全文HTML
--> --> -->近年来, ACSM作为下一代制造技术的主要发展趋势, 受到了科学界与产业界的广泛关注. 许多国家相继启动了“原子制造”相关重大国家战略性研究计划, 旨在科技竞争中取得战略性先机[1,2]. 虽然ACSM技术正处于研究起步阶段, 其广泛的应用前景及将带来的技术性革命, 推动着这一领域的快速发展. 从ACSM技术发展现状来看, 原子制造技术主要包括以下三大类: 1) 基于高能粒子束的原子级去除或原子刻蚀; 2) 基于高分辨显微镜的单原子操纵技术; 3) 基于化学方法的可控原子材料合成. 从原子制造的概念及要求可知, 原子制造的核心在于实现原子在三维空间内的可控排布. 高能粒子束原子刻蚀技术具备规模化生产的潜能, 然而其精度有待进一步提高, 且制造成本较高. 基于高分辨显微镜的原子操纵技术其精度可达单个原子级别, 然而较难实现规模化. 基于化学方法的原子制造技术(金属纳米团簇的合成), 其合成均一性和可控性均有待进一步提高. 因此, 开发新型制造技术是ACSM领域的迫切需求. 核酸作为自然界中最常见的原子级精密组装体, 由于其结构特征及核酸分子间特殊的相互作用模式, 使其从生命科学领域跨越至新型材料领域[3,4]. DNA纳米技术的快速发展, 在材料可控组装、合成领域体现出巨大优势, 有望为ACSM技术的发展提供全新的方法.
核酸是由许多核苷酸单体聚合形成的生物大分子化合物, 是生命遗传物质的主要载体. 由磷酸骨架、核糖和含氮碱基三部分基本单元组成, 其电负性磷酸骨架及其杂环碱基上富含具有孤电子对的氮原子等性质, 使其成为金属离子结合的天然靶标[5,6]. 其次, 通过体外筛选技术获得了一系列可结合金属原子的功能核酸分子, 如金属离子特异性核酸适体(aptamer)、脱氧核酶(DNAzyme)等, 并构建了一系列基于核酸材料的分子器件, 包括金属离子传感器[7,8]、光响应金属纳米簇探针[9-11]、金属纳米器件[12]、含金属抗癌药物等[13,14]. 此外, 非天然人工碱基的合成进一步拓展了核酸材料的功能, 含金属非天然人工碱基的开发为核酸-金属离子相互作用提供了一种全新的策略, 实现了金属离子在核酸中更加精确的定位和组装.
本综述从核酸分子的基本结构与功能出发, 分析了核酸材料与金属离子的作用机制; 随后根据作用机制差异分类论述了核酸-金属相互作用的原理和经典实例, 包括核酸与金属离子和金属离子配合物相互作用、核酸介导的金属纳米簇的生长调控、含金属人工碱基单体的开发与应用等; 接下来详细介绍了DNA纳米技术的发展及其在原子制造领域的优势与潜力; 最后总结了核酸材料在原子制造领域存在的关键技术瓶颈, 并对其发展前景做了进一步展望.
2.1.核酸的基本结构
核酸是一类生物大分子聚合物, 其基本结构单元是由含氮碱基、五碳糖和磷酸基团组成的核苷酸, 天然含氮碱基包括腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)、胞嘧啶(C)、尿嘧啶(U)五种, 分子结构式如图1(a)所示. 核酸的结构通常可分为一级结构、二级结构和三级结构. 核苷酸单体通过磷酸二酯键缩合形成寡聚核苷酸序列, 核苷酸在分子内的排列顺序, 即碱基的排列顺序构成了核酸分子的一级结构. 1953年美国科学家Watson和Crick共同提出了DNA分子双螺旋二级结构模型, 其分子结构具有如下特点[15,16]: 1) 双链DNA分子(dsDNA)由两条脱氧核苷酸长链反向平行盘旋成稳定的双螺旋结构(图1(b)); 2) dsDNA分子中脱氧核糖和磷酸交替连接排列在外侧, 碱基排列在内侧; 3) 嘌呤和嘧啶之间遵循Watson-Crick碱基互补配对原则形成A-T和G-C两种配对形式, 其中A与T以两对氢键相连, 而G与C以三对氢键相连(图1(c)). 值得注意的是, 单链核酸分子也可通过分子内部碱基间互补配对等相互作用形成茎、环等二级结构, 从而形成具有特定功能的“结构域”. 此外, DNA双螺旋进一步扭曲盘绕可形成具有特定三维空间结构、构象的三级结构.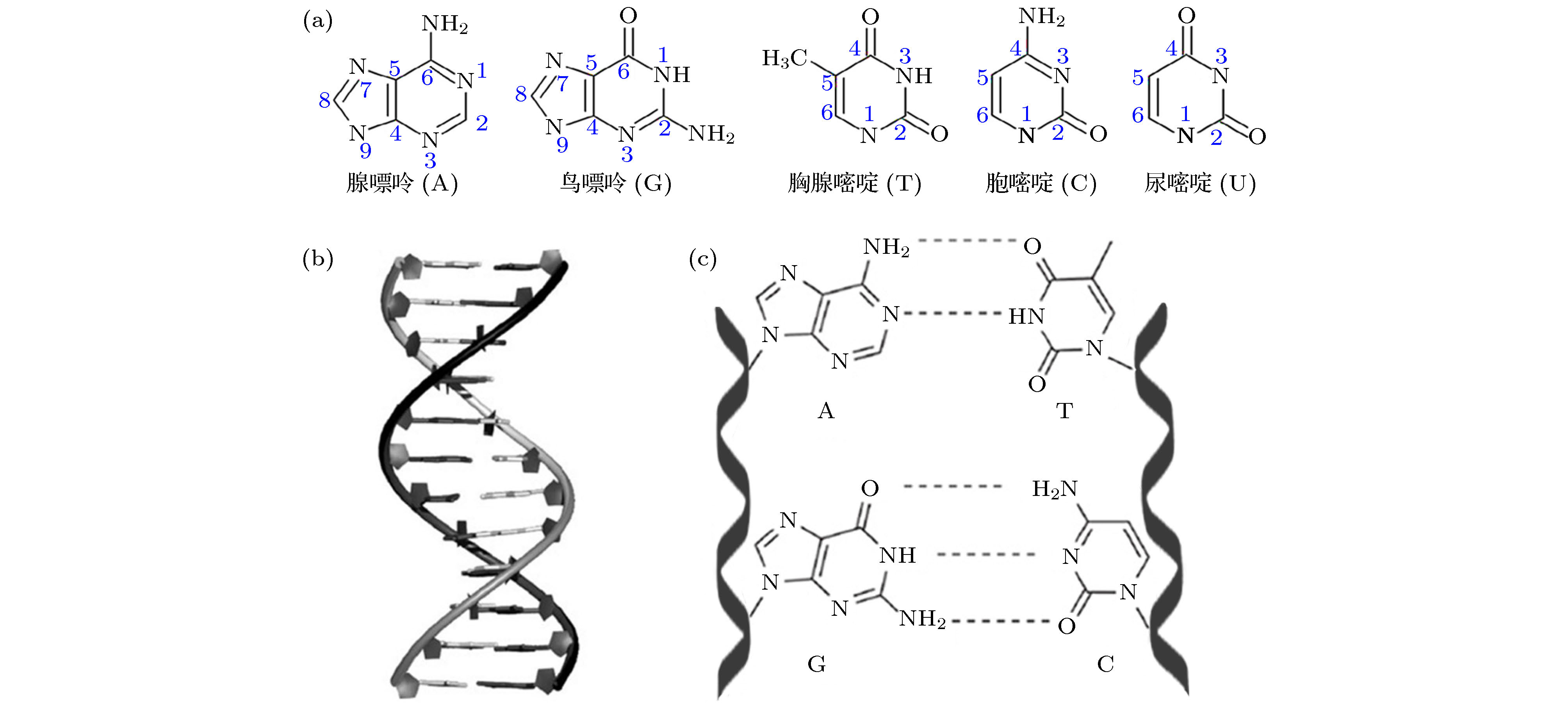 图 1 (a) 5种天然核酸碱基单体结构; (b) B-DNA双螺旋结构模型; (c) 通过氢键相连的DNA碱基互补配对形式
图 1 (a) 5种天然核酸碱基单体结构; (b) B-DNA双螺旋结构模型; (c) 通过氢键相连的DNA碱基互补配对形式Figure1. (a) Structure of five natural nucleic acid bases; (b) structure model of B-DNA; (c) DNA base pairing through hydrogen bonds.
2
2.2.核酸与金属离子相互作用的机制
在生理条件下, 核酸表现出聚阴离子性质, 其电负性磷酸骨架以及杂环碱基上的氮原子富含孤对电子等性质, 为金属离子提供了天然的结合位点[4,5]. 如图2所示, 金属离子一般采用下列两种方式与DNA作用[17,18]: 1) 金属直接与磷酸基团、糖环的氧原子以及碱基杂环上的原子(N, C, O)作用; 2) 通过金属配体的间接相互作用, 如通过金属配体分子与碱基间的氢键作用、π-π堆积作用和疏水相互作用等方式实现金属离子与核酸的连接. X射线晶体结构解析结果显示, 在核酸二级结构中, 碱金属或碱土金属离子(Na+, K+, Mg2+等)通过与磷酸骨架上的氧原子结合, 中和磷酸骨架的强电负性质, 稳定其二级、三级结构. 糖环氧原子与金属离子作用较弱, 但研究表明其仍具备与过渡金属离子(Cu2+, Sn4+, Os4+等)螯合形成核酸-金属离子复合物的能力. X射线晶体衍射和NMR实验结果表明, 杂环碱基可提供多个含孤电子对的N, O等金属离子耦合位点, 且与金属配体间存在氢键、π-π堆积等多种作用力, 是金属离子结合的最佳位点. 研究结果显示, 多种金属离子及金属离子配合物与杂环碱基的结合位点包括: 嘌呤碱基G和A的N7原子, G碱基的O6原子和A碱基的N1原子, 嘧啶碱基T和C的O2和N3原子. 值得注意的是, 不同碱基与金属原子的结合位点及作用力不同, 且可通过以上位点协同作用加强与金属离子的结合. 此部分内容在多篇综述性论文中有详细介绍[6,17,18], 在此不再赘述. 图 2 金属离子与DNA之间的两种作用方式
图 2 金属离子与DNA之间的两种作用方式Figure2. Two modes of interaction between metal ions and DNA.
3.1.天然核酸介导的金属原子组装
金属离子可以通过共价相互作用与DNA分子中的特定碱基结合, 形成以金属离子为媒介的稳定碱基对结构[19,20]. 2004年, Ono等[21]发现了核酸序列中的T-T错配碱基对能够与Hg2+结合形成稳定的T-Hg2+-T配合物, 其他人利用上述性质构建了一系列基于核酸Hg2+的灵敏检测分子探针[22-24]. 2008年, Ono等[25]报道了Ag+可以通过与C碱基对中N3位置相结合而形成C-Ag+-C结构以稳定双螺旋DNA中的C-C碱基错配. Zhao等[26]和Yang[27]等利用此性质并通过不同方法实现了Ag+的特异性检测. 图3(a)展示了T-Hg2+-T和C-Ag+-C配对结构示意图. 此外, DNA还能够与其他金属离子发生共价相互作用, 例如DNA与Cu2+结合形成的DNA金属酶可以催化不对称合成, 其中DNA结构为反应的发生提供了合适的手性环境[28,29]. 图 3 (a) T-Hg2+-T和C-Ag+-C结构示意图; (b) 顺铂与DNA相互作用形成的1, 2-链内加合物[18]; (c) 用于铂药物靶向递送的纳米抗体偶联DNA纳米平台示意图[34]
图 3 (a) T-Hg2+-T和C-Ag+-C结构示意图; (b) 顺铂与DNA相互作用形成的1, 2-链内加合物[18]; (c) 用于铂药物靶向递送的纳米抗体偶联DNA纳米平台示意图[34]Figure3. (a) Illustration of T-Hg2+-T and C-Ag+-C complexes induced fluorescence quenching; (b) 1, 2-intrastrand adducts formed between cisplatin and DNA[18]; (c) illustration of a nanobody-conjugated DNA nanoplatform for targeted platinum drug delivery[34].
此外, 除金属离子直接与核酸碱基相互作用, 金属离子还可通过阳离子金属配合物与核酸分子间接结合. 研究表明, 阳离子金属配合物与DNA之间的共价相互作用是众多金属类抗癌药物发挥作用的基础[13]. 1969年, 化学治疗性金属基药物顺铂的发现是抗癌药物中DNA-金属相互作用的主要实例之一, 其通过共价结合的方式与DNA上的G碱基结合使得dsDNA解旋并抑制随后的转录等一系列过程, 最终诱导癌细胞的凋亡[30,31]. 顺铂能够与DNA结合形成多种加合产物, 其中最常见的类型是1, 2-链内加合物, 如图3(b)所示. 其他一些中心原子为Pt, Ru, Ti, Os, Co, Ni, Cu和Zn等金属配合物药物也被报道具有抗癌活性, 并展示出与顺铂类药物类似抗癌原理[14,18,32,33]. 此外, 利用阳离子金属配合物能够通过凹槽缔合或嵌入的方式与dsDNA相结合这一特点, Ding等[34]实现了含Pt金属配合物抗癌药物的靶向递送和癌症治疗(图3(c)); Wang等[35]和Li等[36]构建了基于Ru(II)配合物发光探针的电致化学发光生物传感平台. 阳离子金属配合物还能够与具有特定空间构型的DNA相互作用, 其中最典型的是氯化血红素(hemin)/G-四链体结构[37,38]. Hemin/G-四链体通常表现出辣根过氧化物酶(HRP)活性, 例如Wang等[39]利用hemin/G-四链体催化3, 3′, 5, 5′-四甲基联苯胺(TMB)氧化实现了K+的比色检测; Huang等[40]通过hemin/G-四链体结构催化H2O2还原产生电化学信号实现了胃癌相关外泌体的检测. Golub等[41]发现hemin/G-四链体还具有烟酰胺腺嘌呤二核苷酸(NADH)氧化酶/过氧化物酶活性, 这一发现为NAD+的再生提供了一种生物型催化剂.
2
3.2.天然核酸介导的金属纳米簇的合成与应用
荧光金属纳米簇(metal nanoclusters, MNCs)具有尺寸小、稳定性高和生物相容性好等优势, 在生物检测和成像领域的应用非常广泛. 基于DNA序列中特定碱基杂环上的N, O功能基团与金属离子之间具有相互作用强, 且DNA的碱基序列和长度可调、二级结构多样、生物相容性好, 使得DNA成为调控MNCs生长有效模板之一[11,42]. 以DNA为模板调控MNCs合成的基本步骤如下: 金属离子首先与DNA结合, 然后还原成核, 进一步生长, 最后在DNA的保护下稳定存在. 2004年, Petty等[43]首次在磷酸盐缓冲溶液中以12个碱基的ssDNA为模板合而成了荧光AgNCs. 随后的研究表明C碱基的N3位置与Ag+之间具有较强亲和力, 因而富C碱基的ssDNA序列通常被用作AgNCs生长的模板[44,45]. Gwinn等[46]利用发夹DNA制备得到了AgNCs, 他们发现Ag-NCs的荧光强度与发夹环上DNA碱基的种类相关. Feng等[47]以三链DNA(tsDNA)作为模板制备得到了荧光AgNCs, 实验表明AgNCs的成核与tsDNA的CGC位点有关. 以上研究结果表明, 通过调控核酸分子的序列和二级结构等参数, 可实现金属纳米团簇的可控合成. 结合原位DNA介导的金属纳米团簇生长及功能核酸分子, Liu等[48]和Liu等[49]利用荧光DNA-AgNCs作为信号探针分别构建了microRNA和蛋白的痕量分析检测平台; Lyu等[50]通过静电吸附的方式在DNA-AgNCs表面修饰阳离子聚电解以提高其荧光强度、稳定性和细胞穿透能力, 从而实现了NIH/3 T3细胞的快速荧光成像(图4). Thomas[51]以含有30个碱基的ssDNA为模板合成了发射蓝光的AuNCs, 研究结果表明, pH以及HAuCl4和DNA的浓度比会影响碱基与Au3+的结合, 从而通过改变以上参数可实现金属纳米团簇的可控制备. 研究表明T碱基的N3位置与Cu2+之间的作用力较强, 因而可通过富T碱基的核酸序列调控CuNCs的制备[9,52,53]. 此外, 科研工作者还报道了G-四链体[54]和i-motif[55]结构也可以作为MNCs生长的模板, 调控金属纳米团簇的生长及其光学性质. 值得关注的是, 通过DNA模板成功合成了Cu/AgNCs[56]和Ag/PtNCs[57]等双金属纳米簇, 解决了化学湿法合成多金属纳米团簇的技术难点.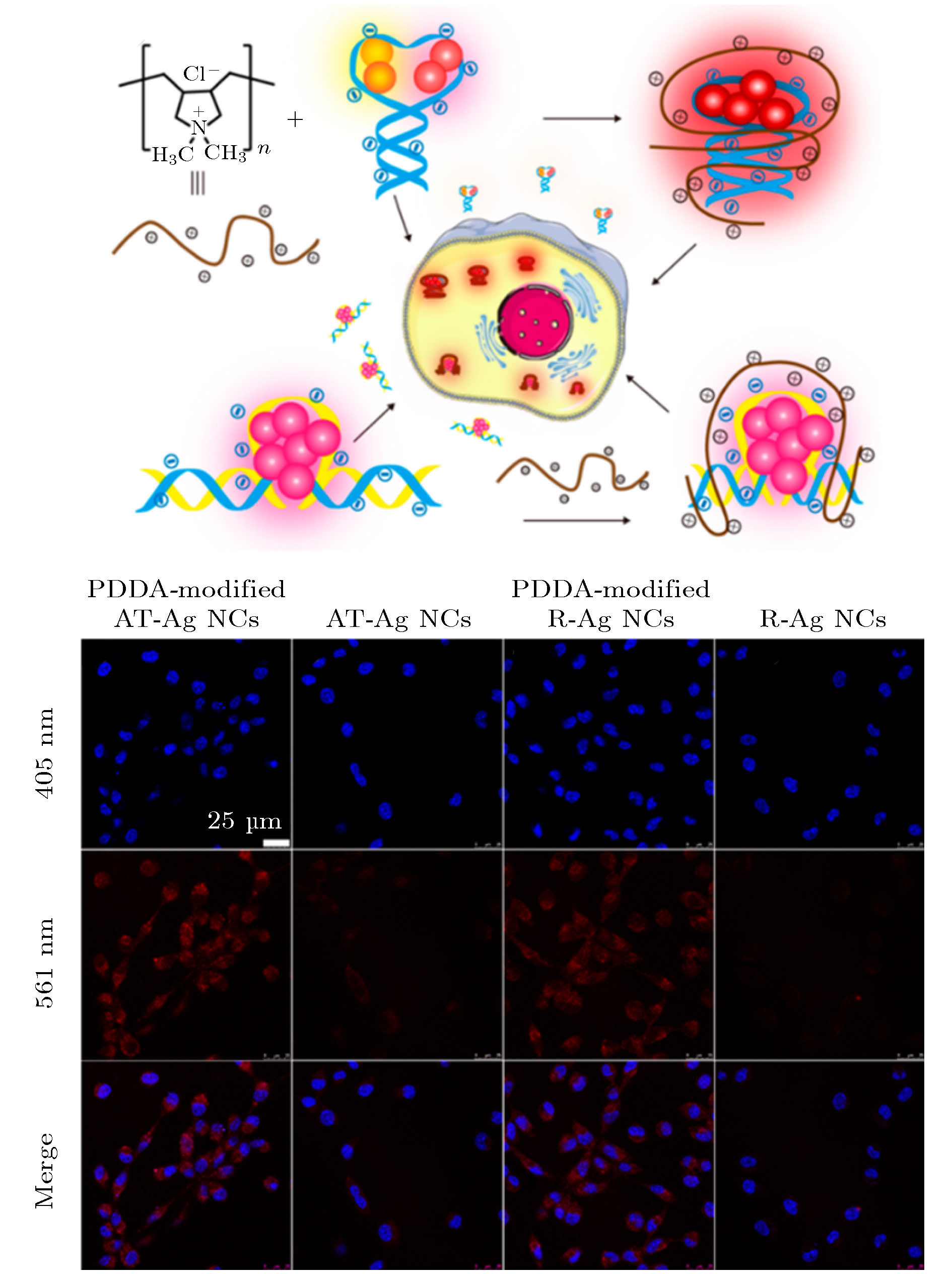 图 4 制备不同DNA@Ag-NCs-阳离子聚电解质复合物用于细胞成像以及NIH/3T3细胞的共聚焦激光扫描显微镜成像图[50]
图 4 制备不同DNA@Ag-NCs-阳离子聚电解质复合物用于细胞成像以及NIH/3T3细胞的共聚焦激光扫描显微镜成像图[50]Figure4. Preparation of different fluorescent DNA-AgNCs-cationic polyelectrolyte complexes for cell imaging and confocal laser scanning microscopy images of NIH/3T3 cells[50].
2
3.3.功能核酸与金属相互作用及应用
功能核酸是通过配体指数富集系统进化技术(systematic evolution of ligands by exponential enrichment, SELEX)筛选得到的具有靶向结合、催化等特定功能的单链寡核苷酸序列[58,59]. 自1990年SELEX技术首次报道以来, 通过体外筛选技术获得了一系列可结合金属离子的功能核酸分子, 主要包括金属离子特异性核酸适体(aptamer)和脱氧核酶(DNAzyme)等, 并构建了一系列基于功能核酸材料的分子器件, 在生物检测、成像和癌症治疗等方面有着广泛的应用[60-63].Aptamer是通过SELEX技术筛选得到能与靶标分子特异性、高亲和力结合的单链寡核苷酸序列[64-67]. 以金属离子作为筛选靶标可得到与特定金属离子结合的aptamer分子, aptamer常通过折叠形成特定二级/三级结构与靶标分子特异性结合[68,69]. 研究表明, K+, Na+和Pb2+与aptamer的作用机理均与G-四链体结构有关[70]有关. 图5(a)展示了由中心阳离子稳定、以Hoogsteen氢键结合4个G碱基和人端粒G-四链体DNA的X射线结构. 1994年, Williamson[71]发现G-四链体结构在K+和Na+等阳离子存在时比较稳定, 机理研究表明金属离子与碱基上的氧原子配位结合, 从而发挥结构稳定作用. 后来特异性结合Pb2+的aptamer被开发, 且基于此构建了一系列Pb2+检测的传感平台[69,72,73].
 图 5 (a) 由中心阳离子稳定的4个G碱基和人端粒G-四链体DNA的X射线结构[18]; (b) 金属离子依赖性DNAzyme的结构示意图[62]
图 5 (a) 由中心阳离子稳定的4个G碱基和人端粒G-四链体DNA的X射线结构[18]; (b) 金属离子依赖性DNAzyme的结构示意图[62]Figure5. (a) Illustration of four guanine bases stabilized by a central cation and X-ray structure of human telomeric G-quadruplex DNA[18]; (b) illustration of metal ions dependent DNAzyme[62].
DNAzyme是一种通过体外筛选技术得到的具有折叠成复杂二级结构的ssDNA序列, DNAzyme通过与金属离子活性中心结合, 可催化特异性核酸切割/连接等反应, 包括RNA或DNA的裂解和连接以及DNA磷酸化等[74,75]. 图6(b)展示了金属离子依赖性DNAzyme的结构, 由一条到底物链和一条酶链组成. 目前为止, Pb2+, Cu2+, Ag+, Zn2+, Mg2+, Ce3+, UO22+和Ln3+等金属离子特异性识别的DNAzyme相继被筛选出来[62,63], 在金属离子、核酸或者蛋白等的检测以及细胞内基因沉默和癌症治疗等方面[7,76-80]有着广泛的应用前景.
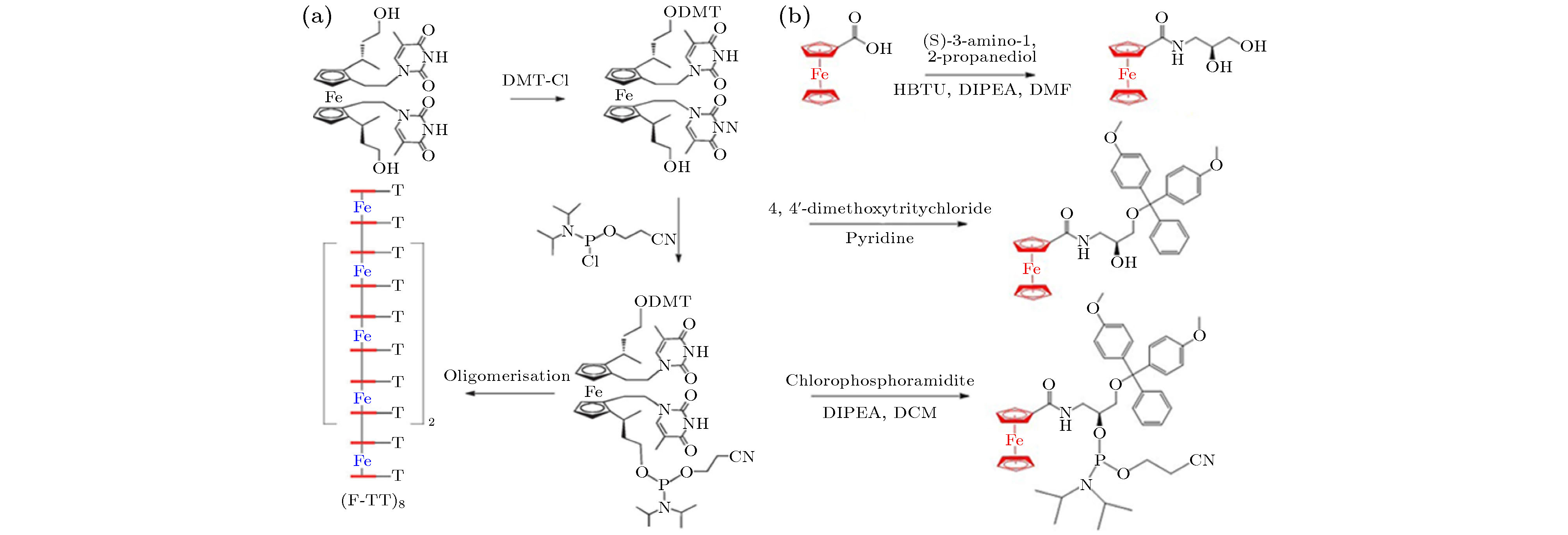 图 6 (a) (Fc-TT)8序列的合成步骤[86]; (b) Fc人工碱基单体的合成步骤[91]
图 6 (a) (Fc-TT)8序列的合成步骤[86]; (b) Fc人工碱基单体的合成步骤[91]Figure6. (a) Synthesis procedure of (Fc-TT)8[86]; (b) synthesis procedure of Fc artificial base monomer[91].
功能核酸分子的核心优势在于可特异性识别特定元素的金属离子, 结合核酸分子序列的可设计性及核酸分子间的自组装能力, 为单原子操纵及组装提供了一种全新的分子工具. 在单原子器件和多原子组装等领域具有巨大的应用潜能.
2
3.4.人工碱基“分子元素”介导的原子制造
在过去的几十年间, 化学家和合成生物学家们一直致力于人工核酸的开发与应用[81,82]. 从核酸的基本结构和性质出发, 通过使用合适的替代物替换核酸聚合物五碳糖、磷酸骨架和碱基中的一个或多个基本单元, 可获得具有区别于天然核酸的物理化学性质的人工核酸分子[83,84]. 随着核酸化学的不断发展, 一系列有机合成的人工核酸相继被报道, 其中金属人工核酸作为金属聚合物多元化领域的重要组成部分, 赋予了核酸材料新功能和新应用, 引起了科研工作者广泛的研究兴趣[85]. 金属人工核酸的制备方法主要有以下两种[86-88]: 1) 不改变杂环碱基基本结构, 用金属化合物来代替DNA中的糖磷酸骨架; 2) 合成含金属配体的亚磷酰胺单体, 通过DNA合成仪实现亚磷酰胺单体聚合形成寡核苷酸序列.对于第一种策略, 最受关注的金属配合物是具有良好电化学活性的二茂铁(Fc), 其环戊二烯基环之间的间隙(3.3 ?)与B-DNA中相邻碱基对之间的间隙(3.4 ?)非常相似, 因而成为替代DNA中的糖磷酸骨架的最佳选择[89,90]. 2012年, James等[86]基于DNA中两个呋喃糖环被Fc分子中两个环戊二烯基单元取代, 通过一系列复杂的有机反应将T碱基标记在Fc单元上, 进一步形成亚磷酰胺单体后, 通过DNA自动合成仪合成了包含8个Fc单元和16个T碱基的(Fc-TT)8序列(图6(a)). 合成的(Fc-TT)8序列易溶于磷酸盐缓冲溶液并且约260 nm处有吸收峰, 且Fc的d-d跃迁的存在使得约435 nm处产生较弱的吸收峰. 通过人工核酸分子的合成实现了Fe原子的一维可控组装, 研究人员虽未进一步探究该人工核酸分子间的杂交性质, 基于其类天然核酸的结构及保留了T碱基的氢键形成能力, 其有望进一步通过分子间杂交形成复杂的二维及三维原子结构.
区别于糖环结构的替换, 湖南大学谭蔚泓院士课题组[91-95]率先提出了通过替换核苷酸分子中碱基基元形成一系列“分子元素”的基本概念. 在这一基本概念的指导下, 合成了一系列基于金属配体、药物分子、疏水分子、刺激响应分子的亚磷酰胺单体, 并通过DNA合成仪实现了分子元素的可控组装. 2018年, Abdullah等[91]通过简单的三步法成功合成了Fc人工碱基单体(图6(b)), 并首次实现了二茂铁人工碱基“分子元素”的制备. 基于Fc分子一个电子转移氧化还原的特性, 将Fc人工碱基单体嵌入DNA序列的不同位置, 通过电化学测试发现随着互补碱基的不同, 电荷转移速率明显不同, 表明合成的Fc人工碱基可以作为检测任意靶序列中单个碱基的变化, 有望成为DNA测序的有力工具. 研究结果表明进一步研究发现, 通过调控Fc人工碱基的数量, 可有效地调控其组装性质. Tan等[96]将修饰了多个Fc人工碱基DNA序列自组装成尺寸可控的DNA胶束(ApFAs), 结果表明通过改变末端Fc碱基的数量可实现组装纳米颗粒的尺寸, 且利用Fc引发类芬顿反应可实现纳米材料的组装与解组装行为(图7(a)), 充分展示了该策略在原子制造领域的有效性. Zhang等[97]设计了嵌入有Fc人工碱基的DNA纳米花结构(DOX-Sgc8-NFs-Fc), 进一步构建了从单原子到纳米材料的制造策略(图7(b)).
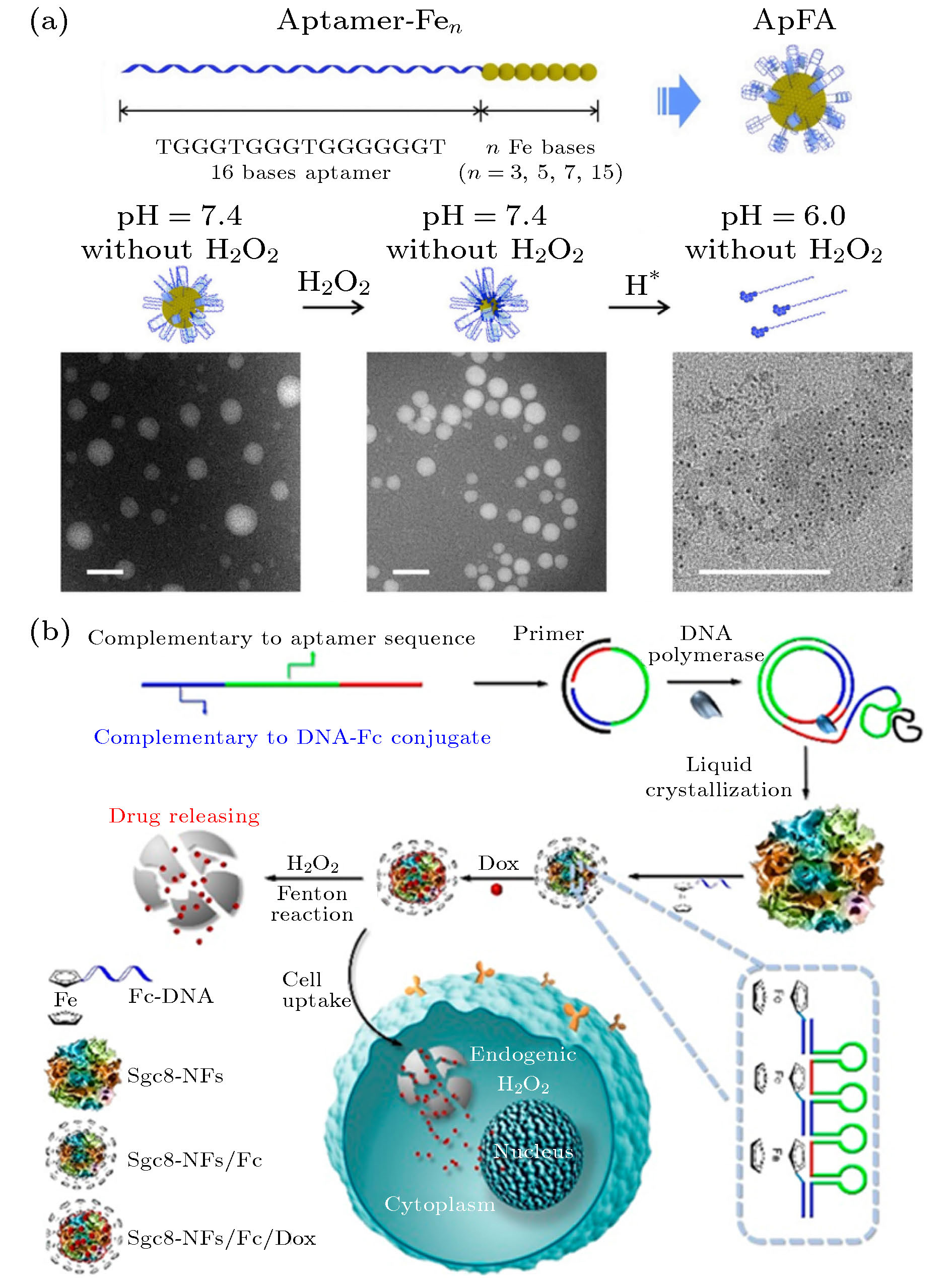 图 7 (a) 适体-Fen两亲分子的示意图以及在不同条件下的ApFAs的TEM图像[96]; (b) DOX/Sgc8-NFs-Fc的制备及其通过类芬顿反应在癌细胞中的自降解过程[97]
图 7 (a) 适体-Fen两亲分子的示意图以及在不同条件下的ApFAs的TEM图像[96]; (b) DOX/Sgc8-NFs-Fc的制备及其通过类芬顿反应在癌细胞中的自降解过程[97]Figure7. (a) Schematic of aptamer-Fen amphiphilic molecules and TEM images of ApFAs at different conditions[96]; (b) preparation of DOX/Sgc8-NFs-Fc and its autodegradation process in cancer cells through Fenton-like reaction[97].
4.1.核酸纳米结构
自从1983年美国纽约大学Seeman教授[98]首次提出DNA纳米技术以来, 科研工作者不断利用DNA这种典型的原子级精准自组装体构筑成特定的纳米结[99-103]. 基于Waston-Crick碱基互补配对原则高特异性、可预测性和高精确性, 通过“自下而上(bottom-up)”的自组装策略, 可以精准设计并合成具有不同形貌、尺寸的DNA纳米结构, 人们设计和制备得到了大小和形状各异的精美DNA纳米结构, 包括三棱柱[104,105]、四面体[106,107]、多面体[108,109]、水凝胶[110,111]、DNA tile[112,113]和DNA折纸[114,115]等. 2006年, Rothemund[116]提出了DNA折纸的概念, 由此发展起来的DNA折纸技术引起了广泛的研究兴趣. 将216条30—40碱基长度的ssDNA订书钉链与含有7249个碱基的环形闭合长ssDNA骨架链紧密有序地钉在一起形成了具有特定形貌的二维DNA折纸(图8(a)). 2009年, Douglas等[114]通过在二维DNA折纸平面之间建立连接, 使其层层堆叠形成三维DNA折纸结构, 进一步拓展了DNA纳米结构的维度. 2016年, Douglas等[117]利用“自上而下(top-down)”策略构建了三维多面体DNA折纸(图8(b)), 标志着DNA纳米技术迈向了新的高度. DNA折纸不仅具有形貌、尺寸可控的优势, 还能精确设计每个碱基的类型和位置, 使其具有纳米(亚纳米)级的空间可寻址能力. 因此, 可以在DNA折纸的特定位置上实现纳米尺度精确定位的基团修饰, 使其成为单分子精准定位、纳米材料可控排布和生长的理想模板[118-123]. 图 8 (a) 二维DNA折纸[116]; (b) 三维多面体DNA折纸[117]
图 8 (a) 二维DNA折纸[116]; (b) 三维多面体DNA折纸[117]Figure8. (a) 2D DNA origami[116]; (b) 3D polyhedral DNA origami[117].
2
4.2.DNA折纸介导的自组装
调控纳米颗粒的空间排布是纳米技术领域长期以来存在的挑战之一, 具有形貌、尺寸可控且空间寻址能力强的DNA折纸为解决这一问题提供了新的工具. 以DNA折纸为模板的纳米颗粒自组装, 通常利用DNA功能化的纳米颗粒与DNA折纸上特定区域的互补链进行杂交实现纳米颗粒的可控排布. 2010年, Ding等[120]首次利用三角形DNA折纸作为模板, 使得不同粒径的AuNPs在折纸的其中一条边上特定的位点进行有序的排布(图9(a)), 由此开辟了基于DNA纳米技术实现金属纳米材料精确组装的新天地. 随后, 一系列具有特殊结构、光学性质的金属纳米结构相继被报道[124,125]. 除了实现纳米颗粒的精确空间排布以外, 科研工作者还利用DNA折纸实现了生物蛋白分子、单分子荧光染料等的精确组装[126,127]. Fu等[128]将葡萄糖氧化酶(GOx)和辣根过氧化物酶(HRP)精确定位在DNA折纸的不同位置, 通过对蛋白分子间的距离的精细调控, 研究了距离对着两种酶协同催化效果的影响(图9(b)). Zhan等[119]使用基于DNA折纸的自下而上组装策略成功构建了等离子领结纳米结构, 进而将单个拉曼报告分子限域在领结的间隙处, 实现了单个纳米结构中单分子表面增强拉曼散射分析(图10(a)). Liu等[129]报告了一种“Action-PAINT”策略, 利用DNA-PAINT实时监控和定位DNA结合事件, 通过光交联以固定分子信标进行特定位置的可视化, 实现了在单个分子上进行可视化时进行超分辨率标记(图10(b)).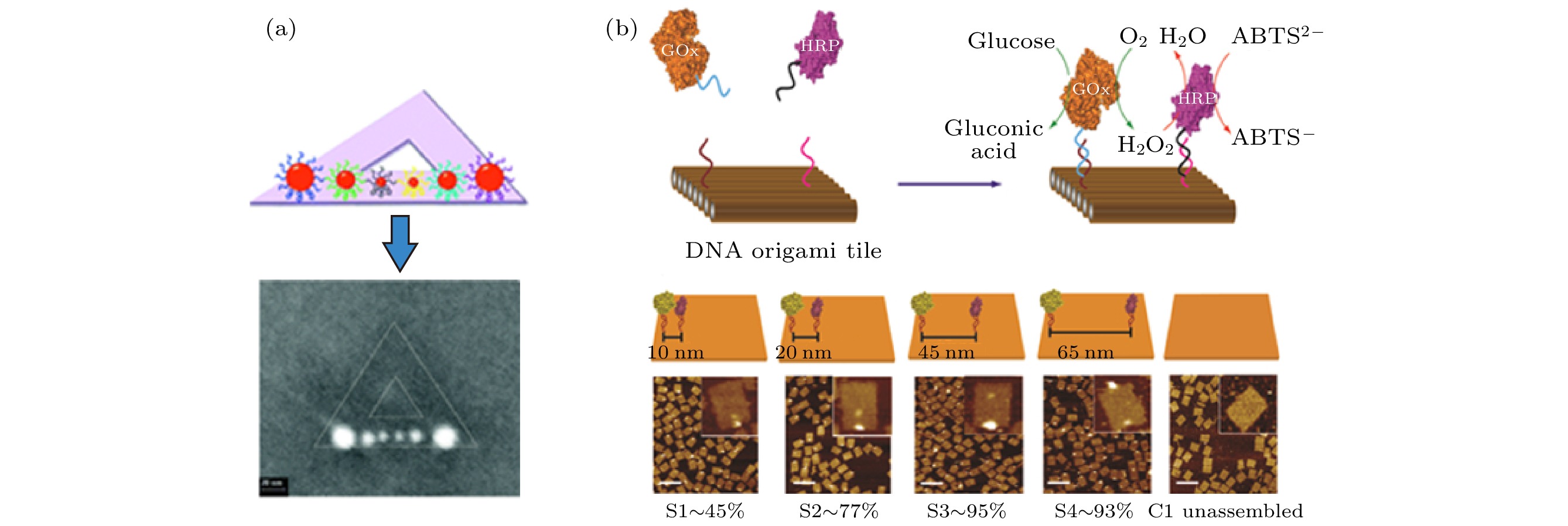 图 9 (a) 利用三角折纸对不同大小AuNPs进行空间排布[120]; (b) DNA折纸介导的GOx和HRP的距离可控共组装[128]
图 9 (a) 利用三角折纸对不同大小AuNPs进行空间排布[120]; (b) DNA折纸介导的GOx和HRP的距离可控共组装[128]Figure9. (a) 2D arrangement of Au NPs using triangle DNA origami[120]; (b) DNA nanostructure-directed coassembly of GOx and HRP enzymes with control over interenzyme distances[128].
 图 10 (a) 基于DNA折纸的金领结纳米结构用于单分子表面增强拉曼散射分析[119]; (b) 用“Action-PAINT”实现单分策略的多点超分辨图案[129]
图 10 (a) 基于DNA折纸的金领结纳米结构用于单分子表面增强拉曼散射分析[119]; (b) 用“Action-PAINT”实现单分策略的多点超分辨图案[129]Figure10. (a) Gold bowtie nanostructures based on DNA origami for single-molecule surface-enhanced Raman scattering analysis[119]; (b) multipoint super-resolution patterning using “Action-PAINT” strategy[129].
2
4.3.DNA折纸介导纳米材料的合成
基于核酸材料与金属离子强相互作用, 核酸分子材料作为模板与金属离子前体结合后, 通过合适的还原剂原位还原金属离子前体, 形成特定形状的金属纳米结构[11,42]. Braun等[130]利用充分利用具识别能力和机械性能的DNA模板桥连两个金电极, 然后沿着DNA分子搭建的模板方向使Ag进行矢向沉积, 从而制备得到仅沿DNA骨架方向沉积的长12 μm、宽100 nm的Ag纳米导线, 这一创新性工作标志着DNA模板为金属纳米材料的精确组装指明了新的方向. 除简单的DNA以外, DNA纳米结构的发展为金属化提供了结构多样的模板. 2011年, Liu等[131]实现了DNA折纸表面的金属化, 以Y型DNA折纸为生长模板, 通过选择性表面生长金属Ag种子, 然后在Ag种子上生长AuNPs. 同年, 他们报道了一种基于Pd种子的快速DNA折纸金属化的新方法, 减少了金属化过程的时间且增加金属化颗粒的密度[132]. 随后, 他们在组装有Pd种子的环形DNA折纸上进行了Au和Cu的金属化, 首次实现了DNA折纸模板上制造导电铜纳米结构[133]. 为进一步提高DNA折纸上金属化位点的可控性, Pilo-Pais等[134]通过延长订书钉链并利用碱基互补配对将成核种子偶联在DNA折纸骨架上, 再进行成核生长也可以在DNA折纸上实现精准的定位金属化, 制备了不同图案的AuNPs结构. Jia等[122]开发了一种基于DNA折纸的高度局部化金属化反应策略, 实现了在全DNA基底上对字母、数字和几何形状进行自由样式的金属绘画, 并模拟制造了单层和双层纳米级印刷电路板, 进一步提高了以DNA折纸为模板的金属化的精准度和可控性, 为纳米电子和纳米光子应用指明了新的方向(图11(a)). 2014年, Helm等[135]和Sun等[136]几乎同时提出了DNA 模具法, 在三维尺度上实现了基于DNA折纸的金属限域生长. 他们设计了具有不同形状的DNA空腔折纸作为模板, 通过还原金属离子制备得到立方体、三角片和圆片等不同形貌的金属纳米颗粒. 除常见金属离子, Fan等[123]利用DNA折纸诱导的团簇预水解策略在纳米尺度上将DNA序列编码的自组装结构复制成具有刚性结构的精确二氧化硅构型(图11(b)). 这一工作不仅突破了传统硅化学合成在结构尺度上的限制, 实现了纳米尺度的精确二氧化硅结构的制备; 而且赋予基于DNA的固态纳米孔在保持精确结构的同时还具备了更好的力学性能, 进一步拓展了基于核酸材料的原子制造的适用领域. 图 11 (a) 使用DNA缩合和固有的金属化图案模拟纳米印刷电路板[122]; (b)利用不同DNA折纸为模板生长SiO2[123]
图 11 (a) 使用DNA缩合和固有的金属化图案模拟纳米印刷电路板[122]; (b)利用不同DNA折纸为模板生长SiO2[123]Figure11. (a) Fabricating nano-printed circuit boards mimics with DNA condensation and intrinsic metallization patterning[122]; (b) growth of SiO2 with different morphology by various DNA origami[123].
