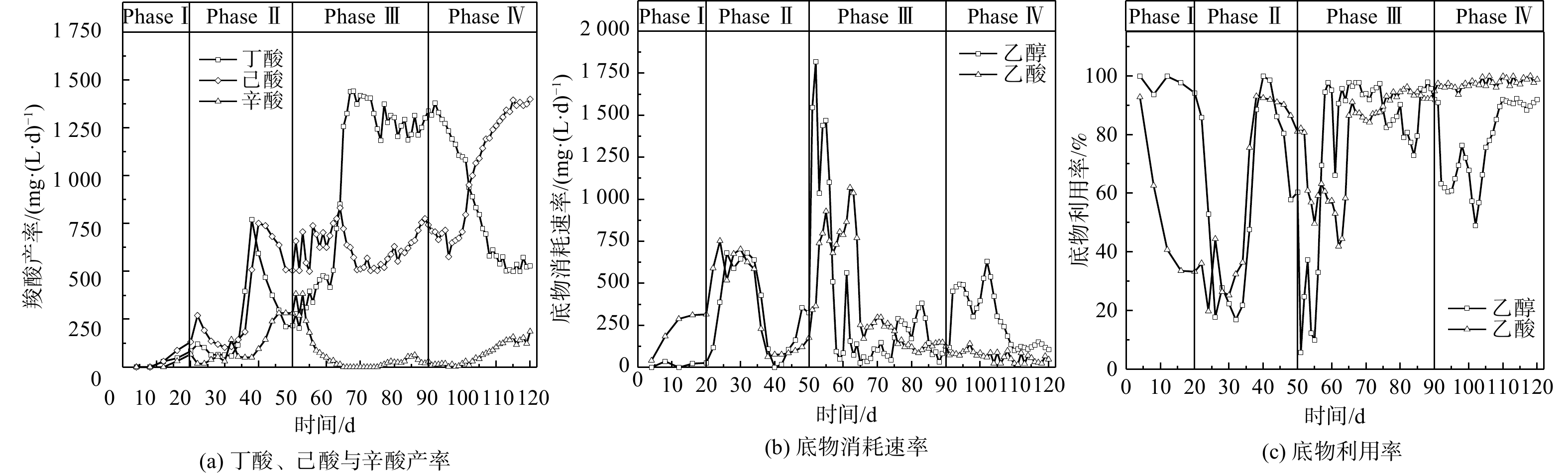
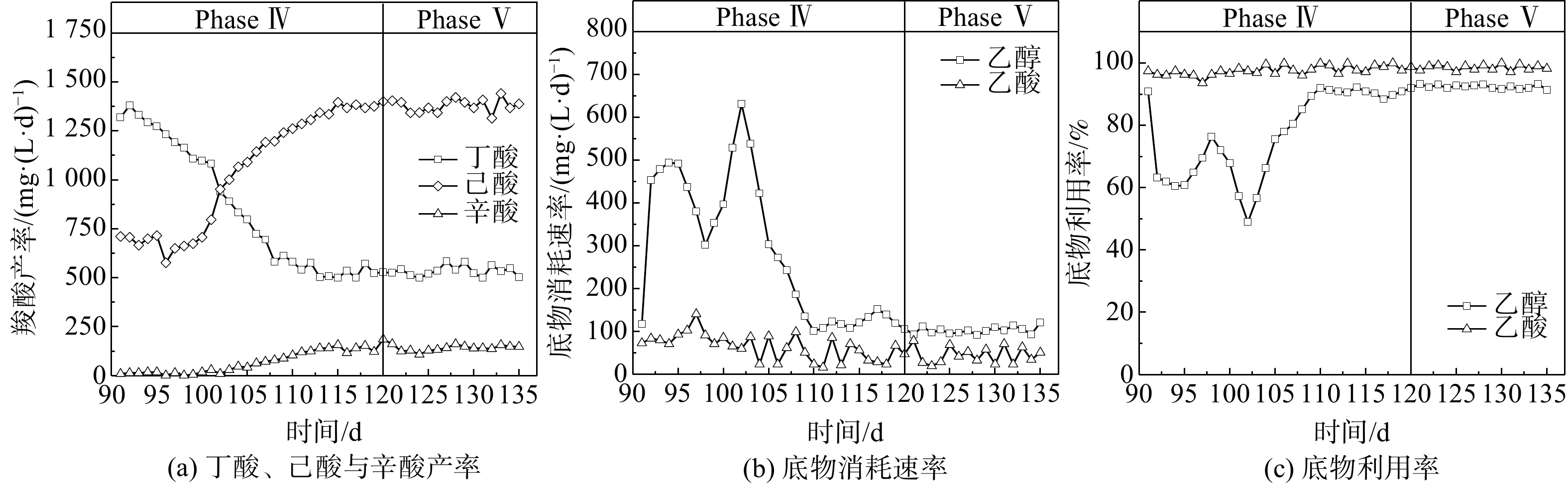
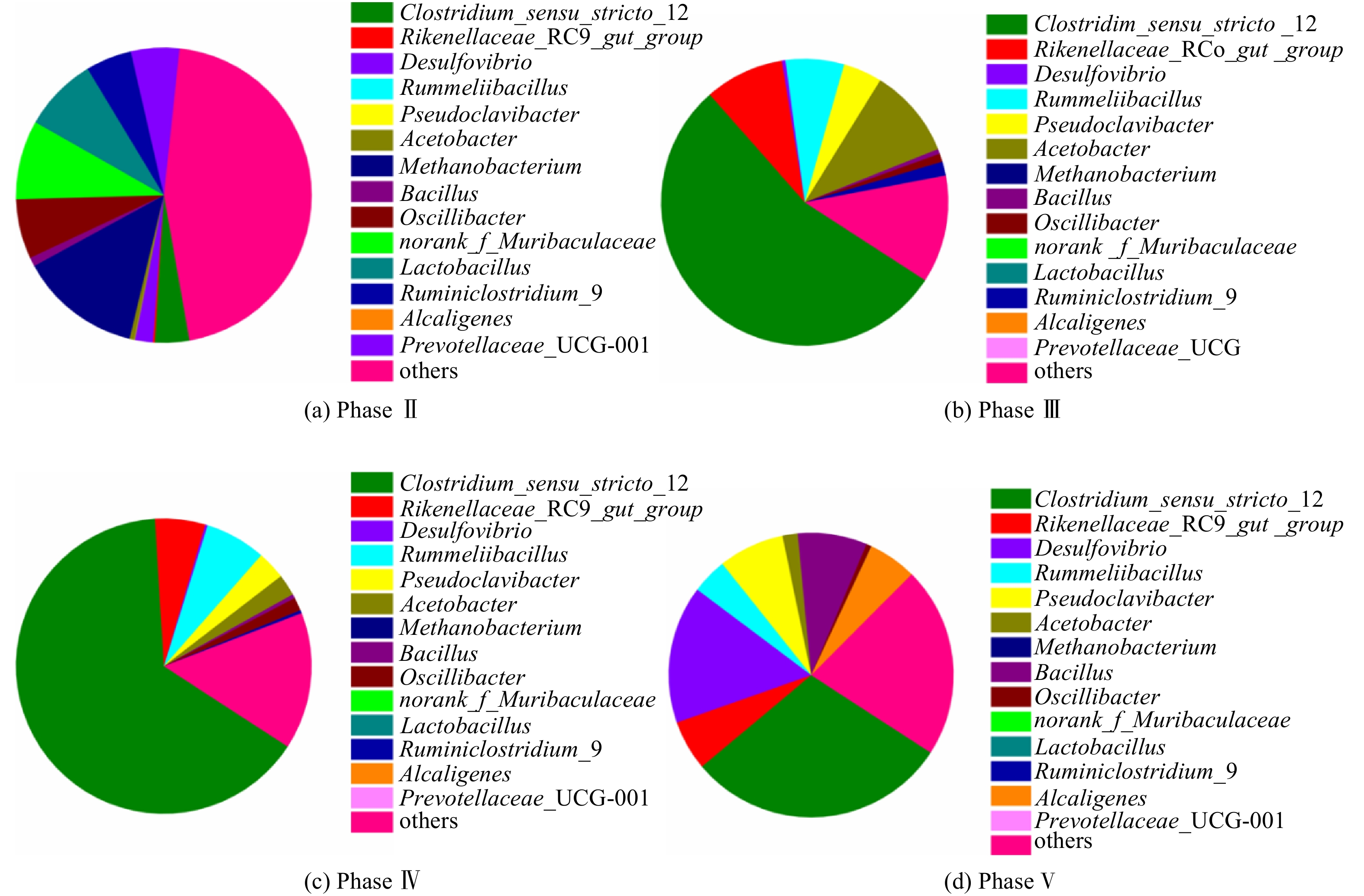
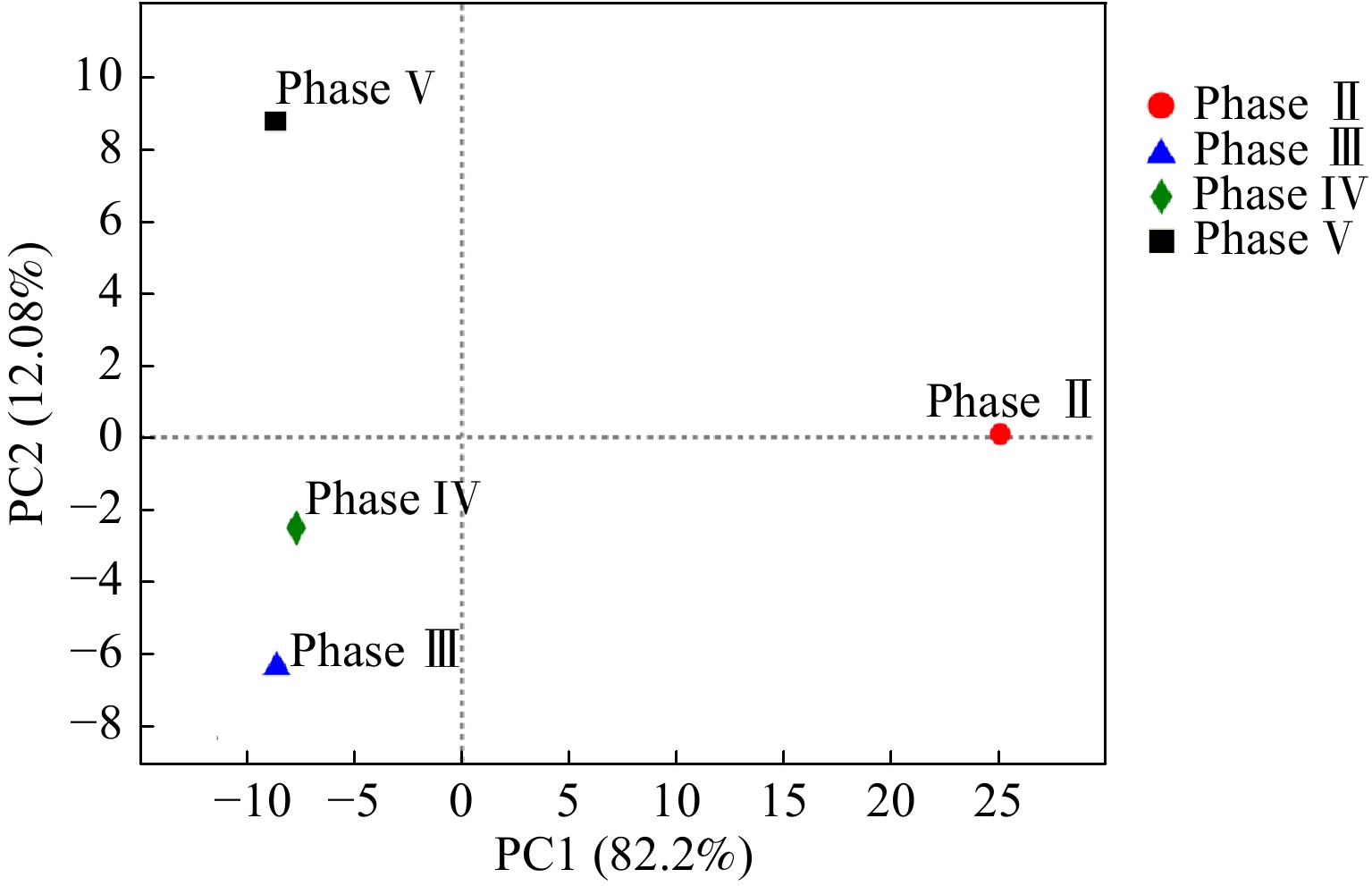
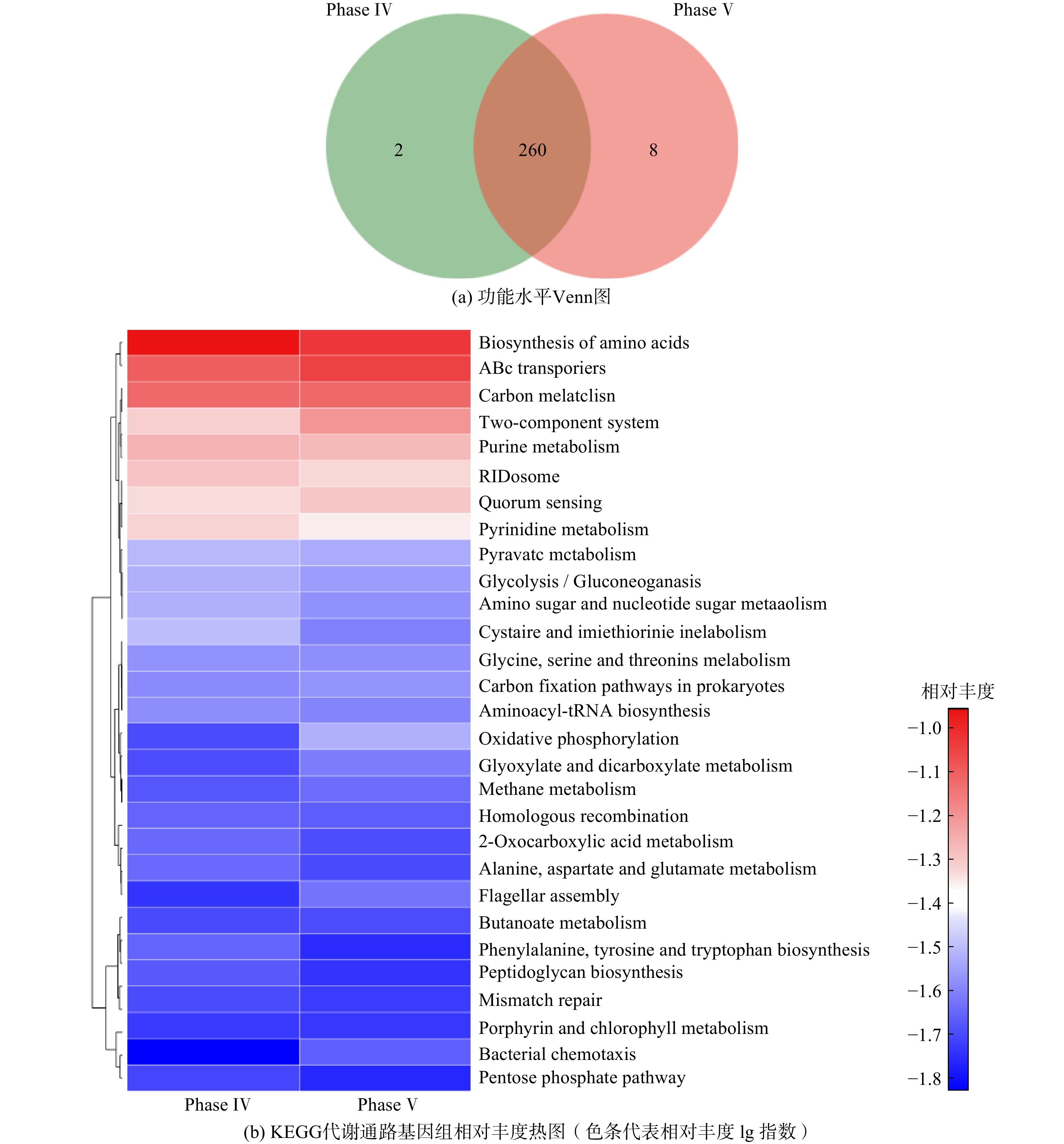
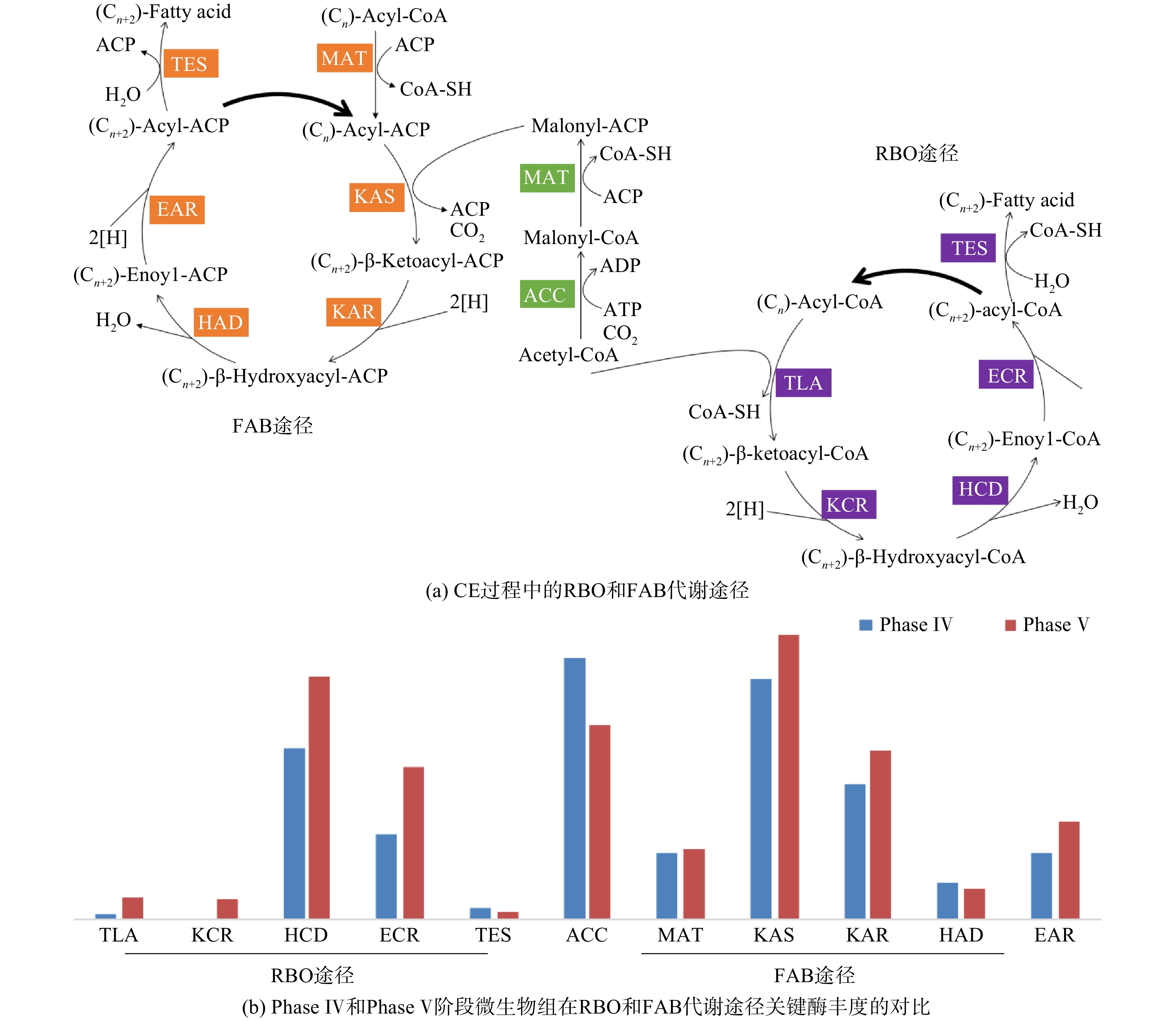
Abstract:To determine the feasibility of producing medium-chain fatty acids (MCFAs) from waste activated sludge in mixed culture, this study firstly optimized bioreactor parameters, i.e. hydraulic retention time (HRT) and ethanol : acid ratio, to assimilate anaerobes and later adopted the “two-stage fermentation” strategy, in which alkaline pretreated sludge was subjected to short-term acidogenesis and microbial chain elongation The results showed that during the 135-day chain elongation (CE) over the long term, with ethanol∶acid ratio=2∶1, when HRT was shortened from 20 d to 5 d (Phase Ⅰ~Ⅲ), the CE functional microbe, Clostridium sensu stricto_12, evolved as the dominant genus (relative abundance 65.21%). However, the maximal productivity of n-caproate was merely 775 mg?(L?d)?1. Subsequently, with ethanol∶acid ratio increased to 3∶1 (Phase Ⅳ), the productivity of n-caproate boosted to 1 402 mg?(L?d)?1, demonstrating an increased product selectivity towards MCFAs. In PhaseⅤ (test phase), during which the substrate swamped from synthetic ethanol/acetate wastewater to sludge fermentation liquor (SFL), the n-caproate productivity maintained at 1 400 mg?(L?d)?1. Based on the metagenomics analysis, both reverse β-oxidation (RBO) and fatty acid biosynthesis (FAB) pathways were involved in microbial chain elongation for MCFAs production. Moreover, as compared to synthetic ethanol/acetate wastewater, SFL increased the relative abundance of some key functional enzymes for the RBO and FAB pathways. The present study provided the practical evidence for continuous production of MCFAs from waste activated sludge, and more importantly, it elucidated the microbial and ecological mechanisms. Taken together, it shed a light on the sludge-derived value-added chemicals for its valorization. Key words:waste activated sludge/ medium-chain fatty acids/ anaerobic fermentation/ chain elongation/ sludge valorization.
图1驯化期微生物碳链延长代谢表现 Figure1.Performance of chain elongation under varied operational conditions during acclimation period
表1市政污水处理厂剩余污泥的基本理化性质 Table1.Characteristics of waste activated sludge from a municipal wastewater treatment plant
pH
TS/(g?L?1)
VS/(g?L?1)
COD/(mg?L?1)
SCOD/(mg?L?1)
TKN/(mg?L?1)
TAN/(mg ?L?1)
6.72±0.05
29.66±0.60
18.63±0.38
21 100±420
346±46
2 560±94
12.4±1.5
pH
TS/(g?L?1)
VS/(g?L?1)
COD/(mg?L?1)
SCOD/(mg?L?1)
TKN/(mg?L?1)
TAN/(mg ?L?1)
6.72±0.05
29.66±0.60
18.63±0.38
21 100±420
346±46
2 560±94
12.4±1.5
下载: 导出CSV 表2不同运行周期的反应器运行条件 Table2.Operational conditions of different phases of bioreactor for chain elongation
运行周期
HRT/d
醇酸比
运行时间/d
Phase Ⅰ
20
2∶1
0~20
Phase Ⅱ
10
2∶1
21~50
Phase Ⅲ
5
2∶1
51~90
Phase Ⅳ
5
3∶1
91~120
Phase Ⅴ
5
3∶1
121~135
运行周期
HRT/d
醇酸比
运行时间/d
Phase Ⅰ
20
2∶1
0~20
Phase Ⅱ
10
2∶1
21~50
Phase Ⅲ
5
2∶1
51~90
Phase Ⅳ
5
3∶1
91~120
Phase Ⅴ
5
3∶1
121~135
下载: 导出CSV 表3驯化期不同运行条件下的总烷基量、平均碳链长度与碳效率 Table3.Total alkyl groups, average chain length and carbon conversion efficiency of microbial chain elongation process under different operational conditions during the acclimation period
运行周期
总烷基量/(mmol·L?1)
平均碳链长度
碳效率/%
Phase Ⅰ
291.8
6.82
33.4
Phase Ⅱ
543.4
6.83
62.1
Phase Ⅲ
507
5.63
72.9
Phase Ⅳ
535.6
6.51
64.8
运行周期
总烷基量/(mmol·L?1)
平均碳链长度
碳效率/%
Phase Ⅰ
291.8
6.82
33.4
Phase Ⅱ
543.4
6.83
62.1
Phase Ⅲ
507
5.63
72.9
Phase Ⅳ
535.6
6.51
64.8
下载: 导出CSV 表4碳链延长(CE)过程涉及到的关键酶及其在Phase Ⅳ和Phase Ⅴ阶段微生物组中的丰度 Table4.Key enzymes involved in chain elongation pathways and their abundance in Phase Ⅳ and Ⅴ
MEERBURG F A, BOON N, VAN WINCKEL T, et al. Optimizing retention times in high-rate contact stabilization for maximal recovery of organics from wastewater[J]. Environmental Science & Technology, 2016, 50(17): 9781-9790.
[2]
ALLOUL A, GANIGUE R, SPILLER M, et al. A three-step approach for the valorization of sewage organics as commodities[J]. Environmental Science & Technology, 2018, 52(12): 6729-6742.
[3]
ANGENENT L T, USACK J G, XU J, et al. Integrating electrochemical, biological, physical, and thermochemical process units to expand the applicability of anaerobic digestion[J]. Bioresource Technology, 2018, 247(9): 1085-1094.
[4]
CAGNETTA C, COMA M, VLAEMINCK S E, et al. Production of carboxylates from high rate activated sludge through fermentation[J]. Bioresource Technology, 2016, 217(3): 165-172.
[5]
ANGENENT L T, RICHTER H, BUCKEL W, et al. Chain elongation with reactor microbiomes: open-culture biotechnology to produce biochemicals[J]. Environmental Science & Technology, 2016, 50(6): 2796-2810.
[6]
AGLER M T, SPIRITO C M, USACK J G, et al. Chain elongation with reactor microbiomes: Upgrading dilute ethanol to medium-chain carboxylates[J]. Energy & Environmental Science, 2012, 5(8): 8189-8192.
[7]
GE S J, USACK J G, SPIRITO C M, et al. Long-term n-caproic acid production from yeast-fermentation beer in an anaerobic bioreactor with continuous product extraction[J]. Environmental Science & Technology, 2015, 49(13): 8012-8021.
[8]
KUCEK L A, XU J J, NGUYEN M, et al. Waste conversion into n-caprylate and n-caproate: Resource recovery from wine lees using anaerobic reactor microbiomes and in-line extraction[J]. Frontiers In Microbiology, 2016, 7: 1892.
[9]
VASUDEVAN D, RICHTER H, ANGENENT L T. Upgrading dilute ethanol from syngas fermentation to n-caproate with reactor microbiomes[J]. Bioresource Technology, 2014, 151(9): 378-382.
[10]
RICHTER H, MOLITOR B, DIENDER M, et al. A narrow pH range supports butanol, hexanol, and octanol production from syngas in a continuous co-culture of Clostridium ljungdahlii and Clostridium kluyveri with in-line product extraction[J]. Frontiers in Microbiology, 2016, 7: 1773.
[11]
LIU C, LUO G, LIU H P, et al. CO as electron donor for efficient medium chain carboxylate production by chain elongation: Microbial and thermodynamic insights[J]. Chemical Engineering Journal, 2020, 390: 124577. doi: 10.1016/j.cej.2020.124577
[12]
LI X, CHEN Y G, ZHAO S, et al. Efficient production of optically pure L-lactic acid from food waste at ambient temperature by regulating key enzyme activity[J]. Water Research, 2015, 70(11): 148-157.
[13]
XU J J, HAO J X, GUZMAN J J L, et al. Temperature-phased conversion of acid whey waste into medium-chain carboxylic acids via lactic acid: No external e-donor[J]. Joule, 2018, 2(2): 280-295. doi: 10.1016/j.joule.2017.11.008
[14]
LE C C, STUCKEY D C. Colorimetric measurement of carbohydrates in biological wastewater treatment systems: A critical evaluation[J]. Water Research, 2016, 94: 280-287. doi: 10.1016/j.watres.2016.03.008
[15]
LE C C, KUNACHEVA C, STUCKEY D C. "Protein" measurement in biological wastewater treatment systems: A critical evaluation[J]. Environmental Science & Technology, 2016, 50(6): 3074-3081.
[16]
Lü C X, SHEN Y W, LI C, et al. Redox-active biochar and conductive graphite stimulate methanogenic metabolism in anaerobic digestion of waste-activated sludge: Beyond direct interspecies electron transfer[J]. Sustainable Chemistry & Engineering, 2020, 8(33): 12626-12636.
[17]
LIU Y H, Lü F, SHAO L M. Alcohol-to-acid ratio and substrate concentration affect product structure in chain elongation reactions initiated by unacclimatized inoculum[J]. Bioresource Technology, 2016, 218: 1140-1150. doi: 10.1016/j.biortech.2016.07.067
[18]
LONKAR S, FU Z H, HOLTZAPPLE M. Optimum alcohol concentration for chain elongation in mixed-culture fermentation of cellulosic substrate[J]. Biotechnology and Bioengineering, 2016, 113(12): 2597-2604. doi: 10.1002/bit.26024
[19]
STEINBUSCH K J J, HAMELERS H V M, PLUGGE C M, et al. Biological formation of caproate and caprylate from acetate: Fuel and chemical production from low grade biomass[J]. Energy & Environmental Science, 2011, 4(1): 216-224.
[20]
WU S L, WEI W, SUN J, et al. Medium-chain fatty acids and long-chain alcohols production from waste activated sludge via two-stage anaerobic fermentation[J]. Water Research, 2020, 186: 116381. doi: 10.1016/j.watres.2020.116381
[21]
HAN W H, HE P J, SHAO L M, et al. Metabolic interactions of a chain elongation microbiome[J]. Applied and Environmental Microbiology, 2018, 84(22): 01614-01618.
[22]
GROOTSCHOLTEN T I, STEINBUSCH K J, HAMELERS H V, et al. High rate heptanoate production from propionate and ethanol using chain elongation[J]. Bioresource Technology, 2013, 136: 715-718. doi: 10.1016/j.biortech.2013.02.085
[23]
GANIGUE R, NAERT P, CANDRY P, et al. Fruity flavors from waste: A novel process to upgrade crude glycerol to ethyl valerate[J]. Bioresource Technology, 2019, 289: 121574. doi: 10.1016/j.biortech.2019.121574
[24]
CANDRY P, ULCAR B, PETROGNANI C, et al. Ethanol: Propionate ratio drives product selectivity in odd-chain elongation with Clostridium kluyveri and mixed communities[J]. Bioresource Technology, 2020, 313: 123651. doi: 10.1016/j.biortech.2020.123651
[25]
AGLER M T, SPIRITO C M, USACK J G, et al. Development of a highly specific and productive process for n-caproic acid production: Applying lessons from methanogenic microbiomes[J]. Water Science and Technology, 2014, 69(1): 62-68. doi: 10.2166/wst.2013.549
[26]
SEEDORF H, FRICKE W F, VEITH B, et al. The genome of Clostridium kluyveri, a strict anaerobe with unique metabolic features[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(6): 2128-2133. doi: 10.1073/pnas.0711093105
[27]
WU S L, SUN J, CHEN X M, et al. Unveiling the mechanisms of medium-chain fatty acid production from waste activated sludge alkaline fermentation liquor through physiological, thermodynamic and metagenomic investigations[J]. Water Research, 2020, 169: 115218. doi: 10.1016/j.watres.2019.115218
[28]
DE VRIEZE J, REGUEIRO L, PROPS R, et al. Presence does not imply activity: DNA and RNA patterns differ in response to salt perturbation in anaerobic digestion[J]. Biotechnology for Biofuels, 2016, 9: 244. doi: 10.1186/s13068-016-0652-5
1.School of Environmental Science and Engineering, Shanghai Jiao Tong University, Shanghai 200240, China 2.Hunan BISEN Environmental & Energy Co. Ltd., Changsha 410100, China Received Date: 2021-01-12 Accepted Date: 2021-04-25 Available Online: 2021-11-16 Keywords:waste activated sludge/ medium-chain fatty acids/ anaerobic fermentation/ chain elongation/ sludge valorization Abstract:To determine the feasibility of producing medium-chain fatty acids (MCFAs) from waste activated sludge in mixed culture, this study firstly optimized bioreactor parameters, i.e. hydraulic retention time (HRT) and ethanol : acid ratio, to assimilate anaerobes and later adopted the “two-stage fermentation” strategy, in which alkaline pretreated sludge was subjected to short-term acidogenesis and microbial chain elongation The results showed that during the 135-day chain elongation (CE) over the long term, with ethanol∶acid ratio=2∶1, when HRT was shortened from 20 d to 5 d (Phase Ⅰ~Ⅲ), the CE functional microbe, Clostridium sensu stricto_12, evolved as the dominant genus (relative abundance 65.21%). However, the maximal productivity of n-caproate was merely 775 mg?(L?d)?1. Subsequently, with ethanol∶acid ratio increased to 3∶1 (Phase Ⅳ), the productivity of n-caproate boosted to 1 402 mg?(L?d)?1, demonstrating an increased product selectivity towards MCFAs. In PhaseⅤ (test phase), during which the substrate swamped from synthetic ethanol/acetate wastewater to sludge fermentation liquor (SFL), the n-caproate productivity maintained at 1 400 mg?(L?d)?1. Based on the metagenomics analysis, both reverse β-oxidation (RBO) and fatty acid biosynthesis (FAB) pathways were involved in microbial chain elongation for MCFAs production. Moreover, as compared to synthetic ethanol/acetate wastewater, SFL increased the relative abundance of some key functional enzymes for the RBO and FAB pathways. The present study provided the practical evidence for continuous production of MCFAs from waste activated sludge, and more importantly, it elucidated the microbial and ecological mechanisms. Taken together, it shed a light on the sludge-derived value-added chemicals for its valorization.
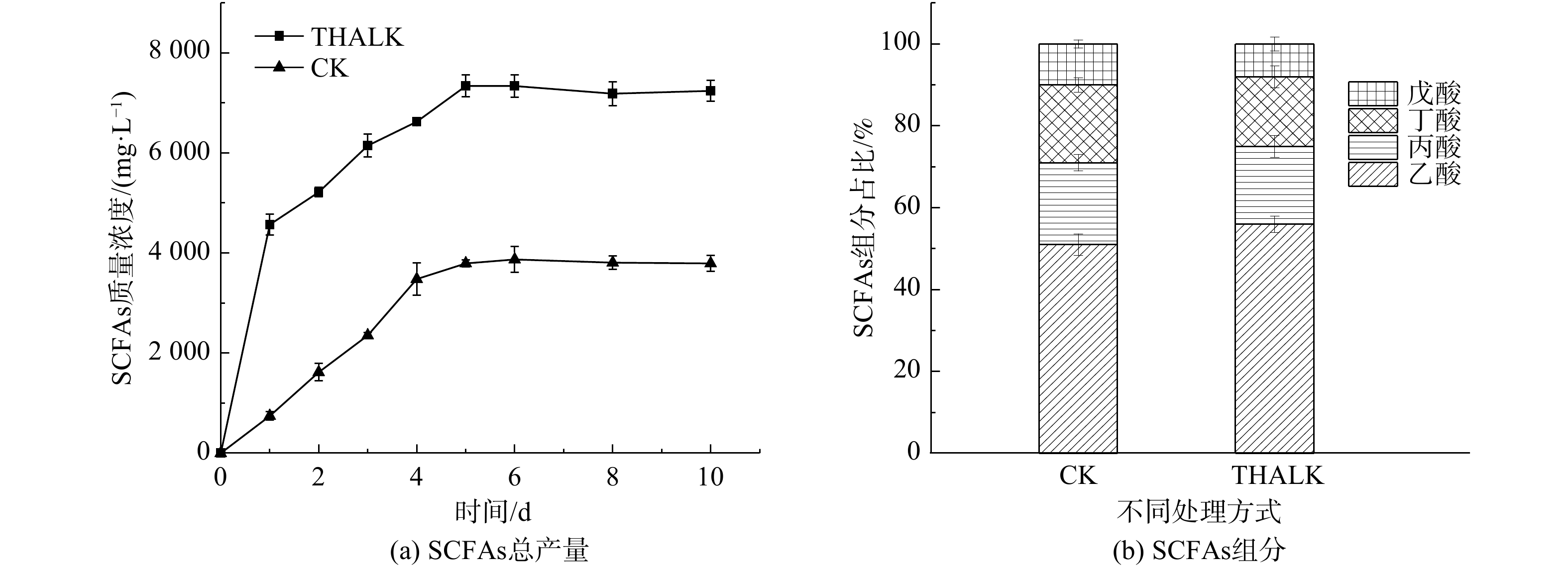
剩余污泥pH使用FE28标准pH计(Mettler-Toledo,瑞士)测定;其余理化指标(包括TS、VS、COD、TKN、TAN),采用国标法测定。污泥厌氧发酵液经离心15 min (转速离心力为12 000g)后,取上清液过膜(滤膜孔径0.45 μm),分析其SCOD、多糖、蛋白质和SCFAs浓度等。多糖和蛋白质分别采用蒽酮-硫酸比色法(以葡萄糖为标准物质)和考马斯亮蓝法(以牛血清蛋白为标准物质)[14-15]测定。SCFAs(乙酸、丙酸、正丁酸、异丁酸、正戊酸、异戊酸)的组分和浓度采用气相色谱法测定,即在对其酸化预处理后(添加适量3%磷酸溶液将样品pH降至4.0以下),使用装配有火焰离子化检测仪(FID)和DB-FFAP型毛细管柱的GC-7890B气相色谱(安捷伦,美国)进行定量分析。微生物连续发酵产MCFAs实验中的底物(乙醇、SCFAs)与产物(丁酸、己酸、辛酸)浓度的定量分析方法与SCFAs相同。
 下载:
下载:  点击查看大图
点击查看大图