National Key Laboratory of Science and Technology on Vacuum Electronics, School of Electronic Science and Engineering (National Exemplary School of Microelectronics), University of Electronic Science and Technology of China, Chengdu 610054, China
Fund Project:Project supported by the National Natural Science Foundation of China (Grant No. 61921002) and the National Basic Science Center of National Natural Science Foundation of China (Grant No. 61988102)
Received Date:16 September 2021
Accepted Date:12 October 2021
Available Online:22 October 2021
Published Online:20 December 2021
Abstract:Potassium channels play an important role in repolarizing the nerve cell action potentials. There are many types of potassium channel proteins, and potassium channels allow potassium ions to specifically pass through the cell membrane, thereby maintaining the resting potential of nerve cells. In this paper, molecular dynamics simulation method is used to simulate the effects of 53.7 THz terahertz wave with different amplitudes on the secondary structure of KcsA potassium channel protein and the potassium ions rate. It is found in this study that under the action of the 53.7 THz terahertz wave, the number of alpha helices in KcsA potassium channel protein decreases, and the number of beta sheets and the number of coils increase. In addition, the 53.7 THz terahertz wave can accelerate potassium ions through the KcsA potassium channel. In this article, the effects of terahertz waves on potassium channel proteins are analyzed through the secondary structure of proteins, and a new perspective for the interaction between terahertz waves and biological functional molecules is presented as well. Keywords:potassium channel/ protein secondary structure/ potassium ion rate/ resonance absorption
4.不同强度太赫兹波对钾离子通道蛋白二级结构的影响二级结构允许简单描述蛋白质的复杂三维结构. 分子动力学软件Gromacs采用DSSP程序将通过X射线晶体学或核磁共振确定的原子坐标转换为供体和受体之间的主链氢键, 然后将重复键合模式转换为螺旋、片和匝等类别, 进一步可细分为α螺旋、卷曲、β折叠、β桥、弯曲、转角、回折等二级结构. 本文计算了在不同幅值太赫兹波作用下钾离子通道蛋白二级结构(总数 = α螺旋 + β折叠 + β桥 + 转角)的变化趋势. 如图3所示, 在经过近700 ps的稳定后, 钾离子通道蛋白的二级结构数目也趋于稳定, 从图3可以看出, 当外加太赫兹波时, 钾离子通道蛋白二级结构数目比不加太赫兹电磁波时多, 外加太赫兹电磁波会影响蛋白质二级结构的变化从而影响其功能. 并且在外加太赫兹波幅值为0.2 V/nm时, 钾离子通道蛋白二级结构的数目增加最多, 此时我们认为是钾离子通道蛋白和太赫兹波形成频率和幅值匹配才造成钾离子通道蛋白结构变化显著. 图 3 太赫兹波强度不同时钾离子通道蛋白中二级结构数目的变化 Figure3. Changes in the number of secondary structures in potassium channel proteins under different terahertz intensities.
蛋白质的二级结构又可进一步分为α螺旋、β折叠、卷曲和转角等结构. α螺旋是蛋白质肽链主链骨架借助氢键卷曲形成的一种周期性螺旋结构, α螺旋跨膜结构域的相互作用和动力学在膜蛋白的折叠、组装和功能中起着重要作用[33-35]. 氢键是α螺旋稳定的主要次级键, 当氢键破坏时, α螺旋构象即被破坏. 借助VMD, 我们发现α螺旋主要集中于S5和S6的孔道处, 即包围了钾离子通道最窄的地方. 不同幅值的太赫兹波作用下α螺旋的变化趋势如图4所示, 可以看出, 在体系平衡后, 外加太赫兹波时, 钾离子通道蛋白中α螺旋的数目要比未加太赫兹波时少, 并且外加太赫兹波的幅值增大时, 对应的α螺旋的数目也在缓慢减小. 因为太赫兹波的能量极低, 不能直接破坏共价键, 只能通过共振形式破坏氢键. 图4中的结果表明, 太赫兹波会破坏钾离子通道蛋白中α螺旋中的氢键, 使其数目减少. 图 4 太赫兹波强度不同时钾离子通道蛋白中α螺旋数目的变化 Figure4. Changes in the number of α-helices in potassium channel protein under different terahertz intensities.
β折叠是由两条以上肽链或同一条肽链之间的不同部分平行或反平行排列形成股, 股与股之间通过氢键固定, 氢键主要在股间. β折叠主要存在于S1—S4序列, 影响通道电压感应和门控. 不同幅值的太赫兹波作用下β折叠的变化趋势如图5所示, 可以看出, 外加太赫兹波时的β折叠数量比未加太赫兹波时多. 说明太赫兹波可能会促进不同肽链之间或同一条肽链之间不同部分形成新的β折叠, 也有可能是太赫兹波导致部分α螺旋展开形成β折叠. 从图4和图5可知, 太赫兹波造成α螺旋减少, β折叠增多. β折叠往往含有较多的非极性残基, 并埋在蛋白质内部形成疏水核心, 而α螺旋通常是两性的, 亲水面位于表面, 疏水一侧朝向蛋白质内部. α螺旋的减少和β折叠的增多会导致钾离子通道中疏水核的暴露和亲水面的减少, 从而影响钾离子通过速率. 图 5 太赫兹波强度不同时钾离子通道蛋白中β折叠数目的变化 Figure5. Changes in the number of β-sheet in potassium channel protein under different terahertz intensities.
卷曲通过主链间的氢键或主链与侧链间的氢键稳定其构象, 其是柔性的, 可以使肽链改变走向, 且有利于连接结构相对刚性的α螺旋和β折叠. 从图6可以看出, 外加太赫兹波时钾离子通道蛋白中的卷曲数目减少, 并且在外加太赫兹波幅值为0.2 V/nm时, 卷曲的数目变化最大, 此时蛋白质的构象变化可能最大. 图 6 太赫兹波强度不同时钾离子通道蛋白中卷曲数目的变化 Figure6. Changes in the number of coil in potassium channel protein under different terahertz intensities.
5.不同强度太赫兹波对离子通过速率的影响钾离子通过钾通道的速率对于调节神经细胞的静息电位和动作电位至关重要, 并且钾离子通道对一些疾病起着调节作用, 是治疗的新靶点. 本文分别计算了无太赫兹电场、太赫兹电场强度为0.1, 0.2, 0.3, 0.4, 0.5 V/nm情况下钾离子通过电压门控钾离子通道的速率, 从图7可知, 最开始随着外加太赫兹波幅值的增大, 钾离子通过钾离子通道的速率也随之增大, 但是当幅值增加到0.3 V/nm及以上时, 钾离子通过的速率不再增加, 我们推测已经达到了钾离子通道的饱和, 这是由于孔道蛋白分子不是电中性的, 其对通过的钾离子具有一定的电吸引, 限定了它的最大运输速度. 并且从图4—图6可知, 当外加太赫兹波的幅值为0.2 V/nm时, 对应的二级结构变化最大, 但是此时的钾离子通过速率却没有达到最大. 静电分析表明, 当钾离子沿着狭窄的孔道穿过膜时它必须穿过一个能量闸, 太赫兹波除了和钾离子通道蛋白中的羰基形成共振外, 还对模拟盒子中的钾离子有库仑力的作用, 两者共同作用使钾离子快速通过钾离子通道. 图 7 太赫兹波强度不同时钾离子通过钾离子通道的数目 Figure7. Number of potassium ions passing through potassium channels under different intensities of terahertz.
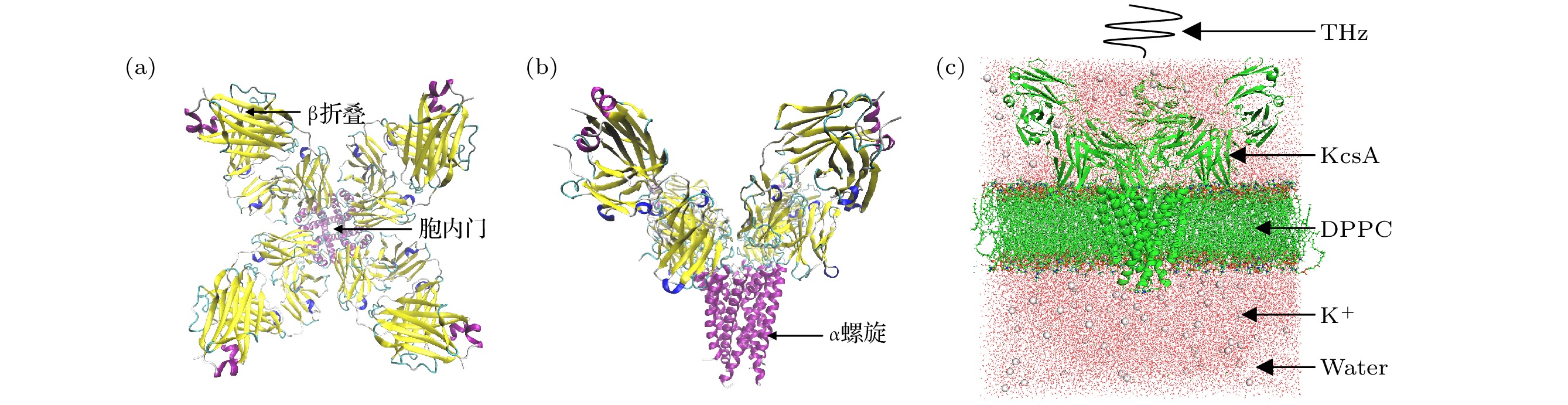 图 1 (a) KcsA四聚体结构(从上往下); (b) KcsA四聚体结构(从前往后); (c)钾离子通道模型
图 1 (a) KcsA四聚体结构(从上往下); (b) KcsA四聚体结构(从前往后); (c)钾离子通道模型


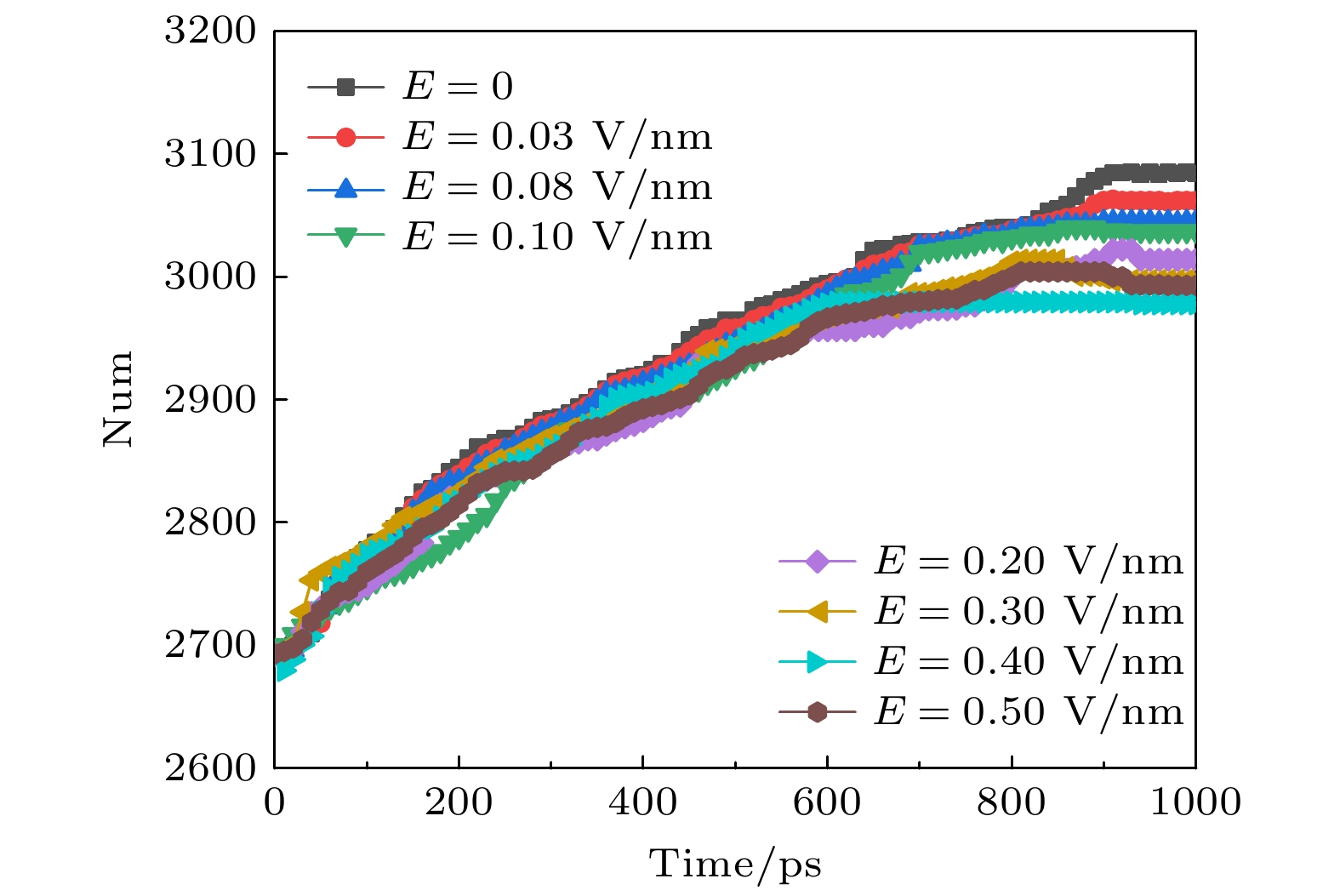 图 2 太赫兹波强度不同时钾离子通道蛋白中氢键的变化
图 2 太赫兹波强度不同时钾离子通道蛋白中氢键的变化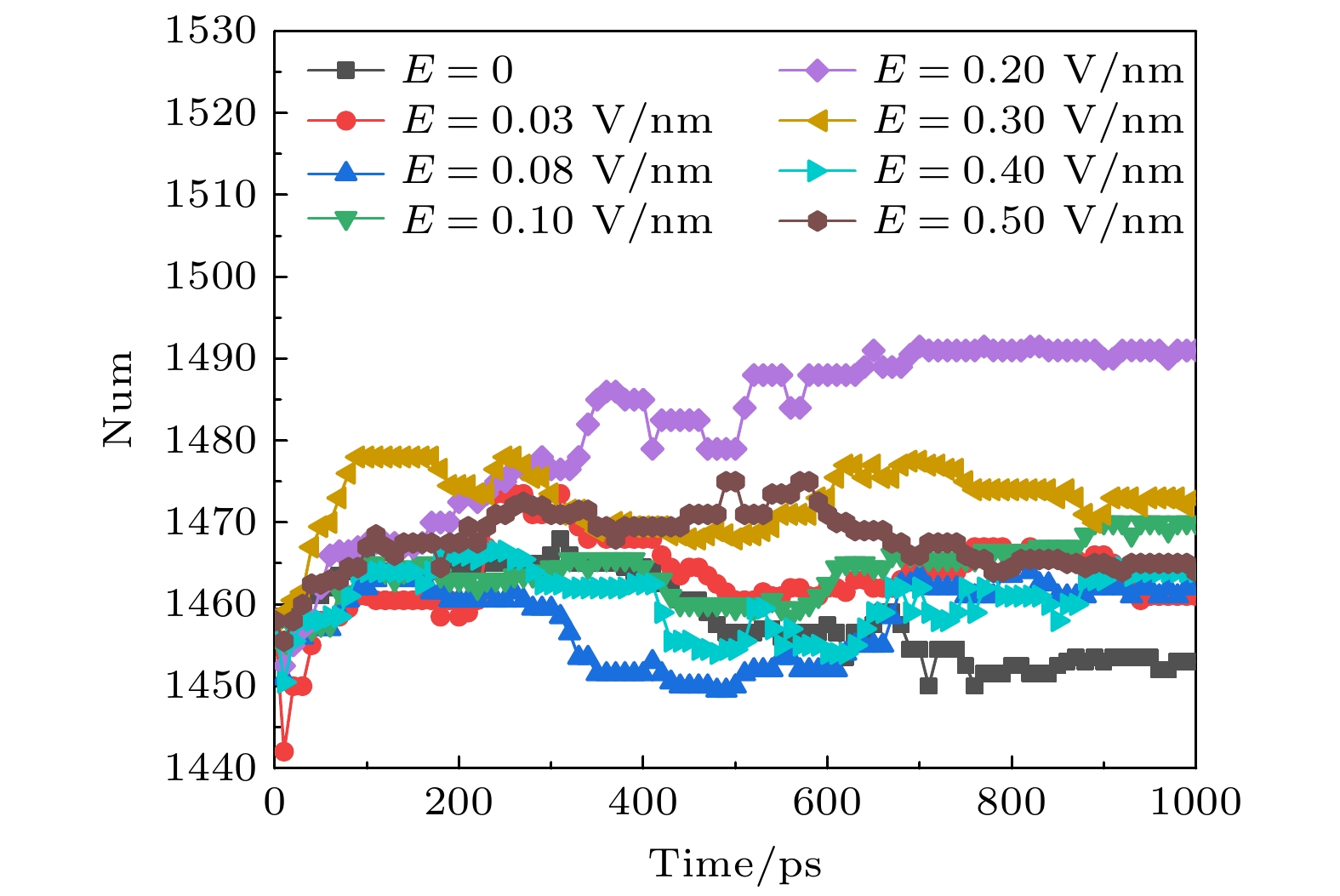 图 3 太赫兹波强度不同时钾离子通道蛋白中二级结构数目的变化
图 3 太赫兹波强度不同时钾离子通道蛋白中二级结构数目的变化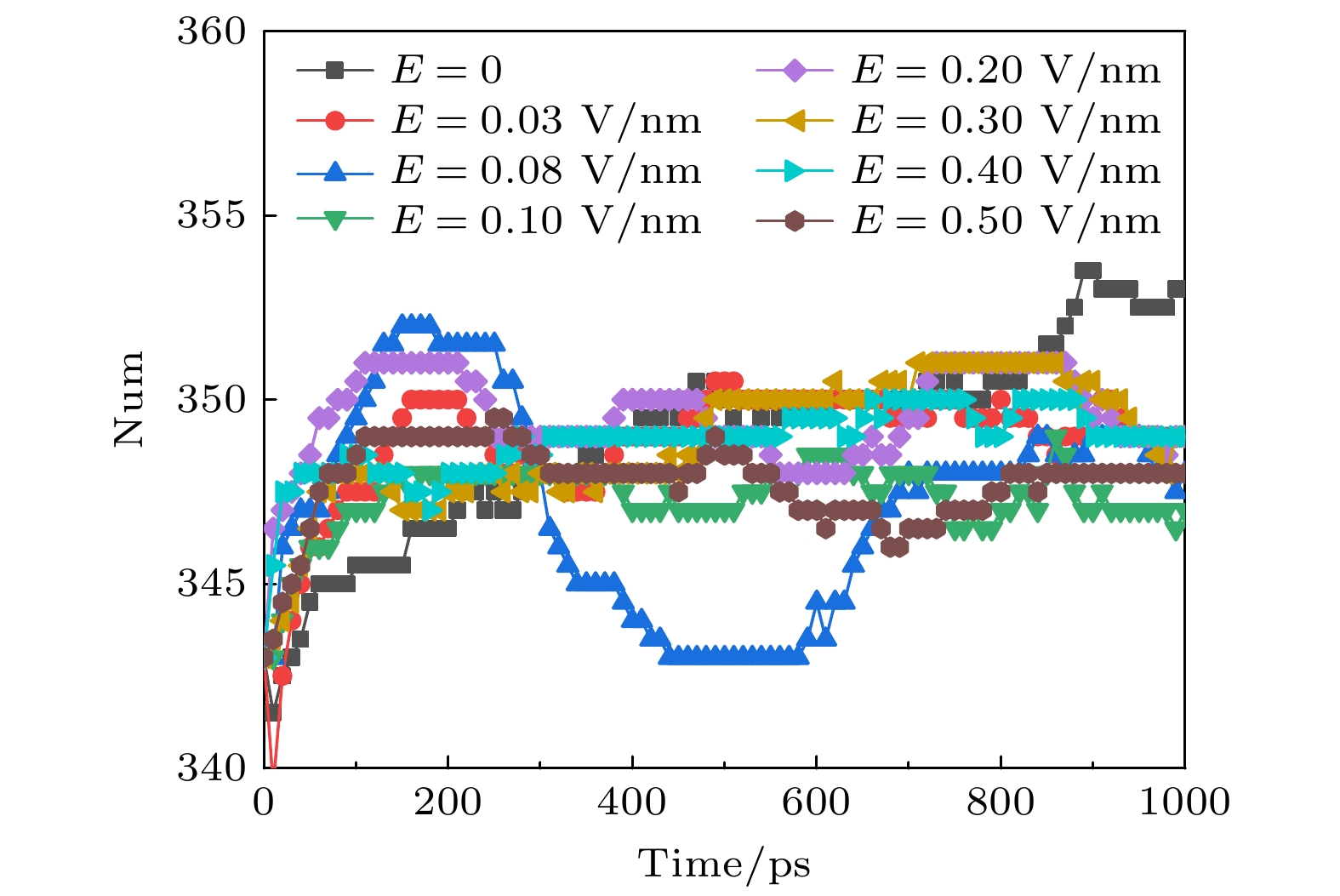 图 4 太赫兹波强度不同时钾离子通道蛋白中α螺旋数目的变化
图 4 太赫兹波强度不同时钾离子通道蛋白中α螺旋数目的变化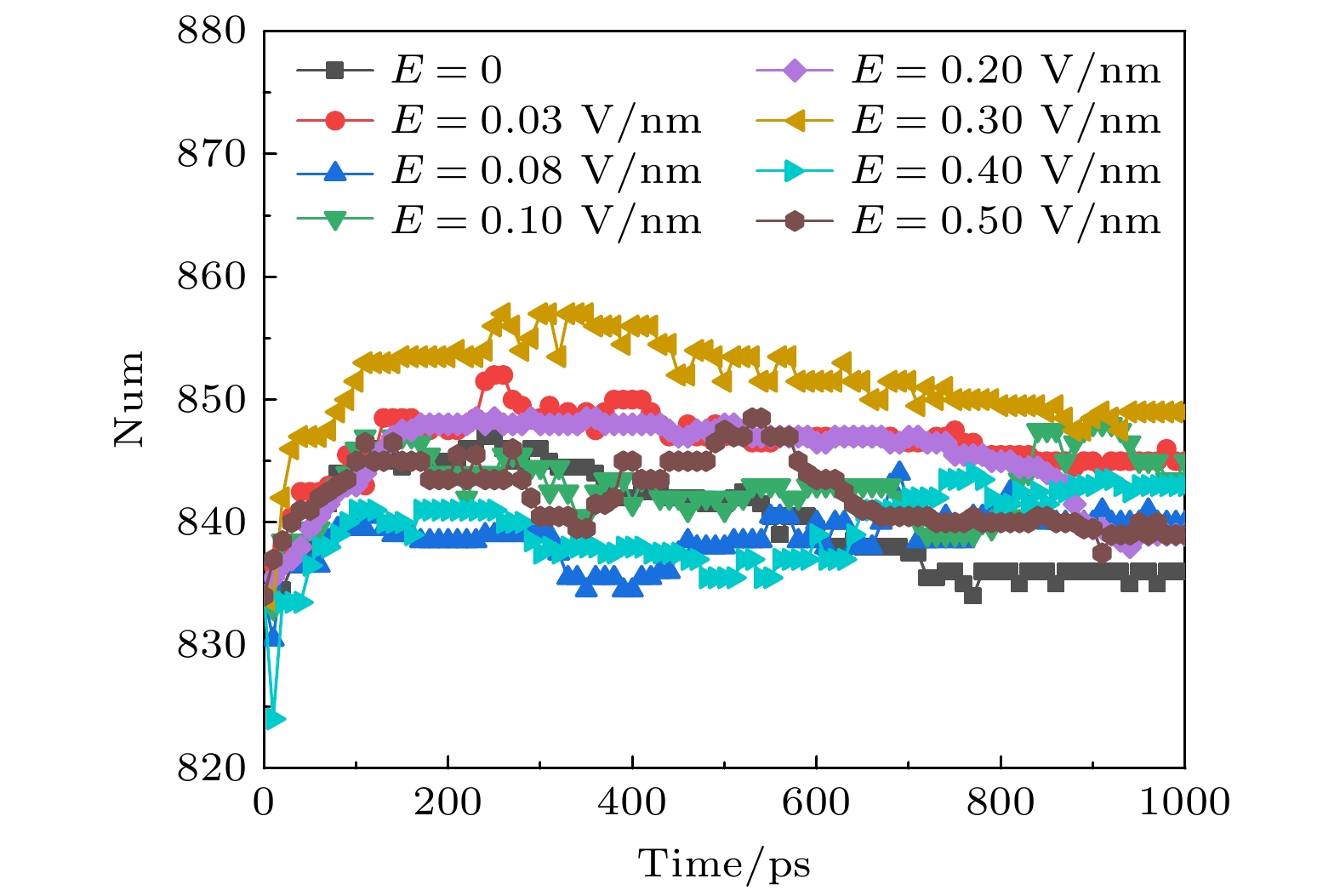 图 5 太赫兹波强度不同时钾离子通道蛋白中β折叠数目的变化
图 5 太赫兹波强度不同时钾离子通道蛋白中β折叠数目的变化 图 6 太赫兹波强度不同时钾离子通道蛋白中卷曲数目的变化
图 6 太赫兹波强度不同时钾离子通道蛋白中卷曲数目的变化 图 7 太赫兹波强度不同时钾离子通过钾离子通道的数目
图 7 太赫兹波强度不同时钾离子通过钾离子通道的数目