关键词:单核苷酸位点多态性(SNP); 光合作用; 产量; 大豆; 自然群体 Correlation and Association Analysis between Biomass and Yield Components in Soybean CHAO Mao-Ni1,**, HAO De-Rong2,**, YIN Zhi-Tong3, ZHANG Jin-Yu1, SONG Hai-Na1, ZHANG Huai-Ren1, CHU Shan-Shan1, ZHANG Guo-Zheng1, YU De-Yue1,* 1National Center for Soybean Improvement, National Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China
2 Jiangsu Yanjiang Institute of Agricultural Sciences, Nantong 226541, China
3 Jiangsu Provincial Key Laboratory of Crop Genetics and Physiology, Yangzhou University, Yangzhou 225009, China
** Contributed equally to this work
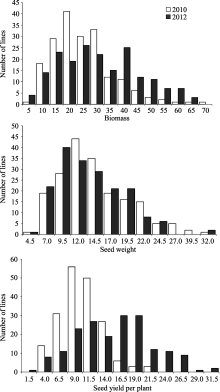
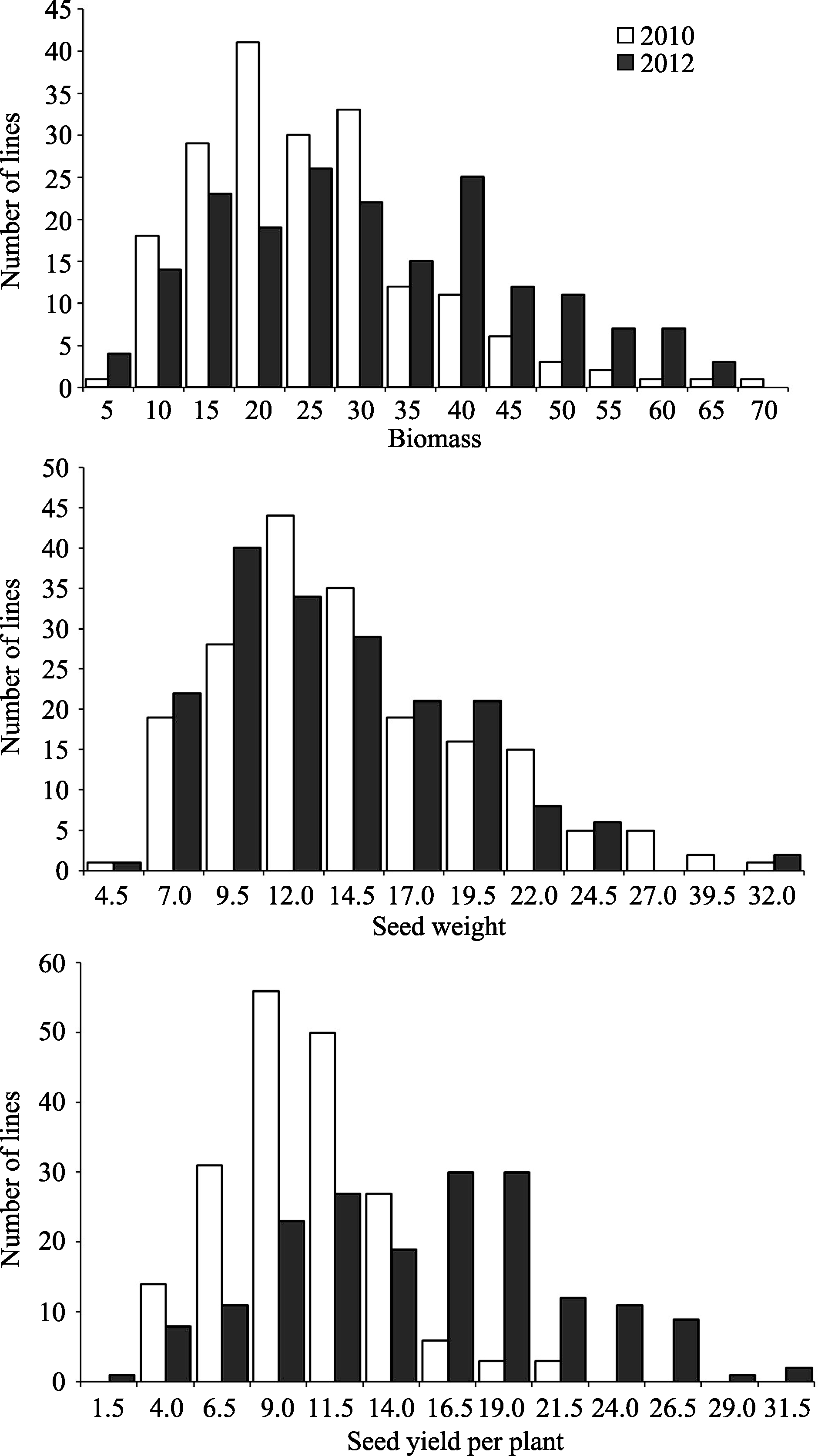
AbstractBiomass, one of the main factors that determine the effective economic yield, has an important effect on the final seed yield. In this study, a genome-wide association analysis was conducted to detect key single-nucleotide polymorphisms (SNPs) associated with biomass and yield components using 1142 SNPs in a soybean landraces panel. There existed abundant phenotypic and genetic diversities and significant correlations among biomass and yield components in the population, and the correlation between biomass and seed yield was slightly higher than that between biomass and seed weight. Genome-wide association analysis using a mixed linear model detected 41, 56, and 29 SNPs associated with biomass, seed weight and seed yield respectively. Among them, 6, 19, and 1 SNPs were detected in two environments. In addition, 15 SNPs were found co-associated with two or more different traits and BARC-029051-06057 on chromosome 19 was associated with the three traits, which implies a partially common genetic basis for the three traits. Many SNPs detected in our study were found co-associated with soybean chlorophyll, chlorophyll fluorescence parameters and yield components in our previous study. The identification of these significant SNPs will be helpful to better understand the genetic basis of biomass and yield components, and facilitate the pyramiding of favorable alleles for future high-yield breeding by marker-assisted selection in soybean.
Keyword:Single nucleotide polymorphisms (SNP); Photosynthesis; Yield; Soybean; Natural population Show Figures Show Figures
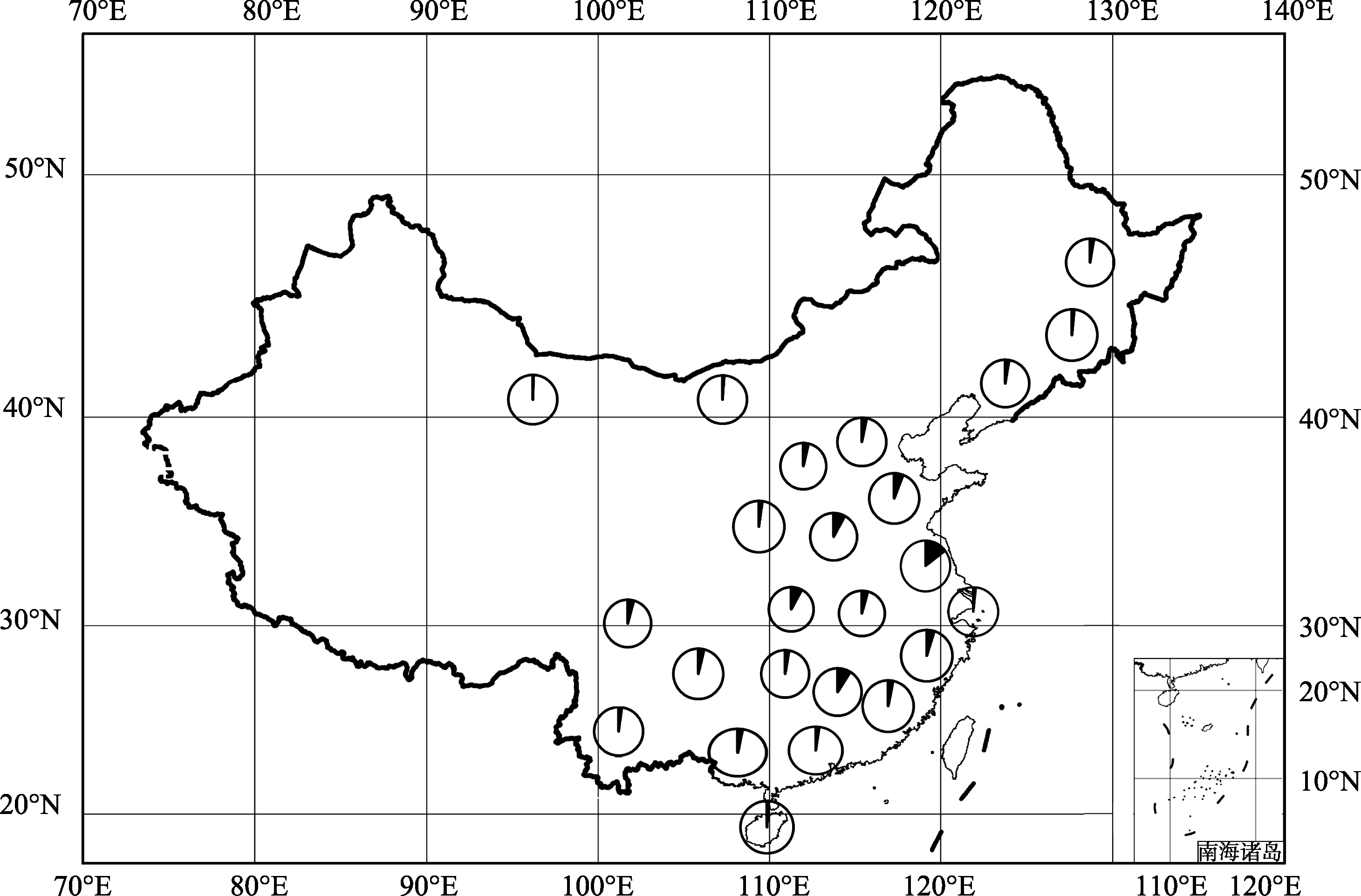
图1 191份栽培大豆地方品种地理分布每个圆圈代表不同的省份, 每个省份的材料数目用圆圈的实心部分表示。Fig. 1 Geographical distribution of 191 soybean landraces in our studyCircles represent different provinces, the number of soybean landraces per province is indicated by closed portion of the circle.
表1 大豆自然群体中生物量与产量组分的描述统计 Table 1 Descriptive statistics of soybean biomass and yield components in soybean natural population
性状 Trait
环境 Environment
均值 Mean
标准差 SD
最小值 Min.
最大值 Max.
显著性 Significance
Ga
G×Eb
生物量 Biomass
2011
22.80
11.26
3.70
68.63
**
**
2012
28.50
14.46
3.22
63.15
百粒重 100-seed weight
2011
13.20
5.44
4.10
30.63
**
**
2012
12.42
5.03
4.37
30.78
单株籽粒产量 Seed yield per plant
2011
8.80
3.47
2.18
20.12
**
**
2012
13.92
6.10
1.33
29.26
a多个环境基因型之间的显著性;b基因型与环境之间的互作。**表示在0.01水平上显著。 aSignificant difference between genotype across different environments;bGenotype × Environment. ** Significant at 0.01 probability level.
表1 大豆自然群体中生物量与产量组分的描述统计 Table 1 Descriptive statistics of soybean biomass and yield components in soybean natural population
表2 大豆生物量与产量组分间的相关系数 Table 2 Correlation coefficients between biomass and yield components in soybean natural population
性状 Trait
生物量 Biomass
百粒重 100-seed weight
单株籽粒产量 Seed yield per plant
生物量 Biomass
0.330**
0.661**
百粒重 100-seed weight
0.253**
0.502**
单株籽粒产量 Seed yield per plant
0.331**
0.368**
对角线下为2011年相关系数,对角线上为2012年相关系数。**表示在0.01水平上显著。 Correlation coefficients for 2011 were listed in down left, and those for 2012 in upper right.** Significant at the 0.01 probability level.
表2 大豆生物量与产量组分间的相关系数 Table 2 Correlation coefficients between biomass and yield components in soybean natural population
表3 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3 SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population
标记 Marker
染色体 Chr.
图位 Position
显著性 -lg P
贡献率 R2
共位性状 Co-located traits
2011
2012
2011
2012
生物量Biomass
BARC-013749-01246
1
3134522
2.89
ns
3.15
nd
BARC-030807-06945
1
53063797
ns
3.03
nd
2.61
SYa PNc SNc
BARC-025791-05069
2
1071247
ns
2.11
nd
1.7
BARC-013995-01298
2
1892774
ns
2.11
nd
1.71
BARC-044739-08781
2
2350068
ns
2.72
nd
2.97
BARC-029969-06761
2
2454551
ns
2.62
nd
2.89
CCIb
BARC-029969-06762
2
2454551
ns
2.59
nd
2.86
CCIb
BARC-018187-02537
2
4344760
2.19
ns
1.79
nd
BARC-014855-01911
4
718393
ns
2.19
nd
2.45
BARC-040651-07807
5
619849
ns
2.05
nd
1.64
BARC-014463-01558
5
34202590
2.08
ns
1.68
nd
BARC-027546-06605
5
41414453
2.10
ns
2.33
nd
BARC-045081-08872
8
10954129
ns
2.62
nd
2.89
BARC-014967-01939
8
11214962
ns
2.09
nd
2.48
BARC-013333-00485
8
45285760
2.01
3.41
2.25
3.74
SYa
BARC-022031-04262
9
812264
3.54
ns
3.87
nd
BARC-013601-01173
10
37792789
ns
3.07
nd
2.64
BARC-029531-06209
10
37848327
ns
2.66
nd
2.24
BARC-024519-04954
11
1054437
2.17
ns
2.4
nd
BARC-018713-03241
11
5089229
2.11
ns
1.71
nd
BARC-015917-02015
11
5104751
2.11
ns
1.71
nd
BARC-028289-05814
11
10175104
ns
2.23
nd
2.45
CCIb
BARC-016279-02316
11
10804671
2.60
5.49
2.2
5.05
BARC-042035-08159
13
43467832
ns
2.20
nd
2.42
PNc SNc
BARC-020519-04649
13
43730737
ns
2.22
nd
2.45
BARC-014917-01920
15
1848113
2.52
ns
2.78
nd
BARC-039087-07438
15
9556913
4.11
ns
4.44
nd
BARC-018461-02916
15
12255921
3.75
2.03
4.06
2.23
BARC-030059-06796
15
20686406
2.42
ns
2.66
nd
BARC-013055-00431
15
23168999
2.60
ns
2.87
nd
BARC-039931-07614
15
36576527
2.22
ns
2.45
nd
BARC-017679-03103
15
38000598
2.33
ns
2.58
nd
BARC-022037-04263
17
7416485
ns
2.72
nd
2.99
BARC-025681-04992
18
61043717
ns
2.08
nd
2.33
BARC-029051-06057
19
47906580
2.74
2.17
2.34
1.78
SYaSWa
BARC-028335-05833
20
694198
2.15
ns
2.38
nd
BARC-042685-08347
20
41982750
ns
2.10
nd
2.31
BARC-025969-05190
20
43678501
ns
2.06
nd
2.31
BARC-031459-07097
20
43897353
ns
2.82
nd
2.44
BARC-041439-07983
20
46530282
2.70
2.03
2.98
2.26
BARC-013645-01207
20
46623915
2.92
2.04
3.21
2.25
表3 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3 SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population
(续表3-1) 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3(Continued) SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population
(续表3-1) 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3(Continued) SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population
(续表3-2) 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3-2(Continued)SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population
标记 Marker
染色体 Chr.
图位 Position
显著性 -lg P
贡献率 R2
共位性状 Co-located traits
2011
2012
2011
2012
BARC-040087-07654
15
47628308
2.02
2.82
1.48
2.18
ETo/ABSb ETo/Trob PIABSb
BARC-042895-08450
16
31292708
ns
2.11
nd
2.11
CCIb
BARC-032251-08930
17
15485936
2.24
ns
2.24
nd
BARC-018479-02918
18
1243529
2.24
ns
2.24
nd
BARC-038795-07333
18
50205922
2.02
ns
2.02
nd
BARC-018117-02529
19
5531602
2.44
ns
2.52
nd
BARC-013481-00498
19
42334572
3.34
3.26
3.3
3.19
BARC-044913-08838
19
43125170
2.15
ns
2.17
nd
BARC-029051-06057
19
47906580
ns
2.24
nd
1.63
SYa Biomassa
BARC-021827-04218
19
48091800
ns
2.41
nd
2.38
SWc
单株籽粒产量Seed yield per plant
BARC-030807-06945
1
53063797
ns
4.51
nd
4.35
Biomassa PNcSNc
BARC-014927-01924
3
39794924
ns
2.02
nd
2.39
BARC-028709-05992
3
40654334
2.92
3.88
2.73
3.71
SYcSWacABS/RCbETo/ABSbPIABSb
BARC-016977-02171
4
7533029
2.07
ns
2.5
nd
BARC-015061-02551
4
31994543
ns
2.60
nd
2.34
SWaCCIb
BARC-041047-07901
4
48958879
3.61
ns
3.48
nd
BARC-040033-07641
5
34734702
ns
2.21
nd
2.61
BARC-028171-05783
5
37928020
ns
2.47
nd
2.21
BARC-020259-04536
5
37999262
ns
2.40
nd
2.15
BARC-027726-06646
8
9489476
2.03
ns
2.45
nd
BARC-028297-05816
8
14653588
ns
2.10
nd
1.83
BARC-038877-07374
8
17940650
2.24
ns
2.7
nd
BARC-013333-00485
8
45285760
ns
2.89
nd
3.42
Biomassa
BARC-021491-04133
9
6593074
ns
2.70
nd
2.46
BARC-018911-03272
10
3519980
2.08
ns
2.5
nd
BARC-032347-08952
10
43920755
2.24
ns
2.02
nd
BARC-018693-02992
10
47121190
2.66
ns
3.2
nd
CCIb
BARC-025709-05013
12
39889639
2.31
ns
2.09
nd
BARC-028887-06033
13
36556810
ns
2.92
nd
3.41
BARC-025561-06521
13
36822597
ns
2.66
nd
2.41
BARC-045151-08895
14
7917464
ns
2.54
nd
2.98
SWaCCIb
BARC-013235-00458
15
50333637
ns
2.49
nd
2.93
BARC-016069-02054
16
1351860
2.40
ns
2.9
nd
BARC-039151-07458
17
41489651
4.07
3.99
BARC-031583-07115
19
36278536
2.59
ns
2.39
nd
BARC-022451-04330
19
37951038
ns
2.07
nd
3.27
BARC-029051-06057
19
47906580
ns
2.77
nd
2.54
Biomassa SWa
BARC-021827-04218
19
48091720
3.06
ns
3.65
nd
BARC-021343-04041
20
2049166
2.39
ns
2.18
nd
a: 参照本研究结果共位的性状;b: 参照郝德荣等对叶绿素荧光定位结果, 与本研究共位的性状[ 19];c: 参照郝德荣等[ 15]对多个环境产量定位结果, 与本研究共位的性状; R2: 贡献率; ns: 相应的环境中关联不显著; nd: 没有数据; CCI: 叶绿素含量; Fv/ Fm: PSII最大光化学产量; ABS/RC: 单位反应中心吸收的光能; ETo/Tro: 用于电子传递的量子产额; ETo/ABS: 用于电子传递的量子产额; PIABS: 以吸收光能为基础的性能指数; SW: 百粒重; PN: 单株荚数; SN: 单株粒数; SY: 单株籽粒产量。 a: The traits co-located with the results of the present study;b: The traits co-located with the previous results of the chlorophyll and chlorophyll fluorescence[19]; c: The traits co-located with the previous results of yield and yield components in soybean across multiple environments[ 15]; R2: Contribution rate; ns: Marker was not detected at significant level in corresponding environment; nd: no data; CCI: Chlorophyll content; Fv/ Fm: quantum efficiency of open reaction center; ABS/RC: energy absorbed by RC of photosynthetic apparatus; ETo/TRo: probability that a trapped exciton moves an electron into the electron transport chain beyond QA-; ETo/ABS: Quantum yield for electron transport; PIABS: performance index; SW: 100-seed weight; PN: number of pods per plant; SN: number of seeds per plant; SY: seed yield per plant.
(续表3-2) 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3-2(Continued) SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population
周恩远, 刘丽君, 祖伟, 孙聪姝. 春大豆农艺性状与品质相关关系的研究. 东北农业大学报, 2008, 39: 145-149ZhouE Y, LiuL J, ZuW, SunC S. Study on relationship between agronomic traits and quality traits in spring soybean. J Northeast Agric Univ, 2008, 39: 145-149 (in Chinese with English abstract)[本文引用:1][CJCR: 0.1053]
[2]
马占峰, 赵淑文, 杨琪, 邹玉梅. 生物产量──大豆高产育种的物质基础. 东北农业大学学报, 1995, 26: 125-130MaZ F, ZhaoS W, YaoQ, ZhouY M. Biological yeild─physical basis of soyeban high yeild breeding. J Northeast Agric Univ, 1995, 26: 125-130 (in Chinese with English abstract)[本文引用:1][CJCR: 0.1053]
[3]
杨胜荣, 黄宗洪, 向关伦, 甘雨, 杨占烈, 潘建慧, 郭慧. 以提高生物产量为途径选育杂交水稻新组合. 农技服务, 2010: 1267-1269YangS R, HuangZ H, XiangG L, GanY, YangZ L, PanJ H, GuoH. Selestive new combinations of hybrid rice by raising biological production. Agric Technol Ser, 2010, 1267-1269 (in Chinese)[本文引用:1]
[4]
AustinR B, FordM A, MorganC L. Genetic improvement in the yield of winter wheat: a further evaluation. J Agric Sci, 1989, 112: 295-301[本文引用:1][JCR: 2.041]
[5]
FischerR A, ReesD, SayreK D, LuZ M, CondonA G, SaavedraA L. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci, 1998, 38: 1467-1475[本文引用:1][JCR: 1.641]
[6]
JinJ, LiuX, WangG, MiL, ShenZ, ChenX, HerbertS J. Agronomic and physiological contributions to the yield improvement of soybean cultivars released from 1950 to 2006 in Northeast China. Field Crops Res, 2010, 115: 116-123[本文引用:1][JCR: 2.474]
[7]
Parry M A J, ReynoldsM, SalvucciM E, RainesC, AndralojcP J, ZhuX G, PriceG D, CondonA G, FurbankR T. Raising yield potential of wheat: II. Increasing photosynthetic capacity and efficiency. J Exp Bot, 2011, 62: 453-467[本文引用:1][JCR: 5.364]
[8]
黄中文, 赵团结, 盖钧镒. 大豆不同产量水平生物量积累与分配的动态分析. 作物学报, 2009, 35: 1483-1490HuangZ W, ZhaoT J, GaiJ Y. Dynamic analysis of biomass accumulation and partition in Soybean with different yield levels. Acta Agron Sin, 2009, 35: 1483-1490 (in Chinese with English abstract). [本文引用:1][CJCR: 1.8267]
[9]
BoardJ E, ModaliH. Dry matter accumulation predictors for optimal yield in soybean. Crop Sci, 2005, 45: 1790-1799[本文引用:1][JCR: 1.641]
[10]
OrfJ H, ChaseK, AdlerF R, MansurL M, LarkK G. Genetics of soybean agronomic traits: II. Interactions between yield quantitative trait loci in soybean. Crop Sci, 1999, 39: 1652-1657[本文引用:1][JCR: 1.641]
[11]
YuanJ, NjitiV N, MeksemK, IqbalM J, TriwitayakornK, KassemM A, DavisG T, SchmidtM E, LightfootD A. Quantitative trait loci in two soybean recombinant inbred line populations segregating for yield and disease resistance. Crop Sci, 2002, 42: 271-277[本文引用:1][JCR: 1.641]
[12]
KabelkaE A, DiersB W, FehrW R, LeRoyA R, BaianuI C, YouT, NeeceD J, NelsonR L. Putative alleles for increased yield from soybean plant introductions. Crop Sci, 2004, 44: 784-791[本文引用:1][JCR: 1.641]
[13]
GuzmanP S, DiersB W, NeeceD J, St Martin S K, LeRoyA R, GrauC R, HughesT J, NelsonR L. QTL associated with yield in three backcross-derived populations of soybean. Crop Sci, 2007, 47: 111-122[本文引用:1][JCR: 1.641]
[14]
PalomequeL, LiuL J, LiW B, HedgesB, CoberE R, RajcanI. QTL in mega-environments: I. Universal and specific seed yield QTL detected in a population derived from a cross of high- yielding adapted × high-yielding exotic soybean lines. Theor Appl Genet, 2009, 119: 417-427[本文引用:1][JCR: 3.297]
[15]
HaoD, ChengH, YinZ, CuiS, ZhangD, WangH, YuD. Identification of single nucleotide polymorphisms and haplotypes associated with yield and yield components in soybean (Glycine max) land races across multiple environments. Theor Appl Genet, 2012, 124: 447-458[本文引用:7][JCR: 3.297]
[16]
黄中文, 赵团结, 喻德跃, 陈受宜, 盖钧镒. 大豆生物量积累、收获指数及产量间的相关与QTL分析. 作物学报, 2008, 34: 944-951HuangZ W, ZhaoT J, YuD Y, ChenS Y, GaiJ Y. Correlation and QTL mapping of biomass accumulation, apparent harvest index, and yield in soybean. Acta Agron Sin, 2008, 34: 944-951 (in Chinese with English abstract). [本文引用:5][CJCR: 1.8267]
[17]
印志同, 宋海娜, 孟凡凡, 许晓明, 喻德跃. 大豆光合气体交换参数的QTL分析. 作物学报, 2009, 36: 92-100YinZ T, SongH N, MengF F, XuX M, YuD Y. QTL mapping for photosynthetic gas-exchange parameters in soybean. Acta Agron Sin, 2009, 36: 92-100 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[18]
AinsworthE A, YendrekC R, SkoneczkaJ A, LongS P. Accelerating yield potential in soybean: potential targets for biotechnological improvement. Plant Cell Environ, 2011, 35: 38-52[本文引用:3][JCR: 5.215]
[19]
HaoD, ChaoM, YinZ, YuD. Genome-wide association analysis detecting significant single nucleotide polymorphisms for chlorophyll and chlorophyll fluorescence parameters in soybean (Glycine max) land races. Euphytica, 2012: 1-13[本文引用:3][JCR: 1.554]
[20]
张贤泽, 马占峰, 赵淑文, 庞士铨. 大豆不同品种光合速率与产量关系的研究. 作物学报, 1986, 12: 43-48ZhangZ X, MaZ F, ZhaoS W, PangS C. The relationship between net photosynthetic rate and yield formation in soybean. Acta Agron Sin, 1986, 12: 43-48 (in Chinese with English abstract)[本文引用:2][CJCR: 1.8267]
[21]
杜维广, 张桂茹, 满为群, 栾晓燕, 陈怡, 谷秀芝. 大豆科学, 1999, 18: 154-159DuW G, ZhangG R, ManW Q, LuanX Y, ChenY, GuX Z. Study on relationship between soybean photosynthesis and yield. Soybean Sci, 1999, 18: 154-159 (in Chinese with English abstract)[本文引用:2]
[22]
MehetreS S, JamadagniB M. Biomass partitioning and growth characters in relation to plant architecture in soybean. Soybean Genet Newsl, 1996, 23: 92-97[本文引用:1]
[23]
SchneebergerK, WeigelD. Fast-forward genetics enabled by new sequencing technologies. Trends Plant Sci, 2011, 16: 282-288[本文引用:1][JCR: 11.047]
[24]
FultonT M, Beck-BunnT, EmmattyD, EshedY, LopezJ, PetiardV, UhligJ, ZamirD, TanksleyS D. QTL analysis of an advanced backcross of lycopersicon peruvianum to the cultivated tomato and comparisons with QTLs found in other wild species. Theor Appl Genet, 1997, 95: 881-894[本文引用:1][JCR: 3.297]
[25]
ThummaB R, NaiduB P, Chand raA, CameronD F, BahnischL M, LiuC. Identification of causal relationships among traits related to drought resistance in Stylosanthes scabra using QTL analysis. J Exp Bot, 2001, 52: 203-214[本文引用:1][JCR: 5.364]
[26]
龚月桦, 高俊凤. 高等植物光合同化物的运输与分配. 西北植物学报, 1999, 19: 564-570GongY H, GaoJ F. Transport and partitioning of photoassimilate in higher plant. Acta Bot Boreal-Occident Sin, 1999, 19: 564-570 (in Chinese with English abstract)[本文引用:1]
[27]
王玲玲, 杜吉到, 郑殿峰, 宋微微, 陈丽霞, 田静斋, 吕美芳. 大豆源库流关系的研究进展. 大豆科学, 2009, 28: 167-171WangL L, DuJ D, ZhengD F, SongW W, ChenL X, TianJ Z, LüM F. Advances in the studies of relation among source sink and flux of soybean. Soybean Sci, 2009, 28: 167-171 (in Chinese with English abstract)[本文引用:2]
[28]
LiuW, FuY, HuG, SiH, ZhuL, WuC, SunZ. Identification and fine mapping of a thermo-sensitive chlorophyll deficient mutant in rice (Oryza sativa L. ). Planta, 2007, 226: 785-795[本文引用:1][JCR: 3.0]
[29]
印志同, 孟凡凡, 宋海娜, 晁毛妮, 许晓明, 邓德祥, 喻德跃. 大豆开花盛期快速叶绿素荧光参数的 QTL 分析. 中国农业科学, 2011, 44: 4980-4987YinZ T, MengF F, SongH N, ChaoM N, XuX M, DengD X, YuD Y. QTL mapping for fast chlorophyll fluorescence parameters in soybean. Sci Agric Sin, 2011, 44: 4980-4987 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4522]
[30]
FloodP J, HarbinsonJ, Aarts M G M. Natural genetic variation in plant photosynthesis. Trends Plant Sci, 2011, 16: 327-335[本文引用:1][JCR: 11.047]
[31]
LefebvreS, LawsonT, FryerM, ZakhleniukO V, LloydJ C, RainesC A. Increased sedoheptulose-1, 7-bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol, 2005, 138: 451-460[本文引用:1][JCR: 6.535]
[32]
OrfJ H, ChaseK, JarvikT, MansurL M, CreganP B, AdlerF R, LarkK G. Genetics of soybean agronomic traits: I. Comparison of three related recombinant inbred populations. Crop Sci, 1999, 39: 1642-1651[本文引用:1][JCR: 1.641]
[33]
黄中文, 赵团结, 喻德跃, 陈受宜, 盖钧镒. 大豆产量有关性状QTL的检测. 中国农业科学, 2009, 42: 4155-4165HuangZ W, ZhaoT J, YuD Y, ChenS Y, GaiJ Y. Detection of QTLs of yield related traits in soybean. Sci Agric Sin, 2009, 42: 4155-4165 (in Chinese with English abstract)[本文引用:2][CJCR: 1.4522]
[34]
KimH K, KangS T, SuhD Y. Analysis of quantitative trait loci associated with leaflet types in two recombinant inbred lines of soybean. Plant Breed, 2005, 124: 582-589[本文引用:2][JCR: 1.596]
[35]
SpechtJ E, ChaseK, Macrand erM, GraefG L, ChungJ, MarkwellJ P, GermannM, OrfJ H, LarkK G. Soybean response to water: a QTL analysis of drought tolerance. Crop Sci, 2001, 41: 493-509[本文引用:2][JCR: 1.641]
[36]
Mian M A R, BaileyM A, TamulonisJ P, ShipeE R, CarterT E, ParrottW A, AshleyD A, HusseyR S, BoermaH R. Molecular markers associated with seed weight in two soybean populations. Theor Appl Genet, 1996, 93: 1011-1016[本文引用:1][JCR: 3.297]
[37]
CsanadiG, VollmannJ, StiftG, LelleyT. Seed quality QTLs identified in a molecular map of early maturing soybean. Theor Appl Genet, 2001, 103: 912-919[本文引用:1][JCR: 3.297]
[38]
LeeS H, ParkK Y, LeeH S, ParkE H, BoermaH R. Genetic mapping of QTLs conditioning soybean sprout yield and quality. Theor Appl Genet, 2001, 103: 702-709[本文引用:1][JCR: 3.297]
[39]
GuzmanP S, DiersB W, NeeceD J, St Martin S K, LeRoyA R, GrauC R, HughesT J, NelsonR L. QTL associated with yield in three backcross-derived populations of soybean. Crop Sci, 2007, 47: 111-122[本文引用:1][JCR: 1.641]
[40]
FunatsukiH, KawaguchiK, MatsubaS, SatoY, IshimotoM. Mapping of QTL associated with chilling tolerance during reproductive growth in soybean. Theor Appl Genet, 2005, 111: 851-861[本文引用:1][JCR: 3.297]
[41]
HoeckJ A, FehrW R, ShoemakerR C, WelkeG A, JohnsonS L, CianzioS R. Molecular marker analysis of seed size in soybean. Crop Sci, 2003, 43: 68-74[本文引用:1][JCR: 1.641]
[42]
HytenD L, PantaloneV R, SamsC E, SaxtonA M, Land au-EllisD, StefaniakT R, SchmidtM E. Seed quality QTL in a prominent soybean population. Theor Appl Genet, 2004, 109: 552-561[本文引用:1][JCR: 3.297]
 , 郝德荣
, 郝德荣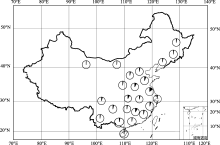

{kind=link}
{kind=link}