全文HTML
--> --> -->最近的研究表明, 水的动力学过程与太赫兹之间有着密切的关系. 太赫兹(terahertz, THz) 波的波段能够覆盖生物大分子、有机体、半导体和等离子体等物质的特征谱, 利用该频段可以加深和拓展人类对物理学、化学、天文学、信息学和生命科学中一些基本科学问题的认识[20-24]. 在生物医学上, 生物大分子相互作用是重大生命现象与病变产生的关键动因, 然而太赫兹光子的能量覆盖了生物大分子空间构象的能级范围 (如图1(a) 所示), 该频段包含了其他电磁波段无法探测到的直接代表生物大分子功能的空间构象等重要信息[25]. 2018年, 刘国治院士[26]推断生物神经信号物理场应为太赫兹到红外区域的高频电磁场, 最可能频率范围应在0.5—100 THz, 并且将此波段称为广义太赫兹. 这预示着广义太赫兹波有望调控生物分子的结构和功能. 生物分子在水溶液中发挥活性和功能, 然而水是极性液体, 它在部分太赫兹频率下有异常高的吸收损失, 表现出热效应, 给太赫兹技术在生物体系中的应用蒙上了一层乌云[27]. 通常来说, 水的太赫兹光谱有3个主峰 (图1(b)), 第1个峰位于5—30 THz区间, 对应水中氢键网络的振动模式(图1(c)), 第2个峰位于45—50 THz, 对应水分子内部键角的弯曲振动 (图1(c)), 第3个峰则位于90—105 THz, 它对应于水分子内部键长的拉伸振动 (图1(c)). 这意味着在广义太赫兹波段存在4个水的太赫兹弱吸收窗口, 可用于非热地调控生物大分子的结构和功能. 得益于此, 近3年, 太赫兹在水溶液中非热地调控细胞动力学过程取得重要的进展[28-35] (图1(d)). 2019年, 我们提出1.39或4.66 THz的电磁刺激能够非热地引发一维水通道内受限水的超级渗透, 促使受限水由一维冰相向一维相干气相转变[28]. 2020年, Wu等[29]发现44.0 THz的电磁刺激能够共振地加速DNA分子的解旋过程, 并且有效地降低DNA的熔解温度. 同年, Wang等[30]观察到太赫兹辐射能够引起DNA碱基对中氢键的瞬态质子转移. Li等[31]证实细胞内ATP水解能够释放频率约为34 THz的内源光子, 进而调控生化反应. 2021年, Li等[32]发现42.55 THz的电磁刺激能够非热地加速钙离子通道内钙离子的渗透. 同年, Liu等[33]演示了53.7 THz的电磁刺激能够非热、可逆地调控神经信号和动物的行为. 以上研究表明, 为了避免太赫兹电磁波被体相水强吸收, 理论上预测既能够调控生物大分子的功能, 又对体相水产生有限热效应的太赫兹波段非常关键, 因此, 有必要深入地对体相水的太赫兹吸收谱进行理论研究.
 图 1 太赫兹与生物分子的密切关系以及太赫兹调控细胞动力学 (a) 生物大分子的转/振动的频率在THz频段; (b) 水的太赫兹吸收谱, 绿色区域是有望非热地调控生物分子的广义太赫兹频率的4个窗口; (c) 水的太赫兹吸收谱的振动模式[36]; (d) 太赫兹波非热调控细胞动力学, 涉及发挥细胞生物功能的水通道蛋白、DNA、钾离子通道、钙离子通道
图 1 太赫兹与生物分子的密切关系以及太赫兹调控细胞动力学 (a) 生物大分子的转/振动的频率在THz频段; (b) 水的太赫兹吸收谱, 绿色区域是有望非热地调控生物分子的广义太赫兹频率的4个窗口; (c) 水的太赫兹吸收谱的振动模式[36]; (d) 太赫兹波非热调控细胞动力学, 涉及发挥细胞生物功能的水通道蛋白、DNA、钾离子通道、钙离子通道Figure1. Close relationship between terahertz and biomolecules and the regulation of cell dynamics by terahertz: (a) Frequency of rotation/vibration of biological macromolecules is in the THz frequency band; (b) terahertz absorption spectrum of water, the green region is the four frequency windows in which electromagnetic wave is expected to non-thermally regulate biomolecules; (c) vibration modes of water corresponding to its terahertz absorption spectrum[36]; (d) terahertz waves non-thermally regulate the dynamics of a cell, involving aquaporins, DNA, potassium and calcium channels that perform biological functions of the cell.
本文利用分子动力学模拟方法基于不同水模型和温度, 研究了水的太赫兹吸收谱以及与氢键网络动力学的关系, 发现水形成的氢键网络越强, 其太赫兹吸收峰越倾向于蓝移, 进一步发现了水的氢键寿命与其太赫兹吸收峰的频率之间存在线性关系, 并利用弹簧振子模型对这一现象的机理加以解释, 也就是将水的氢键等同于弹簧, 如果水的氢键越强, 其等效弹簧的劲度系数就越大, 进而使得氢键网络具有较大的振动频率.
2.1.模拟方法
首先, 建立大小为4 nm × 4 nm × 4 nm的立方体模拟盒子, 然后在常温常压的情况下分别用水分子个数为2165的柔性SPC, SPC/E, TIP3P水分子模型和水分子个数为2211的柔性的TIP4P, TIP4P-Ew水分子模型将模拟盒子填充满. 水分子间的相互作用为




| Water model | σO—O/? | εOO/(kJ·mol–1) | qH/e | qO or qvir/e | θ/(°) | rH—O/? |
| SPC | 3.166 | 0.650 | 0.410 | –0.820 | 109.470 | 1.000 |
| SPC/E | 3.166 | 0.650 | 0.424 | –0.848 | 109.470 | 1.000 |
| TIP3P | 3.151 | 0.636 | 0.417 | –0.834 | 104.520 | 0.957 |
| TIP4P | 3.154 | 0.649 | 0.520 | –1.040 | 104.520 | 0.957 |
| TIP4P-Ew | 3.164 | 0.681 | 0.524 | –1.048 | 104.520 | 0.957 |
表1所用水模型的力场参数的比较
Table1.Comparison of the force field parameters of different water models employed.
2
2.2.分析方法
32.2.1.频谱解析
在经典近似的情况下, 分子与频率相关的光谱强度(I?)可以通过对水的总电荷流的自相关函数进行傅里叶变换获得, 它的表达式为
3
2.2.2.径向分布函数
径向分布函数表示了与某个原子距离为r的单位体积元内出现的分子数密度, 表达式为[39,40]
3
2.2.3.自扩散系数
根据Einstein关系, 自扩散系数与粒子的均方根位移(MSD)成正比, 表达式为[41]
3
2.2.4.氢键寿命
通常氢键采用纯几何准则进行定义: 若两个水分子的氧原子 (O···O) 间的距离小于0.35 nm, O···O—H之间的夹角小于30°, 则认为两水分子之间形成氢键. 氢键的自相关函数为[42-44]



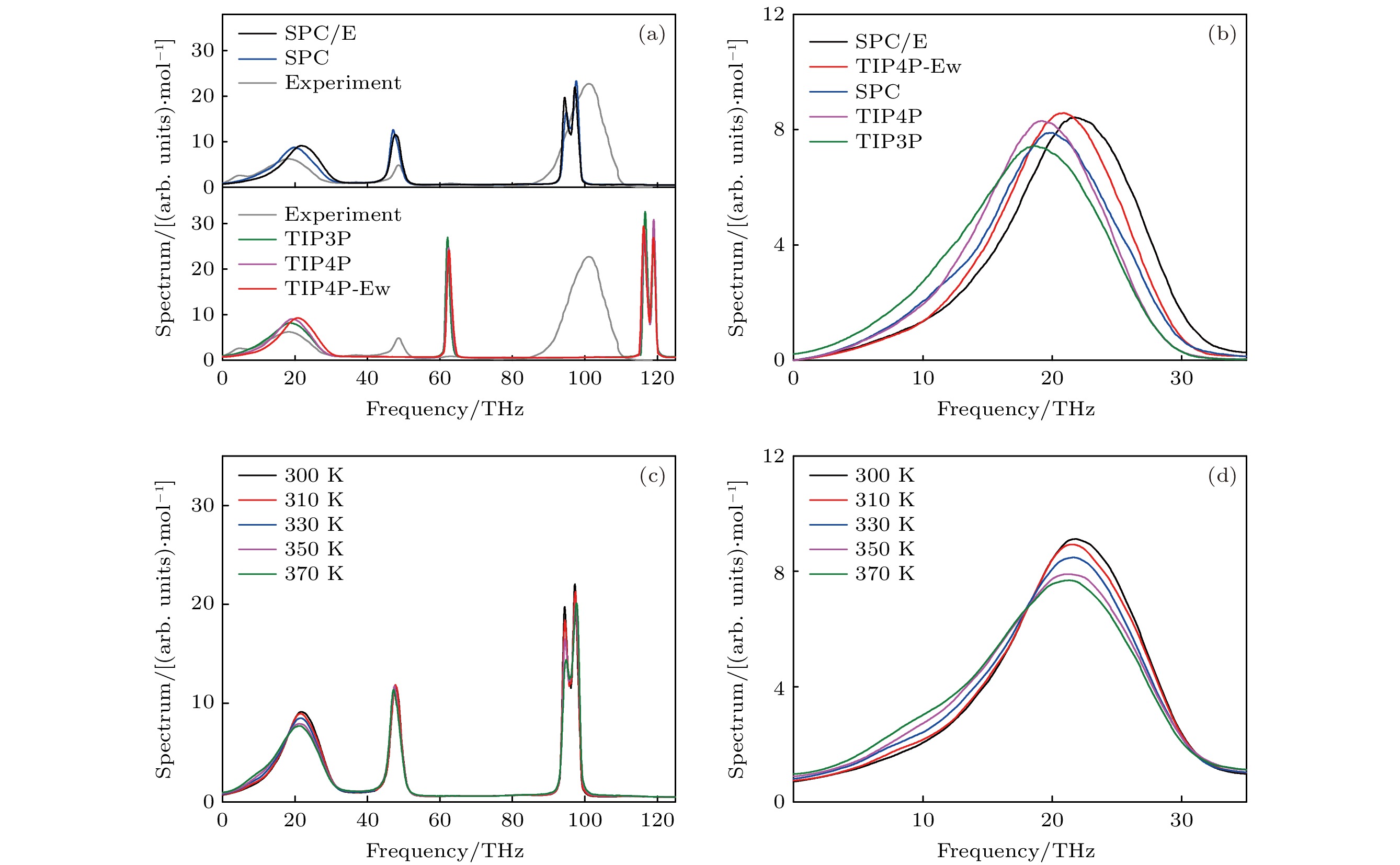 图 2 不同水模型建模的体相水的太赫兹吸收谱和温度对其太赫兹吸收谱的影响 (a) 不同水模型建模的体相水的太赫兹吸收光谱以及实验测量所得体相水的太赫兹吸收谱之间的对比; (b) 对于不同水模型构成的体相水, 其氢键网络的太赫兹吸收谱; (c) 不同温度下, SPC/E水模型建模的体相水的太赫兹吸收谱; (d) 不同温度下, 对于SPC/E水模型构成的体相水, 其氢键网络的太赫兹吸收谱
图 2 不同水模型建模的体相水的太赫兹吸收谱和温度对其太赫兹吸收谱的影响 (a) 不同水模型建模的体相水的太赫兹吸收光谱以及实验测量所得体相水的太赫兹吸收谱之间的对比; (b) 对于不同水模型构成的体相水, 其氢键网络的太赫兹吸收谱; (c) 不同温度下, SPC/E水模型建模的体相水的太赫兹吸收谱; (d) 不同温度下, 对于SPC/E水模型构成的体相水, 其氢键网络的太赫兹吸收谱Figure2. THz absorption spectra of bulk water modeled by different water models and effects of temperature on its spectra: (a) Comparison of THz absorption spectra of bulk water under different water models and its spectra from experimental measurement; (b) THz absorption spectra of hydrogen bond network of bulk water under different water models; (c) THz absorption spectra of bulk water modeled by SPC/E water model at different temperatures; (d) THz absorption spectra of hydrogen bond network of bulk water modeled by SPC/E water model at different temperatures.
因为温度会改变氢键网络的结构, 进而影响氢键网络的相互作用. 较高的温度更利于破坏氢键网络, 降低氢键的相互作用. 为了研究氢键网络的振动吸收峰的中心频率与水的相互作用之间的关系, 选择特定的水模型(SPC/E), 进一步计算了体相水在不同温度下的太赫兹吸收谱. 从图2(c)可知, 温度对水的氢键网络的振动吸收峰有显著的影响, 然而对水的弯曲和拉伸振动的吸收峰的影响有限. 特别地, 随着温度从300 K逐渐升高到370 K, 水的氢键网络振动的吸收峰的中心频率逐步发生了红移(如图2(d) 所示), 也就是300 K下为(21.83 ± 0.04) THz, 310 K下为(21.73 ± 0.05) THz, 330 K下为(21.39 ± 0.05) THz, 350 K下为(21.03 ± 0.06) THz, 370 K下为(20.81 ± 0.07) THz. 因此, 可以初步得出结论: 水具有越强的氢键, 其氢键网络振动的吸收峰的中心频率越大.
为了进一步研究水的氢键网络强弱与其振动吸收峰的中心频率之间的关系, 分析了体相水的结构和动力学性质(图3). 首先, 计算了由不同水模型构成的体相水的径向分布函数g(r), 如图3(a)所示. 可以看出, 随着r的增加, g(r)函数值逐渐减小并收敛到1.0, 这说明水分子表现出短程有序、长程无序的微观结构特征, 符合液体分子的排列规则. 此外, 由SPC/E, TIP4P和TIP4P-Ew构成的体相水的g(r)具有两个峰, 跟实验结果一致[48], 但由SPC, TIP3P构成的体相水的g(r)只有1个峰. 特别地, 由不同水模型构成的体相水的第一水合层的密度是不一样的, 具体表现为g(r)的第1个峰的峰值高度的差异, 其峰值高度的大小关系依次为TIP3P (2.86) < TIP4P (3.13) < SPC (3.18) < TIP4P-Ew (3.43) < SPC/E(3.51); 然而, 由不同水模型构成的体相水的g(r)的第1个峰值所处的位置(r值)具有相反的关系: TIP3P (0.274 nm) > TIP4P (0.273 nm) = SPC (0.273 nm) > TIP4P-Ew (0.272 nm) > SPC/E (0.271 nm). 值得注意的是, g(r)的第1个峰值高度值越大以及g(r)的第1个峰值所处的r值越小, 水的氢键网络振动的太赫兹吸收峰的中心频率越大. 这归因于水分子与周围的第一水合层内的水分子形成氢键, g(r)的第1个峰的峰值高度越大以及它的r越小, 说明水分子与第一水合层内的水分子结合得越紧密, 从而具有越强的氢键相互作用, 对周围水分子的束缚也越强. 粒子的扩散系数能够说明周围粒子对其的束缚情况. 然后, 计算了由不同水模型构成的体相水的扩散系数D, 如图3(b)所示, 各D值的大小关系为TIP3P > TIP4P ≈ SPC > TIP4P-Ew > SPC/E, D的值依次是(3.97 ± 0.09) ×10–5, (2.36 ± 0.04) × 10–5, (2.34 ± 0.07) × 10–5, (1.25 ± 0.07) × 10–5和 (1.12 ± 0.01) × 10–5 cm2/s. 将各D值的大小关系与其氢键网络振动的吸收峰的中心频率的大小关系联系起来, 发现水的氢键对水的束缚越强 (D值越小), 水的氢键网络振动的太赫兹吸收峰的中心频率越大. 同样地, 基于SPC/E水模型, 计算了温度对体相水g(r)和D的影响. 如图3(c) 所示, 随着温度的升高, g(r)的第1个峰值高度不断减小(300 K下为3.51, 310 K下为3.39, 330 K下为3.20, 350 K下为3.01, 370 K下为2.88), 它的第1个谷值却不断升高, 以及第2个峰的峰值也逐步降低甚至消失, 这表明水的长程有序性逐渐减小. 不仅如此, 随着温度升高, 体相水的D逐渐增大, 如图3(d) 所示, 表现为: D的值在300 K时为(1.12 ± 0.01) × 10–5 cm2/s, 310 K时为 (1.54 ± 0.07) × 10–5 cm2/s, 330 K时为 (2.47 ± 0.07) × 10–5 cm2/s, 350 K时为 (3.54 ± 0.03) × 10–5 cm2/s, 370 K时为 (4.80 ± 0.12) × 10–5 cm2/s. 这是因为温度升高使得水分子的运动加剧, 削弱了水分子之间的氢键. 以上结果表明: 水分子具有越紧束缚的氢键网络, 其氢键网络的振动吸收峰的中心频率越大.
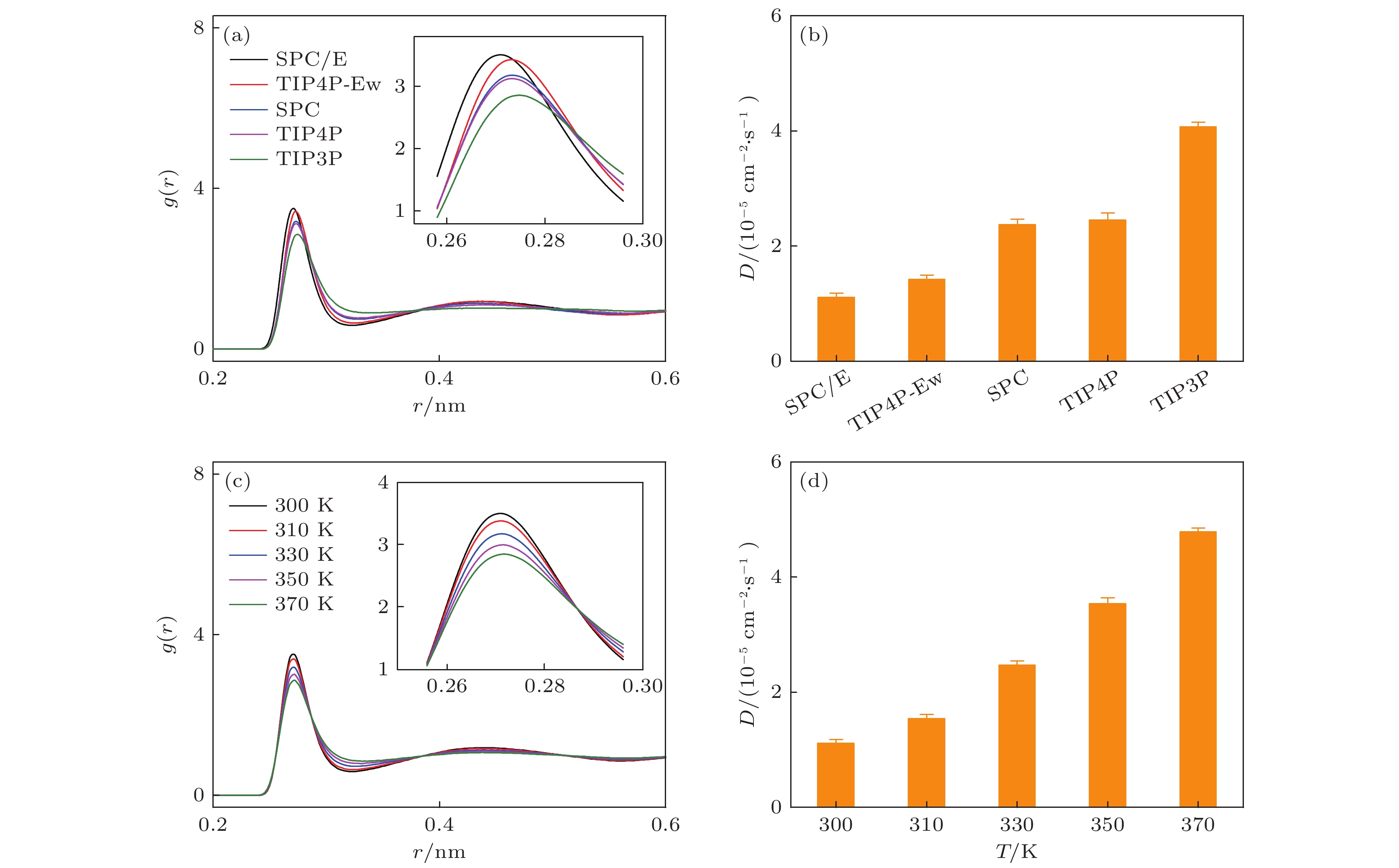 图 3 不同水模型构成的体相水的结构和动力学性质的分析 (a) 不同水模型构成的体相水的径向分布函数; (b) 不同水模型构成的体相水的扩散系数; (c) 特定的水模型(SPC/E)下, 温度对体相水的径向分布函数的影响; (d) 不同温度下SPC/E的扩散系数
图 3 不同水模型构成的体相水的结构和动力学性质的分析 (a) 不同水模型构成的体相水的径向分布函数; (b) 不同水模型构成的体相水的扩散系数; (c) 特定的水模型(SPC/E)下, 温度对体相水的径向分布函数的影响; (d) 不同温度下SPC/E的扩散系数Figure3. Analyses of structure and dynamic behavior of bulk water modeled by different water models: (a) Radial distribution function of bulk water under different water models; (b) diffusion coefficient of bulk water under different water models; (c) under specific water model (SPC/E), the effect of temperature on the radial distribution function of bulk water; (d) diffusion coefficient of bulk water at different temperatures.
最后, 定量研究水的氢键强弱与其振动吸收峰的中心频率之间的关系(如图4所示). 水分子之间的氢键处于动态变化, 它可能在某一时刻断裂, 随后又与其他水分子形成氢键, 氢键动态变化导致体系总偶极距的变化, 因此, 根据涨落耗散定理[49], 氢键动态变化的快慢将直接影响氢键网络振动的太赫兹吸收谱. 氢键的平均寿命能够衡量氢键动态变化的快慢, 它由氢键的自相关函数C(t)拟合得到. 如图4(a)所示, 不同水模型的C(t)都近似于指数衰减, 但是它们衰减的快慢不一样, 表明氢键寿命大小不一样. 从图4(b)可知, SPC/E, TIP4P-Ew, SPC, TIP4P, TIP3P的氢键寿命依次为(7.09 ± 0.03), (5.46 ± 0.02), (3.23 ± 0.02), (3.11 ± 0.02)和(1.66 ± 0.01) ps. 将由不同水模型构成的体相水的氢键网络振动的吸收谱的中心频率v与它们的氢键寿命 τ联系起来 (如图4(c)所示), 发现它们之间呈线性关系



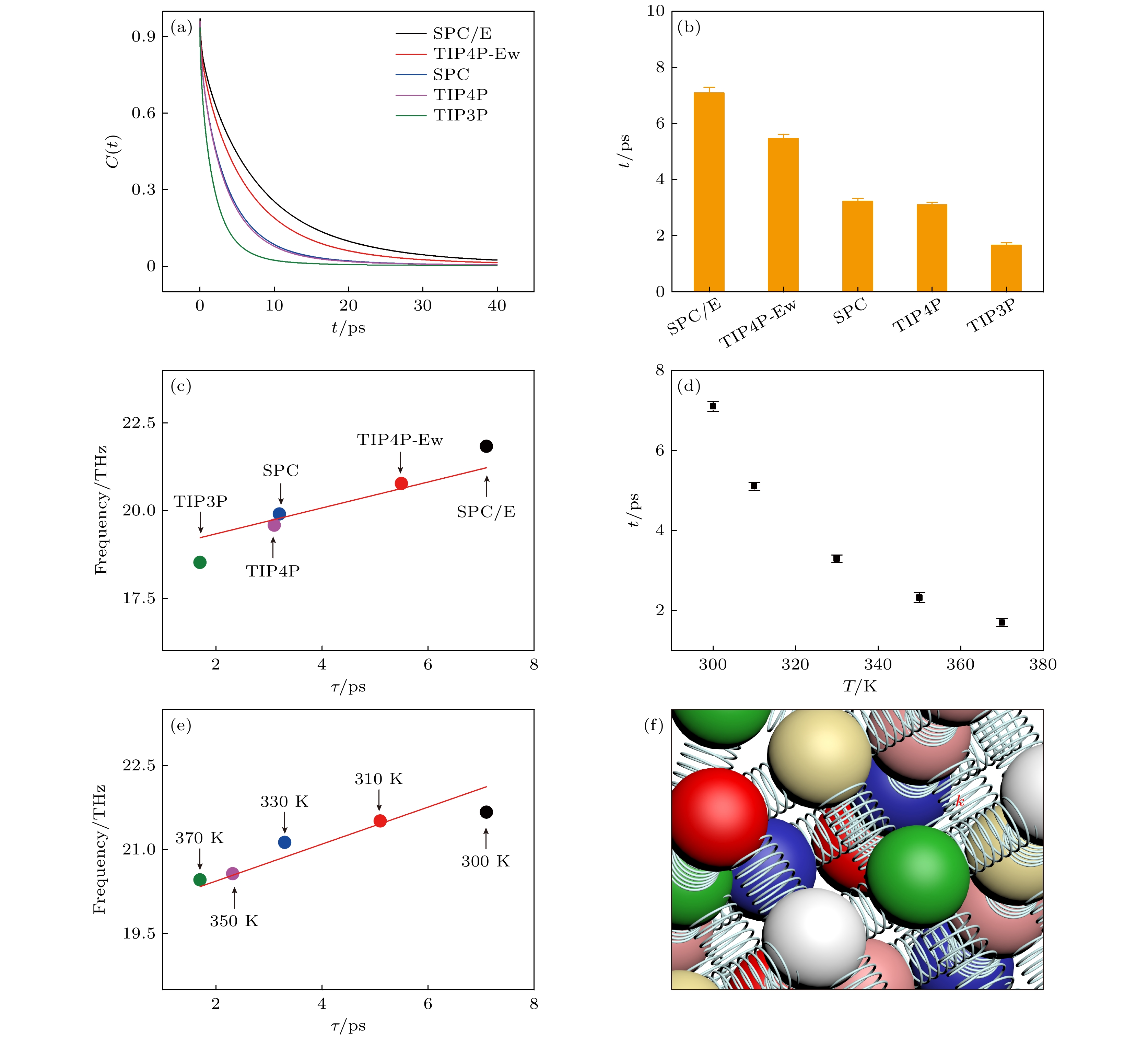 图 4 氢键动力学与氢键网络振动的太赫兹吸收峰的中心频率之间的关系 (a) 由不同水模型构成的体相水的氢键的自相关函数; (b) 由不同水模型构成的体相水的氢键寿命; (c) 不同水模型下, 氢键网络振动的太赫兹吸收谱的中心频率与氢键寿命的对应关系; (d) 不同温度下, 体相水的氢键寿命; (e) 不同温度下, 水的氢键网络振动的太赫兹吸收峰的中心频率与氢键寿命的关系; (f) 氢键网络示意图
图 4 氢键动力学与氢键网络振动的太赫兹吸收峰的中心频率之间的关系 (a) 由不同水模型构成的体相水的氢键的自相关函数; (b) 由不同水模型构成的体相水的氢键寿命; (c) 不同水模型下, 氢键网络振动的太赫兹吸收谱的中心频率与氢键寿命的对应关系; (d) 不同温度下, 体相水的氢键寿命; (e) 不同温度下, 水的氢键网络振动的太赫兹吸收峰的中心频率与氢键寿命的关系; (f) 氢键网络示意图Figure4. Relationship between hydrogen bond dynamics and the center frequency of THz absorption spectra for the vibration of hydrogen bond network: (a) Hydrogen bond autocorrelation functions of bulk water under different water models; (b) lifetime of hydrogen bond of bulk water under different water models; (c) for bulk water under different water models, the relationship between the center frequency of THz absorption spectra for the vibration of hydrogen bond network and the lifetime of hydrogen bond; (d) lifetime of hydrogen bond for bulk water at different temperatures; (e) at different temperatures, the relationship between the center frequency of THz absorption spectra for the vibration of the hydrogen bond network and the lifetime of the hydrogen bond; (f) schematic diagram of the hydrogen bond network.
