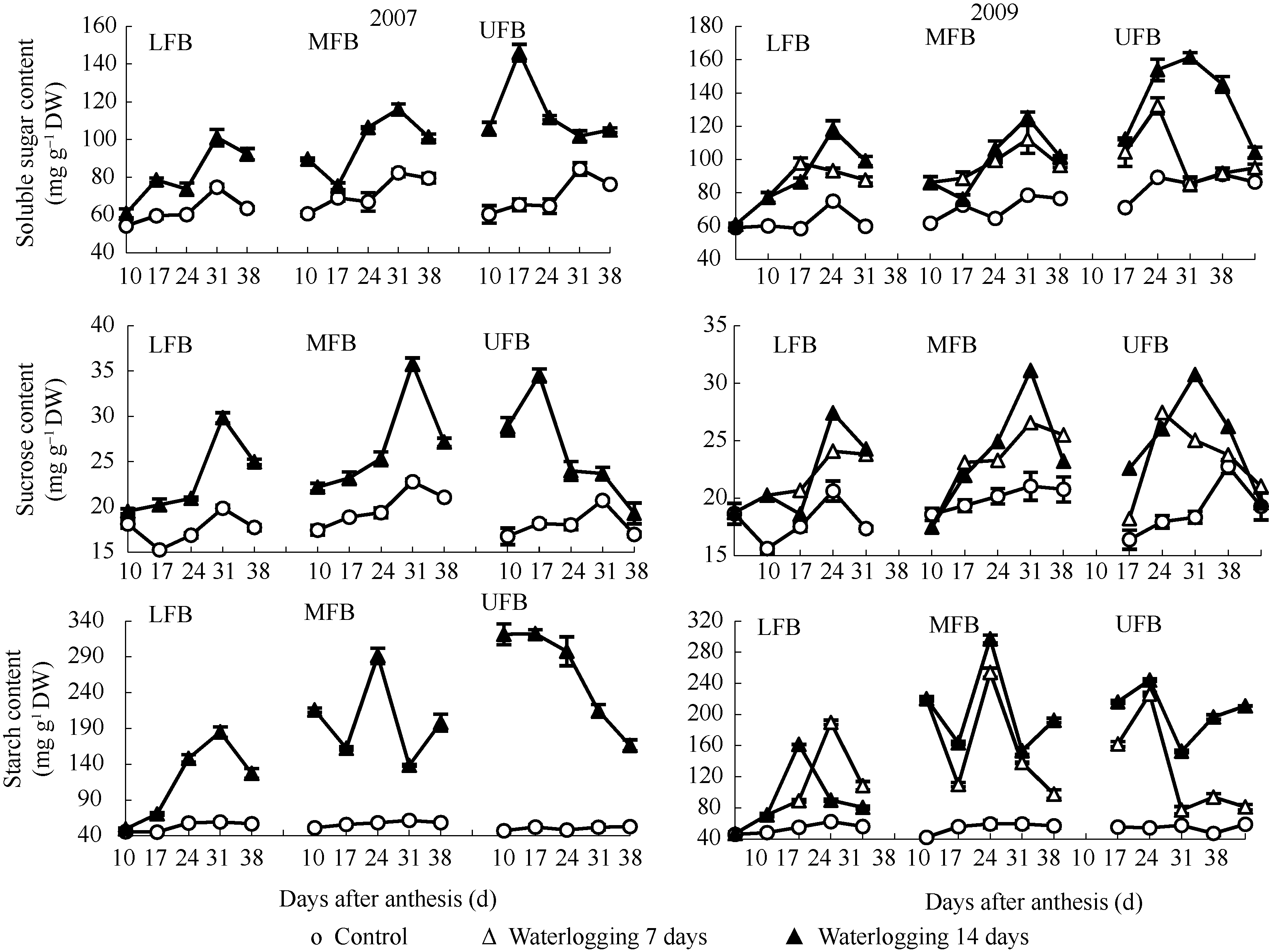
图1 花铃期渍水对棉铃对位叶碳水化合物含量的影响(2007年和2009年)LFB、MFB和UFB分别代表下部、中部和上部果枝。Fig. 1 Effects of waterlogging on carbohydrate contents of subtending leaves of bolls during flowering and boll-forming stage in 2007 and 2009LFB, MFB, and UFB stand for low fruiting branch, middle fruiting branch, and upper fruiting branch, respectively.
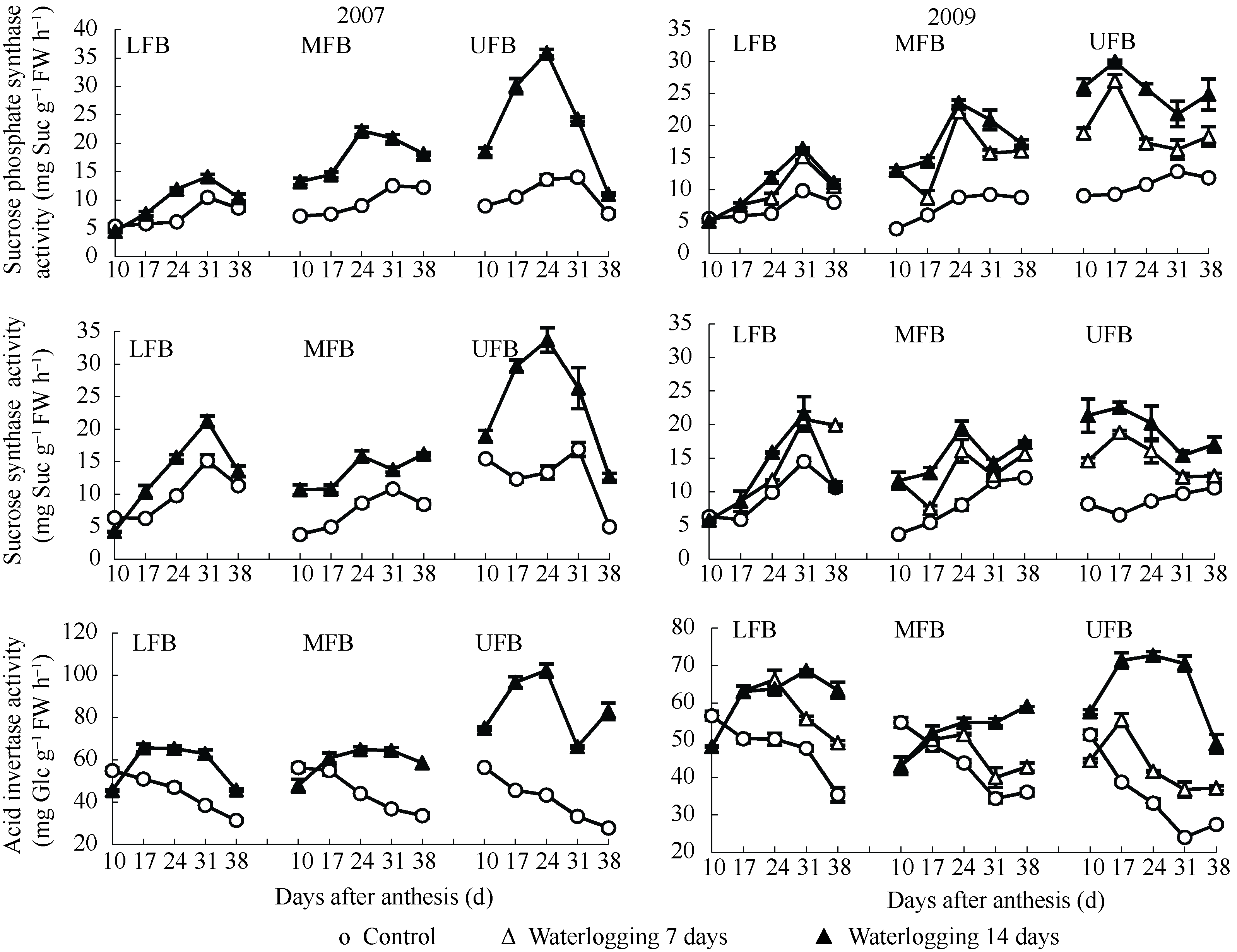
图2 花铃期渍水对棉铃对位叶蔗糖代谢相关酶活性的影响(2007年和2009年)LFB、MFB和UFB分别代表下部、中部和上部果枝。Fig. 2 Effects of waterlogging on sucrose metabolism enzyme activity of subtending leaves of bolls during flowering and boll-forming stage in 2007 and 2009LFB, MFB, and UFB stand for low fruiting branch, middle fruiting branch, and upper fruiting branch, respectively.
表2 棉铃对位叶碳水化合物含量与蔗糖代谢相关酶活性相关关系 Table 2 Relationship between carbohydrate contents and enzyme activities related to sucrose metabolism of subtending leaves of bolls
处理 Treatment
蔗糖磷酸合酶 Sucrose phosphate synthase
蔗糖合酶 Sucrose synthase
酸性转化酶 Acid invertase
可溶性糖 Soluble sugar
对照Control
0.694**
0.240
-0.738**
渍水Waterlogging
0.785**
0.638**
0.459**
蔗糖 Sucrose
对照Control
0.602**
0.350
-0.508**
渍水Waterlogging
0.526**
0.496**
0.368*
淀粉 Starch
对照Control
0.371*
0.320
-0.422*
渍水Waterlogging
0.734**
0.668**
0.444**
*和**分别表示在0.05和0.01水平上显著。* and** mean significant differences at 0.05 and 0.01 probability levels, respectively.
表2 棉铃对位叶碳水化合物含量与蔗糖代谢相关酶活性相关关系 Table 2 Relationship between carbohydrate contents and enzyme activities related to sucrose metabolism of subtending leaves of bolls
BangeM P, MilroyS P, ThongbaiP. Growth and yield of cotton in response to waterlogging. Field Crops Res, 2004, 88: 129-142[Cited within: 1][JCR: 2.474]
[2]
郭文琦, 赵新华, 陈兵林, 刘瑞显, 周治国. 氮素对花铃期短期渍水棉花根系生长的影响. 作物学报, 2009, 35: 1078-1085GuoW Q, ZhaoX H, ChenB L, LiuR X, ZhouZ G. Effects of nitrogen on cotton (Gossypium hirsutum L. ) root growth under short-term waterlogging during flowering and boll-forming stage. Acta Agron Sin, 2009, 35: 1078-1085 (in Chinese with English abstract)[Cited within: 1][CJCR: 1.667]
[3]
郭文琦, 张思平, 陈兵林, 周治国. 水分和氮肥运筹对棉花花后生物量和养分累积及氮素利用率的影响. 西北植物学报, 2008, 28: 2270-2277GuoW Q, ZhangS P, ChenB L, ZhouZ G. Effects of different water and nitrogen application on biomass and nutrients accumulation and nitrogen fertilization recovery rate of cotton after anthesis. Acta Bot Boreal-Occident Sin, 2008, 28: 2270-2277 (in Chinese with English abstract)[Cited within: 1]
ShiM F, ZengB, ShenJ H, LeiS T, ZhuZ, LiuJ H. A review of the correlation of flooding adaptability and carbohydrates in plants. Chin J Plant Ecol, 2010, 34: 855-866 (in Chinese with English abstract)[Cited within: 1][CJCR: 1.989]
[6]
KochK E. Carbohydrate-modulated gene expression in plants. Annu Rev Plant Physoil Mol Biol, 1996, 47: 509-540[Cited within: 1]
[7]
LoeweA, EinigW, ShiL, DizengremelP, HamppR. Mycorrhiza formation and elevated CO2 both increase the capacity for sucrose synthesis in source leaves of spruce and aspen. New Phytol, 2000, 145: 565-574[Cited within: 2][JCR: 6.736]
[8]
MalikA, ColmerT D, LambersH, SetterT L, SchortemeyerM. Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol, 2002, 153: 225-236[Cited within: 2][JCR: 6.736]
[9]
ChenH J, QuallsR G, BlankR R. Effect of soil flooding on photosynthesis, carbohydrate partitioning and nutrient uptake in the invasive exotic Lepidium latifolium. Aquat Bot, 2005, 82: 250-268[Cited within: 1][JCR: 1.593]
[10]
LiaoC T, LinC H. Physiological adaptation of crop plants to flooding stress. Proc Natl Sci Counc Roc (B), 2001, 25: 148-157[Cited within: 1]
[11]
SinglaN K, JainV, JainS, SawhneyS K. Activities of glycolytic enzymes in leaves and roots of contrasting cultivars of sorghum during flooding. Biol Plant, 2003, 47: 555-560[Cited within: 2][JCR: 2.32]
[12]
利容千, 王建波. 植物逆境细胞及生理学. 武汉: 武汉大学出版社, 2002. pp69-70LiR Q, WangJ B. Plant Stress Cell and Physiology. Wuhan: Wuhan University Press, 2002. pp69-70(in Chinese)[Cited within: 1]
GrofC P L, KnightD P, McNeilS D, LunnJ E, CampbellJ A. A modified assay method shows leaf sucrose-phosphate synthase activity is correlated with leaf sucrose content across a range of sugarcane varieties. Aust J Plant Physiol, 1998, 25: 499-502[Cited within: 1]
[15]
李永庚, 于振文, 姜东, 余松烈. 冬小麦旗叶蔗糖和籽粒淀粉合成动态及与其有关的酶活性的研究. 作物学报, 2001, 27: 658-664LiY G, YuZ W, JiangD, YuS L. Studies on the dynamic changes of the synthesis of sucrose in the flag leaf and starch in the grain and related enzymes of high-yielding wheat. Acta Agron Sin, 2001, 27: 658-664 (in Chinese with English abstract). [Cited within: 1][CJCR: 1.667]
[16]
GeigenbergerP, StittM. Sucrose synthase catalyses a reversible reaction in vivo in developing potato tubers and other plant tissues. Planta, 1993, 189: 329-339[Cited within: 1][JCR: 3.347]
[17]
DéjardinA, RochatC, WuillèmeS, BoutinJ P. Contribution of sucrose synthase, ADP-glucose pyrophosphorylase and starch synthase to starch synthesis in developing pea seeds. Plant Cell Environ, 1997, 20: 1421-1430[Cited within: 1][JCR: 5.135]
[18]
PraxedesS C, DaMattaF M, LoureiroM E, FerrãoM A G, CordeiroA T. Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffee canephora Pierre var. Kouillou) leaves. Environ Exp Bot, 2006, 56: 263-273[Cited within: 1][JCR: 2.578]
[19]
WellsR. Response of leaf ontogeny and photosynthetic activity to reproductive growth in cotton. Plant Physiol, 1988, 87: 274-279[Cited within: 1][JCR: 6.555]
[20]
张文静, 胡宏标, 陈兵林, 王友华, 李文峰, 周治国. 棉铃对位叶生理特性的基因型差异及其与铃重形成的关系. 棉花学报, 2007, 19: 296-303ZhangW J, HuH B, ChenB L, WangY H, LiW F, ZhouZ G. Relationship between genotypic difference of physiological characteristics in leaf subtending boll and boll weight forming. Cotton Sci, 2007, 19: 296-303 (in Chinese with English abstract)[Cited within: 1][CJCR: 1.462]
[21]
周青, 王友华, 许乃银, 张传喜, 周治国, 陈兵林. 温度对棉铃对位叶生理特性及铃重形成的影响. 西北植物学报, 2009, 29: 518-527ZhouQ, WangY H, XuN Y, ZhangC X, ZhouZ G, ChenB L. Effect of temperature on physiological characteristics in leaf subtending boll and boll weight forming. Acta Bot Boreali-Occident Sin, 2009, 29: 518-527 (in Chinese with English abstract)[Cited within: 1]
[22]
刘敬然, 孟亚利, 王友华, 陈兵林, 张国伟, 周治国. 外源6-BA和ABA对不同播种期棉花产量和品质及其棉铃对位叶光合产物的影响. 作物学报, 2013, 39: 1078-1088LiuJ R, LiuJ J, MengY L, WangY H, ChenB L, ZhangG W, ZhouZ G. Effect of 6-BA and ABA applications on yield, quality and photosynthate contents in the subtending leaf of cotton with different planting dates. Acta Agron Sin, 2013, 39: 1078-1088[Cited within: 3][CJCR: 1.667]
[23]
中国科学院上海植物生理研究所编. 现代植物生理学实验指南. 北京: 科学出版社, 1999. p127Shanghai Institute of Plant Physiology, Chinese Academy of Sciences. Modern Laboratory Manual of Plant Physiology. Beijing: Science Press, 1999. p127 (in Chinese)[Cited within: 1]
[24]
汤章城. 现代植物生理学实验指南. 北京: 科学出版社, 1999. pp126-128TangZ C. Modern Laboratory Manual of Plant Physiology. Beijing: Science Press, 1999. pp126-128(in Chinese)[Cited within: 1]
[25]
HuberS C. Biochemical mechanism for regulation of sucrose accumulation in leaves during photosynthesis. Plant Physiol, 1989, 91: 656-662[Cited within: 1][JCR: 6.555]
[26]
郭文琦, 刘瑞显, 周治国, 陈兵林. 施氮量对花铃期短期渍水棉花叶片气体交换参数和叶绿素荧光参数的影响. 植物营养与肥料学报, 2010, 16: 362-369GuoW Q, LiuR X, ZhouZ G, ChenB L. Effects of nitrogen fertilization on gas exchange and chlorophyll fluorescence parameters of leaf during the flowering and boll-forming stage of cotton under short-term waterlogging. Plant Nutr Fert Sci, 2010, 16: 362-369 (in Chinese with English abstract)[Cited within: 1][CJCR: 1.883]
[27]
AlbrechtG, MustrophA, FoxT C. Sugar and fructan accumulation during metabolic adjustment between respiration and fermentation under low oxygen conditions in wheat roots. Physiol Plant, 2004, 120: 93-105[Cited within: 1][JCR: 6.555]
[28]
IslamM A, MacdonaldS E. Ecophysiological adaptations of black spruce (Picea mariana) and tamarack (Larix laricina) seedlings to flooding. Trees, 2004, 18: 35-42[Cited within: 1][JCR: 1.925]
[29]
YangJ, ZhangJ, WangZ, ZhuQ. Activities of starch hydrolytic enzymes and sucrose-phosphate synthase in the stems of rice subjected to water stress during grain filling. J Exp Bot, 2001, 52: 2169-2179[Cited within: 1][JCR: 5.242]
[30]
王维, 张建华, 杨建昌, 朱庆森. 水分胁迫对贪青迟熟水稻茎贮藏碳水化合物代谢及产量的影响. 作物学报, 2004, 30: 196-204WangW, ZhangJ H, YangJ C, ZhuQ S. Effect of water stress on metabolism of stored carbohydrate of stem and yield in rice grown under unfavorable- delayed senescence. Acta Agron Sin, 2004, 30: 196-204 (in Chinese with English abstract)[Cited within: 1][CJCR: 1.667]
[31]
HiranoT, UchidaN, AzumaT, YasudaT. Relationship between export rate of photoassimilates and activation state of sucrose phosphate synthase in submerged floating rice. Jpn J Crop Sci, 1997, 66: 675-681[Cited within: 1]
[32]
HaradaT, IshizawaK. Starch degradation and sucrose metabolism during anaerobic growth of pondweed (Potamogeton distinctus A. Benn. ) turions. Plant Soil, 2003, 253: 125-135[Cited within: 1][JCR: 2.638]
[33]
RosaM, PradoC, PodazzaG, InterdonatoR, GonzálezJ A, HilalM, PradoF E. Soluble sugars-Metabolism, sensing and abiotic stress. Plant Signal Behav, 2009, 4: 388-393[Cited within: 1]
[34]
王文泉, 张福锁. 高等植物厌氧适应的生理及分子机制. 植物生理学通讯, 2001, 37: 63-70WangW Q, ZhangF S. The physiological and molecular mechanism of adaptation to anaerobiosis in higher plants. Plant Physiol Commun, 2001, 37: 63-70 (in Chinese with English abstract)[Cited within: 2]
[35]
HookD D, BrownC L. Root adaptations and relative flood tolerance of five hardwood species. Forensic Sci, 1973, 19: 225-229[Cited within: 1]
[36]
StancatoG C, MazzaferaP, BuckeridgeM S. Effect of a drought period on the mobilization of non-structural carbohydrates, photosynthetic efficiency and water status in an epiphytic orchid. Plant Physiol Biochem, 2001, 39: 1009-1016[Cited within: 1]
[37]
CrawfordR M M, BraendleR. Oxygen deprivation stress in a changing environment. J Exp Bot, 1996, 47: 145-159[Cited within: 1][JCR: 5.242]
[38]
SchlüterU, CrawfordR M M. Long term anoxia tolerance in leaves of Acorus calamus L. and Iris pseudacorus L. J Exp Bot, 2001, 52: 2213-2225[Cited within: 1][JCR: 5.242]
 , 刘瑞显
, 刘瑞显

{kind=link}
{kind=link}